
Abstract
Aims:
Eukaryotic typical 2-cysteine (Cys) peroxiredoxins (Prxs) are multifunctional proteins subjected to complex regulation and play important roles in oxidative stress resistance, hydrogen peroxide (H2O2) signaling modulation, aging, and cancer, but the information on the biochemical functions and regulation mechanisms of prokaryotic atypical 2-Cys Prxs is largely lacking.
Results:
In this study, we show that at low peroxide concentrations, the atypical 2-Cys Prx in Corynebacterium glutamicum (CgPrx) mainly exists as monomers and displays thioredoxin (Trx)-dependent peroxidase activity. Moderate oxidative stress causes reversible S-mycothiolation of the H2O2-sensing Cys63 residue, which keeps CgPrx exclusively in dimer form with neither peroxidase nor chaperone activity. Then, the increased levels of H2O2 could act as a messenger to oxidize the redox-sensitive regulator hydrogen peroxide-inducible gene activator, leading to activation of expression of the more efficient mycothiol peroxidase and catalase to eliminate excessive peroxide. If oxidative stress is too severe, the H2O2-sensing Cys63 becomes hyperoxidized to sulfonic acid, which irreversibly inactivates the peroxidase activity, and most of CgPrx will be converted to multimeric chaperones for salvage of damaged proteins.
Innovation:
We demonstrate for the first time that atypical 2-Cys CgPrx acts as both a Trx-dependent peroxidase and a molecular chaperone and plays a regulatory role in modulating the peroxide-mediated signaling cascades.
Conclusion:
These results reveal that CgPrx functions as a multifunctional protein crucial for adapting appropriate responses to different levels of oxidative challenge in C. glutamicum. Antioxid. Redox Signal. 26, 1–14.
Introduction
P
In addition to their peroxidase activity, Prxs have been reported to be involved in roles such as protecting DNA against mutation, suppressing tumor formation, promoting longevity, defending pathogens against host immune responses, acting as molecular chaperones to stabilize proteins, and regulating redox signal transduction (18, 24, 33, 34, 40, 50, 51). Under oxidative stress conditions, the peroxidatic Cys-SOH of 2-Cys Prxs is occasionally further oxidized to sulfinic or sulfonic derivatives, which have lost the peroxidase activity. Oxidization also triggers the structure of Prx to transit from low-molecular-weight (LMW) oligomers with peroxidase activity to high-molecular-weight (HMW) complexes with molecular chaperone activity (8, 18). The equilibrium of Prx oligomers is regulated by post-translational modifications and by cues such as redox state, ionic strength, and pH (5, 29, 30).
In this study, we demonstrate that the atypical 2-cysteine (Cys) thioredoxin-dependent peroxidase 2-Cys peroxiredoxin in Corynebacterium glutamicum (CgPrx) also functions as a molecular chaperone. The switch between these two functions is regulated by oxidative stress-triggered S-mycothiolation, which is concomitant with significant changes in its protein structure. We also demonstrate that CgPrx plays a regulatory role in modulation of peroxide-mediated signaling cascades by controlling the activation of hydrogen peroxide (H2O2)-inducible gene activator and expression of catalase (katA) and mycothiol peroxidase (mpx). Our data establish that CgPrx, by acting as a delicate H2O2 sensor and the primary scavenger of H2O2, is central for making appropriate responses to different levels of H2O2 faced by C. glutamicum.
As a reversible post-translational thiol modification, glutathionylation has been reported to protect the active site Cys residues of certain Prxs from irreversible overoxidation to sulfonic acids, regulate the quaternary structure of some typical 2-Cys Prxs, or to provide reducing equivalents to support peroxidase activity (6, 25, 32, 36). Moreover, inactivation of eukaryotic 2-Cys Prx by hyperoxidation or phosphorylation has been proposed to promote the localized accumulation of H2O2 and thus regulate the activity of other proteins (10, 50, 51). However, most of the current knowledge on Prxs was obtained in eukaryotic cells; information about bacterial Prxs was largely lacking because of their robust response to oxidative inactivation and appeared to be required only to degrade peroxides.
Corynebacterium glutamicum, one of the most important industrial bacteria widely used for production of amino acids and nucleotides, is also an excellent laboratory model for systems biology study and for the investigation of its pathogenic relatives such as C. diphtheriae, C. jeikeium, and Mycobacterium tuberculosis (44). C. glutamicum exhibits remarkable resistance to H2O2, a feature that has drawn considerable attention for mechanistic studies (27). C. glutamicum produces mycothiol (MSH; AcCys-GlcN-Ins) as the main nonenzymatic antioxidant. MSH is functionally equivalent to glutathione (γ-
Recently, Chi et al. found that S-mycothiolation of the atypical 2-Cys peroxidase in C. glutamicum (CgPrx) inhibits the peroxidase activity and that such inhibition can be reversed by reduction mediated by the Mrx1/MSH/mycothiol disulfide reductase electron pathway (7). While the dual functions and switching mechanisms between different functions and structures of typical 2-Cys Prxs have been well documented, the molecular chaperone activity has not yet been reported for any atypical 2-Cys Prxs. This study contributes to a deeper characterization of the atypical 2-Cys Prx, CgPrx, in C. glutamicum. A model in which CgPrx functions as a multifunctional protein crucial for adapting appropriate responses to different levels of oxidative challenge in C. glutamicum was proposed.
Results
CgPrx is required for resistance to low oxidative stress
Sequence analyses revealed that CgPrx belongs to the atypical 2-Cys Prx group, whose members have been shown to function as Trx-dependent monomeric peroxidases (20) (Supplementary Fig. S1; Supplementary Data are available online at
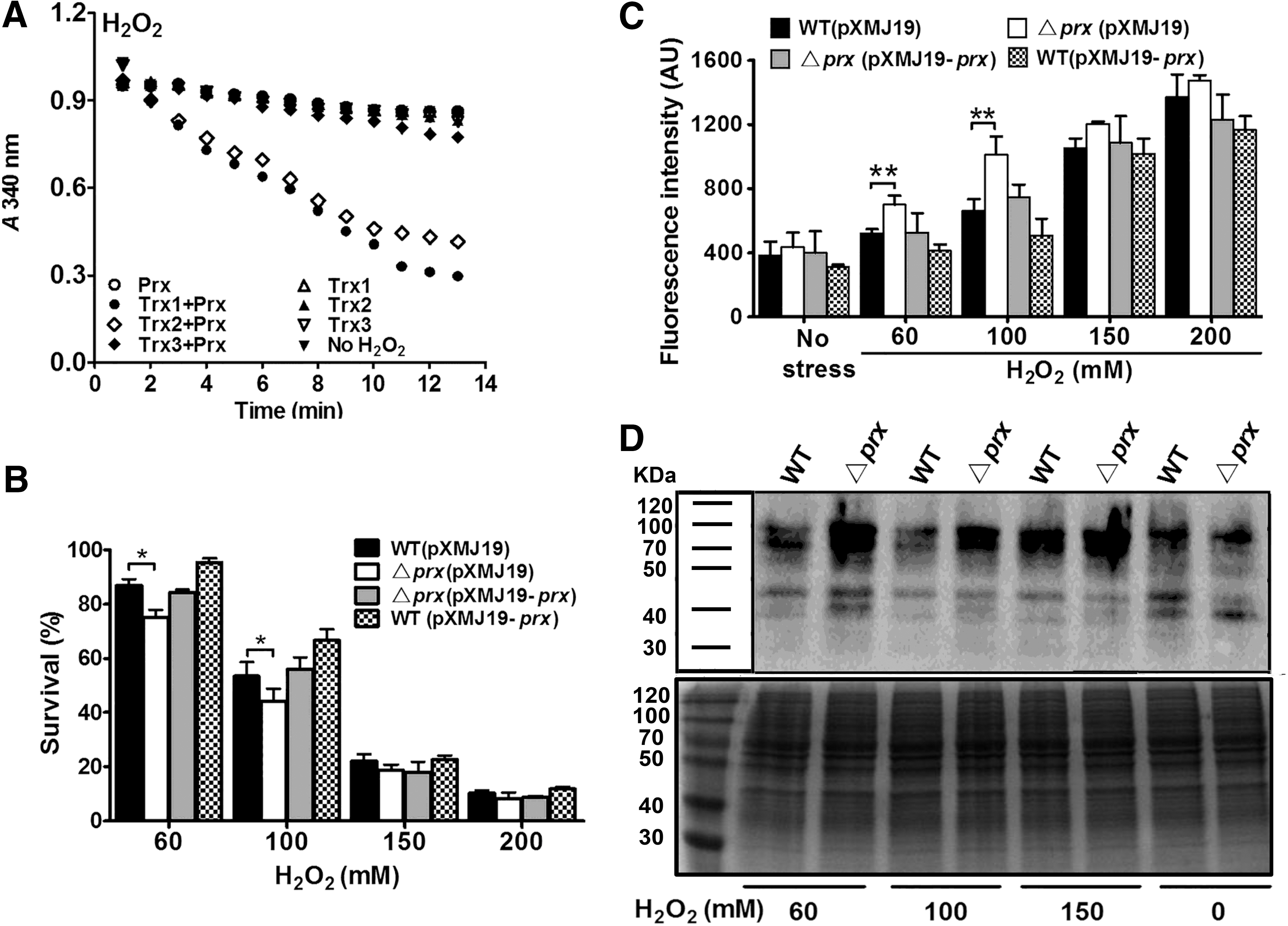
To investigate the physiological function of CgPrx, we tested the effects of deleting this gene on the sensitivity of C. glutamicum to H2O2. Compared with the wild type, the Δprx mutant was more sensitive to low concentrations of H2O2 (60 and 100 mM), and the sensitive phenotype can be almost completely complemented by expressing the prx gene (Fig. 1B). Interestingly, the mutant exhibited similar sensitivity to high concentrations of H2O2 (150 and 200 mM) to that of the wild type (Fig. 1B). In agreement with this observation, overexpression of CgPrx increased the resistance of the wild type to low, but not high, concentrations of H2O2 (Fig. 1B).
Since Prx is known to eliminate ROS under oxidative stress in various organisms (14, 21), we next examined the ability of CgPrx in ROS removal by fluorometrically measuring intracellular ROS levels with the membrane permeate fluorogenic probe, 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) (48). Comparing with the wild type, the Δprx mutant contained significantly higher ROS after being exposed to low concentrations of H2O2 for 30 min. This higher ROS levels in the mutant can be reversed by expressing the prx gene (Fig. 1C). The higher level of ROS in the mutant was not detected when the cells were exposed to higher concentrations of H2O2 (Fig. 1C). To test whether CgPrx is capable of reducing oxidative damage to proteins under low H2O2 concentrations, we performed carbonyl assays with proteins isolated from the Δprx mutant and wild-type cells that had been treated with H2O2. A 30-min exposure to <100 mM H2O2 caused more carbonylation in proteins from the mutant (Fig. 1D). Such difference was not observed when cells were treated with 150 mM H2O2. Taken together, these results demonstrate that CgPrx plays an important role in protecting C. glutamicum cells against oxidative damage caused by low levels of H2O2.
Cys63 and Cys97 form an intramolecular disulfide bond during the reduction of H2O2
CgPrx contains three Cys residues in which Cys63 and Cys97 likely are the peroxidatic and the resolving Cyss (Supplementary Fig. S1), respectively, essential for the activity of atypical 2-Cys Prxs (51). In reaction, the peroxidatic Cys residue first is oxidized by peroxide to a Cys sulfenic acid (CP-SOH) intermediate (12); this peroxidatic Cys-SOH is then attacked by the resolving Cys of Prx to form a redox-active disulfide. To stabilize and trap the Cys-SOH, the C-terminal Cys of the active site disulfide pair must be removed (12). To test this, we produced CgPrx variants, CgPrx:C84SC97S, CgPrx:C63SC97S, and CgPrx:C63SC84S, and treated the proteins with 4-chloro-7-nitrobenzofurazan (NBD-Cl) under anaerobic condition with or without prior exposure to H2O2. H2O2-treated CgPrx:C84SC97S labeled with NBD gave a maximal absorbance at 347 nm characteristic of the NBD-modified product [R-S(O)-NBD], indicating the detection and trapping of approximately stoichiometric amounts of SOH by Cys63, the only Cys in this variant (Fig. 2A). In contrast, CgPrx:C84SC97S treated with NBD-Cl, but not H2O2, gave a maximal absorbance at 420 nm, typical for the thiol adducts with NBD-Cl (Cys-S-NBD) (42). In contrast, the absorbance of the CgPrx:C63SC97S and CgPrx:C63SC84S variants remained unchanged upon incubation with NBD-Cl either before or after exposure to H2O2. These results further support that Cys63 is the peroxidatic Cys residue in CgPrx, a conclusion that was strengthened by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-TOF MS/MS) analysis, which detected a peak of 2135.7 Da in the H2O2-treated CgPrx:C84SC97S (labeled [B] in inset panel), 16 Da higher compared with the untreated protein resulting from the addition of an oxygen atom and formation of Cys sulfenic acid (Cys-SOH) on the Cys63 residue (Fig. 2B and Supplementary Fig. S4A).

To investigate whether Cys97 acts as the resolving Cys residue of CgPrx, we analyzed the formation of disulfide bonds in single-Cys variants of CgPrx (CgPrx:C63S, CgPrx:C84S, and CgPrx:C97S) in response to H2O2 using the 4-acetamido-4′-maleimidyldystilbene-2, 2′-disulfonic acid (AMS) covalent modification assay. AMS covalently modifies free thiol groups, leading to slower electrophoretic mobility proportional to the number of free thiol groups in the protein (11). AMS-treated oxidized CgPrx:C84S migrated more rapidly compared with similarly treated reduced CgPrx:C84S, but similar to that of oxidized untreated CgPrx:C84S (Fig. 2C), indicating that CgPrx:C84S was fully oxidized by H2O2 to form a disulfide bond between Cys63 and Cys97. However, CgPrx:C97S, treated with H2O2 and modified with AMS, migrated faster than its reduced form (AMS modified) and slower than that of the oxidized (unmodified) form (Fig. 2C), indicating that despite modification, the protein was not fully oxidized. Oxidized and reduced CgPrx:C63S were modified by AMS and have the same electrophoretic mobility, indicating that these proteins were not oxidized by H2O2 and Cys84/Cys97 still existed in the thiol state. These indicate that CgPrx:C84S is fully oxidized by H2O2 to form a disulfide bond between Cys63 and Cys97. This conclusion was confirmed by MS analysis. The protein bands from H2O2-treated CgPrx:C84S (labeled [D] in Fig. 2C) have a mass of 3945.9 Da, which corresponds to the 50-to-69 peptide (calculated and observed mass, 2119.5 Da) and the 96-to-112 peptide (calculated and observed mass, 1828.2 Da) of the CgPrx:C84S variant connected by a disulfide bridge that does not appear in non-H2O2-treated CgPrx:C84S (Fig. 2D and Supplementary Fig. S4B). These results demonstrated that CgPrx is capable of forming a disulfide bond between the peroxidatic Cys63 and the resolving Cys97 during H2O2 decomposition.
CgPrx switches from a peroxidase to a molecular chaperone upon sulfonation of Cys63 by high concentrations of H2O2
Several typical 2-Cys Prxs have been reported to have dual physiological functions as peroxidase and molecular chaperone (1, 2, 8, 18). To investigate whether the atypical 2-Cys Prx CgPrx possesses molecular chaperone activity, we examined its ability to inhibit thermal aggregation of malate dehydrogenase (MDH) at high temperature. Incubation of more than 7.5 mM H2O2-treated CgPrx and MDH at a 1:1 subunit molar ratio prevented the latter from aggregation (Fig. 3A). Similarly, CgPrx treated with higher than 7.5 mM H2O2 effectively protected insulin from dithiothreitol (DTT)-induced protein precipitation (Fig. 3B), further suggesting that CgPrx acts as an effective molecular chaperone. Importantly, CgPrx not treated with H2O2 displayed high peroxidase activity, but not chaperone activity. The peroxidase and molecular chaperone activities of CgPrx were regulated by H2O2 (Fig. 3C). CgPrx exhibits high peroxidase activity in the presence of low concentrations of H2O2 (0.1 mM) for 30 min; its molecular chaperone activity is negligible under this condition. As the concentration of H2O2 increases, its peroxidase activity declines, accompanied by increased molecular chaperone activity. The chaperone activity of CgPrx peaks in the presence of 25 mM H2O2 and is about 2–2.5-fold weaker compared with the well-known molecular chaperone, heat shock protein 70 (Fig. 3C).
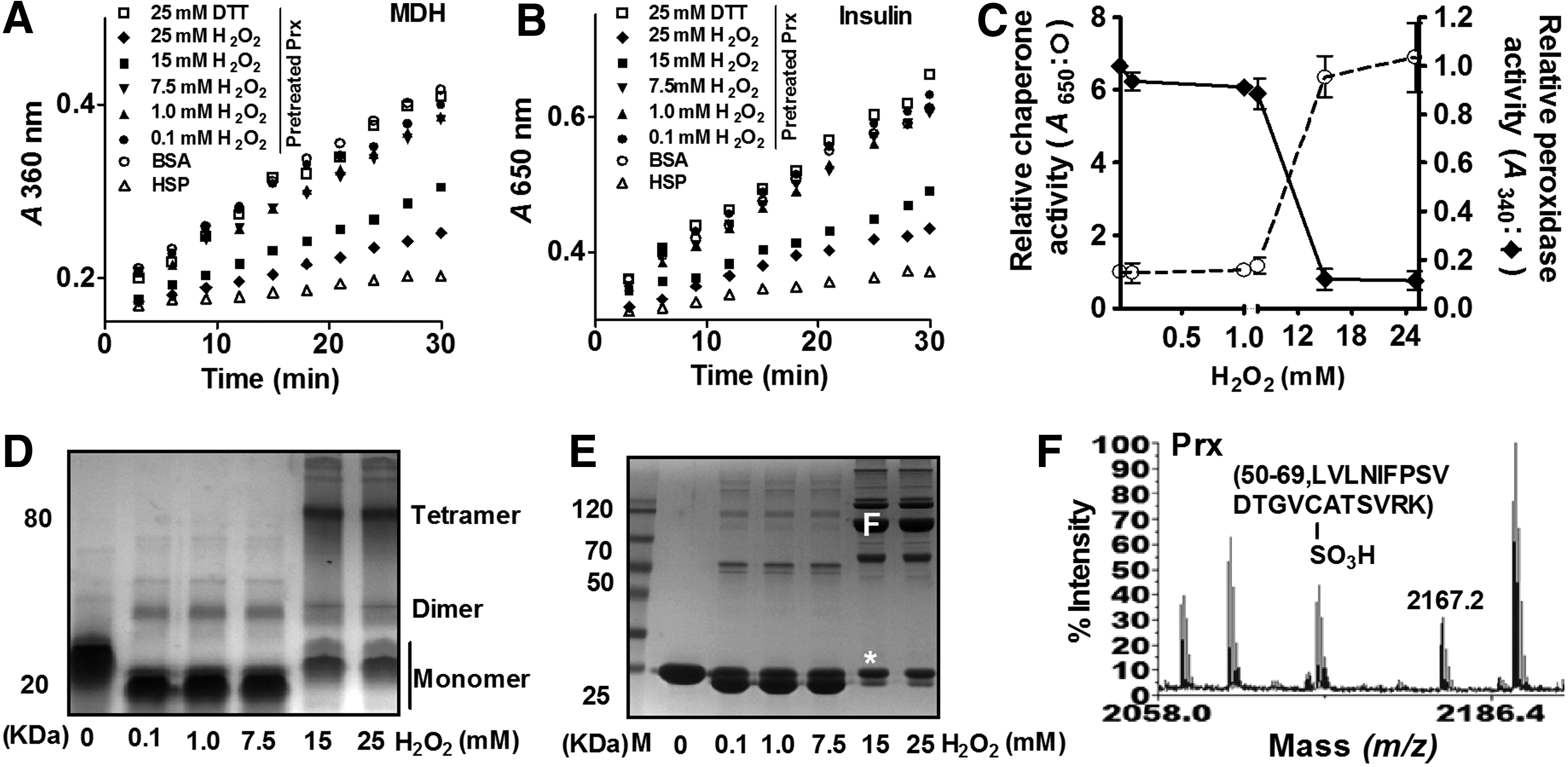
Interestingly, H2O2-induced switch in the biochemical activity of CgPrx occurs concomitant with significant changes in its protein structure. H2O2-treated CgPrx migrated as multiple bands in both native polyacrylamide gel electrophoresis (PAGE) and nonreducing sodium dodecyl sulfate (SDS)-PAGE gels (Fig. 3D, E). The abundance of protein bands corresponding to oligomers increased as the concentration of H2O2 increases (Fig. 3D, E). In contrast, CgPrx not treated with H2O2 existed as a monomer (Fig. 3D, E).
CgPrx appeared to exist mainly as intramolecular disulfide bond-containing monomers when treated with low concentrations of H2O2 (0.1, 1, and 7.5 mM) (Fig. 3D, E). High concentrations of H2O2 (15 and 25 mM) induced the formation of multimers, including dimers, tetramers, and hexamers (Fig. 3D, E). A novel peak of 2167.2 Da peptide that was 48 Da higher than that of the Cys-63-containing 56–99 peptide (LVLNIFPSVDTGVCATSVRK) in reduced CgPrx was observed in tetrameric and monomeric CgPrx (labeled [F] and [*] in Fig. 3E), suggesting that the CgPrx was sulfonated on the Cys63 residue when treated with high concentrations of H2O2 (Fig. 3F and Supplementary Fig. S4C, D). Thus, CgPrx functions as a peroxidase in the form of monomers in low concentrations of H2O2 and switches to oligomers with chaperone activity upon being sulfonated on the Cys63 residue when treated with high concentrations of H2O2.
Oxidative stress-induced structural changes of CgPrx in vivo
To examine whether the structure of CgPrx is regulated by oxidative stress in vivo, we treated cells of the Δprx mutant overexpressing His6-CgPrx with various concentrations of H2O2 and probed the forms of His6-CgPrx by immunoblotting after nonreducing SDS-PAGE separation. Our results indicate that in the absence of oxidative stress, CgPrx existed as reduced monomers and became intramolecular disulfide bond-containing monomers when exposed to oxidative stress conditions (<120 mM H2O2) (Fig. 4A, B). One hundred twenty micromolar H2O2 induced most proteins into its dimer form (labeled [C] in Fig. 4A). Interestingly, an extra mass of 2605.2 Da was identified in this dimer, which was 484 Da higher than the Cys-63-containing (56–99) peptide of reduced CgPrx (Fig. 4C and Supplementary Fig. S4D). This observation is consistent with the result from the addition of MSH, indicating that Cys63 was mycothiolated under certain oxidative stress. This observation was confirmed by the reversible biotin switch assay. His6-CgPrx overexpressed in the Δprx mutant pretreated with 120 mM H2O2 exhibited a strong band of mycothiolation. However, no mycothiolation signal was observed in the non-H2O2-treated Δprx(pXMJ19-His6-prx) strain. Similarly, no mycothiolation signal was observed for His6-CgPrx overexpressed in the MSH null mutant, ΔprxΔmshC, regardless of 120 mM H2O2 treatment (Fig. 4E).
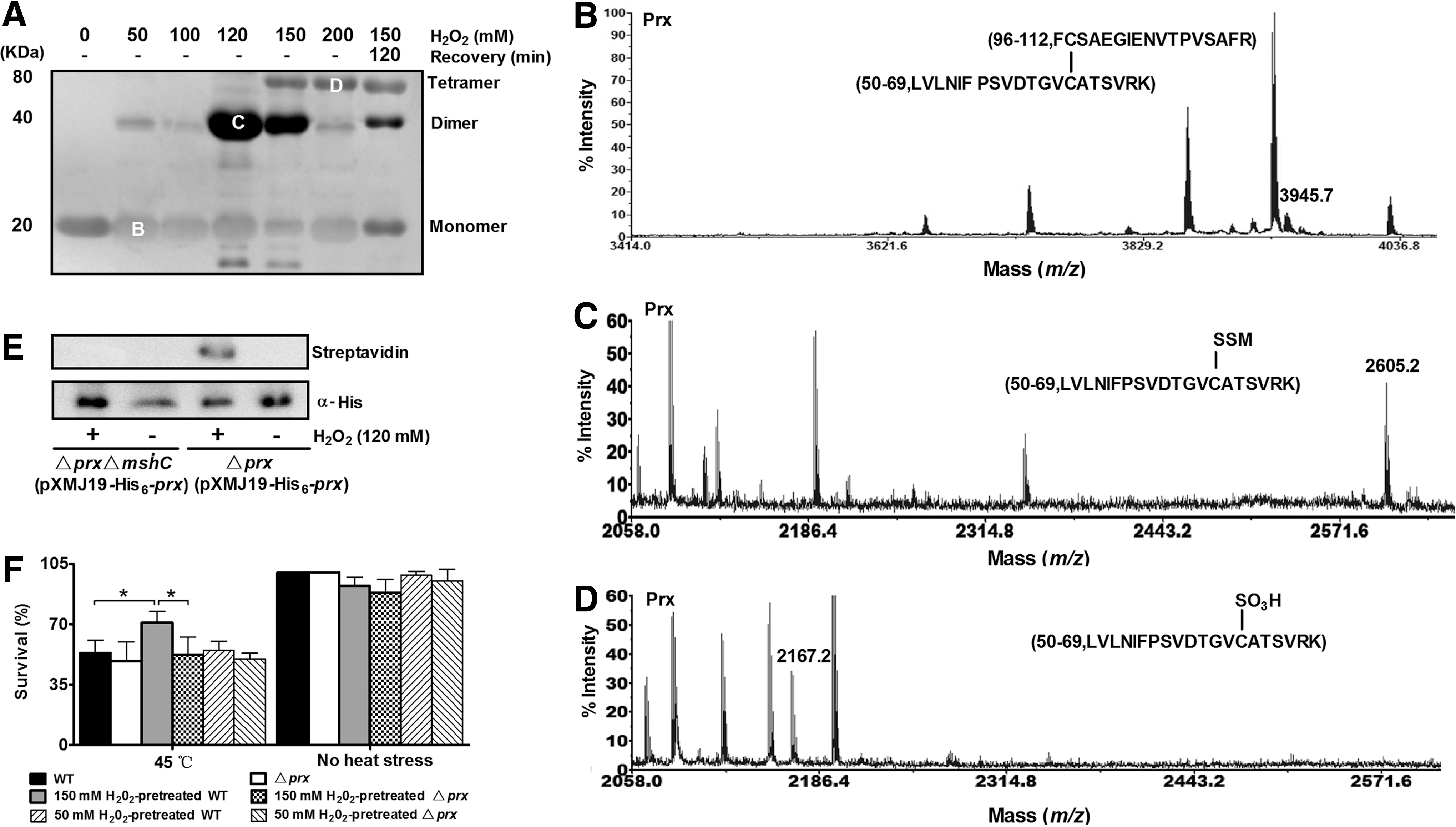
Treatment with 150 mM H2O2 resulted in the formation of sulfonated CgPrx tetramers as well as mycothiolated dimers. However, increasing H2O2 to 200 mM caused more production of sulfonated CgPrx tetramers (Fig. 4D). Withdrawing H2O2 for 120 min led to reduction of mycothiolated CgPrx (CgPrx-SSM) dimers, but not sulfonated CgPrx tetramers, indicating that CgPrx tetramers were irreversibly overoxidized to form sulfonic acid (Fig. 4A). Pretreatment of wild-type C. glutamicum with 150 mM H2O2 endowed significant resistance to subsequent heat stress treatment (45°C for 1h); such resistance did not occur in the Δprx mutant (Fig. 4F). In contrast, treatment with 50 mM H2O2 had no detectable effect in improving the heat stress resistance to either strain (Fig. 4F). Thus, sulfonated CgPrx tetramers induced by high levels of H2O2 possess chaperone activity.
Similar structure changes were observed for endogenous CgPrx treated under the same conditions visualized by a specific anti-CgPrx rabbit polyclonal antibody (Supplementary Fig. S5). These indicate that CgPrx confers different activities in different structural forms when the bacterial cells meet different levels of oxidative challenges.
Mycothiolation of Cys63 regulates the oligomerization and catalytic activity of CgPrx under moderate H2O2 stress
To further investigate the function of mycothiolated CgPrx under oxidative stress, we first analyzed the oligomeric status of His6-CgPrx expressed in Δprx(pXMJ19-His6-prx) and ΔprxΔmshC(pXMJ19-His6-prx) under 120 mM H2O2 treatment. His6-CgPrx from strain Δprx(pXMJ19-His6-prx) exposed to 120 mM H2O2 showed a major band at about 40 kDa position (dimers), whereas His6-CgPrx from similarly treated ΔprxΔmshC(pXMJ19-His6-prx) hardly contained that 40 kDa band; instead, most of them existed as tetramers (Fig. 5A, lane 4). Consistently, dimer formation was also detected by treating purified CgPrx with 20 mM H2O2 and 0.5 mM MSH, but not with H2O2 alone (Fig. 5B). Instead, a band about 80 kDa corresponding to CgPrx tetramers was detected (Fig. 5B). A novel peak of 2605.9 Da that was 484 Da higher than that of the Cys-63-containing 56–99 peptide (2119.5 Da) in reduced CgPrx was observed in dimeric CgPrx (labeled [D] in Fig. 5A), indicating the addition of MSH on the Cys63 residue (Fig. 5D and Supplementary Fig. S4D).
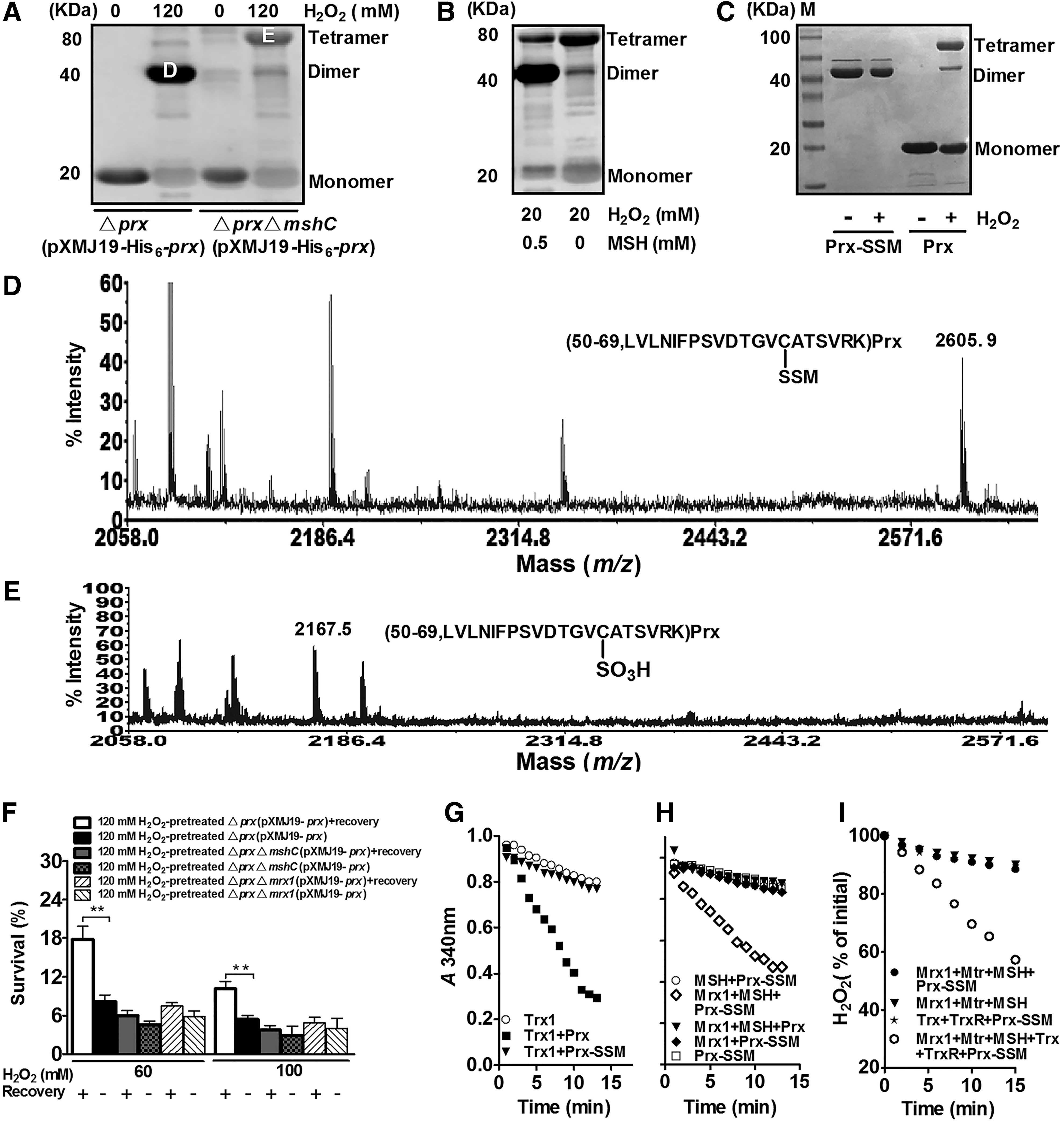
In contrast, mycothiolation of Cys63 was not observed in CgPrx tetramers. Instead, the Cys63 residue was sulfonated in CgPrx tetramers, with a mass of 2167.5 Da (labeled [E] in Fig. 5A) that was 48 Da higher than Cys63-containing 56–99 peptide of the reduced CgPrx (Fig. 5E). The above results suggest that mycothiolation of CgPrx prevents the formation of overoxidized tetramers under moderate oxidative stress. Consistent with this conclusion, while 20 mM H2O2-treated CgPrx appeared as both dimers and tetramers, similarly treated CgPrx-SSM existed only as dimers (Fig. 5C).
Protein S-thiolation is a reversible post-translational thiol modification that protects active site Cys residues of key enzymes against irreversible overoxidation and restores their functions under normal conditions (9). To investigate the role of S-mycothiolation in protecting the peroxidase activity of CgPrx, we determined the effects of recovery on the survival rates of low concentrations (60 and 100 mM) of H2O2-treated strains, Δprx(pXMJ19-prx) and ΔprxΔmshC(pXMJ19-prx). A 120-min recovery allowed the MSH-containing Δprx(pXMJ19-prx) strain pretreated with H2O2 to survive better. On the contrary, similar recovery time had very marginal effects on restoring the sensitivity phenotype of the MSH lacking ΔmshCΔprx(pXMJ19-prx) treated with H2O2 (Fig. 5F). Moreover, withdrawing H2O2 for 120 min led to a decrease of mycothiolated CgPrx dimers formed in the bacterial cells treated with 150 mM H2O2, which was accompanied by an increase in CgPrx monomers (Fig. 4A). These results indicate that S-mycothiolation not only regulates the oligomerization status of CgPrx but also reversibly protects its peroxidase activity under moderate H2O2 stress.
Mrx1-catalyzed demycothiolation reactivates S-mycothiolated CgPrx
S-mycothiolation inhibits the peroxidase activity of CgPrx in a way that can be restored by Mrx1-catalyzed demycothiolation. In agreement with this notion, no peroxidase activity was detected in S-mycothiolated CgPrx (CgPrx-SSM) (Fig. 5G). To reactivate mycothiolated CgPrx, we reconstituted the CgMrx1/CgMtr/MSH electron transfer pathway with CgPrx-SSM as a substrate. Only samples containing 50 μM CgPrx-SSM showed consumption of NADPH (Fig. 5H). No reduction of CgPrx-SSM was observed in the absence of CgMrx1, indicating that high levels of MSH (500 μM) alone were not sufficient to demycothiolate CgPrx-SSM in the absence of CgMrx1. As expected, demycothiolated CgPrx was able to restore the peroxidase activity in the CgTrx/CgTrxR and CgMrx1/CgMtr/MSH-coupled enzyme assay system (Fig. 5I). These results indicate that S-mycothiolation of Cys63 inhibits the peroxidase, and such inhibition can be reversed by demycothiolation with the CgMrx1/CgMtr/MSH-reducing system.
Consistent with this conclusion, a 120-min recovery significantly increased the survival rates of H2O2-pretreated Δprx(pXMJ19-prx) to low concentrations of H2O2 (60 and 100 mM) and had very marginal effects on restoring the sensitivity phenotype of H2O2-pretreated ΔprxΔmrx1(pXMJ19-prx) to the same treatment (Fig. 5F). In conclusion, S-mycothiolation reversibly protects CgPrx from overoxidation under moderate oxidative stress.
CgPrx modulates the activation of hydrogen peroxide-inducible gene activator and the expression of katA and mpx
In addition to Prx, KatA and MPx are two other major H2O2 scavengers in C. glutamicum. To distinguish the potentially different roles of Prx, KatA, and MPx in removing H2O2 by C. glutamicum, we investigated the survival rate of Δprx, Δmpx, and ΔkatA challenged with different H2O2 concentrations. Deletion of prx, mpx, or katA did not affect bacterial growth under normal condition without H2O2 stress (Supplementary Fig. S6A). Notably, the Δprx mutant was slightly, but substantially, more sensitive to low concentrations of H2O2 (45 mM) than the wild type, whereas the ΔkatA and Δmpx mutants showed wild-type-level resistance under the same condition. Conversely, the ΔkatA and Δmpx mutants were hypersensitive to high concentrations of H2O2 (100–200 mM) compared with the wild type, whereas the Δprx mutant showed a wild-type-level resistance under the same conditions (Fig. 6A). Consistent with the survival results, the ΔmpxΔkatA double mutant containing only Prx scavenged 5 mM H2O2 almost as effectively as the wild type; the ΔmpxΔprxΔkatA triple mutant completely failed to scavenge 120 mM H2O2. Whereas strains, ΔprxΔkatA and ΔprxΔmpx, were less effective as ΔmpxΔkatA in demolishing 5 mM H2O2, both were clearly more effective in abolishing 120 mM H2O2 than ΔmpxΔkatA (Fig. 6B). These results indicate that CgPrx, KatA, and MPx play distinct roles: CgPrx is more effective in scavenging very low concentrations of H2O2, while KatA and MPx are the more effective enzymes at higher concentrations. This conclusion was supported by the fact that the survival rates of the ΔmpxΔkatA mutant were significantly lower compared with the wild type after exposure to 120 mM H2O2 for 10 min; this mutant survived at rates similar to that of the wild type after exposure to 5 mM H2O2 for 30 min. In contrast, the ΔprxΔkatA and ΔprxΔmpx mutants lacking Prx survived at rates similar to that of the wild type after exposure to 120 mM H2O2 for 10 min; such rates were substantially lower compared with the wild type after exposure to 5 mM H2O2 for 30 min (Fig. 6C). Note that the growth of ΔprxΔkatA, ΔprxΔmpx, and ΔkatAΔmpx was nearly identical to the wild type under normal condition without H2O2 stress (Supplementary Fig. S6B).
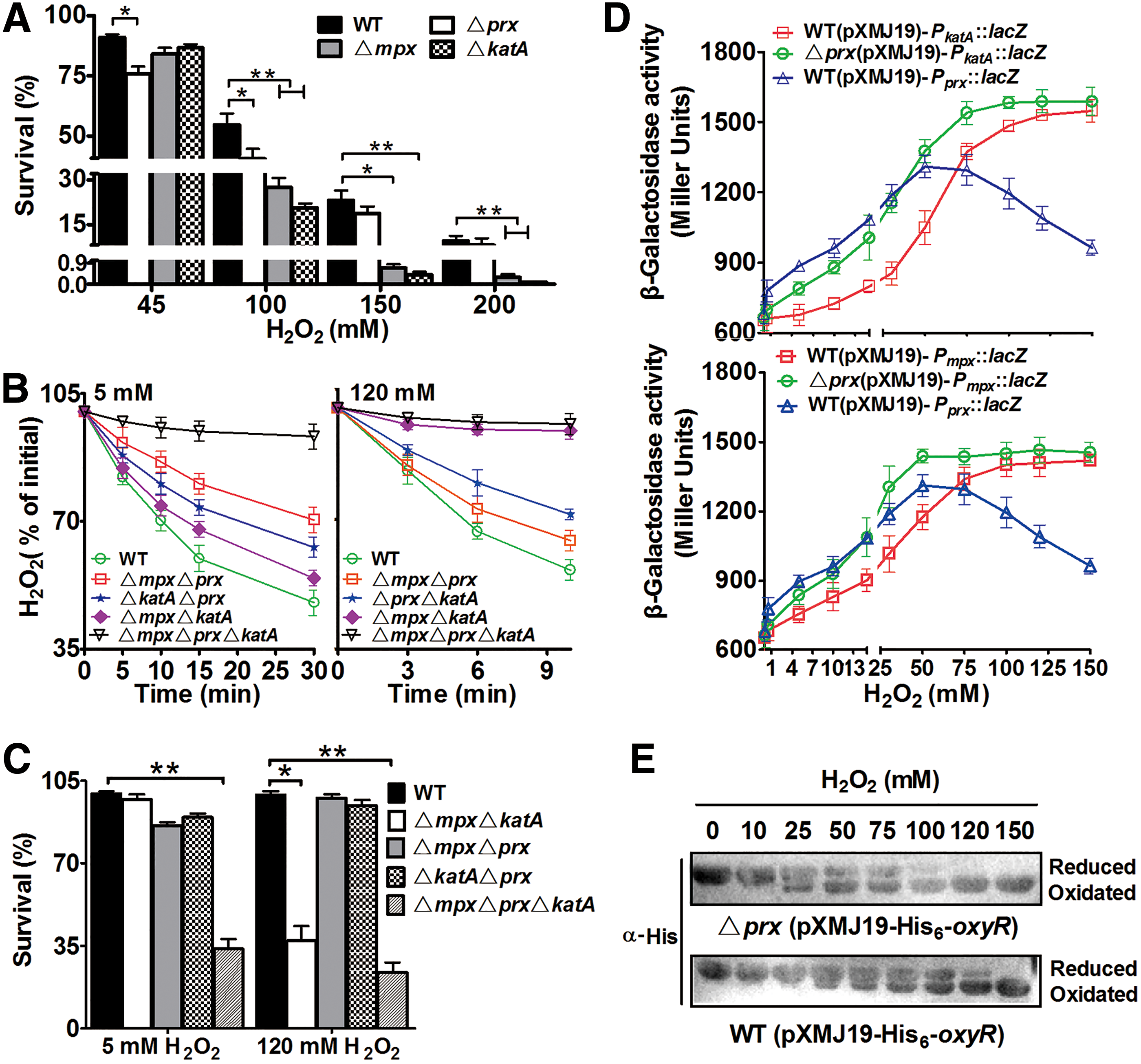
Consistent with the above conclusions, the expression level of katA and mpx was lower than that of prx under low concentrations of H2O2, but became significantly higher than that of prx in the presence of high concentrations of H2O2 (Fig. 6D). Strikingly, the expression level of katA and mpx increased to that of prx level at low concentrations of H2O2 in a strain without the prx gene (Fig. 6D), suggesting that Prx inhibits the expression of katA and mpx under low H2O2 concentration, possibly by keeping an extremely low intracellular H2O2 concentration through its peroxidase activity. An increase in intracellular H2O2 levels is known to signal the activation of the primary redox-sensitive regulator hydrogen peroxide-inducible gene activator (OxyR) by oxidation, leading to the induction of the expression of the more powerful KatA (41) and also possibly MPx. Indeed, at least 50 mM H2O2 was required for OxyR activation in wild-type bacteria, but only 25 mM was required for such activation in the Δprx mutant, which was sufficient to induce the expression of katA and possibly mpx (Fig. 6E).
Discussion
The results herein revealed that CgPrx, an atypical bacterial 2-Cys Prx, by acting as a delicate H2O2 sensor and the primary scavenger of H2O2 in C. glutamicum, indeed plays central roles for making appropriate responses to different levels of H2O2 (Fig. 7). (i) Under low peroxide concentrations, the H2O2-sensing peroxidatic Cys63 forms an intramolecular disulfide with the resolving Cys97, which allows monomeric CgPrx to reduce peroxide as a Trx-dependent peroxidase. This activity prevents KatA and MPx from unnecessary expression. (ii) Moderate peroxide challenge induces the reversible S-mycothiolation of the H2O2-sensing Cys63, converting CgPrx into an inactive dimer form. Thus, increased levels of H2O2 oxidize redox-sensitive regulators such as OxyR to induce the expression of the more efficient KatA and MPx to eliminate the excess peroxide. (iii) Under hyperoxidative stress condition, the H2O2-sensing Cys63 will be hyperoxidized to Cys-sulfonate, leading to irreversible inactivation of peroxidase activity. Most CgPrx molecules will transit to tetrameric and higher molecular weight chaperones for the salvage of damaged proteins.

The typical 2-Cys Prx is able to assume multiple oligomeric structures in response to cellular redox states and the transition between these structures dynamically regulates the switch between the peroxidase and chaperone activities (5). For example, the 2-Cys Prx from yeast transits from LMW species to oligomeric and HMW complexes upon being challenged by oxidative stress or heat shock, whose activity is switched from peroxidase to chaperone (18). Although atypical 2-Cys Prxs appear to exist in the lower polymerization degree (dimers, tetramers, and hexamers) compared with typical 2-Cys Prxs (decamers and HMW species), increasing evidence indicates that they also undergo dynamic conformation transitions with functional implications (4, 5). Our finding that high concentrations of H2O2 induce CgPrx into dimers, tetramers, and even HMW species is in line with these observations. While most of typical 2-Cys Prxs and even some 1-Cys Prxs have been found to function as both peroxidases and molecular chaperones (1, 2, 8, 18, 22), this is the first report on the dual activity of an atypical 2-Cys Prx.
Besides being hyperoxidized to induce the peoxidase-to-chaperone functional switch under high H2O2 concentrations, the Cys63 sensor was modified by S-mycothiolation under moderate concentrations of H2O2 treatment (Fig. 4). S-mycothiolation clearly protects CgPrx from being damaged under oxidative stress, which is in line with the fact that a reversible thiol modification often protects active Cys of key enzymes against irreversible overoxidation to sulfonic acids. In addition, S-mycothiolation plays a crucial role in regulating the oligomerization status of CgPrx in a way different from S-glutathionylation. While S-glutathionylation induced the dissociation of Prx oligomers (6, 32, 36), S-mycothiolation maintains CgPrx exclusively in its dimeric form, defective in both peroxidase and chaperone activity, and prevents the CgPrx dimer to form tetramer and HMW complexes (Fig. 5). Thus, S-mycothiolation of CgPrx not only maintains CgPrx in its dimeric form, defective in both peroxidase and chaperone activity, but also protects CgPrx from irreversible overoxidation.
In eukaryotes, 2-Cys Prxs containing the oxidative susceptible GGLG and YF motifs utilize a unique regulatory mechanism to modulate the peroxide-mediated signaling cascades. According to the floodgate hypothesis, sensitive 2-Cys Prxs act as a peroxide floodgate to keep peroxides away from susceptible molecules until it is opened by inactivation via overoxidation, which allows a rapid increase in H2O2 levels and thus permits its signaling activity (51). In contrast, because of the lack of the GGLG and YF motifs and their robust response to oxidative inactivation, bacterial 2-Cys Prxs have never been attributed to signaling events. Strikingly, our results clearly demonstrate that CgPrx uses a more complex floodgate mechanism to modulate the peroxide signal in C. glutamicum. Inactivation of CgPrx by S-mycothiolation induced the expression of katA and mpx via activating the OxyR regulator. This mechanism provides a rapid and reversible way to open the floodgate without the need to produce the irreversibly inactivated protein. The CgPrx floodgate can also be completely opened by irreversible overoxidation, which occurs possibly under severe oxidative conditions that threaten the viability of bacterial cells. Thus, it appears that bacterial Prx has evolved as a more complex mechanism than ever thought before for adapting appropriate responses to different levels of oxidative challenge.
Materials and Methods
Bacterial strains and culture conditions
Bacterial strains and plasmids used in this study are listed in Supplementary Table S1. C. glutamicum and Escherichia coli strains were cultured in mineral salts medium (MM) or Luria-Bertani (LB) medium as previously reported (43). In-frame deletions were generated by means of the method described (45). Sensitivity assays for peroxides were performed as described (45).
Cloning, expression, and purification of recombinant proteins
The genes encoding C. glutamicum Trx2 (NCgl2874), Trx3 (NCgl0289), CgPrx (NCgl1041), and OxyR (NCgl1850), were amplified by polymerase chain reaction (PCR). These DNA fragments were digested and subcloned into similar digested vectors, obtaining pET28a-trx2, pET28a-trx3, pET28a-prx, pGEX6p-1-prx, pXMJ19-prx, pXMJ19-His6-prx, and pXMJ19-His6-oxyR, respectively. Site-directed mutagenesis was constructed as described (53). The plasmids, pK18mobsacB-Δprx and pK18mobsacB-ΔkatA, used to construct deletion mutants were made by overlap PCR (53). The lacZ fusion reporter vectors, pK18mobsacB-PkatA::lacZ and pK18mobsacB-Pprx::lacZ, were made by fusion of the katA and prx promoter to the lacZY reporter gene via overlap PCR (53). For overexpression or complementation in C. glutamicum strains, corresponding pXMJ19 derivatives were transformed into C. glutamicum by electroporation. The transformants were selected on LB plates supplemented with nalidixic acid and chloramphenicol (45). Expression in C. glutamicum was induced by addition of 0.5 mM isopropyl β-
Analysis of sulfenic acid and disulfide bond formation
The formation of Cys-SOH as a reaction intermediate was examined by the assays of 50 mM DTT-treated or 10 mM H2O2-treated CgPrx:C84SC97S, CgPrx:C63SC84S, and CgPrx:C63SC97S proteins labeled with NBD-Cl (12, 42). The formation of disulfide bond was measured with AMS (Molecular Probes, Eugene, OR) as described (11).
Enzyme assays
Peroxidase activity of CgPrx was determined by monitoring the rate of NADPH oxidation (3). By using the Trx-based regeneration system, the reaction mixture comprised 50 mM Tris-HCl buffer (pH 8.0), 2 mM ethylenediaminetetraacetic acid (EDTA), 250 μM NADPH, 5 μM CgTrxR, 1 μM CgPrx, and 40 μM CgTrx. In alternative assays by using the CgMrx1/CgMtr/MSH-based system, CgTrxR was replaced by 5 μM CgMtr and 500 μM MSH and CgTrx was replaced by 40 μM CgMrx1. The reaction was started by addition of 100 μM peroxide, following a 5-min preincubation, and NADPH oxidation was monitored at 340 nm. Molecular chaperone activities were determined by assessing the ability of recombinant CgPrx to inhibit the aggregation of MDH and insulin substrate as described previously (1, 2). β-Galactosidase assay was performed with o-nitrophenyl β-
Measurement of intracellular ROS levels and determination of cellular protein carbonylation
Intracellular ROS levels were measured using H2DCFDA (48). Protein carbonylation assays were performed as described (47).
Oxidative stress-dependent structural switch of CgPrx in vivo
To determine oxidative stress-dependent structural switch of CgPrx in vivo, C. glutamicum wild-type, Δprx(pXMJ19-His6-prx), and ΔmshCΔprx(pXMJ19-His6-prx) strains were cultured in MM containing 1% glucose at 30°C. Bacterial cells were grown to mid-exponential phase and split into 80-ml aliquots for H2O2 treatment (50–200 mM, 10 min). The stressed samples were harvested immediately by centrifugation. For recovery, the stressed samples were inoculated into fresh MM containing 25 μg/ml streptomycin at 30°C for 2 h and then harvested. Crude cell lysates were subjected to nonreducing SDS-PAGE, and the structural properties of CgPrx were visualized by immunoblotting using anti-His antibody and analyzed by MALDI-TOF-TOF MS/MS (Voyager-DE STR; Applied Biosystems, Waltham, MA). Structure properties were also visualized for endogenous CgPrx by using an anti-CgPrx rabbit polyclonal antibody generated and affinity purified according to the method described previously (16).
Mycothiolation of CgPrx in vivo and in vitro
Iodoacetamide (IAM)-biotin-tagged demycothiolated proteins were obtained as described (17). Briefly, Δprx(pXMJ19-His6-prx) and ΔmshCΔprx(pXMJ19-His6-prx) strains were grown aerobically to an A 600 of 0.9. Each culture was divided into two parts, one was exposed to 120 mM H2O2 for 10 min at 30°C, while the other was grown without H2O2 (negative control). Cells were harvested and resuspended in urea/3-(3-cholamidopropyl) dimethylammonio-1-propanesulfonate (CHAPS) alkylation buffer (100 mM Tris-HCl, pH 8.0, 1 mM EDTA, 8 M urea, 1% CHAPS, 100 mM IAM) for 30 min in the dark before sonication on ice. Protein extracts were obtained by centrifugation and continued to be alkylated for 30 min in the dark. His6-Prx was purified using His·Bind Ni-NTA resin (Novagen, Madison, WI) in accordance with the manufacturer's instructions. Purified His6-Prx was demycothiolated using 25 μM recombinant Mrx1 in the presence of 1 mM MSH, 250 μM NADPH, and 5 μM Mtr for 30 min at room temperature. The mixture was then treated with 5 mM biotin-maleimide dissolved in dimethyl sulfoxide for 30 min. Unreacted biotin-maleimide was removed by ice-cold acetone precipitation three times for 1 h, followed by centrifugation at 10,000 g for 30 min. The pellet was redissolved in Tris-HCl buffer, separated using nonreducing SDS-PAGE, and detected by Western blotting with streptavidin-horseradish peroxidase (1:300; Thermo, Rockford, IL). The resulting His6-Prx (10 μM) was also separated by reducing SDS-PAGE and detected by Western blotting with anti-His antibody or stained with Coomassie Brilliant Blue. Protein bands were excised from the gel, in-gel digested with trypsin, and analyzed using MALDI-TOF-TOF MS/MS. In vitro mycothiolation of CgPrx was conducted according to the method of Chi et al. (7). MSH was purified from C. glutamicum as described (13).
H2O2 scavenging by whole cells
C. glutamicum strains were grown to stationary phase in MM containing 1% glucose at 30°C. Then, 5 and 120 mM H2O2 was added, respectively, and continued to incubate at 30°C in 100 rpm. At intervals, aliquots were immediately harvested, diluted when necessary, and assayed for H2O2 content by the ferrous xylenol orange assay (49).
Redox response of OxyR
WT(pXMJ19-His6-oxyR) and Δprx(pXMJ19-His6-oxyR) strains were grown to stationary phase and treated with different concentrations of H2O2 for 10 min at 30°C. Proteins extracted from the cells with TissueLyser II (Qiagen, Hilden, Germany) were reacted with 15 mM AMS in the dark (11), resolved on nonreducing SDS-PAGE, and then detected by Western blotting with anti-His antibody.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31270078 and 31500087), Key Science and Technology R&D Program of Shaanxi Province, China (2014K02-12-01), and the Natural Science Foundation of Shandong Province, China (ZR2015CM012).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.