
Abstract
It has been almost two decades since the first demonstration of hydrogen sulfide (H2S) as a physiological mediator of cognitive function and vascular tone. H2S is physiologically important because it protects various organs from ischemia–reperfusion injury besides regulating inflammation, oxygen sensing, cell growth, and senescence. The production, metabolism, and regulation of H2S have been studied extensively. H2S modulates target proteins through sulfhydration (or sulfuration) or by the reduction of cysteine disulfide bonds. A large number of novel H2S-donating compounds are being developed owing to the therapeutic potential of H2S. Recently, polysulfides, rather than H2S, have been identified as molecules that sulfhydrate (or sulfurate) their target proteins. Antioxid. Redox Signal. 22, 347–349.
T
One of the modes through which H2S carries out signaling is by the addition of sulfur to the cysteine residues of target proteins, thereby producing bound sulfane sulfur. This elicits a conformational change in the proteins and modifies their activities. The above process is called sulfhydration or sulfuration, and it is analogous to the nitrosylation reaction carried out by NO (5). Recently, however, polysulfides, but not H2S, have been shown to induce sulfhydration (or sulfuration). Molecules with the same oxidation state do not react with each other. Because the oxidation state of S in both H2S and cysteine residues is −2, H2S and cysteine residues are unable to react with each other. In contrast, because the oxidation state of S in polysulfides is 0, it easily reacts with cysteine by adding bound sulfane sulfur to this residue (sulfhydration). Polysulfides activate the transient receptor potential ankyrin 1 (TRPA1) channels by sulfhydrating (sulfurating) two cysteine residues at the amino terminus (2). They also facilitate the translocation of Nrf2 (which upregulates the transcription of antioxidant genes) to the nucleus by sulfhydrating an accompanying protein called Keap1. The activity of the phosphatase and tensin homolog (PTEN), a lipid phosphatase, which regulates the growth of cancer cells, is also regulated by polysulfides (2). The physiological roles of polysulfides are therefore quite robust (Fig. 1).
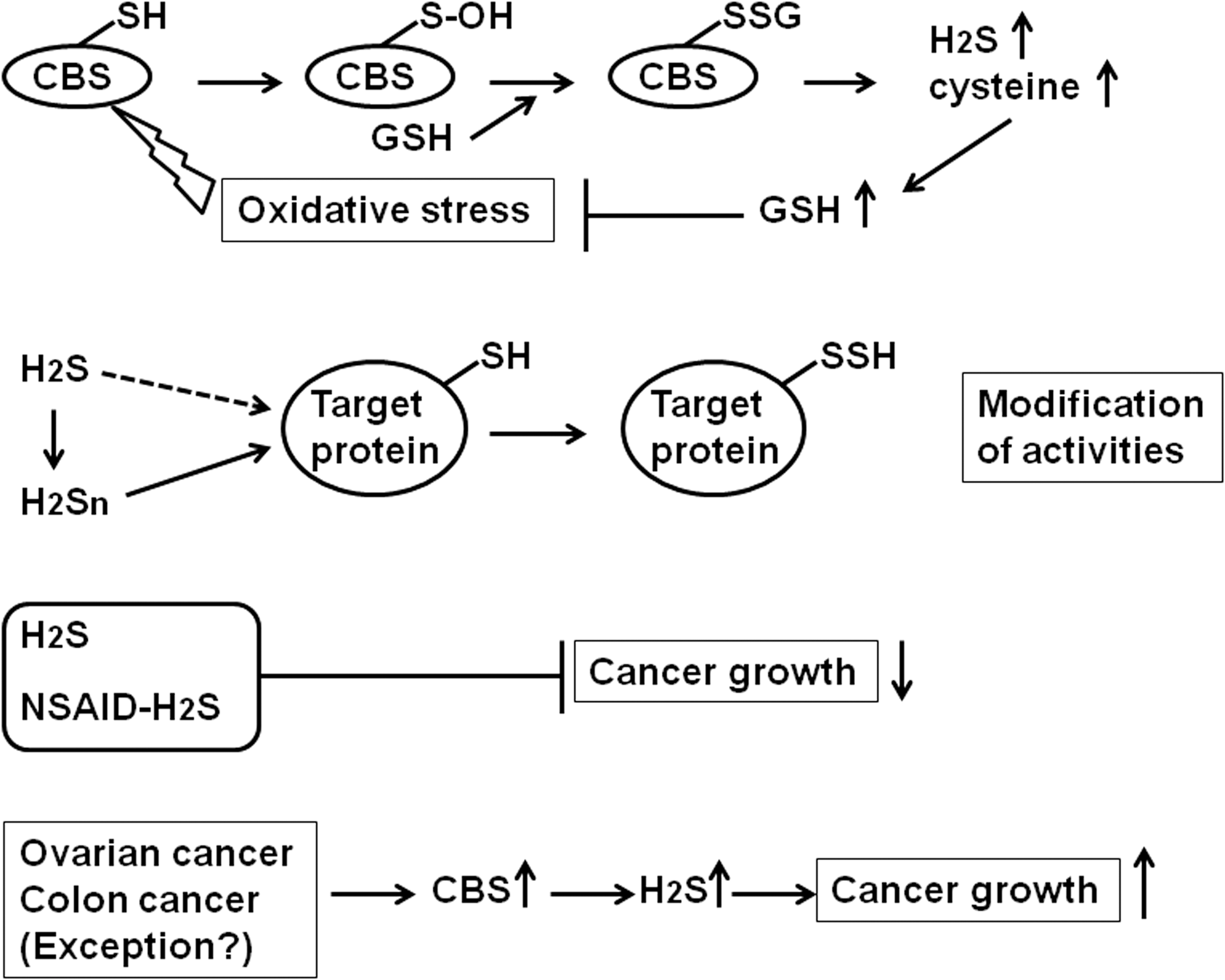
H2S has been reported to inhibit cancer development at various stages (6). H2S donors are cytotoxic to human cancer cells. They also induce cell cycle arrest in the G2/M phase and promote apoptosis. However, they do not affect the survival of normal human cells. Several recent studies have shown that H2S-releasing NSAIDs, which were initially developed by Wallace et al. as anti-inflammatory drugs having no side effects on the gastrointestinal tract, exhibit enhanced chemopreventive effects in cancer cells (7). These compounds inhibit cancer cell proliferation and promote apoptosis, as well as cell cycle arrest. The parental NSAIDs alone do not confer any beneficial effects in cancer cells. H2S donors and H2S-releasing NSAIDs show great promise as therapeutic agents against several forms of cancer. Contrary to these observations, CBS is upregulated in colorectal and ovarian cancer cells, where H2S is involved in promoting cellular bioenergetics, proliferation, and migration (1). In these cancer cells, the suppression of CBS activity can be used as a potential therapy (Fig. 1).
Banerjee and colleagues, in their original article, demonstrated that the CBS activity is enhanced by the S-glutathionylation of a cysteine residue of CBS in cells under oxidative stress (3). Cells experiencing oxidative stress decrease their glutathione levels to prevent further damage. S-glutathionylation increases the CBS activity, which in turn generates cysteine (a substrate for the production of glutathione) as well as H2S. The generation of these two products makes cells resistant to oxidative stress (Fig. 1).
In their review article, Paul and Snyder discussed the physiological role of sulfhydration (sulfuration), a process that involves the modification of protein activities and produces effects opposite to those elicited by nitrosylation (5). Sulfhydration of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) increases its catalytic activity, whereas nitrosylation suppresses it. Parkin, an E3 ubiquitin ligase, is a protein that is affected during Parkinson's disease. It is less sulfhydrated and shows a reduced catalytic activity in patients suffering from Parkinson's disease. However, the same protein is nitrosylated and inactivated to a higher degree after the onset of this neurodegenerative disease (5). Paul and Snyder also gave one more example of the involvement of H2S in neurodegenerative disorders: the levels of H2S and CSE are lower in patients suffering from Huntington's disease than in normal individuals. H2S thus plays an important role in the pathophysiology of this disease (5).
H2S induces conditions similar to hypoxia and acts on effecter molecules known to mediate hypoxic responses. The metabolism of H2S is coupled to that of O2, and the concentrations of H2S are inversely related to those of O2 in tissues. Based on these observations, H2S has been proposed as an O2 sensor (4). In his review article, Olson discussed that the O2-dependent metabolism of H2S is an O2 sensor (4). The highly reactive sulfur is able to form bonds with its own species as well as with nitrogen and oxygen. It regulates protein functions through sulfhydration (sulfuration) and nitrosylation, as described in the Forum article by Paul and Snyder (5). O2-dependent H2S inactivation can be effectively used in accurately determining O2 availability and for coordinating the supply and demand at O2-sensing tissues and cells. The development of specific inhibitors of H2S metabolism and the development of H2S donors that release H2S in a controlled manner will enhance the understanding of the O2-sensing process of this molecule.
The effects of H2S and related compounds on various types of cancers and their ability to suppress cancer development have been described previously (6, 7). According to Szabo and colleagues, the production of H2S by CBS is increased in colorectal and ovarian cancers and both play an important role in promoting cellular bioenergetics, proliferation, and migration (1). H2S induces angiogenesis and local vascular relaxation, which increases the blood flow and nutrient supply to the tumor. It also enhances oxygen consumption, ATP production, and glutathione production to support tumor growth. The therapeutic potential of CBS inhibitors in cancer therapy has been discussed previously (1). The unavoidable side effects of these inhibitors include the accumulation of cellular homocysteine, a cytotoxic molecule and a cardiovascular risk factor.
H2S exerts regulatory effects on inflammatory processes and promotes their resolution. According to Wallace et al., several H2S-releasing anti-inflammatory drugs, which are being developed for the treatment of a wide range of diseases, show considerable promise in relevant animal models. Many of these drugs are undergoing human trials (7). H2S-releasing NSAIDs show therapeutic potential because they can be used as chemopreventive agents in several forms of cancer.
In his review article, Kimura discussed the regulation of H2S-producing enzymes, described a novel pathway that produces H2S from
Some of these Forum articles were presented and discussed in the Third International Conference on H2S in Biology and Medicine, held in Kyoto and organized by Hideo Kimura in June, 2014 (Nitric Oxide 39: Supplement 1, 2014).
Footnotes
Acknowledgments
This work was supported by a grant from the National Institute of Neuroscience, KAKENHI (23659089) Grant-in-Aid for Challenging Exploratory Research, and KAKENHI (26460115) Grant-in-Aid for Scientific Research to H.K.