
Abstract
Alteration in neurotrophic factors support and antioxidant defenses in the central nervous system (CNS) along with deficit of ferritin have been associated with idiopathic Parkinson's disease (PD). The objectives were to analyze in the cerebrospinal fluid (CSF) of patients with PD and controls the following: (i) the levels of the neuroprotectant factors glial cell line-derived neurotrophic factor, persephin, neurturin, and brain-derived neurotrophic factor, (ii) the levels of transforming growth factor-β1 (TGFβ1) and transforming growth factor-β2 (TGFβ2), proinflammatory factors, (iii) the activity of the antioxidant enzymes glutathione peroxidase (GPx), glutathione reductase (GR), glutathione-S-transferase (GST), catalase, superoxide dismutases (SODs), and peroxiredoxins (PRDxs), and (iv) ferritin levels. The study revealed that, among neurotrophic factors, only TGFβ1 levels were found to be enhanced in patients with PD (early, p < 0.05; advanced, p < 0.02). Regarding antioxidant enzymes, the activity of GPx, catalase, and PRDxs, all hydrogen peroxide scavengers, was found to be significantly reduced in patients (GPx, p < 0.001; catalase, p < 0.01; PRDxs, p < 0.01, one-way analysis of variance). Finally, ferritin content in CSF was significantly diminished over time in patients (early, p < 0.01, −49%; advanced, p < 0.001, −80.7%). Our observations lead to the hypothesis that parkinsonian patients suffer from a serious disturbance of redox state in the CNS, as evaluated through the CSF, characterized by reduced hydrogen peroxide scavenging and iron storage. Antioxid. Redox Signal. 23, 893–898.
Introduction
A
Regarding neurotrophic support, neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and ligands belonging to the glial cell line-derived neurotrophic factor (GDNF) family are considered to facilitate the survival of dopaminergic cells, neurons that die in the course of PD (1, 2). In contrast, members of the transforming growth factor (TGF) family such as TGFβ1 and TGFβ2 are potent regulators of inflammatory responses in the CNS (6) and they seem to be produced in astrocytes and microglial cells as part of the neuroinflammatory response of the disease. Changes in all of these ligands in the cerebrospinal fluid (CSF), a fluid in close contact with nerve tissue and considered as a good witness of neurodegeneration, deserve to be further analyzed.
Oxidative stress, defined as a disbalance between the production of reactive oxidative species and antioxidant mechanisms, has been postulated to underlie parkinsonian neurodegeneration (7). Oxidative stress can be detected in blood, brain tissue, and CSF of PD patients. CSF antioxidant defenses are known to be seriously affected because the activity of important enzymes such as glutathione-S-transferase (GST), the ferroxidase ceruloplasmin, and the peroxidase DJ-1 or PARK7 is reduced (8). However, either conflicting results or no changes have been reported as to other CSF antioxidant enzymes such as superoxide dismutases (SODs), superoxide anion scavengers, or the hydrogen peroxide (H2O2) scavengers, glutathione peroxidase (GPx), glutathione reductase (GR), and catalase (8).
Alterations in neurotrophic factors support and antioxidant defenses along with deficit of ferritin have been associated with idiopathic Parkinson's disease. After analyzing the cerebrospinal fluid, less neurotrophic support was not observed as far as glial cell line-derived neurotrophic factor family ligands and brain-derived neurotrophic factor levels are concerned. Only the proinflammatory ligand transforming growth factor-β1 (TGFβ1) was enhanced. We also note that the enzymatic activity of hydrogen peroxide scavengers, but not superoxide anion scavengers, is reduced, and ferritin content is reliably diminished. Our observations indicate that parkinsonian patients suffer from a serious disturbance of redox state in the central nervous system, with altered hydrogen peroxide scavenging and iron storage.
In addition, it has been proposed that cell death in the substantia nigra in PD patients is associated with increased iron content together with decreased levels of ferritin, iron storage protein (4). Ferritin is known to bind about one third of iron in the brain, and low ferritin levels would enhance free iron. Changes of ferritin in CSF in PD patients are not well known. The objectives of this study were to analyze in the CSF of PD patients and controls the following: (i) the levels of the neuroprotectant factors GDNF, persephin, neurturin, and BDNF, (ii) the levels of TGFβ1 and TGFβ2, proinflammatory TGF family ligands, (iii) the activity of the antioxidant enzymes GPx, GR, GST, catalase, SODs, and peroxiredoxins (PRDxs), and (iv) ferritin levels. Clinical situation was evaluated through the Hoehn and Yahr scale, Unified Parkinson's Disease Rating Scale (UPDRS), and years of disease.
Results and Discussion
First, we verified whether the main clinical characteristics were similar between the PD and control groups. All parameters were similar except for hypertension (p < 0.01), more frequent in patients with PD relative to controls, as shown in Table 1.
Mean ± SEM. Statistical comparisons were carried out with the χ2 test (qualitative variables) or the Student's t-test (quantitative variables).
PD, Parkinson's disease; SEM, standard error of the mean.
Second, we quantified levels of GDNF family ligands (GDNF, persephin, and neurturin) and BDNF, all of them considered as neuroprotectant trophic factors, through the enzyme-linked immunosorbent assay (ELISA). Levels of GDNF, persephin, and BDNF in CSF were not found to be different among groups, and neurturin was undetectable likely because its level is below the detection range of the ELISA kit, as shown in Table 2. Then, levels of the TGF family ligands, TGFβ1 and TGFβ2, considered as neuroinflammatory factors, were also measured through ELISA. One-way analysis of variance (ANOVA) indicated a significant group effect on TGFβ1 values (F 2,36 = 4.7, p < 0.02). TGFβ1 values in CSF were significantly enhanced in both early and advanced PD patients relative to controls (early PD, p < 0.05; advanced PD, p < 0.01, Newman–Keuls), as shown in Table 2. CSF changes in patients were found to be enhanced over time, because TGFβ1 values in early PD patients were significantly lower than those observed in advanced PD patients (p < 0.05). Following statistical correlations between TGFβ1 levels and clinical parameters, a positive correlation was found between TGFβ1 and motor UPDRS III (r = 0.55, p < 0.001, Pearson's test), as shown in Figure 1. No significant differences between patients and control subjects were observed on TGFβ2 values.

Mean ± SEM.
p < 0.01 versus controls.
p < 0.05 versus advanced patients.
p < 0.001 versus controls.
BDNF, brain-derived neurotrophic factor; GDNF, glial cell line-derived neurotrophic factor; nd, not detectable; TGFβ1, transforming growth factor-β1; TGFβ2, transforming growth factor-β2.
Third, to further evaluate the role of CSF antioxidant enzymes in PD patients, the activity of GPx, GR, GST, catalase, SODs, and PRDxs was measured through ELISA or the horseradish peroxidase (HRP) method (9). GPx activity was found to be significantly reduced in PD patients, as indicated by the one-way ANOVA (group effect, F 2,36 = 44.1, p < 0.001). PD patients showed lower GPx levels relative to controls (early PD, t = 6.8, p < 0.001; advanced PD, t = 6.8, p < 0.001), as shown in Figure 2. Percentage mean GPx activity was found to be strongly reduced in both groups of PD patients (early PD, −84.6%; advanced PD, −85.5%). Catalase activity was also found to be significantly reduced in PD patients (one-way ANOVA, F 2,36 = 4.7, p < 0.015). PD patients showed lower activity levels than controls (early PD, t = 2.1, p < 0.05; advanced PD, t = 2.8, p < 0.017). Mean catalase activity was reduced by 59.4% and 78.2% in early and advanced PD patients, respectively. Finally, activity of PRDxs was also found to be significantly reduced in PD patients (one-way ANOVA, group effect, F 2,36 = 4.9, p < 0.01). Activity of PRDxs was found to be reduced in both groups of PD patients relative to controls (early PD, t = 2.2, p < 0.05; advanced PD, t = 2.7, p < 0.01). Mean PRDxs activity was reduced by 60% and 68% in early and advanced PD patients, respectively. No changes were observed in the activity of the remainder enzymes, GR, GST, and SODs, as observed in Figure 2. No correlations with clinical parameters were observed.

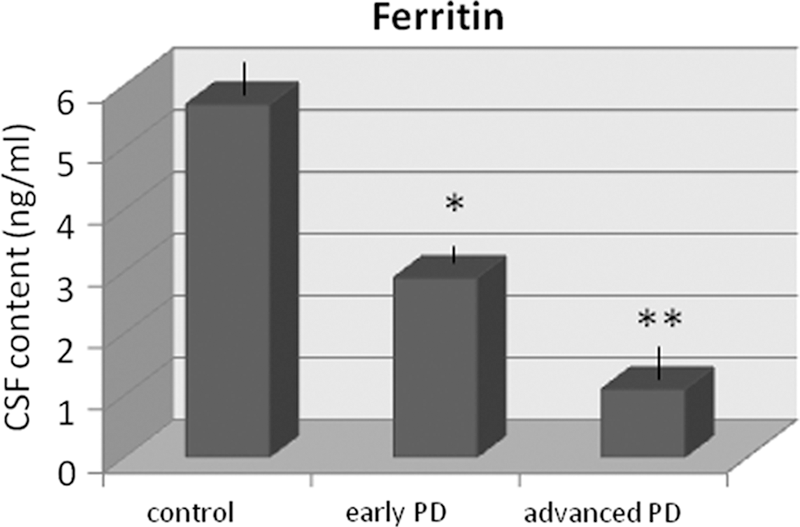
Four, to further study the metabolism of iron storage in PD patients, ferritin content was quantified in CSF of patients and controls through ELISA. Ferritin level in CSF was significantly altered in patients (one-way ANOVA, group effect, F 2,36 = 14.3, p < 0.001). Thus, CSF ferritin was significantly reduced in early (t = 2.7, p < 0.013) and advanced PD patients (t = 5.9, p < 0.001) relative to controls. Percentage levels were reduced by 49% and 80.7% in early and advanced PD patients, respectively (Fig. 3), indicating that changes worsened over time. No correlations with clinical parameters were observed, except for the Hoehn–Yahr stage (r = −0.51, p < 0.05; Pearson's test).
Finally, the Tibbling-Link index was quantified to confirm that CSF protein levels were not influenced by blood levels due to blood–brain barrier inflammation. The Tibbling-Link index in the PD group was lower than 0.7 in every case, and it was observed not to be different relative to the control group (0.45 ± 0.01 PD; 0.42 ± 0.01 controls). Hence, no inflammation of the blood–brain barrier was detected in PD patients, in accordance with previous data (5).
Idiopathic PD has been associated with altered neurotrophic support. Our study reveals that at least the neuroprotectant mediators GDNF, persephin, and BDNF are not altered in CSF of parkinsonian patients. Hence, it can be concluded that neurotrophic factors support is not altered in PD patients as far as the GDNF family ligands and BDNF are concerned. Regarding GDNF, the most studied GDNF family ligand, several authors have already pointed out that this factor is not modified in PD patients because the content of GDNF in nigrostriatal regions or the expression of its receptors cRet and GRFα1 is not found to be altered in patients (3). In this context, GDNF, persephin, and BDNF are known to exert neuroprotective effects in animal models of PD, a fact that has prompted numerous suggestions that these neurotrophic factors may be therapeutic tools to treat PD (2). In contrast, among all neurotrophic factors which were analyzed in this study, only TGFβ1 was found to be elevated in CSF of patients. Since TGFβ1 is considered as a neuroinflammatory factor, it can be hypothesized that changes of TGFβ1 in CSF could be accounted for by an ongoing neuroinflammatory process (6). Of note is that statistical correlations of this proinflammatory mediator point to an important role for neuroinflammation in the clinical evolution of PD because TGFβ1 levels in CSF are enhanced in parallel with motor deficits, as evaluated through UPDRS III.
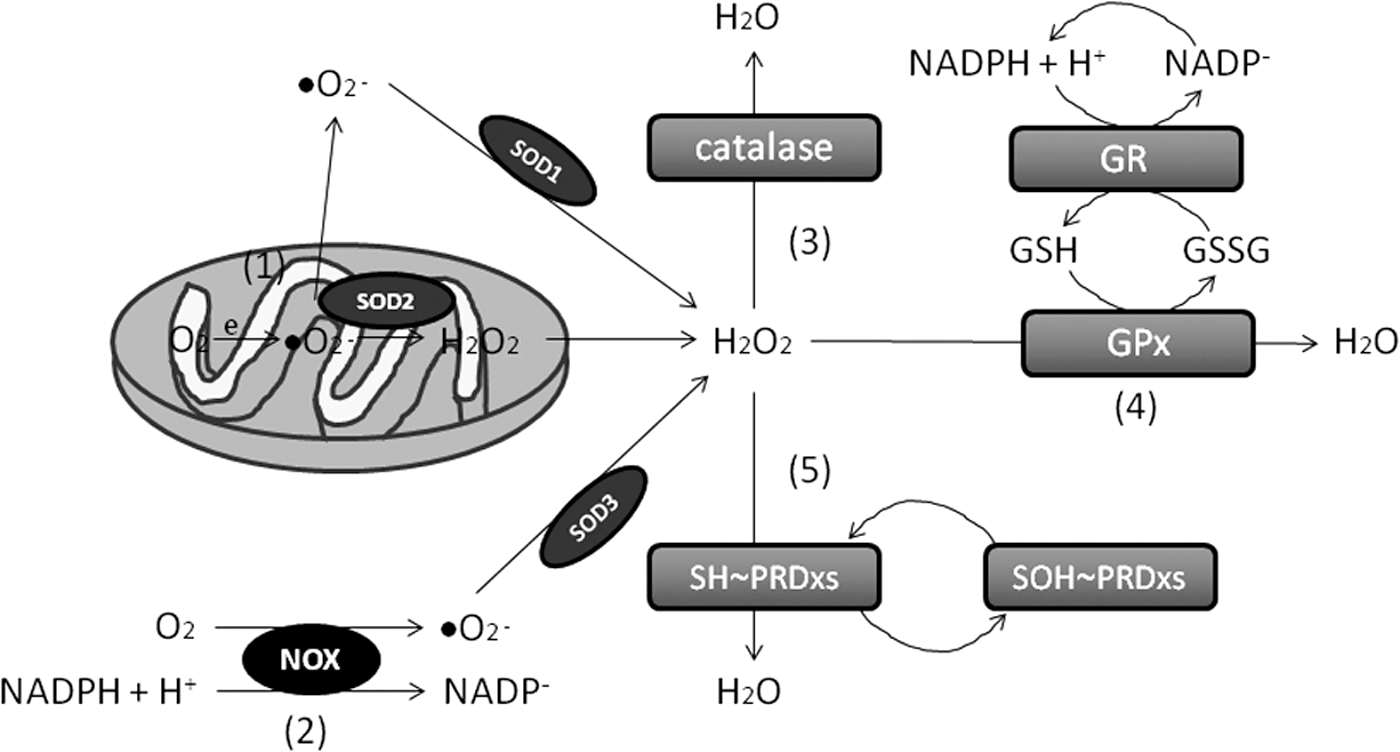
Idiopathic PD has also been associated with oxidative stress (7). After analyzing the enzymatic activity of superoxide anion and H2O2 scavengers, the data reveal a disturbance of H2O2 scavenging, but not superoxide anion scavenging, as manifested through a reliably reduced activity of GPx, catalase, and PRDxs, without changes in SODs (Fig. 4). GPxs are selenoproteins that reduce peroxides in water. They also reduce lipid hydroperoxides to their corresponding alcohols. Catalases are ubiquitous enzymes that mediate the decomposition of H2O2 in water. PRDxs are cysteine-dependent peroxidases that react with hydrogen peroxide and hydroperoxide substrates. Data fit well with the presence of peroxidation stress due to reduced antioxidant defenses related to hydrogen peroxide scavenging, and it can be hypothesized that neurons would be less protected from oxidative damage in patients with PD.
Finally, our data indicate that ferritin content is reliably and progressively reduced in CSF of PD patients, a novel result to our knowledge because ferritin was found to be reduced in the brain tissue of PD patients rather than in CSF (4). Ferritin is known to bind about one third of iron in the brain, and low ferritin levels would then enhance free iron. In this context, it has been proposed that cell death in the substantia nigra in PD patients is associated with increased iron content together with decreased levels of ferritin (4). Taken together, the enzymatic and ferritin data indicate a serious alteration of redox homeostasis in patients' CSF, characterized by deficit of hydrogen peroxide scavenging and iron storage.
To sum up, the findings indicate that CSF analysis of patients with PD does not reveal a deficit of neurotrophic factors support as far as the neuroprotectant GDNF family ligands and BDNF are concerned. Among the studied neurotrophic factors, only the proinflammatory TGFβ1 is observed to be augmented in patients. Regarding antioxidant defenses, the reliable decrease in the activity of GPx, catalase, and PRDxs without changes in SODs indicates altered hydrogen peroxide scavenging, likely reflecting the presence of peroxidation stress. The content of ferritin, iron storage protein, in CSF is also found to be reliably diminished. The enzymatic and ferritin data indicate a serious disturbance of redox homeostasis in patients with PD, characterized by deficit of hydrogen peroxide scavenging and iron storage.
Notes
Study participants
Patients suffering from PD and diagnosed by a neurologist based on single-photon emission computed tomography (SPECT) and clinical information were included in this study. Patients were classified and scored according to the Hoehn–Yahr stages and UPDRS. Following Hoehn–Yahr criteria, patients were divided into the early PD group (Hoehn–Yahr stages 1 and 2) and advanced PD group (Hoehn–Yahr stages 3 and 4). UPDRS comprised total UPDRS, motor symptoms or UPDRS III, and disease complications or UPDRS IV. All subjects were nonsmokers or nonalcohol drinkers (smoking and alcohol drinking modify biochemical CSF parameters) (5). Smoking was defined as current smoker who consumed cigarettes on a daily basis or occasional smokers. Alcohol use was defined as drinking > 210 g per week. Control subjects were recruited from either patients' relatives or volunteers subjected to intradural anesthesia for surgery in the Service of Surgery of Hospital Macarena and without any neurological disorder. Individuals presenting with any of liver, renal, and cardiac dysfunctions, malabsorption, autoimmune diseases, diabetes mellitus, rheumatoid arthritis, AIDS, and infectious conditions (because oxidative stress markers in peripheral fluids may be altered in such conditions), were excluded from both PD and control groups. Diabetes mellitus and rheumatoid arthritis were defined based on WHO criteria.
The following clinical information was gathered from each patient: age, sex, body weight, hypertension, dyslipidemia, coffee drinking, smoking, taking of vitamin A/vitamin E supplement, statins and aspirins, daily levodopa dose, type and dose of dopamine agonists, and rasagiline. Hypertension was diagnosed when blood pressure repeatedly exceeded 140 mmHg (systolic) and/or 90 mmHg (diastolic) or when a subject was taking antihypertensive medication to control hypertension. Use of vitamin A/E was defined as daily intake of either vitamin A or vitamin E. Drinking coffee was defined as daily intake of at least 300 ml of coffee.
CSF collection
CSF was collected using lumbar puncture. Five milliliters of CSF was collected and stored in polypropylene tubes (Eurotube), protected by light, and rapidly aliquoted, coded, and frozen at −80°C for further studies. One-milliliter collection in a glass tube was employed to observe the absence of traumatic puncture and to quantify red cells before storing. CSF with excess of red cells was discarded (>500 red cells/μL).
Tibbling-Link index
CSF was collected in polypropylene tubes. Simultaneously, blood samples were obtained (5). CSF samples were centrifugated at 450 g for 10 min and the supernatant was recovered. Then, paired CSF and serum were aliquoted and stored under biobanking conditions until studies were performed. Albumin and IgG were quantified by standard immunochemical nephelometry in serum and CSF samples (Siemens BN II system). IgG indexes were calculated as previously described (5). It has been defined that IgG oligoclonal bands are not detected in patients with IgG indexes smaller than 0.45; however, indexes higher than 0.77 reflect intrathecal IgG synthesis.
ELISA measurement
CSF aliquots were unfrozen and sonicated with homogenizing solution (150 mM NaCl, 50 mM HEPES, 1 mM phenylmethylsulfonyl fluoride, 0.6 μm leupeptin, and 1% Triton X-100, pH 7.4). For measuring levels of GDNF, persephin, neurturin, BDNF, TGFβ1, and TGFβ2 in CSF, commercial kits were used (GDNF Emax ® ImmunoAssay System, Promega; Human Persephin ELISA Kit, Wuhan EIAab Science Co. Ltd.; Human Neurturin ELISA Kit, Wuhan EIAab Science Co. Ltd.; Human BDNF ELISA kit, Boster Biological Tech; TGFβ1 Emax Immunoassay System, Promega; TGFβ2 Human ELISA kit, Abcam) following manufacturer's instructions.
Commercially available ELISA kits were used for evaluating the activity of the antioxidant enzymes GR, GPx, GST, catalase, and SOD. ELISA kits were purchased from Bioxytech (GR-340 assay kit; Oxis International, Inc.) and Biovision (GPx activity kit 762-100, GST Assay Kit K263-100, Catalase activity Assay Kit, SOD activity Kit K335-100). The GR-340 assay is based on the oxidation of NADPH to NADP+ catalyzed by a limiting concentration of GR. One GR activity unit is defined as the amount of enzyme catalyzing the reduction of 1 μmol of GSSG per minute at pH 7.5 and 25°C. Biovision glutathione peroxidase assay Kit measures GPx activity through a coupled reaction with GR. The GST colorimetric activity assay kit is based on the GST-catalyzed reaction between GSH and CDBN, GST substrate. As for catalase, this enzyme reacts with hydrogen peroxide to produce water and oxygen, and the uncoverted hydrogen peroxide reacts with the probe to produce a product that can be measured at 570 nm. The inhibition activity of SOD of the rate of the reduction with a superoxide anion is measured by the SOD activity assay kit. Finally, peroxiredoxin activity was measured following the HRP method, based on measurement of peroxide-dependent PRDx activity with hydrogen peroxide using the HRP competition assay (9).
Statistics and ethics
We obtained CSF from 29 patients, comprising four Hoehn–Yahr stages (stage 1, n = 12; stage 2, n = 12; stage 3, n = 8; stage 4, n = 5), and 21 controls. Stage 1 and 2 patients were considered as the early PD group (n = 24) and stages 3 and 4 patients were considered as the advanced PD group (n = 13). One-way ANOVA was used for statistical comparisons (n = 13/group), followed by post hoc Newman–Keuls' test or Student's t-test. Correlations were performed with the Pearson's test or the single linear regression test. Two group comparisons were carried out with the Student's t-test. If needed, normalization was verified with the Shapiro–Wilk test. Informed consent forms under a protocol approved by the University of Seville and Macarena Hospital internal ethics and scientific boards were obtained from all the subjects, and the subjects' consent was obtained according to the Declaration of Helsinki (BMJ 1991; 302: 1194).
Footnotes
Acknowledgments
The authors thank Mara Guerra and Silvia Castellano (BIO127 lab, University of Seville) for their excellent technical assistance; Dr. Luis Capitán for his continuous support (Vicedean, School of Medicine of Seville); Dr. Guillermo Izquierdo and Dr. Castiñeiras for allowing the use of the facilities of the Services of Neurology and Surgery, respectively (Hospital Macarena, Seville); Dr. Maria-Isabel Garcia-Sánchez and the Biobanco Hospitalario Macarena (National Biobank Network, Carlos III Health Institute RD09/0076/00080) for their support in the samples collection procedure and storage and Tibbling-Link measures; and Dr. Cinta Calvo and the Service of Nuclear Medicine (Hospital Macarena, Seville) for SPECT analyses. The authors are most grateful to all patients and their partners as well as control subjects who participated in this study. This study was supported by grants to EF by Junta de Andalucia (BIO127).