
Abstract
Introduction
R
The present study is the first to demonstrate the pivotal role of GSH/GSSG imbalance and its mediated oxidative stress in nimbolide (NL)-induced inhibition of signal transducer and activator of transcription 3 (STAT3) signaling axis as well as attenuation of tumor growth and metastasis in prostate cancer (PCa) transgenic mouse model.
A common characteristic of PCa is its dependence on activated STAT3 for proliferation and survival. STAT3 is an oncogenic transcription factor that is constitutively activated in many solid tumors, including PCa (4, 14, 39, 42, 44). Normally, STAT3 is present in an inactive form as a monomer in the cytoplasm; once the monomer is phosphorylated by the receptor-associated tyrosine kinases, it can form active dimers and then migrate into the nucleus to induce gene transcription (42). The receptor-associated tyrosine kinases are activated through ligand engagement; these ligands include the cytokine such as interleukin-6 (IL-6) and growth factors such as epidermal growth factor (EGF).
The persistent activation of STAT3 contributes to the development of PCa through distinct mechanism(s). First, STAT3 is involved in the progression of PCa; it participates in the change from hyperplasia to neoplasia in prostate epithelial cells (3). Second, STAT3 determines the survival of PCa cells; inhibition of STAT3 by antisense STAT3 oligonucleotides can induce apoptosis of PCa (1, 2, 30). Third, STAT3 might be involved in CRPC development as it not only protects PCa cells from apoptosis due to androgen deprivation (28) but may also enhance the function of androgen receptor even in the absence of androgen (6). These important oncogenic characteristics make STAT3 an excellent molecular target for PCa therapy.
NL, a terpenoid limonoid isolated from Neem trees and flowers (5, 10), has been found to exhibit diverse pharmacological activities such as antifeedant (45), antimalarial (36), anti-HIV (46), antimicrobial (37), and also significant anticancer responses (21). The diverse anticancer effects of NL have been reported in multiple tumor types, including those of breast, colorectal, brain, and liver (11, 20, 25, 26). More interestingly, both supercritical extract of Azadirachta indica (Neem) leaves and purified NL have also been reported to exert antiproliferative and proapoptotic effects against PCa (35, 48), but the in-depth molecular mechanisms of its anticancer effects were not completely deciphered in these studies. Considering the critical role of STAT3 in PCa progression as discussed above, we hypothesized that NL may attenuate PCa growth through targeting STAT3 signaling cascade. Our results indeed indicate that NL can induce oxidative stress-mediated blockade of STAT3 activation that contributes to its growth and metastasis inhibitory effects in PCa.
Results
NL inhibits the viability as well as reduces invasion and migration of PCa cells
DU145 and LNCaP were treated with NL (0–20 μM) for 24, 48, and 72 h and thereafter MTT assay was performed. As shown in Figure 1B, there were significant reductions in the viability of two tumor cell lines upon NL treatment and these decreases were both time and dose dependent. Viability inhibition was also observed in PC-3 cells with NL treatment (Supplementary Fig. S1; Supplementary Data are available online at
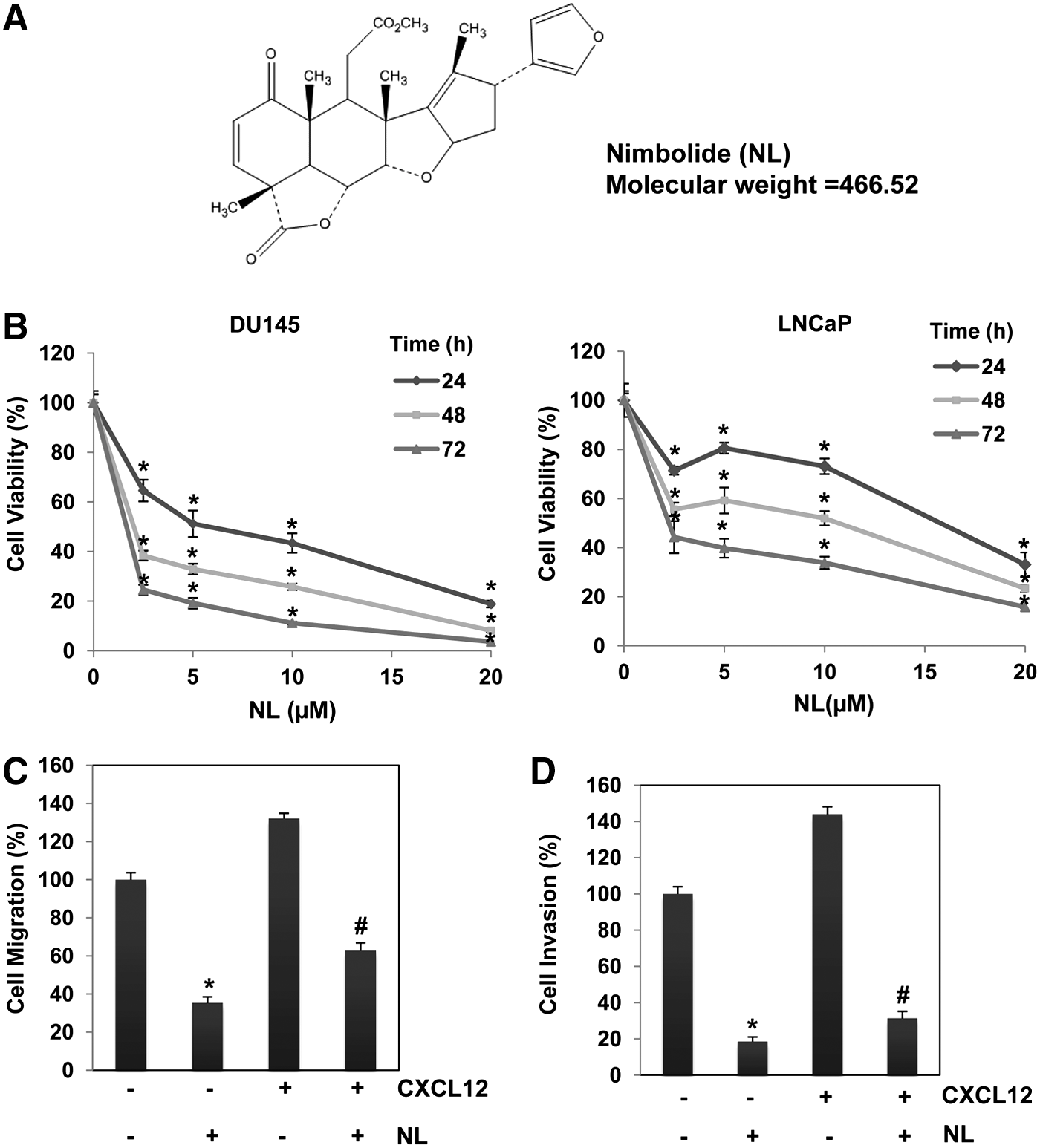
Cell invasion and migration are considered as initial steps during the process of tumor metastasis (47). Figure 1C, D presents the results obtained from wound healing and Boyden chamber assays. These two assays were performed in DU145 cells stimulated with or without chemotactic agent, CXCL12. We observed significant decreases in both cellular migration and invasion following NL treatment with and without CXCL12 stimulation, thereby providing evidence(s) that NL may exhibit antimetastatic effects against PCa cells.
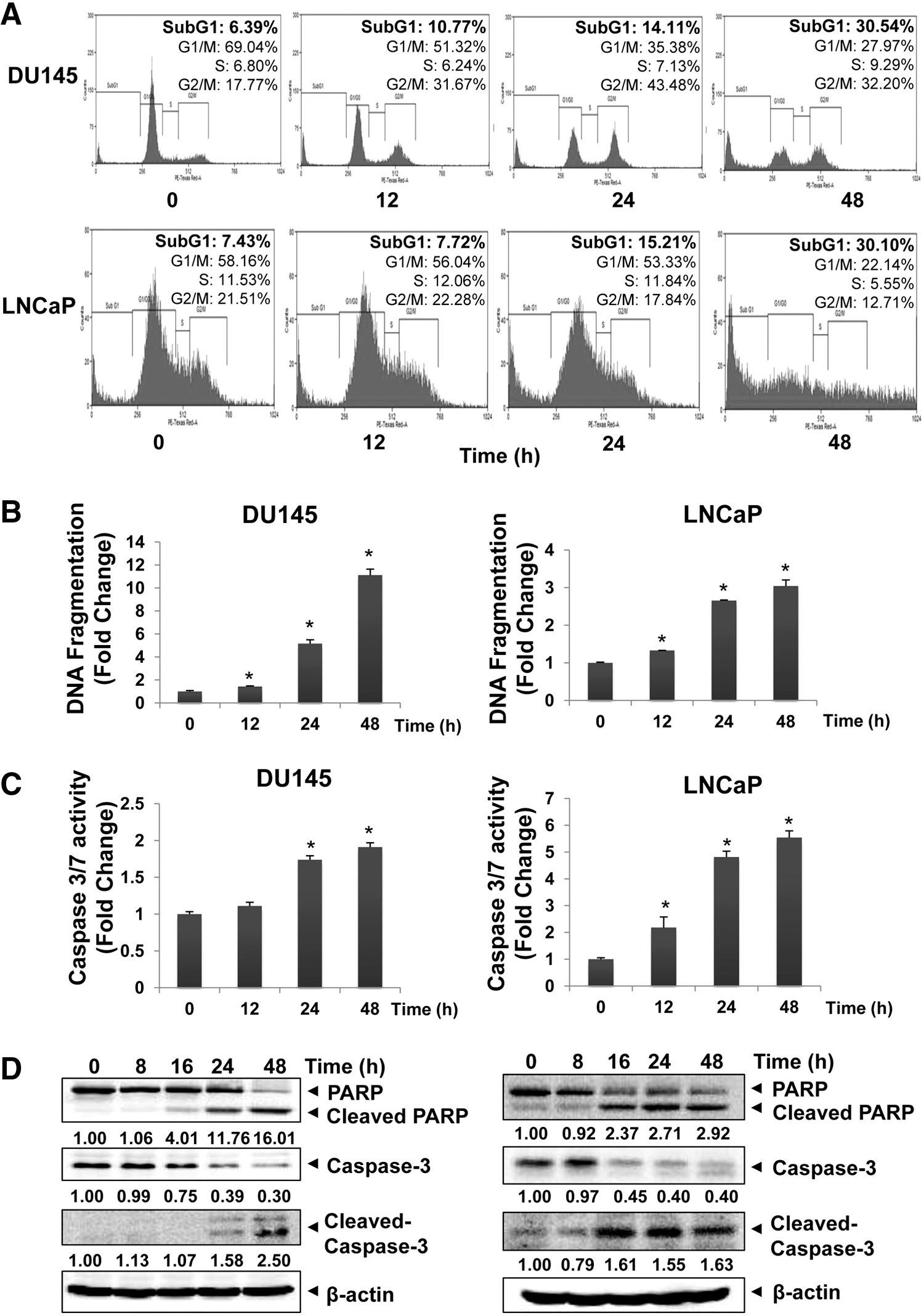
NL induces substantial apoptosis in PCa cells
We next analyzed the potential of NL to induce apoptosis in DU145 and LNCaP using diverse molecular techniques. As cells accumulated in the SubG1 phase represent the apoptotic population (24), flow cytometry was first performed to study the proapoptotic effect of NL in PCa cells. Figure 2A shows a clear increase of cell population percentage in the SubG1 phase following NL treatment and this increase was time dependent in both cell lines. DNA fragmentation is another characteristic marker of cellular apoptosis (9). From Figure 2B, it is apparent that NL increased the DNA fragmentation levels in both DU145 and LNCaP cells in a time-dependent manner. The effects of NL on caspase-3/7 and PARP activation were also determined using luciferase assay and Western blot analysis. Figure 2C, D shows that NL induced a significant increase in caspase-3/7 activities and also elevated cleavage of pro-caspase-3 and PARP in PCa cells. Overall, these results demonstrate the apoptotic effect of NL against PCa cells.
NL inhibits STAT3 activation cascade in PCa cells
We next investigated the potential effect of NL on the STAT3 signaling pathway using diverse molecular biology techniques. First, we analyzed its effect on STAT3 phosphorylation by detecting phospho-STAT3 (p-STAT3) (Tyr 705) levels using Western blot analysis. We found that both constitutive p-STAT3 in DU145 and IL-6-stimulated p-STAT3 levels in LNCaP were substantially reduced upon NL treatment (Fig. 3A, C). Janus kinases (JAK) and Src families are the major upstream tyrosine kinases that can regulate STAT3 activation (42). In DU145 cells, both JAK1 and JAK2 phosphorylation levels were reduced by NL, while Src phosphorylation was not affected (Fig. 3B). Similar trend was also observed in LNCaP cells, in which NL reduced the phosphorylation of both JAK1 and JAK2 stimulated by IL-6 (Fig. 3C). These data indicate that the STAT3 inhibition caused by NL treatment may be due to downregulation of activation of upstream through JAK1 and JAK2 kinases.
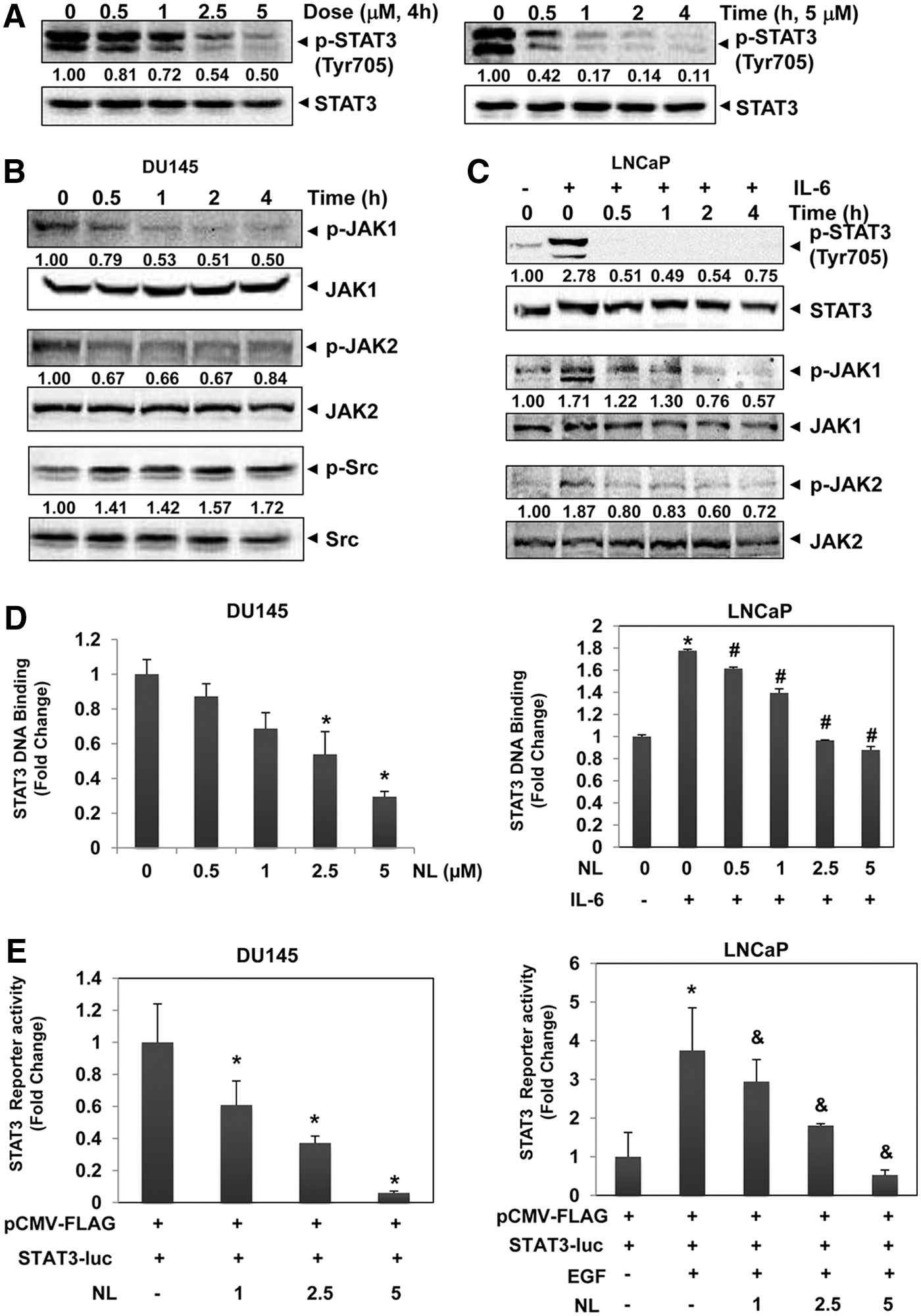
As STAT3 binds to DNA and thereby induces gene transcription after nuclear translocation, we next determined the potential effects of NL on both STAT3 DNA binding and transcriptional activities. As shown in Figure 3D, E, both constitutive and IL-6-stimulated STAT3 DNA binding ability and constitutive and EGF-induced luciferase reporter activities were significantly suppressed upon NL treatment, thereby demonstrating the negative effect of NL on the STAT3 signaling pathway.
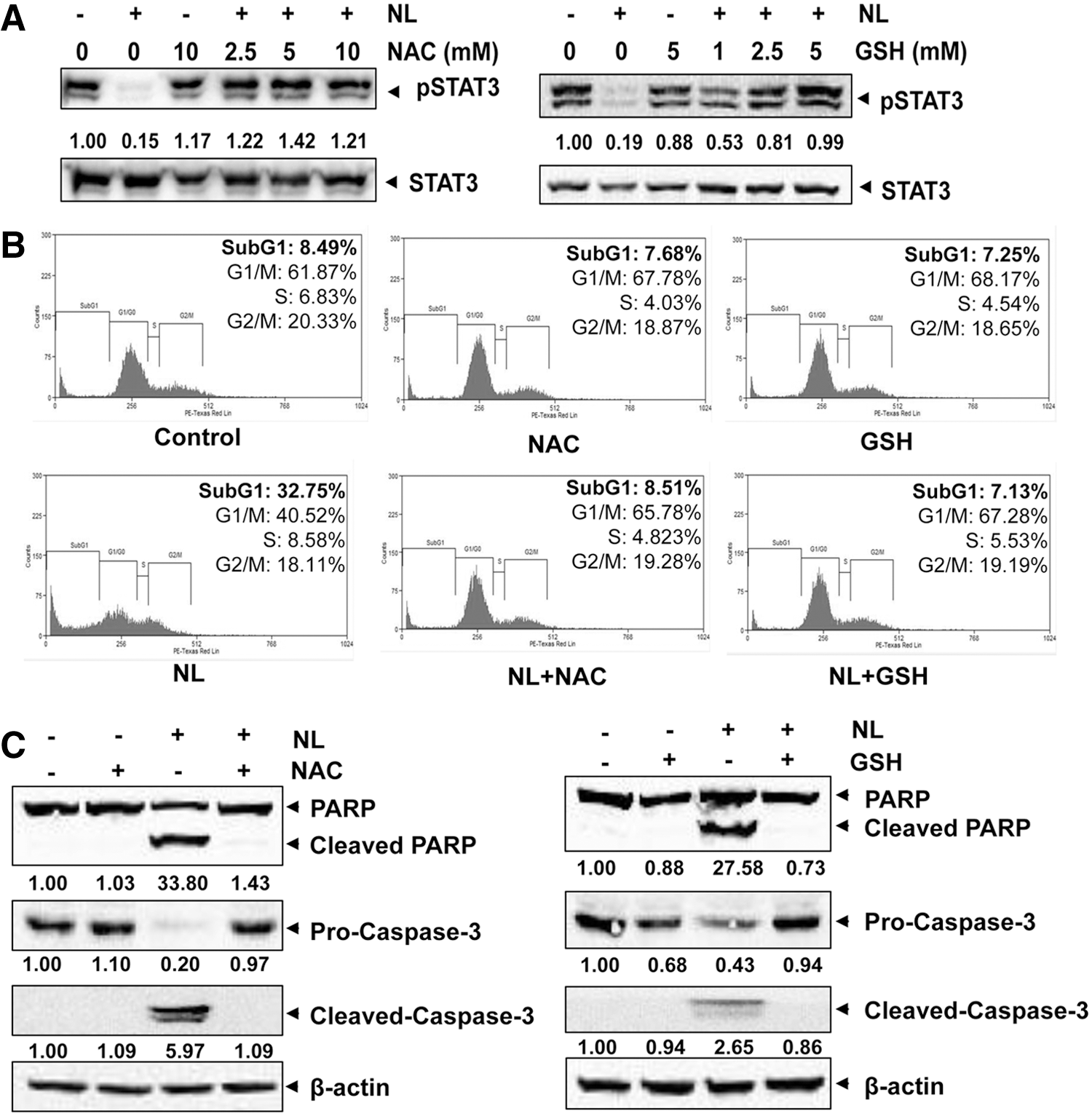
Antioxidants reverse NL-induced STAT3 inhibition and cellular apoptosis
N-acetyl-L-cysteine (NAC) and glutathione (GSH) are two major thiol-related antioxidants (7, 33). Interestingly, pretreatment of these two antioxidants substantially abolished STAT3 abrogation caused by NL treatment (Fig. 4A), thereby indicating that oxidative stress may be involved in the inhibitory effect of NL on STAT3 in PCa cells. Furthermore, NAC/GSH pretreatment was also found to significantly reduce cellular apoptosis induced by NL as observed by flow cytometric and Western blot analyses (Fig. 4B, C), further demonstrating that oxidative stress may also contribute to the observed proapoptotic effects of NL.
GSH/GSSG imbalance contributes to NL-induced reactive oxygen species production
To further validate whether NL can induce oxidative stress in PCa cells, reactive oxygen species (ROS) levels were measured in DU145 cells by staining with H2-DCFDA. We observed a significant increase of ROS production upon NL treatment (Fig. 5A), suggesting that NL can indeed induce oxidative stress in PCa cells.
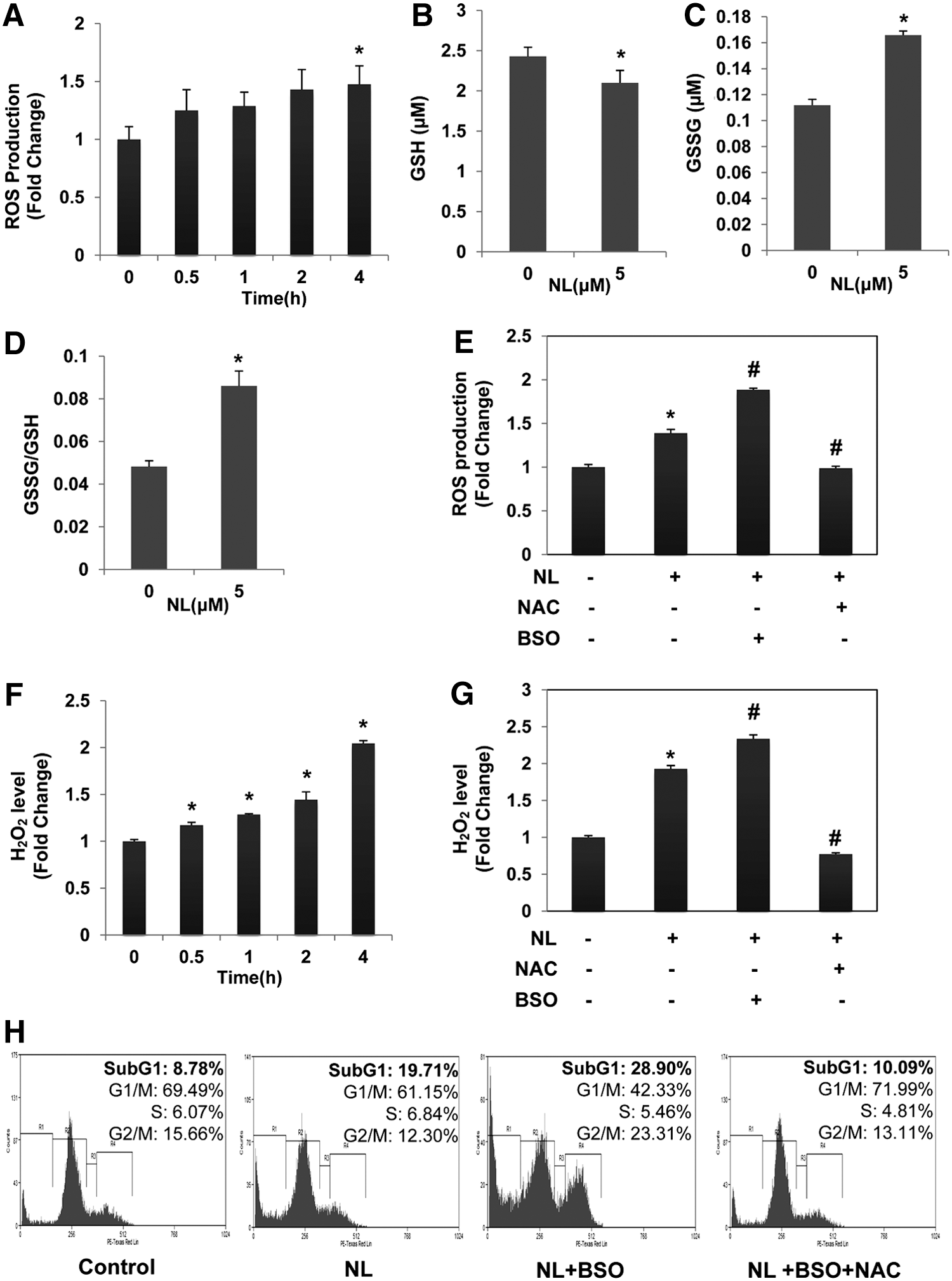
The GSH/GSSG system is one of the major intracellular antioxidant systems (27). The ratio of GSH to oxidized glutathione (GSSG) is an indicator of cellular oxidative stress (13). To explore the possible mechanism(s) of increased ROS production, we analyzed the effect of NL on the GSH/GSSG system. As shown in Figure 5B, C, a significant decrease of GSH and an obvious increase of GSSG were observed in DU145 cells upon treatment, thereby indicating that exposure of the cells to the drug resulted in an imbalance of the GSH/GSSG system. In addition, almost twofold elevation of GSSG/GSH ratio was observed (Fig. 5D), demonstrating that NL can induce oxidative stress in DU145 cells. To further investigate the role of GSH/GSSG imbalance in mediating NL-induced oxidative stress, both GSH synthesis blocker buthionine sulfoximine (BSO) and GSH prodrug NAC were employed. We found that pretreatment with NAC significantly prevented NL-induced ROS production, while BSO enhanced ROS production (Fig. 5E).
To further clarify the type of ROS involved, we measured the H2O2 level in NL-treated cell. As shown in Figure 5F, a time-dependent increase of H2O2 was observed, and the increase reached to almost twofold compared with the control level after 4 h (Fig. 5F). NL has no effect on superoxide (
NL inhibits the activity of glutathione reductase in PCa cells
Glutathione reductase (GR) catalyzes the reduction of GSSG to GSH to resist oxidative stress (8). As we observed a significant alteration in the GSH/GSSG ratio, we next investigated whether this imbalance may be due to the disturbance in GR activity. We treated the cells with NL and collected the cell homogenates to measure cellular GR activity. A time-dependent decrease of GR activity was observed (Fig. 6A). However, no change was observed in the GR protein expression level (Fig. 6B), indicating that GR inhibition by NL is not mediated through negative regulation of its protein expression.

To clarify how NL inhibits GR, we next investigated the interaction between GR and NL in a cell-free assay system. GR was incubated with NL in the presence of NADPH for 30 min, and then the substrate GSSG was added to initiate the reaction. As shown in Figure 6C, 70% of GR activity was lost upon NL treatment. These results demonstrate that NL directly suppresses GR by inhibiting its kinetic activity.
NL inhibits the expression of STAT3-regulated gene products
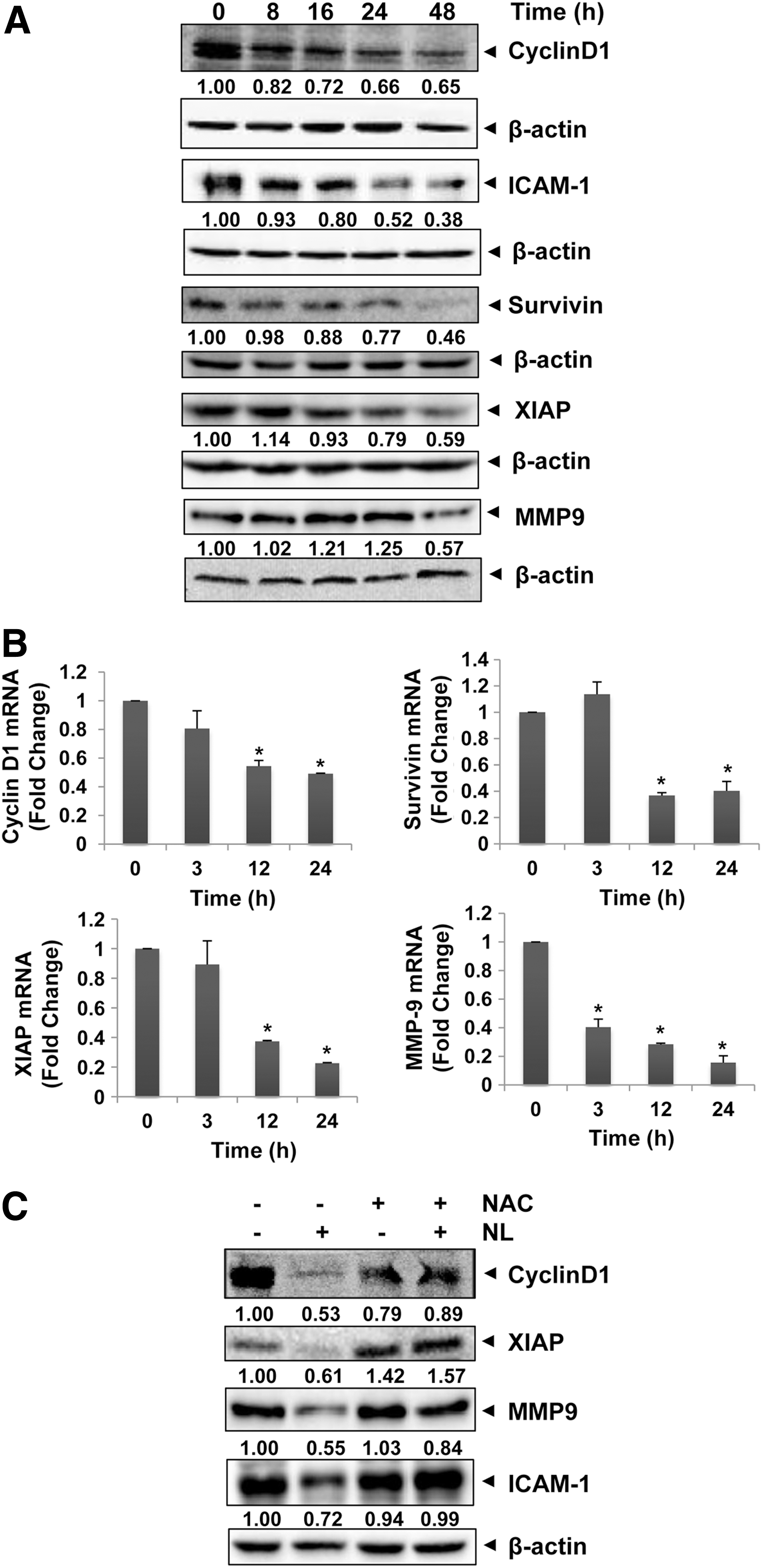
STAT3 regulates the expression of various oncogenic genes involved in tumor growth, angiogenesis, and metastasis (42). To analyze the effect of NL on STAT3-regulated proteins as well as genes, both Western blot analysis and real-time polymerase chain reaction (PCR) assays were performed. As shown in Figure 7A and Supplementary Figure S3, clear decreases were observed in the levels of various proteins such as survivin, Cyclin D1, and MMP-9 upon NL treatment, and similar decreases were also observed in the mRNA levels (Fig. 7B). These results provide evidence that NL treatment downregulates the expression of diverse STAT3-regulated genes. Interestingly, downregulation of these gene products could be reversed upon NAC pretreatment (Fig. 7C), thereby further demonstrating the role of oxidative stress in STAT3 inhibitory effects of NL.
NL suppresses prostatic intraepithelial neoplasia formation in transgenic adenocarcinoma of mouse prostate mouse model
Pharmacokinetics study indicates that NL showed a good exposure in mouse serum at 3 mg/kg with maximum concentration of 800 nM (Supplementary Fig. S4), suggesting that 3 mg/kg could be suitable dose for evaluating in vivo anticancer effect of NL in a TRAMP mouse model.
Prostatic intraepithelial neoplasia (PIN) formation by 4–10 weeks of age has been reported in a transgenic adenocarcinoma of mouse prostate (TRAMP) mouse model (17). To investigate whether NL suppresses PIN formation, 4-week-old mice were administered with vehicle or 3 mg/kg NL by oral gavage, 5 times a week for 8 weeks (n = 8), and then sacrificed. Representative hematoxylin and eosin (H&E) staining in dorsal lateral prostate (DLP) sections from nontransgenic mice, control, and NL-treated mice is shown in Figure 8A (i). DLP from nontransgenic mice exhibited typical acini with abundant eosinophilic intraluminal secretions. Compared with nontransgenic mice, DLP from vehicle-TRAMP exhibit high incidence of PIN, but with treatment of 3 mg/kg NL, PIN was significantly reduced by 35% [Fig. 8A (ii)]. In addition, none of the animals showed well-differentiated carcinoma (WDC) and poorly differentiated carcinoma (PDC) at this stage [Fig. 8A (ii)].

NL prevents the progression from PIN to PCa in a TRAMP mouse model
To study whether NL can prevent progression from PIN to PCa, 12-week-old TRAMP mice were treated with vehicle or 3 mg/kg NL by oral gavage, 5 times a week for 6 weeks (n = 8), and then sacrificed. Representative H&E staining in DLP sections from nontransgenic mice, control, and NL-treated TRAMP mice is shown in Figure 8B (i). Compared with nontransgenic mice, DLP from vehicle-TRAMP exhibit higher incidence of PIN and WDC. NL administration resulted in a 20% decrease of WDC incidence and a modest increase (30%) of PIN incidence compared with vehicle-TRAMP [Fig. 8B (ii)]. These results indicated that NL significantly suppressed the progression from PIN to WDC in TRAMP mice.
NL suppresses the growth of established PCa in vivo
To determine whether NL can suppress the growth of established PCa, 24-week-old TRAMP mice were treated with vehicle or 3 mg/kg NL by oral gavage, 5 times a week for 12 weeks (n = 8), and then sacrificed. At this age, H&E staining revealed greater incidences of PIN, WDC, and PDC in the vehicle-TRAMP mice than that in nontransgenic mice. Compared with vehicle-TRAMP mice, the DLP of the NL-treated TRAMP mice exhibited significantly lower incidence of PIN, WDC, and PDC [Fig. 8C(i) and (ii)]. For example, the incidence of the PIN in the DLP of NL-treated mice was lower by 10% in comparison with vehicle-TRAMP mice, while the incidences of the WDC and PDC were lower by 50% and 20%, respectively. Overall, these results indicate that NL administration significantly inhibited PCa progression as well as the tumor growth.
NL inhibits STAT3 phosphorylation and expression of Ki-67 in DLP of TRAMP mice
We next examined the potential effect of NL on STAT3 phosphorylation in the DLP of TRAMP mice. As shown in Figure 8D, DLP from vehicle-treated TRAMP mice exhibited a higher phosphorylation level of STAT3 compared with nontransgenic prostates, while NL intake resulted in a marked reduction of STAT3 phosphorylation. The inhibition on phosphorylation status of STAT3 in Group 3 was also observed in immunohistochemistry (IHC) experiments (Fig. 8E). We also investigated the effect of NL on the expression of proteins involved in cancer proliferation (Ki-67) and apoptosis (cleaved caspase-3). As shown in Figure 8D, E, we observed an increase in cleaved caspase-3 level and a decrease in proliferation biomarker Ki-67 expression compared with the control TRAMP group.
NL suppresses tumor growth and metastasis in a TRAMP mouse model
To investigate the effect of NL in the tumor growth and metastasis, 24-week-old TRAMP mice were treated with vehicle or 3 mg/kg NL by oral gavage, 5 times a week for 12 weeks (n = 8), and then sacrificed at 36 weeks. A significant (p < 0.05) difference in the tumor volume was observed in the NL-treated mice (Fig. 9A). C57BL/6 mice showed none of the features associated with PCa development as observed in TRAMP mice. In addition, no significant effects were observed in the body weight in TRAMP mice receiving the long-term feeding of NL when compared with vehicle-fed TRAMP mice (Fig. 9B). Interestingly NL reduces the metastasis of prostate tumor to lung and liver in TRAMP mice as evidenced by the reduced number of colonies in these organs (Fig. 9C).

Discussion
The aim of the present study was to elucidate the molecular mechanism(s) of anticancer effects of NL in PCa cell lines and to investigate its effects on the growth as well as metastasis of PCa in vivo in a TRAMP mouse model. NL exerted its potent inhibitory effects on both constitutive and IL-6-inducible STAT3 activation in PCa cells concomitant with suppression of upstream JAK1 and JAK2 kinases. This triterpene also modulated the GSH/GSSG system and thereby caused oxidative stress that mediated its observed STAT3 inhibitory effects in PCa cells. NL further downregulated the expression of various STAT3-regulated genes involved in antiapoptosis, metastasis, and angiogenesis. This downregulation led to the inhibition of proliferation, induction of apoptosis, reduction of invasion, and significant suppression of the growth and metastasis in a TRAMP mouse model (Supplementary Fig. S5).
We first noted that NL suppressed the viability and induced substantial apoptosis in two distinct PCa cell lines as evident by various apoptosis assays. The significant reduction in cellular viability and induction of apoptosis may be mediated through the downregulation of various proliferative and survival genes as observed by both real-time PCR and Western blot analysis. These observations are also consistent with the previous results obtained by Raja Singh et al.(35) who reported that NL can inhibit proliferation and induce apoptosis in PC-3 cells through altering molecules involved in IGF signaling via the PI3K/Akt pathway. In our study, the mitochondria may be involved in the apoptosis included by NL, as we observed a disturbance in the mitochondrial transmembrane potential with NL treatment (Supplementary Fig. S6). Moreover, in the advanced PCa, invasion and migration constitute the initial steps during the cancer metastasis process. We further found that NL treatment reduced the migratory and invasive potential of PCa cells, which can also be explained by the observed downregulation in the expression of various metastasis-related genes (MMP-9 and ICAM1).
We further observed that NL could also mitigate both constitutive and inducible STAT3 activation, respectively, in DU145 and LNCaP cells and these effects of NL on STAT3 phosphorylation correlated with the suppression of upstream protein tyrosine kinases (JAK1 and JAK2). We also observed that NL can also suppress both DNA binding and transcriptional activation of STAT3 in DU145 cells. These results are consistent with a previous study, in which NL was found to abrogate STAT3 activation in glioblastoma multiforme tissues (25), although the detailed molecular mechanism of its action on STAT3 signaling axis was not analyzed in this report.
Interestingly, we found for the first time that NL-induced STAT3 abrogation as well as cellular apoptosis could be reversed by ROS scavengers, NAC and GSH. ROS is maintained at low levels by cellular antioxidative systems under normal physiological conditions (23, 32); this low level of ROS can promote both cell survival and proliferation. However, when ROS accumulates at elevated nonphysiological concentrations, it can lead to apoptotic cell death through damage of cellular macromolecules such as DNA and proteins (38). As we noted that both NL-induced STAT3 suppression and apoptosis were abolished by ROS scavengers, we hypothesized that ROS may contribute to the STAT3 inhibitory and anticancer effects of NL. We indeed observed significant increase of ROS (to be specific, H2O2) in PCa cells upon NL treatment, supporting that NL can induce ROS production in DU145 cells. The ability of NL to induce ROS production was further demonstrated by the twofold increase of the GSSG/GSH ratio, which is an indicator of cellular oxidative stress.
Increased ROS production by NL might result from the imbalance of the GSH/GSSG system. In the GSH/GSSG system, GSH can reduce ROS through its thiol groups and then be oxidized into GSSG (7, 12). Disturbance of this system will limit its ability to scavenge ROS and thereby increase ROS production. We observed a significant GSH decrease and GSSG increase in NL-treated cells compared with untreated cells, suggesting that NL treatment indeed disturbed the GSH/GSSG system. GSH decrease was also observed in NL-treated DLP tissue from 24-week old TRAMP mice (Supplementary Fig. S7). In addition, replenishment of GSH with NAC abolished NL-induced ROS/H2O2 production as well as apoptosis, while inhibition of GSH synthesis with BSO had opposite effects, further demonstrating the contribution of glutathione system disturbances to NL-induced ROS production and observed anticancer effects.
The GSH level could be replenished by GR, which catalyzes the reduction from GSSG to GSH (32). In our study, GR inhibition may contribute to the GSH/GSSG imbalance induced by NL as a significant decrease of cellular GR activity was observed with NL treatment. GR inhibition may be due to a reduction in the GR activity as we did not observe a change in GR protein expression, but its kinetic activity was clearly inhibited by NL when incubated with the substrate NADPH and GSSG. In addition, the inhibition in GR kinetic activity by NL may require GR reduction. The disulfide bridge at the GR active site could be reduced by NADPH to generate two free cysteine thiols that in turn can reduce GSSG to GSH. We found that depletion of NADPH (only added at the point of measuring GR activity) from the assay buffer prevented NL to exert GR inhibitory effect (Supplementary Fig. S8A), suggesting a requirement of GR prereduction for GR inhibition by NL.
To further analyze whether NL binds at or near the active site where the natural substrate GSSG interacts, we also tried to perform a competitive inhibition assay. However, this assay was not successful as NL requires at least 30 min to inhibit GR activity (Supplementary Fig. S8B), while the substrate GSSG can immediately initiate the reaction. Thus, whether NL binds to the active site or other sites needs further in-depth analysis.
It has been also reported that ROS can directly oxidize the cysteine residues of JAK2 in the catalytic domain and therefore is able to suppress its kinase activity as well as downstream STAT3 activation (43). In our study, we found that the major ROS induced by NL, H2O2, can not only inhibit the phosphorylation of JAK2 and STAT3 but also significantly reduce the cell viability of PCa cells (Supplementary Fig. S9), supporting the notion that the oxidization of JAK2 may contribute to ROS-mediated STAT3 inhibition. Tyrosine phosphorylation site (Tyr-705) in STAT3 is surrounded by a number of cysteines in the C-terminal region (49). Protein modification of these cysteine residues with GSH in response to mild oxidative stress, which is also called S-glutathionylation, has also been reported to inhibit STAT3 activation through directly preventing the tyrosine phosphorylation of STAT3 (49). Thus, it is also possible that NL may abrogate STAT3 activation by inducing the S-glutathionylation process in PCa cells.
We next analyzed the potential effects of NL administration on tumor growth and metastasis in a TRAMP mouse model. This model is a well-established, spontaneously developing PCa model that mimics natural PCa development and progression, from PIN to highly invasive metastatic PCa in humans (18). We indeed found for the first time that NL can significantly inhibit tumor progression in 24-week TRAMP mice exposed to NL for 12 weeks and inhibit metastasis to both the lungs and liver. IHC analysis of tumor tissues from TRAMP mice indicated that this triterpene exerted its in vivo anticancer effects by downmodulating p-STAT3 expression that correlates well with its observed in vitro effects. We also noted that NL can decrease the expression of Ki-67 and increase the cleaved caspase-3 in TRAMP tumor tissues, which clearly demonstrates its significant antiproliferative and proapoptotic potential in PCa. These findings are quite interesting in view of a recent study in which supercritical extract of Azadirachta indica leaves was reported to inhibit tumor growth through undefined mechanism(s) in LNCaP-luc2 xenograft (48). Thus, based on our findings in transgenic mouse model, NL can be developed further as a potential chemopreventive and therapeutic agent for PCa.
NL has been extensively studied for its potential anticancer effects over the past decade and accumulating evidences suggest that it can significantly abrogate tumor initiation, progression, and promotion by diverse molecular mechanisms such as inhibition of cellular proliferation, induction of apoptosis, production of increased levels of antioxidants and carcinogen-detoxifying enzymes, suppression of migration, invasion, as well as angiogenesis, and negative regulation of diverse cell signaling pathways such as NF-κB, PI3K/Akt, MAP kinases, and Wnt/βcatenin (5, 10, 21). Interestingly, our data show for the first time that NL not only downregulates the activation of oncogenic STAT3 transcription factor through the induction of oxidative stress but can also inhibit tumor progression and metastasis in a transgenic mouse prostate model.
These findings are of great translational potential considering the critical role of STAT3 family of transcription factor in PCa pathogenesis and the reported efficacy as well as safety of NL as a potent anticancer agent against various malignancies. For example, several prior studies have also indicated that NL is a potential chemopreventive agent that can significantly suppress tumorigenesis without exhibiting any substantial toxicity and hence may form the basis of novel therapy for cancer. (5, 10, 19, 21). Accordingly, in our data on acute toxicity of NL (Supplementary Fig. S10), no significant adverse effects were noticed in the 3 mg/kg NL-treated mice. Overall, the findings of this study further emphasize the significant therapeutic potential of NL against PCa and provide a strong evidence for evaluating the safety and efficacy of NL in future clinical trials.
Materials and Methods
Reagents
RPMI 1640 and DMEM were obtained from Invitrogen (Carlsbad, CA). Fetal bovine serum (FBS) was from BioWest (Miami, FL). H2-DCFDA was from Molecular Probes (Carlsbad, CA). 3-(4,5- Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), propidium iodide (PI), crystal violet, NAC, BSO, GSH, and EGF were purchased from Sigma-Aldrich (St. Louis, MO). Antibodies against Cyclin D1, survivin, PARP, pro-caspase-3, GAPDH, and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies against phospho-STAT3 (Tyr 705), STAT3, phospho-Src (Tyr 416), Src, phospho-JAK1 (Tyr 1022/1023), JAK1, phospho-JAK2 (Tyr 1007/1008), JAK2, cleaved caspase-3, MMP-9, ICAM-1, XIAP, and GR were purchased from Cell Signaling Technology (Beverly, MA). IL-6 was obtained from ProSpec-TanyTechnoGene Ltd., (Rehovot, Israel). NL was purchased from Abcam (Cambridge, United Kingdom). Ten millimolar stock solution of NL was prepared by dissolving in DMSO and diluting in cell culture medium according to the required concentrations before treatment.
Cell culture
Human PCa DU145 and LNCaP cell lines were obtained from American Type Culture Collection (Manassas, VA). DU145 and LNCaP cells were cultured in RPMI-1640 and DMEM, respectively, with 10% FBS at 37°C in a humid atmosphere of 95% air and 5% CO2.
MTT assay
Cells were seeded in a 96-well plate. Twenty microliters of MTT (5 mg/ml) was added to each well and incubated for 2 h at 37°C. Thereafter, the purple formazan dye was dissolved in lysis buffer (20% SDS, 50% dimethylformamide), and the absorbance was measured by a microplate reader (Tecan, Durham, NC) under 570 nm.
Flow cytometric analysis
Cells were collected, fixed with 70% of ice-cold ethanol, and stained with PI (5 μg/ml) for 30 min. Cellular DNA content across the cell cycle was analyzed by a CyAn ADP flow cytometer (Dako Cytomation).
DNA fragmentation assay
DNA fragments were measured by the cell death detection ELISAPLUS kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions.
Wound healing assay
Cells were cultured in the inserts set in the bottom of the well (Ibidi GmbH, Martinsried, Germany). With cells grown to a confluent state, the inserts were removed to create a ∼500 μm gap without cell adhesion. Then, the cells in the nongap area were treated with or without NL for 8 h, followed by a 24-h incubation with 100 ng/ml CXCL12 (Prospec-TanyTechnoGene Ltd., Ness-Ziona, Israel). The widths of the gap were measured at time 0 and 24 h of CXCL12 incubation.
Invasion assay
Cells were seeded in the top chamber of a Bio-Coat Matrigel invasion assay system (BD Biosciences, San Jose, CA) in serum-free medium. After preincubation with or without 2.5 μM NL for 8 h, transwell chambers were then placed into the wells of a 24-well plate where either the basal medium only or basal medium containing 100 ng/ml CXCL12 had been added. After incubation for 24 h, cells on the upper surface of the chambers were removed by wiping the surface with cotton swab, while the cells in the bottom surface were stained with crystal violet solution, and counted under a microscope (Eclipse TS100; Nikon Tokyo, Japan).
STAT3 DNA binding assay
The binding of STAT3 to DNA was measured in nuclear extracts with an ELISA-based TransAM STAT3 assay kit (Active Motif, Inc. Carlsbad, CA) according to the manufacturer's instructions.
STAT3 luciferase reporter assay
Cells were seeded in 96-well plates and transfected with the STAT3 reporter plasmid by using lipofectamine 2000 (Invitrogen) according to the manufacturer's protocols. After 48 h post-transfection, cells were treated with NL and then lysed in luciferase lysis buffer (Promega, Madison, WI). The luciferase activity was measured with the Tecan microplate reader.
Caspase-3/7 activity assay
Caspase-3/7 activities were determined by a Caspase-Glo® 3/7 assay kit (Promega) according to the manufacturer's instructions.
Western Blot analysis
Western Blot analysis was performed as described previously (29).The densitometric analysis of the blots was done using ImageJ software. For phosphorylated proteins, the densitometric values were normalized to nonphosphorylated proteins; thereafter, results were expressed as fold change relative to the control. For other nonphospho proteins, the densitometric value was first normalized to β-actin, and then expressed as fold change relative to the control.
ROS detection
Cells were collected and washed with phosphate-buffered saline, followed by staining with 5 μM H2DCF-DA for 30 min. ROS generation was measured via the developed fluorescence product, dichlorofluorescein, an oxidized product of dye H2DCF-DA. Fluorescence was measured using the Tecan microplate reader at wavelengths according to the manufacturer's instructions.
H2O2 and glutathione measurement
The cellular H2O2 and glutathione levels were measured by ROS-Glo™ H2O2 Assay and GSH/GSSG-Glo Assay (Promega) according to the manufacturer's protocol.
Cellular GR determination
Cellular GR activity was measured with the Glutathione Reductase Assay kit (Abcam) according to the manufacturer's protocol.
Measurement of GR activity
All kinetics were carried out in GR assay buffer (50 mM potassium phosphate buffer, pH 7.6), which contains 200 mM KCl, 1 mM EDTA at 25°C. The standard assay mixture contained 100 μM NADPH and 0.5–1 U/ml GR. NL was allowed to react for at least 30 min before the addition of 1 mM GSSG, and the absorption decrease at 340 nm due to NADPH consumption was monitored over 3 min. Determination of the GR activity was achieved by measurement of the initial rates of disappearance of NADPH determined spectrophotometrically at 340 nm.
Quantitative real-time PCR
Total RNA was extracted from the cells using the Trizol reagent according to the manufacturer's instructions (Invitrogen), and then the real-time PCR was performed as described previously (29).
TRAMP mice breeding and genotyping
TRAMP mice breeding and genotyping were performed as described previously (39).
In vivo antitumor study
The inbred male TRAMP mice, 4 weeks old, were maintained in temperature-controlled rooms with a 12-h light/12-h dark cycle. All mice were weighed before start of experiment. The mice were then randomized into the following NL-treated groups (3 mg/kg b.w., oral gavage) and vehicle control groups (0.1% DMSO) (n = 8): Group 1: 4-week-old TRAMP mice were treated with NL or vehicle for 8 weeks; Group 2: 12-week-old TRAMP mice were treated with NL or vehicle for 6 weeks; and Group 3: 24-week-old TRAMP mice were treated with NL or vehicle for 12 weeks. Age-matched nontransgenic C57BL/6 mice were used as a negative control of TRAMP model through the 36-week period. Body weight and tumor size were recorded at the end of treatment period, and the tumor size was determined by Vernier caliper and calculated using the formula: [length × (width)2]/2. Mice were euthanized by CO2 inhalation, followed by cervical dislocation. Prostate gland was microdissected from the seminal vesicles under a stereomicroscope and fixed in 10% phosphate-buffered formalin for H&E and IHC analysis. A part of the DLP prostate was snap-frozen in liquid nitrogen for Western blot analysis.
Pathological grading of tumor tissues
Five-micrometer tissue sections from the DLP from individual mice of nontransgenic mice, vehicle, and NL-treated groups were stained with H&E. Thereafter, 10 randomly selected fields of H&E-stained sections were independently scored by two researchers for incidence corresponding to each pathological stage. Pathological grading was performed using criteria described and established previously (17, 40).
Statistical analysis
Data are expressed as the mean ± SD and vertical error bars denote the SD in the Figures. For comparing means of two groups, Student's t-test was performed to study the significance of differences between groups. A value of p < 0.05 was considered statistically significant. For comparing more than two groups, one-way ANOVA with Turkey post hoc analysis was performed. Fisher's exact test was used to compare incidence of PIN and adenocarcinoma in control versus NL-fed groups. p < 0.05 was considered significant.
Footnotes
Acknowledgments
This work was supported by an NUHS Bench to Bedside to Product grant to G.S. K.M.H. was supported by grants from the National Medical Research Council of Singapore, Biomedical Research Council of Singapore, and the Singapore Millennium Foundation. K.S.A was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (NRF-2015R1A4A1042399). A.P.K was supported by grants from the National Medical Research Council of Singapore [R-713–000-177–511], the National Research Foundation Singapore, and the Singapore Ministry of Education under its Research Centres of Excellence initiative to Cancer Science Institute of Singapore, National University of Singapore, and by the NCIS Yong Siew Yoon Research Grant through donations from the Yong Loo Lin Trust.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.