Abstract
This editorial shortly summarizes the highlights described in the Forum, novelties about selenoproteins. Two articles describe the selenoprotein biosynthesis and the role of so far identified proteins involved, including that of selenocysteine-β-lyase, which also may link selenoproteins to energy metabolism. Novel and, in part, unexpected functions are reviewed. Thioredoxin reductase 1 (TrxR1) can change from an anti- to a pro-oxidant and appears to be involved in the regulation of the Nrf2/Keap1 system. Methionine sulfoxide reductase B1 (MsrB1) catalyzes a novel posttranslational protein modification. The membrane proteins, Sel K,S,T,N, and I, form selenylsulfide bonds leading to the formation and stabilization of protein complexes required for protein trafficking. By this mechanism, selenoprotein K (SelK) supports palmitoylation of membrane-associated proteins. Thus, selenium and selenoproteins obviously have functions by far exceeding that of counteracting oxidative stress and even also catalyzing oxidoreductive processes. Antioxid. Redox Signal. 23, 757–760.
S
The human genome contains 25 genes for selenoproteins from which the sequence is known. Surprisingly, however, we still do not know the function of all of them, and even the function of relatively well-characterized selenoproteins can change under certain circumstances. In this Forum, therefore, most striking novelties on selenoprotein biosynthesis, including the possible role of selenocysteine-β-lyase (Scly) not only in the biosynthesis of selenoproteins but also in linking selenoproteins to energy metabolism, are summarized. Regarding novel functions, the Forum particularly focuses on thioredoxin reductase 1 (TrxR1) (3), methionine sulfoxide reductase B1 (MsrB1) (6), selenoprotein K (SelK) (5), and other membrane-associated selenoproteins (SelS, SelI, SelN, and SelT) (7).
Selenoprotein Biosynthesis and Related Topics
The second article of this Forum gives an update of selenoprotein biosynthesis (2). With the detection that Sec is encoded by the stop codon TGA in 1986 (4), it had become clear that the incorporation of Sec into proteins must be complex. The requirement of a Secis (Selenocysteine Insertion Sequence) element to recognize UGA in the mRNA as codon for Sec was found relatively soon. In eukaryotes, the Secis is a characteristic stem-loop structure in the 3′-untranslated region (3′UTR) of the selenoprotein mRNAs. Then, the existence of a specific tRNA (tRNA[Ser/Sec]) for Sec was recognized. Since then, many more essential factors have been identified (2). The Secis-binding protein 2 (SBP2) binds to the Secis, triggers the transfer of the 3′UTR to the ribosome, and directs the biosynthetic complex formation with other factors in a still unknown sequence of events. In this study, Bulteau and Chavatte describe two models for selenoprotein biosynthesis comprising the functions of UGA, Secis, SBP2, the Sec tRNA[Ser/Sec]-EFSec-GTP ternary complex, the RNA-binding protein, eL30, and two other Secis-binding proteins, nucleolin and the eukaryotic initiation factor 4A3 (eIF4A3) (2).
An important player in selenoprotein biosynthesis appears to also be the Scly, an enzyme, which specifically breaks down Sec into alanine and selenide (9). This way, selenium derived from Sec of selenoproteins can be recycled. Selenide becomes phosphorylated and is transferred to the seryl residue bound to the tRNA(Ser)Sec, thus generating Sec (2). However, the Se recycling pathway remained neglected until the knockdown of Scly in HeLa cells revealed its involvement in selenoprotein biosynthesis. Disruption of Scly in mice led to even more surprising results (9). Selenoproteins in livers of the selenium-adequately fed mice were not strikingly affected with the exception of SelS, which was decreased. On a low-selenium diet, however, hepatic GPx1, SelS, and plasma SePP1 levels were lower in Scly KO mice than in WT mice. More interestingly, knockout of Scly led to hyperinsulinemia, hyperleptinemia, glucose intolerance, and hepatic steatosis in mice on a selenium adequate diet. This phenotype, which was aggravated under selenium restriction, resembles the metabolic syndrome. In addition, mice developed obesity, fatty liver, and hypercholesterolemia. In their present article, Berry and colleagues raise the question whether the obvious role of Scly in lipid metabolism could be independent of its function in selenoprotein biosynthesis and, indeed, report on numerous findings that point to a link between selenium and energy metabolism (9). The results of the two studies with Scly KO mice are quite unexpected and, in part, provocative and might pave the way for a completely new field in the biology of selenium and selenoproteins.
Taken together, the process of selenoprotein biosynthesis is much more complicated than the usual protein biosynthesis. It requires specific factors, some of which, for example, eIF4A3, are regulated by selenium themselves. It is therefore hard to believe that nature developed such a complex mechanism just to create some antioxidant enzymes. The preoccupation that selenium is primarily in charge of antioxidant defense was first corroborated by the discovery of glutathione peroxidase being a selenoprotein, but soon challenged by the selenoprotein nature of the deiodinases (4). Later, it became clear that even a peroxidase, namely GPx4, does not always act as a peroxidase but also moonlights during sperm development to a nonenzymatic structural protein in the midpiece of mature spermatozoa. More examples of nonantioxidant roles of selenoproteins rise on the horizon and are compiled here: (i) the functional switch of TrxR1 after modification of the Sec (3); (ii) the coordinated action of MsrB1 and Micals in the regulation of the function of actin (6); (iii) the interaction of selenoproteins with proteins in membranes (7); and (iv) the role of SelK in the palmitoylation of proteins in the endoplasmic reticulum (ER) membrane (5).
Thioredoxin Reductase-1
TrxR1 is a selenoprotein well characterized as an oxidoreductase. It is part of the Trx system, which had first been discovered to provide the reduction equivalents for ribonucleotide reductase and, later, to protect against oxidative stress, thereby maintaining cell proliferation and viability. With increasing knowledge about the requirement of redox processes in cell biology, the Trx system was recognized as a regulator in protein folding and signal transduction (3). In the Nrf2/Keap1 system, TrxR1 plays an unexpected and surprising dual role. Clearly, a robust activation of Nrf2 in TrxR1-depleted organisms is observed. Nrf2 is activated by an oxidative/electrophilic modification of Cys151 in Keap1. By keeping Keap1 in the reduced state, TrxR1 counteracts activation of the pathway or, in other words, cooperates with Keap1 in maintaining a balanced Nrf2 activity. To guarantee this balance, however, TrxR1 activity also has to be regulated. An inhibition can be reached by the same agents that modify Keap1. These compounds attack the highly accessible Sec residue in TrxR1, thus inhibiting TrxR1 activity. This way Keap1 remains modified and Nrf2 can work [for details, see Ref. (3)]. Moreover, derivatization by electrophiles not only inactivates TrxR1 but also leads to its transformation from an anti- to a pro-oxidant, the so-called selenium-compromised thioredoxin reductase-derived apoptotic protein (SecTRAP). If it turns out that only Sec-containing TrxR1 is involved in Nrf2 modulation, this aspect, as the authors state, would indeed help to explain why the enzyme has evolved to be a selenoprotein (3).
Methionine Sulfoxide Reductase-B1
A novel post-translational modification for the regulation of protein functions is the reversible oxidation of methionine residues. As an example, the oxidation of actin by Mical and reduction by the selenoprotein, MsrB1, are highlighted in this Forum (6).
Micals (molecules interacting with CasL) contain an N-terminal flavo-mono-oxygenase domain, by which they stereospecifically oxidize Met in proteins. Mammalian Mical1 and Mical2 oxidize Met residues to Met-R-SO. In turn, MrsB1, initially named SelR or SelX, is a stereospecific methionine-R-sulfoxide reductase. It reduces the R enantiomer of oxidized methionines in proteins. An attack of the sulfur in the S = O group in Met is discussed as the first step of the reaction. The proposed formation of an intermediate selenenic acid—as also postulated for the mechanism of GPxs (8)—is tempting and has, however, so far neither been proven for any GPx nor for MsrB1 by chemical analysis. Reversible post-translational modification of protein methionines such as that of protein thiols is obviously a mechanism to regulate protein activity and function and has to be added to the regulation via phosphorylation, acetylation, and ubiquitination. Prominent examples are the processes that depend on the regulated assembly/disassembly of actin. If methionine residues in actin are oxidized by Micals, actin filaments disassemble. Upon reduction by MsrB1, actin can repolymerize. Thus, the control of actin polymerization is guaranteed by an enzymatic process and not by random oxidation. Furthermore, methionine modification contributes to the change of the image of damaging ROS into important cellular signaling molecules. Macrophages utilize this type of redox control by an increase in MsrB1 expression upon activation. Thus, MsrB1 appears to be part of innate immunity, which, inter alia, might be an explanation for the promotion of anti-inflammatory responses by selenium.
In summary, MsrB1 is a selenoprotein taking part in the regulation of the modification of proteins with essential cellular functions, a function clearly different from the prevention of damage by oxidative stress.
Membrane-Bound Selenoproteins
The article by Liu and Rozovsky (7) highlights the interaction of selenoproteins with membrane-embedded protein complexes, either in signal transduction or in fixing the orientation of membrane proteins.
SelI is a polytopic membrane protein, which catalyzes the last step in the biosynthesis of phosphatidyl ethanolamine, that is, it acts as an ethanolamine phosphotransferase. Based on homology models, it was suggested that the Sec residue of SelI sits at the same site as the active center, the cytoplasmic site. A nearby Cys residue for the formation of a selenylsulfide bond has not been identified so far, yet the possibility remains that Sec of SelI forms an intermolecular diselenide bond with a partner protein.
SelT is only bound to one leaflet of the lipid bilayer (monotopic) and accomplishes a permanent membrane attachment. A hydrophobic domain intercalates and Cys residues appear to bind lipid head groups and determine the orientation. Specific helices form an extensive contact with the membrane, positioning SelT at the membrane interface for optimal interaction with its protein partners. Proposed functions are based on its relationship to SelW. SelW binds to 14-3-3, a family of proteins recognizing phosphorylated substrates and modifying their functions, activity, or cellular localization, as well as their incorporation into protein complexes. The association with SelW prevents 14-3-3 from its binding with partners. SelT may have overlapping or similar functions and thus be again a selenoprotein with functions in the regulation of protein trafficking.
SelN is required for normal muscle development and function. Loss of SelN might be responsible for the white muscle disease observed in sheep under selenium deficiency. In humans, mutations in the gene for SelN lead to muscular dystrophy. The correct localization of SelN in the membrane is not yet known. Structural analyses so far revealed features similar to other members of the Trx superfamily, which support its function in redox signaling. Like SelT, SelN regulates the activity of calcium channels, mainly those of ryanodine receptors (RyRs). The interaction sites with RyR, however, still have to be identified.
SelS, also known as Tanis or VIMP, is a bitopic, that is, a single transmembrane domain, membrane protein. It belongs to the SelS/SelK family. The function of SelS was first described as participating in the transport of multiprotein complexes and recruiting partners in the ER-associated degradation of misfolded proteins into a membrane complex, which has recently been confirmed by a large-scale affinity study (7). In ERAD (ER-associated protein degradation), SelS interacts with p97ATPase needed for providing the energy for the disassembly of protein machineries and pulling misfolded proteins out of the ER lumen. The SelS/p97ATPase complex is shifted to Derlin, the channel through which the misfolded protein is retrotranslocated. Another function is that of a disulfide reductase. Oxidized substrates and SelS form a mixed selenyl sulfide, which in turn is cleaved by a Cys in SelS. The resulting intramolecular selenyl sulfide can be reduced by the Trx/TrxR system. SelS, thus, has two possible roles: providing a scaffold for the formation of protein complexes and disulfide reduction. As discussed for TrxR1, the high nucleophilicity of the Se in Sec might contribute to the former role: creating a Sec-Cys bond to stabilize the complex and guarantee an easy disassembly on demand.
A Specific Function of SelK: Protein Palmitoylation
SelK is another member of the SelS/SelK family (7). Like SelS, it is a bitopic membrane protein. Its partners also belong to the ERAD pathway, the p97ATPase, Derlin 1, and Derlin2. It also stabilizes and modulates the activity of larger protein complexes, possibly via the Se-S bond as discussed for SelS.
In this Forum, Fredericks and Hoffmann (5) describe a novel function of SelK, its participation in the palmitoylation of membrane proteins by the protein, acyl transferase DHHC6 (stands for an aspartic acid-histidine-histidine-cysteine motif within the catalytic domain) (5). DHHC6 palmitoylates and thus stabilizes the inositol-1,4,5-triphosphate receptor (IP3R), a Ca2+ channel in the ER membrane. SelK and DHHC6 contain an SH3 domain by which they interact. Four models of how SelK can facilitate such stabilization have been discussed, from which three require the presence of Sec to form a selenyl disulfide bond between SelK and a Cys in the catalytic domain of DHHC6. (i) Palmitoic acid is more efficiently transferred due to a Sec-Cys bond between DHHC6 and SelK. (ii) Sec becomes intermediately palmitoylated and delivers the palmitoyl residue to the DHHC6 active site, from where it is transferred to the substrate. (iii) Alternatively, the inhibitory oligomerization of DHHC6 is prevented by the selenyl disulfide bond between both proteins. Since palmitoylation conventionally is a two-step reaction (transfer of palmitoic acid to the enzyme and then to the target), the third possibility is favored. If this holds true, Sec is used to recruit and fix proteins to a complex in a fast, stable, and reversible way: a completely novel function for selenium in selenoproteins.
What Makes Selenoproteins So Unique?
This often raised question (1) is most commonly answered by referring to the low pKa of Sec (5.2) compared with that of Cys (8.5). However, this difference in the pK values of the free amino acids appears irrelevant as also pK values of Cys residues within proteins can easily reach five or less, as in redoxins and thiol peroxidases. In addition, molecular manipulations such as Sec to Cys exchange did not provide any solid answer. The typical decline of activity due to replacement of Sec by Cys in selenoenzymes is a week argument since the mutation of Sec in native selenoproteins affects activity more drastically, while native Cys-containing orthologues can be surprisingly active.
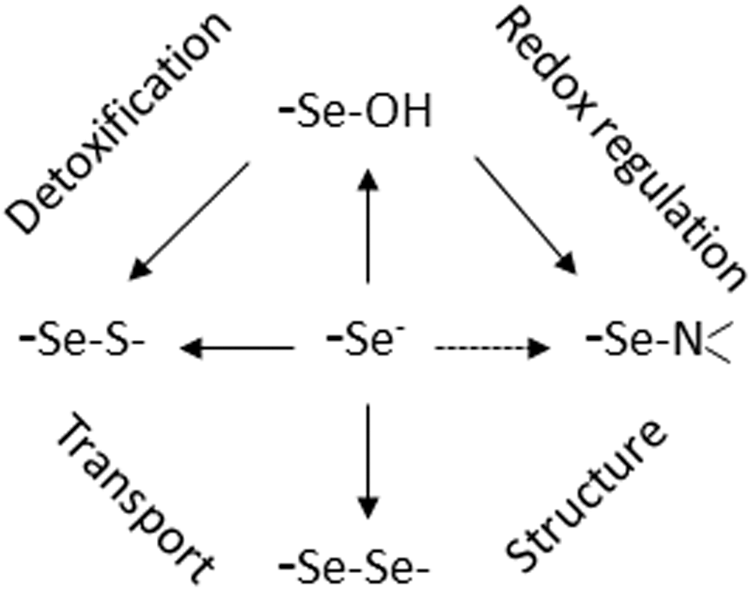
A truly unique feature, nevertheless, of Sec is its high nucleophilicity. This accounts for a much higher reaction rate with electrophiles, which in selenoproteins can reach a 107 times faster rate than that of free cysteine and also makes a thiol (selenol)/disulfide (selenylsulfide) exchange extremely fast (8). Of course, this pronounced nucleophilicity of Sec predisposes selenoproteins for redox reactions, which, as meanwhile amply exemplified, are not always meant to cope with oxidative stress. The biological consequences of these redox reactions reach from detoxification of H2O2 to peroxide sensing and signaling, oxidative protein polymerization, reversible cross-linking of proteins, oxidative protein folding, hormone biosynthesis and degradation, and DNA synthesis and many more can be anticipated. In chemical terms, the known reactions of Sec in selenoproteins are mostly homologous to those we know from sulfur chemistry (Fig. 1) oxidation to selenenic or seleninic acid, diselenide formation, Se-thiolation, S-alkylation, β-cleavage with liberation of H2Se, and the like. However, they generally appear to proceed faster and sometimes the homologous cysteine reactions are not observed at all in a biological context. The deiodination of aromatic rings by the deiodinases may serve as an example. In this respect, the evolving versatility of selenium catalysis reminds of the name-giving goddess, Selene, who loved to surprise with her mysterious abilities (4). The most recent surprise was the discovery of selenium taking an out time from catalytic cycling in GPx (8). In the absence of reducing substrate, the selenium adopts a resting position in the form of a selenenylamide bond and keeps sleeping until it is woken up by a prince's kiss: by its favored thiol substrate.