
Abstract
Aims:
Ischemic postconditioning (iPoC) is known to mitigate ischemia–reperfusion (IR) injury of the liver, the mechanisms of which remain to be elucidated. This study explored the role of microRNA-183 (miR-183) in the protective mechanism of iPoC.
Results:
Microarray analysis showed miR-183 was robustly expressed in rats' livers with iPoC. miR-183 repressed the mRNA expression of Apaf-1, which is an apoptosis promoting factor. Using an oxygen–glucose deprivation (OGD) injury model in Clone 9 cells, hypoxic postconditioning (HPoC) and an miR-183 mimetic significantly decreased cell death after OGD, but miR-183 inhibitors eliminated the protection of HPoC. The increased expression of Apaf-1 and the downstream activation of capsase-3/9 after OGD were mitigated by HPoC or the addition of miR-183 mimetics, whereas miR-183 inhibitor diminished the effect of HPoC on Apaf-1-caspase signaling. In the in vivo experiment, iPoC and agomiR-183 decreased the expression of serum ALT after liver IR in the mice, but antagomiR-183 mitigated the effect of iPoC. The results of hematoxylin and eosin and TUNEL staining were compatible with the biochemical assay. Moreover, iPoC and agomiR-183 decreased the expression of Apaf-1 and 4-HNE after IR injury in mouse livers, whereas the antagomiR-mediated prevention of miR-183 expression led to increased protein expression of Apaf-1 and 4-HNE in the postischemic livers.
Innovation:
Our experiment showed the first time that miR-183 was induced in protective postconditioning and reduced reperfusion injury of the livers via the targeting of apoptotic signaling.
Conclusion:
miR-183 mediated the tolerance induced by iPoC in livers via Apaf-1 repressing. Antioxid. Redox Signal. 26, 583–597.
Introduction
R
Ischemic postconditioning (iPoC) refers to a controlled reperfusion performed at the late phase of ischemia or early reperfusion period to increase the tolerance to reperfusion injury. In 2003, Zhao et al. showed that brief, interrupted reperfusion performed immediately in the initial reperfusion period after ischemia could mitigate myocardial infarct size by 44% (38). Although the protective effects of iPoC on reperfusion injury have also been shown in noncardiac organs, including the liver (6), the mechanism of iPoC is not clear. Because iPoC works within a relatively short time window, its clinical application is limited. A thorough understanding of the protective mechanisms underlying postconditioning would be helpful in the development of novel and practical therapies for reperfusion injury of the liver.
Hepatic postconditioning robustly induces microRNA-183, which represses proapoptotic Apaf-1 and increases the ischemic tolerance. This study demonstrates the translational role that miRNAs play in coordinating adaptation to ischemic diseases.
microRNAs (miRNAs) are a class of endogenously expressed, small, noncoding RNAs that are involved in translational repression of messenger RNA (2). The roles of miRNAs in the regulation of development (25) and malignant disease (7) have been explored. For example, a distinct set of hepatic miRNAs has been found to have regulatory roles in hepatocellular carcinoma (11, 35). Recently, increasing evidence has indicated that miRNAs are associated with ischemic diseases, including stroke (9, 18), heart dysfunction (29), and rejection after liver transplantation (10). Moreover, it has been shown that the injection of endogenous miRNAs isolated from the ischemia-preconditioned heart reduces the infarct size of the myocardium after index ischemia, and miRNA treatment increases the protein expression of eNOS and heat shock proteins (37). Based on these findings, we hypothesize that the modulation of miRNAs and their targets might play a protective role in the postconditioning of livers. In this study, we identified miRNA-183 and its target, Apaf-1, in the protective mechanism of iPoC in the liver.
Results
microRNA expression profiles in rats' liver subjected to ischemia–reperfusion injury only or iPoC
A rat model of partial liver ischemia–reperfusion (IR) was performed. Pairwise comparisons of the miRNA expression between liver samples of the IR group and the iPoC group after 240 min of reperfusion were performed. From the miRNA microarray, 34 miRNAs in the iPoC group were significantly differentially expressed (12 upregulated and 22 downregulated, fold change >20.5) compared with those of the IR group (Table 1). Among the 12 upregulated miRNAs, miR-183 was the most highly upregulated (9.078-fold change). Real-time quantitative polymerase chain reaction (qPCR) further validated the overexpression of miR-183 and the depressed expression of miR-141 in the reperfused livers of the iPoC group compared with the expression in the livers of the IR group (Fig. 1).
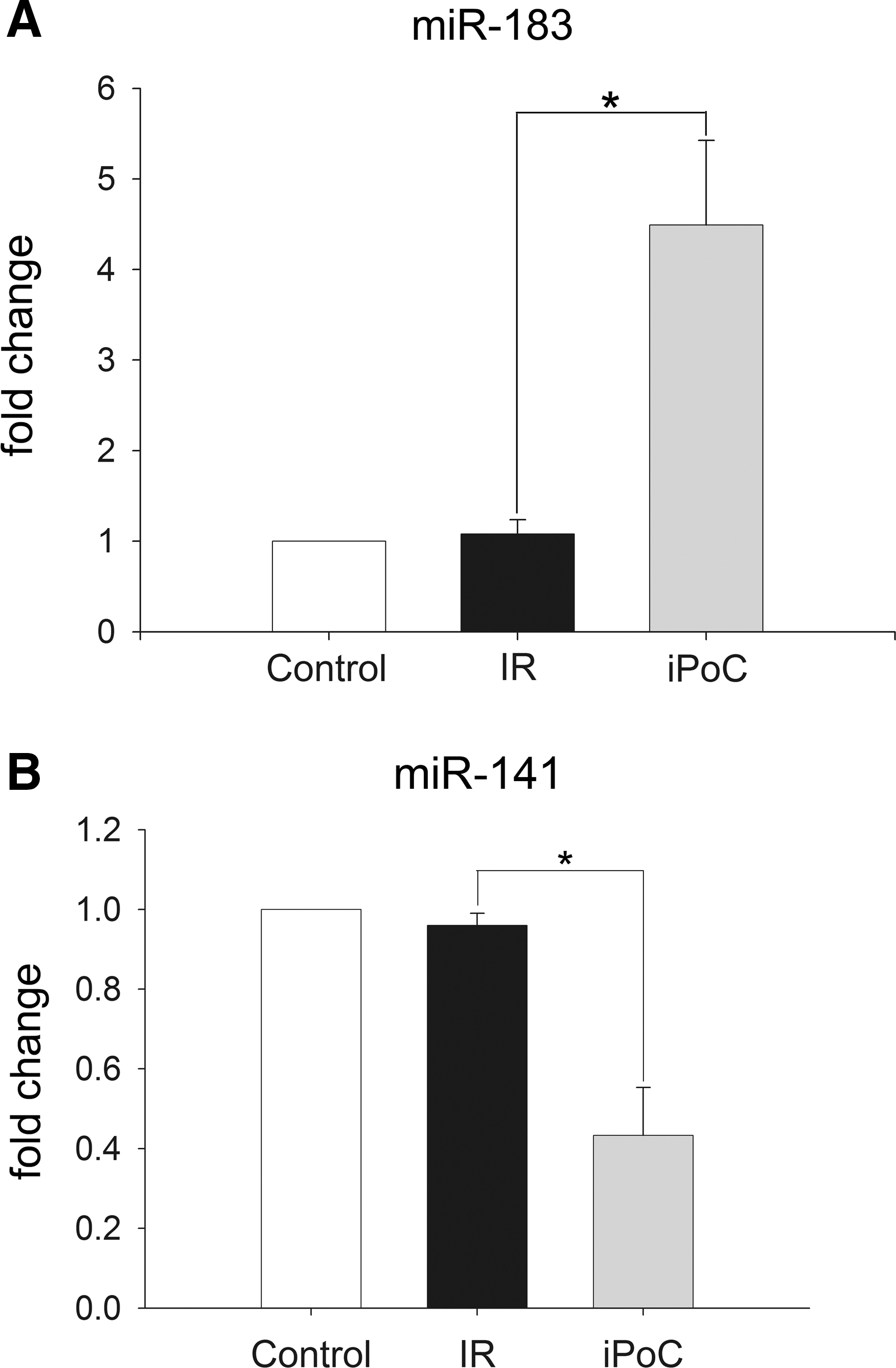
Data were expressed as fold change of microRNA expression compared to that of IR group.
microRNAs expressed in human, rat, and mouse.
microRNAs expressed in rat and mouse.
microRNAs expressed in human and rat.
iPoC, ischemic postconditioning; IR, ischemia–reperfusion.
The expression of Apaf-1, a predicted target of miR-183, in postischemic rat livers
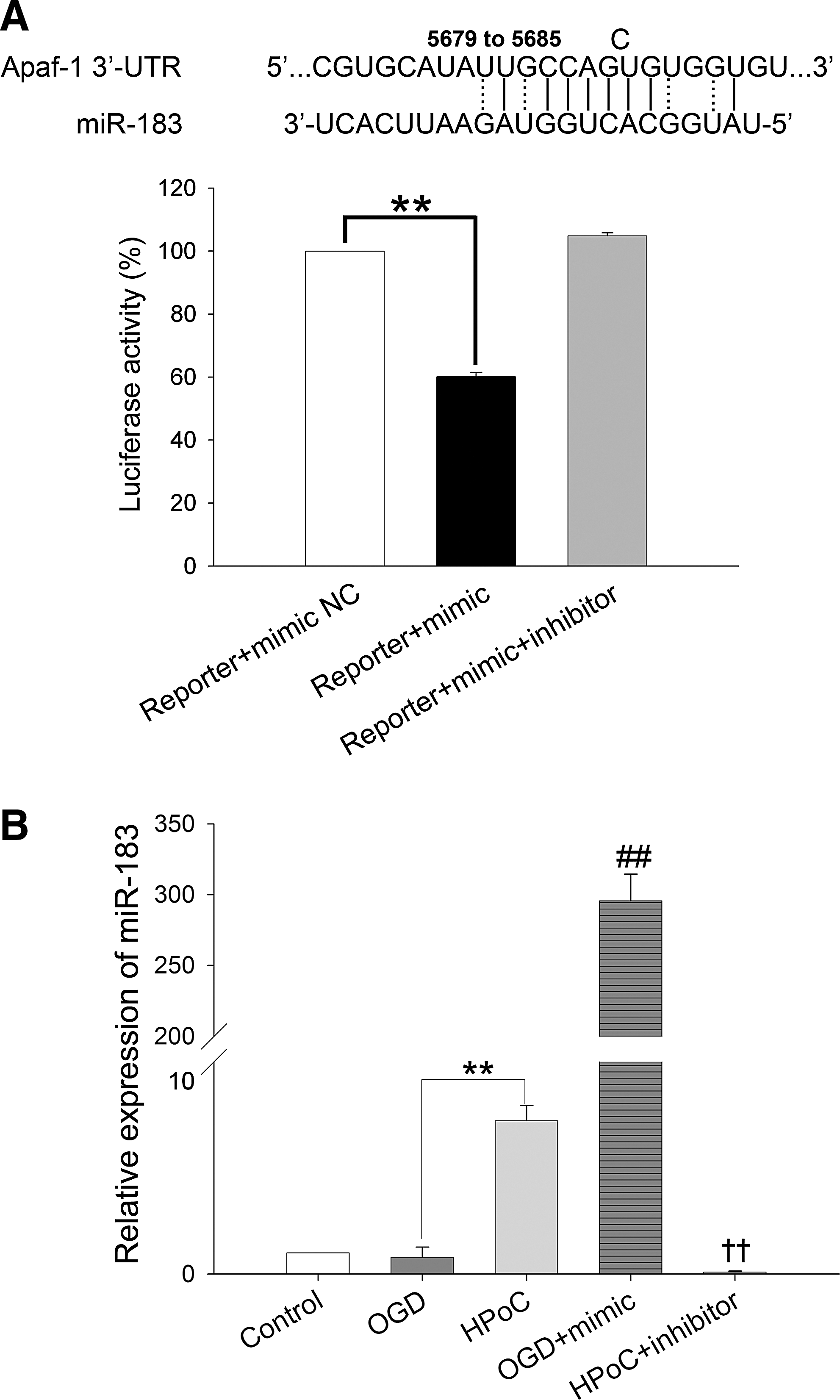
Apaf-1 is one of the predicted targets of miR-183 (

Hypoxic postconditioning and miR-183 mimics attenuate cell death after oxygen–glucose deprivation injury in Clone 9 cells
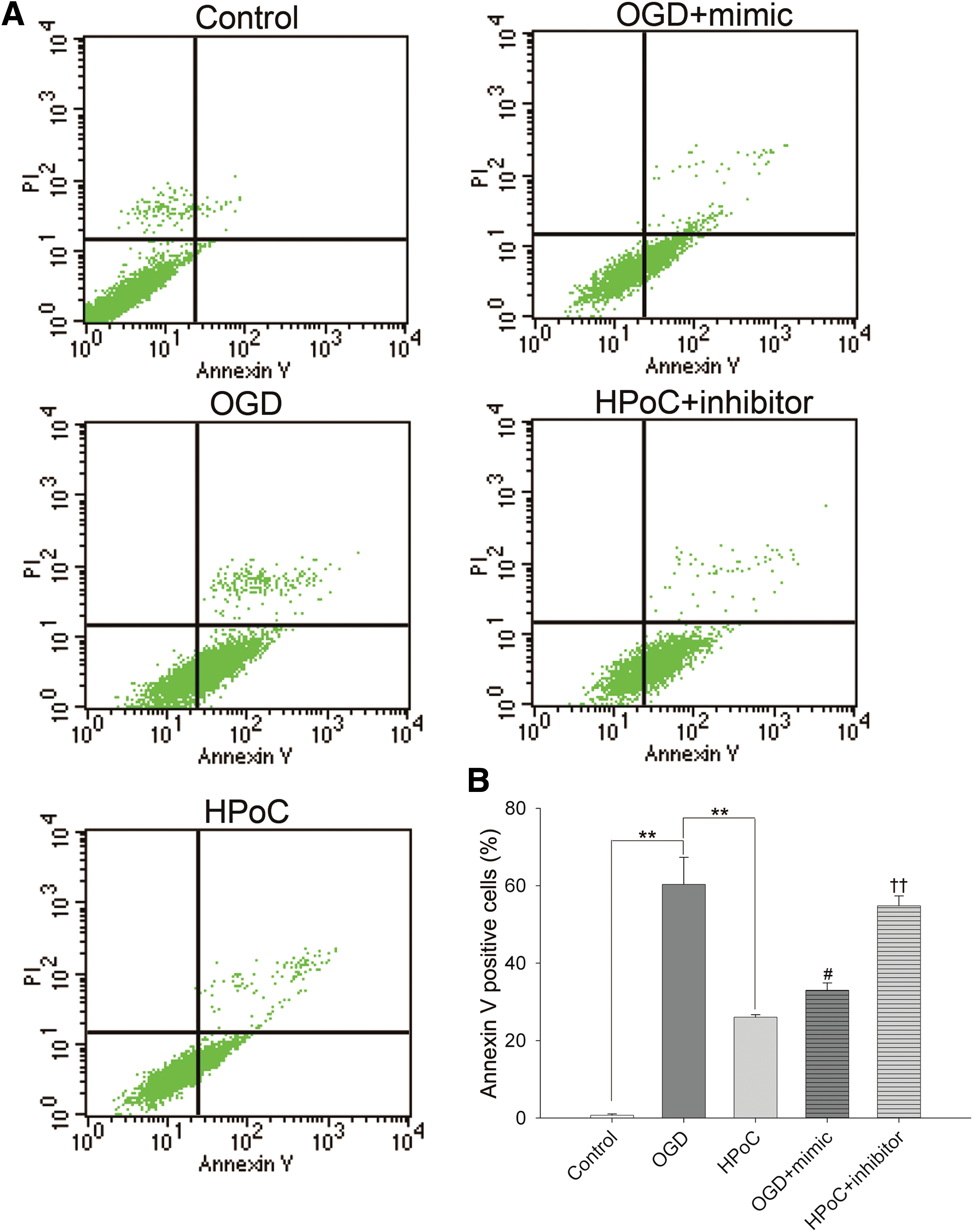
In vitro hypoxic postconditioning (HPoC) in an oxygen–glucose deprivation (OGD) model was established to mimic in vivo iPoC. As evaluated by Annexin V and PI staining (Fig. 4), the percentage of cell death in the HPoC group (26.04% ± 0.66%, p < 0.01) and the OGD + miR-183 mimic group (33.03% ± 1.86%, p < 0.05) significantly decreased when compared with that of the OGD group (60.33% ± 6.98%). The protective effect of HPoC was mitigated by the addition of miR-183 inhibitors (54.82% ± 2.51% in the HPoC + miR-183 inhibitor group vs. 26.04% ± 0.66% in the HPoC group, p < 0.01). Moreover, terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining was performed to further assess cell death. As shown in Figure 5, the percentage of TUNEL-positive cells in the HPoC group (44.85% ± 2.27%) and the OGD + mimic (56.26% ± 1.63%) group was less than in the OGD group (70.46% ± 1.71%, p < 0.01). miR-183 inhibitors eliminated the protective effects of HPoC, which led to increased cell death in the HPoC + inhibitor group (66.74% ± 1.47% vs. 44.85% ± 2.27%, p < 0.01). These findings demonstrated that HPoC protected Clone 9 cells from OGD-induced injury and the protective effects were mimicked by miR-183 mimics or abrogated by miR-183 inhibitors.

The effects of HPoC and miR-183 mimics on the expression of Apaf-1 and apoptosis signaling
The expression of miR-183 and Apaf-1 in the in vitro HPoC model was evaluated. Similar to our observations in the in vivo system, after HPoC, the expression of miR-183 significantly increased compared to that of the OGD group (7.22 ± 0.72 vs. 0.79 ± 0.48, p < 0.01) (the expression of miR-183 in the control group was normalized to 1.0) (Fig. 3B). However, Apaf-1, the predicted target of miR-183, was overexpressed after OGD. HPoC or miR-183 mimics significantly reduced the overexpression of Apaf-1 after OGD, whereas the repressive effect of HPoC on Apaf-1 expression was inhibited by the addition of miR-183 inhibitors (Fig. 6, upper panel). The results showed that the HPoC-induced repression of Apaf-1 was regulated by miR-183.
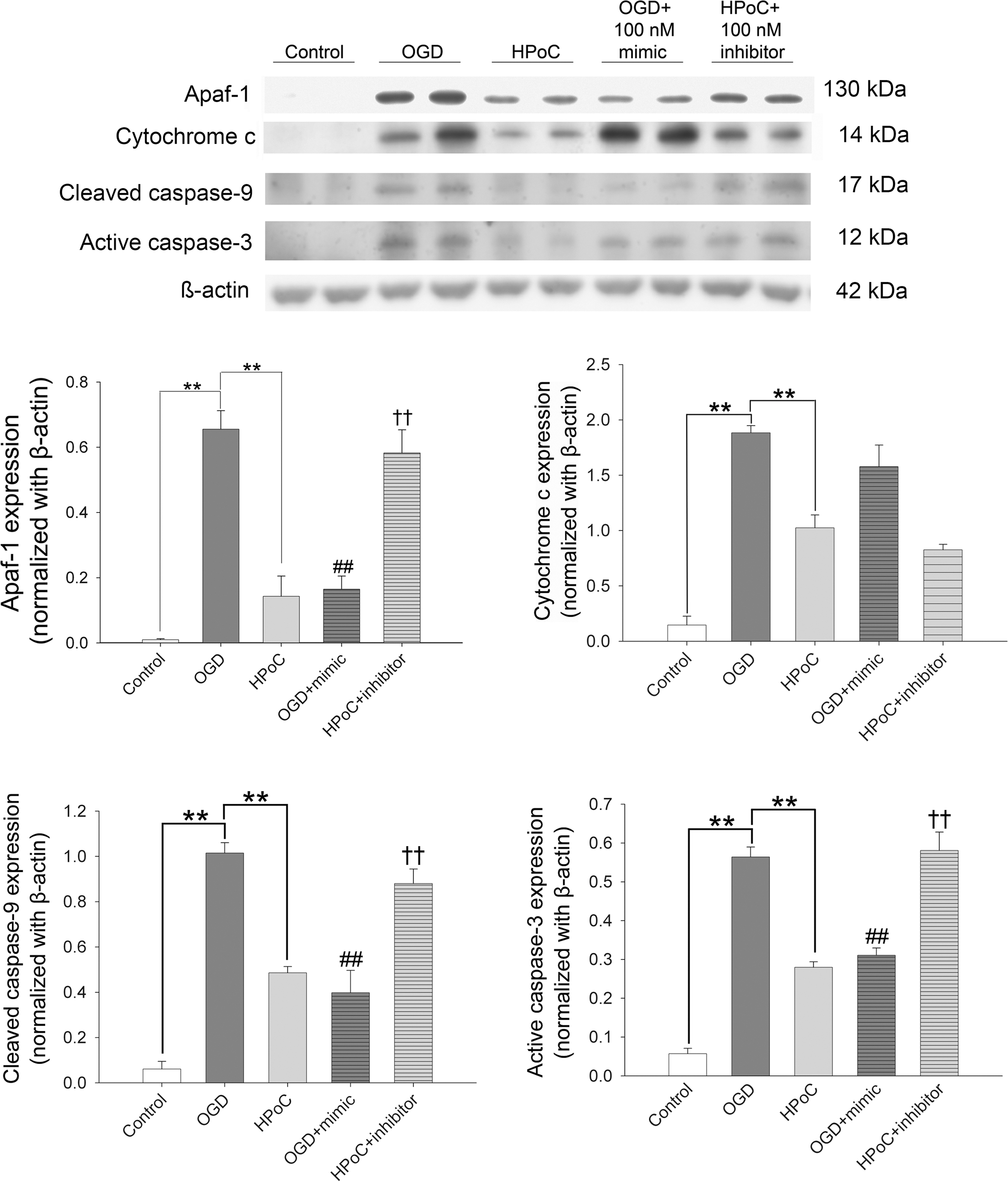
Moreover, the signaling of Apaf-1, including cytochrome c release (upstream), cleavage of caspase-9 and activation of caspase-3 (downstream signaling), after OGD was evaluated by Western blot analysis. As shown in Figure 6, the increased release of cytochrome c into the cytosol after OGD was mitigated by HPoC but not by the addition of miR-183. miR-183 inhibitors did not reverse the repressive effect of HPoC on cytochrome c release. However, the expression of cleaved caspase-9 and active caspase-3, which occurs downstream of Apaf-1 signaling, was regulated by miR-183. The expression of cleaved caspase-9 and active caspase-3 increased after OGD compared with the control group (1.02 ± 0.05 vs. 0.06 ± 0.03 for cleaved caspase-9, p < 0.01; 0.56 ± 0.03 vs. 0.06 ± 0.01 for active caspase-3, p < 0.01). Both HPoC and the addition of miR-183 mimics effectively reduced the increase in the cleavage of caspase-9 (0.49 ± 0.03 in the HPoC group, 0.40 ± 0.10 in the OGD + miR-183 mimic group) and the activity of caspase-3 (0.28 ± 0.01 in the HPoC group, 0.31 ± 0.02 in the OGD + miR-183 mimic group) after OGD, whereas the addition of miR-183 inhibitors (0.88 ± 0.06 of cleaved caspase-9; 0.58 ± 0.05 of active caspase-3) abrogated the effects conferred by HPoC. These results indicated that HPoC mitigated cellular injury partly through miR-183-Apaf-1-caspase signaling.
iPoC and agomiR-183 reduced reperfusion injury in the livers of mice
In the mouse model, the expression of miR-183 in the liver of the iPoC group, as measured by qPCR, was 4.7-fold higher than that of the IR group (p < 0.05) (Fig. 7). Moreover, iPoC and agomiR-183 infusion both decreased the level of serum alanine transferase (ALT) after partial liver IR injury when compared with that of the IR group (145.6 ± 9.5 U/L in the iPoC group and 268.0 ± 8.1 U/L in the IR + agomiR-183 group vs. 513.4 ± 39.4 U/L in the IR group, p < 0.005). Cotreatment with antagomiR-183 diminished the protective effect of iPoC (381.6 ± 10.6 U/L in the iPoC + antagomiR-183 group vs. 145.6 ± 9.5 U/L in the iPoC group, p < 0.001) (Fig. 8).


Morphologically, hepatic IR injury in mice was observed by hematoxylin and eosin (H&E) staining and quantitated by Suzuki's classification. The livers in the IR group showed severe congestion, vacuolation, and hepatocyte necrosis, whereas the livers in both the iPoC and IR + agomiR-183 groups showed only mild changes. The Suzuki scores in congestion, vacuolation, and necrosis were 1.6 ± 0.2, 2.1 ± 0.1, and 2.4 ± 0.1, in the IR group; 0.9 ± 0.2, 1.5 ± 0.1, and 1.5 ± 0.1 in the iPoC group; 0.8 ± 0.2, 1.7 ± 0.1, 1.9 ± 0.1 in the IR + agomiR-183 group; and 1.8 ± 0.1, 2.4 ± 0.1, 2.5 ± 0.1 in the iPoC + antagomiR-183 group, respectively (Fig. 9). Moreover, the increase in TUNEL-positive cells in the livers after partial liver IR injury was significantly reduced by iPoC or the addition of agomiR-183, and the antagomiR-183 mitigated this effect of iPoC. The percentages of TUNEL-positive cells in the livers were 3.2% ± 0.5% in the control group, 62.8% ± 0.3% in the IR group, 40.7% ± 0.5% in the iPoC group, 43.0% ± 0.6% in the IR + agomiR-183 group, and 65.2% ± 0.8% in the iPoC + antagomiR-183 group (p < 0.01) (Fig. 10). These results indicate that iPoC protects the liver partly through the induction of miR-183.

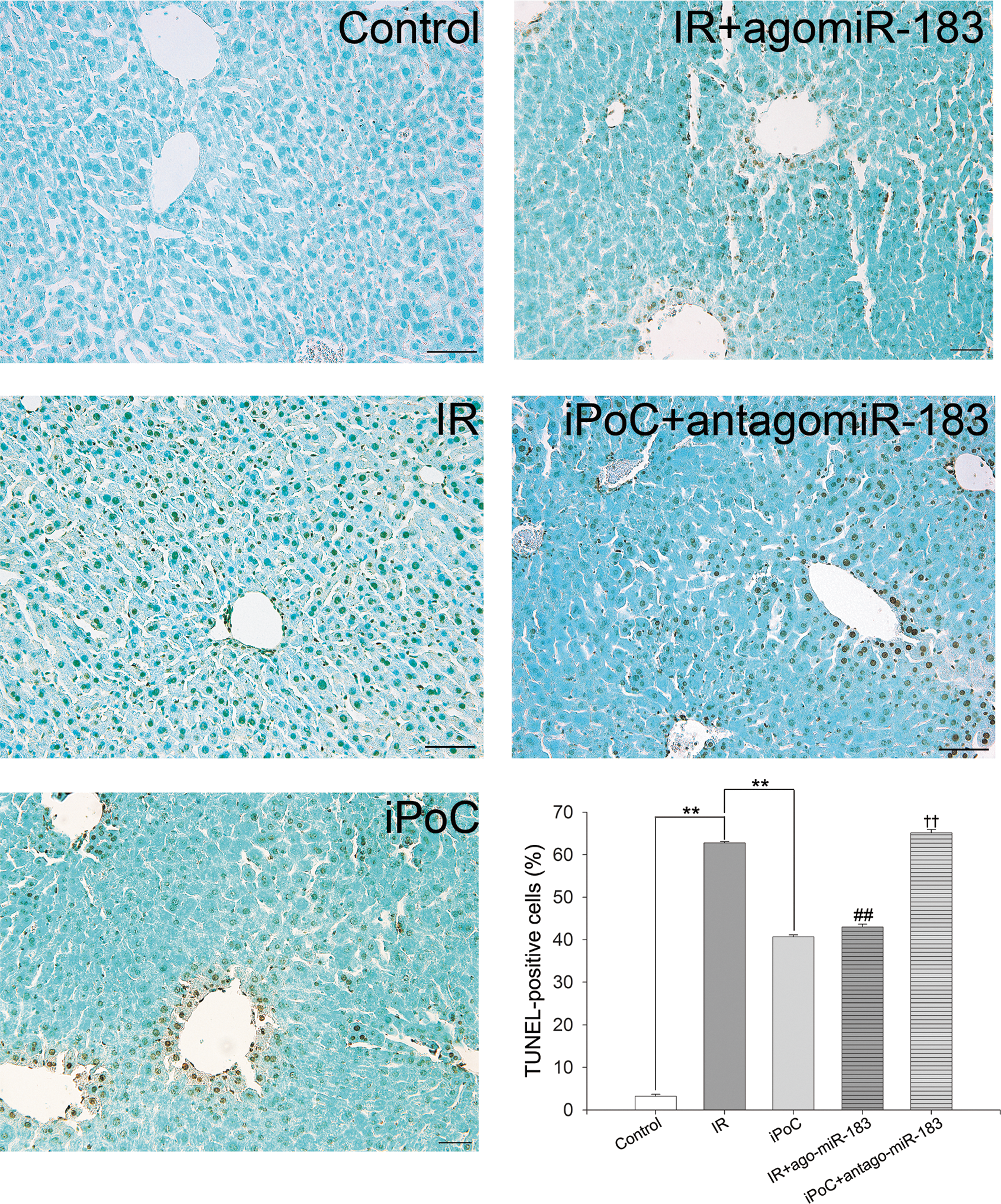
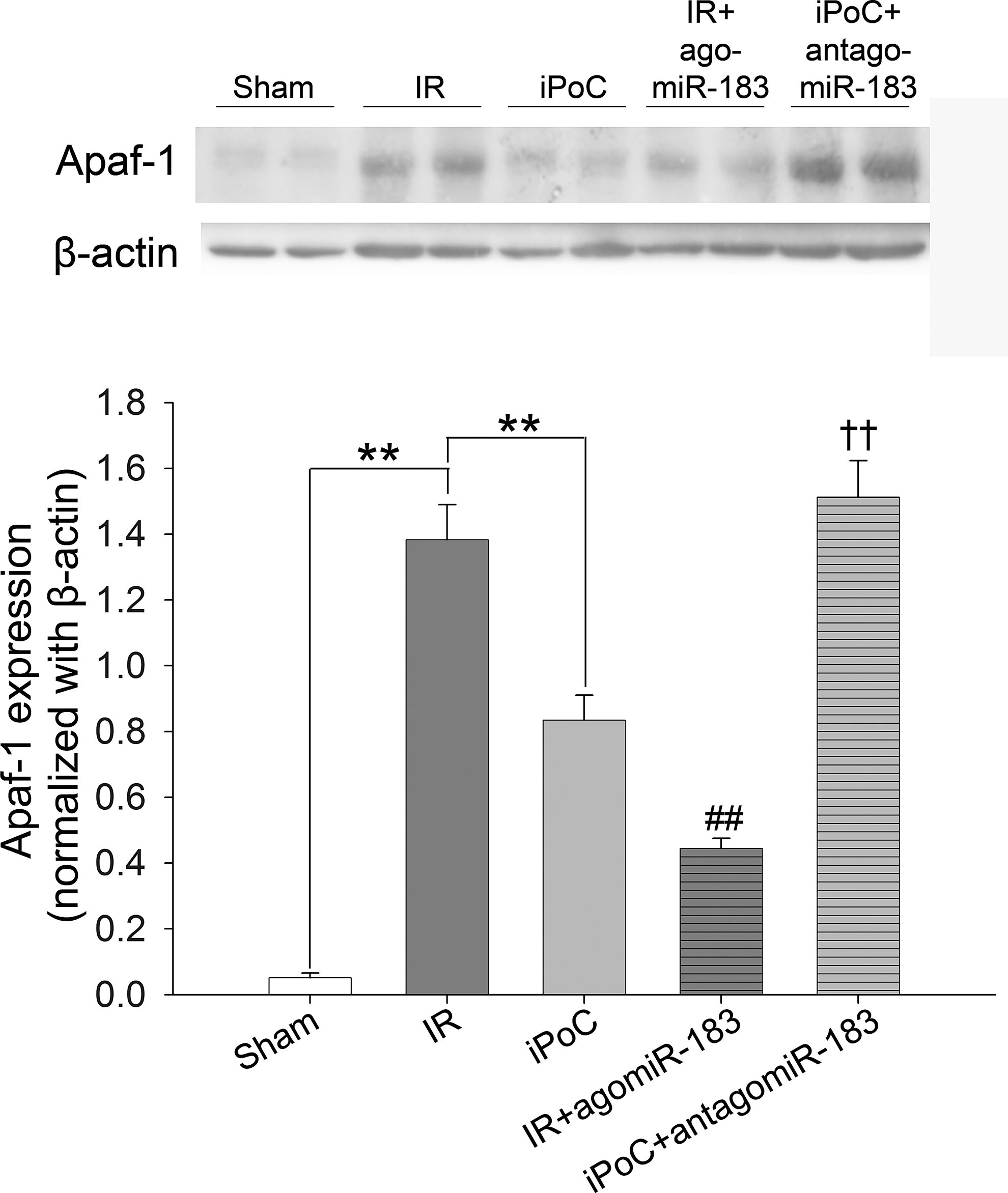
The expression of Apaf-1, the target of miR-183, in the livers of mice was detected by Western blot analysis (Fig. 11). Compared with the IR group, iPoC and agomiR-183 infusion decreased the hepatic expression of Apaf-1 after IR injury, whereas cotreatment with antagomiR-183 abrogated the effect of iPoC on Apaf-1 expression in the livers.
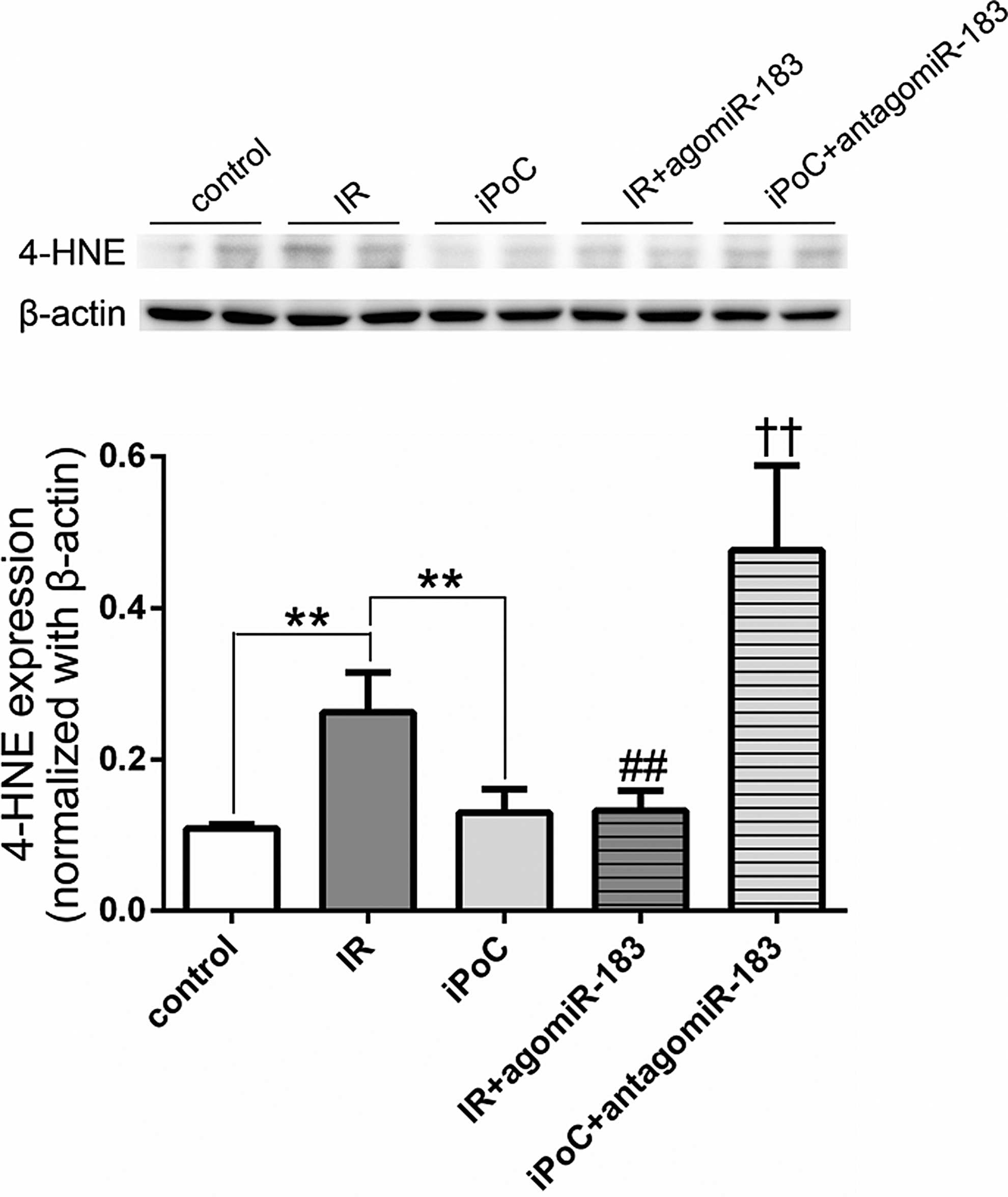
iPoC and agomiR-183 reduced oxidative stress in the livers of mice
To explore the protective mechanism and possible trigger of miR-183 activation in the iPoC, we measured the expression of 4-hydroxy-2-nonenal (4-HNE), an unsaturated hydroxyalkenal produced by lipid peroxidation in cells, in mice undergoing liver IR injury. As shown in Figure 12, iPoC as well as the addition of agomiR-183 reduced the increase of 4-HNE after liver IR (0.12 ± 0.03 in iPoC group; 0.13 ± 0.02 in IR + agomiR-183 group; 0.26 ± 0.05 in IR group, p < 0.01). The addition of antagomiR-183 mitigated the effect of iPoC on the expression of 4-HNE (0.48 ± 0.11 in iPoC + antagomiR-183 group vs. 0.12 ± 0.03 in iPoC group, p < 0.01). The results indicated that iPoC reduced oxidative stress partly through the effect of miR-183.
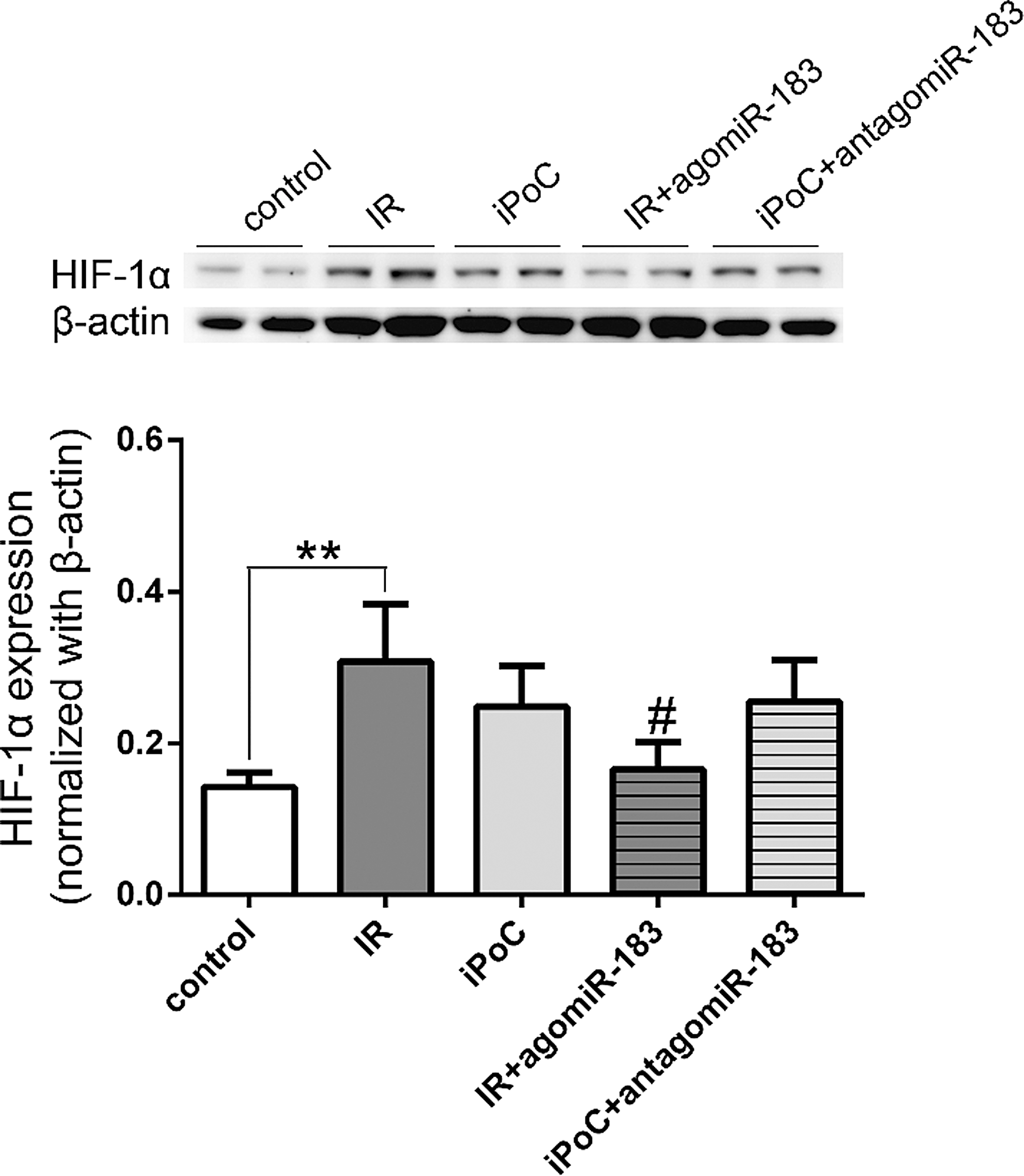
HIF-1α is a main adaptive regulator of cells undergoing oxidative stress. Whether miR-183 worked through the HIF-1α signaling was explored. We examined the expression of HIF-1α in mice undergoing liver IR injury. As shown in Figure 13, the expression of HIF-1α increased in both IR and iPoC groups when compared to that of the control, but the difference between IR and iPoC groups was not significant. The addition of miR-183 to IR group reduced the expression of HIF-1α (0.17 ± 0.03 in IR + agomiR-183 group vs. 0.31 ± 0.08 in IR group, p = 0.015), while antagomir-183 did not markedly reduce the expression of HIF-1α (0.26 ± 0.05 in iPoC + antagomiR-183 group vs. 0.25 ± 0.05 in iPoC group, p = 0.862). The results indicated that miR-183 may regulate the expression of HIF-1α, but other hypoxamirs may have an additive or antagonistic effect on HIF-1α expression in liver IR injury.
Discussion
This study demonstrates the role of miR-183 in the protective mechanisms of iPoC in hepatic IR injury. The main findings are that (a) postconditioning (ischemic or hypoxic) upregulated the expression of miR-183 and decreased the expression of its target, Apaf-1; (b) HPoC mitigated OGD injury in Clone 9 cells, partly through the induction of miR-183; (c) miR-183 reduced OGD-induced cell death by targeting Apaf-1, which is involved in proapoptotic signaling; and (d) in vivo, iPoC protected the livers of mice from IR injury partly by upregulating miR-183, which in turn repressed the expression of proapoptotic Apaf-1. Our results indicated that the tolerance to IR injury resulting from postconditioning is related to the regulation of certain miRNAs.
Although postconditioning in patients undergoing liver resection has been shown to have hepatoprotective effects in a randomized clinical trial (6), its exact mechanisms are under active investigation. Triggering of intracellular signaling, by players such as adenosine (8), the prolonging of the acidosis state in the reperfusion period (15), and the prevention of the opening of mPTP (20) have been proposed to be responsible for the protective effects. miRNAs are known to play important biological roles in the pathology of diseases of the liver, including hepatitis (17) and hepatocellular carcinoma (36). Recently, studies have shown the regulated expression of miRNAs in preconditioned livers (33) and the involvement of protectomiRs in postconditioning-induced cardioprotection (30).Therefore, the modulation of miRNA expression offers a potential therapeutic option for reperfusion injury. miR-183 is a member of a highly conserved miRNA cluster (miR-183, miR-96, and miR-182) located on human chromosome 7q32.3, which was first identified in the sensory organs (34). Previous studies have shown that miR-183 is oncogenic by targeting the tumor-suppressor programmed cell death 4 (PDCD4) in hepatocellular carcinoma (21), gastric cancer (13), and esophageal squamous cell carcinoma (27). However, limited studies about its roles in ischemic diseases have been reported. Our study is the first to show that miR-183 exerts its IR injury-sparing effect through antiapoptotic signaling.
The regulation of miRNAs and their interactions is complex. An miRNA can bind to a large number of mRNAs, in a manner dependent on a signature sequence in the 3′-UTR region of the target mRNAs. A specific mRNA can be controlled by many miRNAs if they contain complementary binding sites. Structural studies have estimated that most miRNAs can inhibit ∼200 mRNAs (20). Although our study has indicated a relationship between miR-183 and its target Apaf-1 in the postischemic liver, the protection afforded by the miR-183 mimetic is only partial, and thus, other miRNAs and mRNAs might also play a role in the damage-sparing mechanism.
We observed that several miRNAs responded rapidly to iPoC, and their expression changed by a significant magnitude. Ischemia seemed to have a greater effect on genes that transcribe miRNAs. However, the exact mechanisms that regulate the transcription of miRNA after ischemia or iPoC are not known. It has been shown that after focal brain ischemia, the synthetic machinery for miRNAs (RNases Drosha and Dicer, and cofactor Pasha) does not change, but the expression of miRNAs changes in a temporal pattern (9). In our in vitro experiment, the expression of miR-183 significantly increased 90 min after OGD and HPoC (data not shown). It has been postulated that ischemia might alter the expression of certain mRNAs or proteins that cross talk or form a feed-forward loop with miRNAs at the transcription level (9).
The reduction of oxidative stress is one of the known protective mechanisms of pre/postconditioning. We measured the association of miR-183 expression and the redox status in our experimental setting, which showed that both iPoc and agomiR-183 mitigated the production of 4-HNE, a lipid peroxidation product, in the liver IR injury. The effect of iPoC on reducing lipid peroxidation was abrogated by the addition of antagomir-183. The results showed that miR-183 protected the livers through the regulation of redox status in the IR injury. On the contrary, it is known that RNA molecules could be modified structurally by reactive oxygen species (ROS). Recent study showed that the changed ROS level could modulate the transcription of microRNAs (16). The temporal relationship between ROS production and microRNA biogenesis was not explored in the present study.
Hypoxia inducible factor (HIF)-1α is crucial for cellular response to oxidative stress. We examined the expression of HIF-1α in mice undergoing liver IR injury. IPoC as well as IR induced the expression of HIF-1α in livers. AgomiR-183 reduced the HIF-1α expression induced by liver IR, but antoagomir-183 barely changes the HIF-1α production in the iPoC setting. The results indicated that miR-183 may regulate the expression of HIF-1α, but agomiR-183 could not modulate the degradation of HIF-1α. In contrast, the transcriptional regulation of miRNA expression under ischemic conditions can be HIF dependent. Recent evidence indicates that HIF may induce the transcription of multiple hypoxamirs through direct binding to HREs in their respective promoters (28). The induced hypoxamirs may reciprocally have an additive or antagonistic effect on HIF-1α expression. For example, a research showed that miR-21 was a transcriptional target of HIF-1α, but the inhibition of miR-21 reduced HIF-1α expression in cardiomyocytes under hypoxia (23). Therefore, the cause and effect relationship of ROS or HIF-1α on the microRNA biogenesis remains to be explored in postconditioning on liver IR injury.
There are issues that remain to be solved in our study. First, reperfusion-induced liver injury may result in necrosis or apoptosis. In our study, both iPoC and the addition of agomiR-183 decreased apoptotic cell death in mice livers through repressing the expression of Apaf-1. Moreover, iPoC mitigated the degree of necrosis in the liver cells after IR injury (Suzuki score: 1.5 ± 0.1 vs. 2.4 ± 0.1; serum ALT: 45.6 ± 9.5 U/L vs. 513.4 ± 39.4 U/L). The administration of agomiR-183 resulted in a significant but less protective effect on hepatic necrosis, through a mechanism not explored in this study. Second, apoptosis is an ATP-dependent cell death program that is related to mitochondria dysfunction and caspase activation. Our results showed that HPoC and the addition of an miR-183 mimetic decreased the expression of Apaf-1 and caspase-3/9 activation. However, whether miR-183 changes mitochondrial function or ATP biogenesis remains to be determined. Third, although the association of miR-183 and Apaf-1 signaling in the postischemic liver was shown, the direct involvement of Apaf-1 in the postconditioning mechanism was not explored in this study. Further Apaf-1 knockdown experiment would be helpful. Fourth, the effects and roles of nonparenchymal cells in postconditioning were not explored in this study. The studies of alternation of sinusoid circulation following iPoC were limited. Arai et al. showed that ischemic preconditioning also protects sinusoid endothelium and suppresses Kupffer cell activation in reperfusion injury (3). Fifth, prolonging acidosis (14) and signaling ROS were proposed possible mechanisms involved in protective postconditioning. Whether miR-183 worked through the maintaining acidosis was not explored in this study. Sixth, the oxygen threshold for the activation of hypoxamirs was not tested in this study. It has been reported that either 0.1% or 1% O2 hypoxia could repress the expression of microRNAs (4) or repress the enzymes involved in biogenesis of microRNAs (28) in breast cancer cell lines. However, there were no reports to define the oxygen threshold for increasing the biogenesis of microRNAs in livers following conditioning maneuver.
In summary, the present study demonstrated that miR-183 has antiapoptotic functions, which include repressing the expression of Apaf-1 in the setting of hepatic postconditioning.
Materials and Methods
Models of IR injury and iPoC of the liver
Partial liver IR and iPoC procedures were established on rats (for microarray study) and mice (for functional assays) separately. The rats were randomly assigned to three groups (n = 6 per group). In the control group, rats underwent laparotomy only. In the IR group, rats underwent 45-min liver ischemia followed by 240-min reperfusion. In the iPoC group, rats received 45-min liver ischemia and three cycles of 1-min reperfusion followed by 1-min ischemia at the end of ischemia. For the functional study, the mice were randomly assigned to five groups (n = 6 per group). In the control group, mice underwent laparotomy only. In the IR group, mice received 30-min liver ischemia followed by 60-min reperfusion. In the iPoC group, mice underwent 30-min liver ischemia and three cycles of 30-s reperfusion followed by 30-s ischemia at the end of ischemia. In the IR + agomiR-183 group, agomiR-183 (1 mg/kg) was infused via splenic injection 1 h before ischemia started. In the iPoC + antagomiR-183 group, antagomiR-183 (2 mg/kg) was infused via splenic injection 1 h before ischemia started. The protocols are depicted in Figure 14.

Animal care and preparation
Male Wistar rats weighing 200–250 g and male C57BL/6 mice weighing 20–25 g were used. All animal experiments and animal care were performed in accordance with the “Guides for the Care and Use of Laboratory Animals” (published by National Academy Press, Washington, DC, 1996). All protocols used in this study were approved by the Laboratory Animal Care Committee of the National Taiwan University College of Medicine.
Surgical procedures to induce partial hepatic IR injury
For the microarray study, a modified partial hepatic IR was performed on rats (22). Wistar rats were anesthetized by intraperitoneal injection of sodium pentobarbital (35 mg/kg). The trachea was intubated to keep airway patent and the right jugular vein was catheterized for infusion of glucose with a rate of 20 ml/kg/h. Body temperature was maintained at 36.5–37.5°C using a heating pad. The carotid artery was catheterized for measurement of blood pressure and blood sampling. After a 30-min stabilization period, the left hepatic artery was isolated and partial hepatic IR injury was produced by placing an atraumatic microvascular clip across the origin of left hepatic artery for 45 min (ischemia period) and by removing the clip for 240 min (reperfusion period). In the postconditioning group, the rats underwent three cycles of 1-min reperfusion by releasing the clip across the left hepatic artery followed by 1-min ischemia of liver by clamping the left hepatic artery after 45-min partial liver ischemia. The animals in the control group received laparotomy and vascular isolation procedure only. The animals of IR group received partial liver IR injury for 45 min. In each group, six rats were used for measurement. Serum samples and hepatic tissues (including the ischemic and nonischemic lobes) were harvested at predetermined time intervals and prepared for assay.
For the functional assay, a mice model of partial liver IR injury was established. Male C57BL/6 mouse was anesthetized with pentobarbital (10 mg/kg, intraperitoneal injection). After laparotomy, the arterial branches to the left lateral and median lobes of the liver were clamped for 30 min (ischemia) and then released for 60 min (reperfusion) in the IR group. Three cycles of 30 s of ischemia (by clamping the vessels) followed by 30 s of reperfusion (by releasing the vascular clip) were performed at the end of ischemia in the iPoC group. The control group underwent laparotomy only. To explore the functional role of microRNA-183 in iPoC, the agonist (agomiR-183) and antagonist of miR-183 (antagomiR-183), complexed with the Invivofectamine 2.0 Reagent (Invitrogen; Life Technologies Corporation), were infused. After laparotomy, the spleen of the mice was exposed. In the IR + agomiR-183 group, agomiR-183 (1 mg/kg) at a volume of 100 μl was infused into the spleen 60 min before liver ischemia began. For the iPoC + antagomiR-183 group, antagomiR-183 (2 mg/kg) at a volume of 100 μl was infused into the spleen 60 min before liver ischemia and iPoC maneuver was performed. In each group, six mice were used for measurement. Serum samples and hepatic tissues were harvested at predetermined time intervals and prepared for assay.
microRNA microarray
Total RNA was extracted from liver samples of each group by the miRNeasy Mini Kit (Qiagen). The quantity and quality of total RNA were measured by NanoDrop 2000 (Thermo Scientific). The qualified RNAs were used for miRNA microarray using the miRCURY LNA™ microRNA Array kit (6th generation human, mouse & rat, #208401; Exiqon, Vedbaek, Denmark). The RNA samples of IR group and the iPoC group were labeled with Hy3 and Hy5 separately. After labeling, the samples were hybridized with the miRCURY LNA™ microRNA Array chips at 56°C for 16 h. The miRNA-hybridized chips were washed and scanned with GenePix 4000B (Molecular Devices). The data were analyzed by GeneSpring GX software (Agilent Technologies).
Target prediction
The miRNA mature sequence database was obtained from miRBase (
Quantitative real-time PCR
Quantitative real-time-PCR (qRT-PCR) assays were performed using the miScript PCR Starter Kit (Qiagen, No. 218193) for the rno-miR-183 and rno-mir-141 expression. Mature miRNAs were reversely transcripted according to the manufacturer's instruction. Briefly, 0.5 μg template RNA, 5 × miScriptHiSpec Buffer, 10 × miScriptNucleics Mix, RNase-free water, miScript Reverse Transcriptase Mix were mixed together and then incubated in a thermocycler (Applied Biosystems) for 60 min at 37°C, 5 min at 95°C, and held at 4°C. Real-time PCR was performed in a 10 μl mixture containing 1 μl 200 × diluted RT products, 1 μl 10 × miScript Universal primer, 5 μl 2 × QuantiTect SYBR Green PCR Master Mix, 1 μl forward primer (U6 and rno-miR-183), and 2 μl RNase-free water. The miRNA expression levels were determined by the 2−ΔΔCt method.
OGD and HPoC
To simulate the in vivo IR injury and iPoC maneuver, an in vitro OGD and HPoC model were established. Clone 9 cells, purchased from Bioresource Collection and Research Center (BCRC, Hsin-Chu, Taiwan), were grown and maintained in Ham's F-12K medium (Invitrogen) supplemented with 10% fetal bovine serum (Hyclone, Logan, Conn) and 1% penicillin G/streptomycin/amphotericin (Hyclone, Logan, Conn) in a humidified atmosphere with 5% CO2 at 37°C. Clone 9 is an epithelial cell line isolated from the normal liver of a young Sprague-Dawley male rat (32). The cell line exhibited uniform properties and was used for in vitro studies for oxidative injuries (5, 12).
For the induction of OGD, the Clone 9 cells were washed twice in balanced salt solution without glucose (BSS0, 118 mM NaCl, 1.8 mM CaCl2, 0.8 mM MgSO4, 5.4 mM KCl, 1 mM NaH2PO4, 14.7 mM NaHCO3, 10 mM HEPES, pH 7.4) and placed in a controlled atmosphere chamber (Plas Labs, Lansing, MI) for 20 h at 37°C. The chamber was preflushed with a gas mixture of 90% N2/5% CO2/5% H2 for 30 min until the oxygen concentration in the chamber was below 1%. Oximetry (Gastech Co., Ltd.) was used to monitor the oxygen concentration. The HPoC group received three cycles of 10 min of reoxygenation (at normoxic incubator) followed by 10 min of hypoxia at the hypoxic chamber after OGD. After OGD was performed, the BSS0 in the culture dish was replaced by Ham's F-12K medium, and the cells were cultured in a normoxic atmosphere for 2 h (reperfusion period). Samples were collected for measurement following 2-h reperfusion. In OGD + mimic group, miR-183 mimic (100 nM) was added to the Ham's F-12K medium at 24 h before OGD. In HPoC + inhibitor group, miR-183 inhibitor (100 nM) was added to the medium at 24 h before OGD. The grouping is depicted in Figure 15.

Transfection of rno-miR-183 mimics and rno-miR-183 inhibitors
The miRNA mimic, miRNA inhibitor, or negative control (abbreviated as NC), purchased from RiboBio (Guangzhou, China), was mixed with Lipofectamine 2000 separately. The mixture was added to dishes at a final concentration of 100 nM, followed by incubating for 6 h at 37°C. The cells were subjected to hypoxia 24 h after transfection, and total RNA, protein, or cells were collected for qPCR, Western blot, or flow cytometry.
Dual-luciferase reporter assay
Clone 9 cells (5 × 104/well) were cultured in 48-well plates for 24 h and then transfected with pMIR-RB-REPORT-Apaf-1-3′-UTR (1 μg/well) (RiboBio Co., Ltd., Guangzhou, China) and rno-miR-183 mimic (100 nM/well) (or a negative control, separately). The plasmid (pMIR-RB-REPORT-Apaf-1-3′-UTR) contained Firefly luciferase (reference gene) and Apaf-1 3′-UTR downstream from Renilla luciferase (reporter gene). After 24 h of transfection, the ratio of Renilla to Firefly luciferase activity was measured by a dual-luciferase assay system (Promega) according to the manufacturer's instruction.
Cell death analysis
The cell death after OGD was assessed by using the Annexin V-FITC Apoptosis Detection Kit (Strong Biotech Corporation). The Clone 9 cells were harvested, washed, and resuspended in binding buffer. Annexin V and propidium iodide were added to stain the cells and incubated for 15 min in the dark at 15–25°C. The stained cells were analyzed using a BD FACSCalibur flow cytometer (BD Biosciences, San Jose, CA). The percentage of the early apoptotic cells was calibrated by CellQuest (BD).
TUNEL assay of Clone 9 cells
An in situ cell death detection kit (Roche) was used to detect DNA fragmentation after OGD. Clone 9 cells cultured on glass coverslips coated with type I collagen (106 cells/well) were used. After treatment, the cells were washed with PBS and fixed in 4% paraformaldehyde at room temperature. The peroxidase activity was blocked by 3% H2O2, and then, the cells were permeabilized with 0.1% Triton-X-100. The cells were immersed in TUNEL reaction mix to label the fragment DNA at 37°C. To identify nuclear morphology, cells were counterstained with DAPI (2 μg/mL). Cell death was quantified by counting the percentage of TUNEL-positive nuclei under a confocal microscope (TCS SP5; Leica, Wetzlar, Germany). Values are shown as the means of counting of 50 fields.
Western blot analysis
The cytosolic and mitochondrial portions of samples were separated by the mitochondria isolation kit (Thermo Scientific) in cultured cells (26) or by differential centrifugation in liver tissue (31). The cytosolic expression of various proteins after OGD in Clone 9 cells or after liver IR was measured by Western blot analysis.
The extracted protein was quantified by Bradford assay. Equal amount (30 μg) of cytosolic proteins of each sample was resolved on gel by electrophoresis, transferred, and blocked in TBST (20 mM Tris–HCl, 137 mM NaCl, and 0.1% Tween 20, pH 7.5). The membrane was incubated with a primary antibody overnight at 4°C, followed by the horseradish peroxidase-conjugated secondary antibody. The primary antibodies used were Apaf-1 (Santa Cruz), cytochrome c (abcam), cleaved caspase-9 (Cell Signaling), active caspase-3 (BioVision), HIF-1α (GeneTex), and β-actin (abcam). Specific bands were detected by the ECL detection system (Millipore). Protein signals were quantified by optical density ratio using the expression of β-actin as a control.
To quantitate oxidative stress in reperfusion injury, we measured the amount of 4-HNE-modified protein, an α,β-unsaturated hydroxyalkenal produced by lipid peroxidation (24), in the partial liver IR model. Briefly, the liver samples were lysed in lysis buffer containing 50 mM Tris–HCl, 0.25% sodium deoxycholate, 150 mM NaCl, 0.1% SDS, and a protease inhibitor cocktail (Roche, Indianapolis, IN). Protein lysates were centrifuged at 4°C for 30 min at 14,000 g. After centrifugation, the supernatant fluid was collected, and protein content in each sample was determined. Equal amounts of proteins (30 μg) were loaded and separated by polyacrylamide gel electrophoresis. Specifically, 4-HNE (46545; Abcam, Cambridge, MA) (1:1000 dilution) was probed as a biomarker of oxidative stress. After exposure to horseradish peroxidase-conjugated secondary antibodies, the blots were imaged and analyzed as mentioned (22).
Serum ALT level
The serum alanine aminotransferase level in mice of each group was measured before and 60 min after reperfusion (n = 5) to ascertain the severity of hepatocellular injury. The ALT activity was determined using a prepared kit (Sigma Diagnostics, St. Louis, MO).
Histology of livers after IR injury
The paraffin-fixed liver sections of 5 μm thickness were stained for H&E. The hepatic injury was graded by Suzuki's classification, which was based on the severity of sinusoidal congestion, cytoplasmic vacuolation, and necrosis of parenchymal cells (19) shown on the liver sections. For each category, 0–3 points were graded based on the severity.
For immunohistochemical staining, the sections were probed with the anti-Apaf-1 antibody (Santa Cruz) and stained by using the UltraVision Quanto Detection System kit (Thermo Fisher Scientific, Waltham, MA). The intensity of Apaf-1 staining was quantified by a microscope-based image analysis program (Image Pro Plus; Media Cybernetics, Silver Spring, MD).
Terminal deoxynucleotidyl transferase dUTP nick end labeling
TUNEL assay was used to evaluate cell deaths after IR injury. Liver sections of 5 μm thickness were used. The labeled cells were detected using the peroxidase–streptavidin conjugate followed by diaminobenzidine staining (TdT-FragEL™ DNA Fragmentation Detection Kit, Calbiochem, San Diego, CA). Apoptotic index was presented as the percentage of TUNEL-positive cells to the totality in one 400 × magnifying field. The yields of apoptotic index in fifty selected fields in each sample were averaged.
Caspase-3 activity assay
The caspase-3 activity in Clone 9 cells was measured by the Caspase-3/CPP32 Colorimetric Assay Kit (BioVision, Palo Alto, CA). The cell lysates were incubated with DEVD-pNA (p-nitroaniline) substrate at 37°C. Cleaved caspase-3 cut the DEVD-pNA to form pNA, which was measured by detecting the absorbance at 405 nm.
Statistical analysis
Data were analyzed by Sigma Plot 8.0 (Systat Software, Inc.). Results are presented as mean ± standard error. The Mann–Whitney U test was used to analyze the differences between experimental groups. All differences were considered to be statistically significant when p value <0.05.
Footnotes
Acknowledgments
We thank Shu-Jen Chou (DNA Microarray Core Lab) and Shu-Hsing Wu (the research fellow and vice director) of Institute of Plant and Microbial biology at Academia Sinica of Taiwan for the technical assistance in performing microRNA arrays in this study. We thank Hsin-Yuan Chen for assisting the measurement of HIF-1α expression. This study was granted by the Ministry of Science and Technology of Taiwan, MOST 103-2320-B-002-005.
Authors' Contributions
H.-C. L. and I.-R.L. were responsible for the original design and primary experiments; T.-K.L. contributed to the microarray and transfection assay; S.-Y.L. performed the biochemical analysis; E.-Y.Y. performed statistical analysis; and I.-R.L. prepared the manuscript.
Author Disclosure Statement
No competing financial interests exist.