
Abstract
Aims:
We asked whether hydrogen sulfide (H2S), as the third gaseous mediator, provided fast antidepressant effect on major depressive disorders and underlying mechanisms.
Results:
The decreased level of H2S was detected in the hippocampus of chronic unpredictable mild stress (CUMS)-treated rats. Acute administration of H2S either by H2S inhalation or by the donor NaHS produced a rapid antidepressant-like behavioral effect. Further investigation demonstrated that this effect of H2S was mediated by reversing the CUMS-induced decrease in dendritic spine density and required the activation of mammalian target of rapamycin (mTOR)C1 and neurotrophic TrkB receptors, which proceeded to increase synaptic protein expression, including postsynaptic density protein 95, synaptophysin, and α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor GluR1/2 subunit.
Innovation:
This study provides the first direct evidence for detecting the decreased H2S in hippocampus of CUMS rats and the biological significance of H2S in treating major depression.
Conclusion:
Our data demonstrate that H2S activates mTORC1 signaling cascades and thereby produces fast-onset antidepressant effect. The study provides a profound insight into H2S or its donors as potent preventive and therapeutic agents for intervention of depression. Antioxid. Redox Signal. 27, 472–488.
Introduction
M
We demonstrated the decreased level of hydrogen sulfide (H2S) in hippocampus of chronic unpredictable mild stress (CUMS) rats and provide evidence that H2S donor or inhaled H2S exerted fast action to improve the depressive behaviors of CUMS rats, which involved the crucial role of H2S in facilitating mammalian target of rapamycin (mTOR)-dependent translation initiation and increasing the expression of GluR1 and GluR2 subunits of AMPA receptors. Our results indicate that H2S may become a potent preventive and therapeutic agent for depression.
Ketamine, a dissociative anesthetic, has been used as a hallucinogen for years owing to the psychotomimetic effects. Increasing evidence shows the rapid-onset and long-lasting antidepressant effect after a single subpsychotomimetic dose of ketamine (3). Ketamine is a noncompetitive blockade of N-methyl-D-aspartate receptor (NMDAR) and activates the mTOR signaling pathway and subsequently promotes synapse-associated protein synthesis, which underlies the mechanism of fast-acting antidepressant effect (25). The study on ketamine-produced antidepressant effect provides an opportunity to explore new antidepressants with rapid-acting effect, sustained relief, and less side effects.
Numerous studies have shown a link between H2S and the mTOR signaling pathway. In a recent study, it has been found that H2S could reduce smoking-induced autophagic cell death via activation of mTOR (47). A novel hydrogen sulfide-releasing molecule, GYY4137, likely protects against high-glucose-induced cytotoxicity by activation of the mTOR signaling pathway in H9c2 cells (43). Besides, H2S ameliorates hepatic ischemia and reperfusion injury through phosphorylation of the PDK-1/Akt/mTOR/p70S6k axis (39). The effect of H2S on activation of mTOR is in line with the mechanism of fast-onset antidepressant.
Increasing studies have demonstrated that there is a direct relationship between the misbalance of the redox system and the pathogenesis of several psychiatric diseases (37). Redox dysregulation occurs following an increased release of reactive oxygen species and reactive nitrogen species, while the antioxidant defense is decreased (29, 37). Clinical and preclinical studies have supported that the oxidative stress-related damage plays a major role in the etiology of depression (29). Altered levels of inflammation and immune response induced by oxidative stress have also been indicated in several types of depression-related diseases (31, 38).
On the other side, antidepressants also show antioxidant properties, for example, treatments with fluoxetine, imipramine, tricyclic antidepressants, or venlafaxine significantly increased the activities of superoxide dismutase, catalase, glutathione S-transferase, glutathione reductase and the level of glutathione (GSH) in the rodents model of depression induced by restraint stress (29). In addition, some studies indicate that antidepressant treatment combined with antioxidant drugs could enhance the antidepressive effect and produce a better clinical outcome (29).
H2S has been recognized as the toxic gas with a characteristic smell of rotten eggs for centuries. The concentration of H2S in water solution generated by NaHS is dependent on protonation equilibria, which could be calculated by the pKa of H2S and HS−. Under physiological conditions, the percentage of distribution of H2S: HS−: S2 − will be 30:70:0.000002, respectively. At physiological pH (37°C), 20–30% of sulfide is present as H2S, and H2S mainly exists as HS− (35, 36). Thus, NaHS (100 μM) solution generates H2S at 20–30 μM.
However, as the third gaseous mediator alongside nitric oxide and monoxide, H2S has been identified as an endogenous modulator of tissue function (11). Previous studies have shown that the level of H2S being released from the desulfhydration of L-cysteine was 30 μM to >100 μM. However, Furne et al. found that H2S concentrations in whole tissue were orders of magnitude lower than conventionally accepted values required to alter tissue function, which was measured by using the method of rapid tissue catabolism, and showed that H2S concentrations in situ tissue were only about 15 nM in the brain and liver of mouse (11).
Physiological concentrations of H2S selectively enhance NMDAR-mediated responses and facilitate the induction of hippocampal long-term potentiation (LTP) (1). Our previous study has also demonstrated that H2S can regulate amygdala-dependent emotional memory by promoting the function of GluN2B-containing NMDARs in the amygdala of rats (42). Pathophysiological concentration (200 pM) of NaHS, a donor of H2S, aggravates seizure-like events of rats in vivo and in vitro, which may be due to an increase in neuronal excitation (27). On the contrary, the decrease in H2S level in the brain is closely related to the occurrence of Alzheimer's disease and Parkinson's disease (10, 17).
A previous study preliminarily reported that H2S might improve depressive- and anxious-like behaviors tested in nonstressed rats and mice (4), but the effect of H2S on recognized MDD animal models as well as the potential mechanism remains unknown. To clarify the action and underlying mechanisms of H2S on the depression-like behavior, we observed the effect of intraperitoneal injection of H2S donor NaHS or inhaled H2S on the depression-like behavior in the CUMS model. The role of mTOR signaling pathway and glutamate receptor in the antidepressant effect of H2S was evaluated.
Results
Acute administration of H2S ameliorates the depressive-like behaviors of normal rats
To examine whether the levels of H2S were changed in depressive rats, the endogenously bound sulfane sulfur in hippocampus of CUMS rats was measured by gas chromatography–mass spectrometry (GC-MS). We found that the concentration of bound sulfane sulfur was significantly reduced when compared with vehicle group (vehicle: 0.0149 ± 0.0011 nM/Mg; CUMS: 0.0104 ± 0.0015 nM/Mg; F
1,16 = 6.011, p < 0.05; Fig. 1A). The standard H2S solutions were analyzed by GC-MS to verify the reliability of our results. The mass spectrum showing ions at m/z = 33, 34, and 36 (Supplementary Fig. S1A; Supplementary Data are available online at

To validate whether H2S produced toxic effect at this dosage, we examined the inflammatory cytokines such as interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and inducible nitric oxide synthase (iNOS) in the serum after intraperitoneal injection of NaHS or inhaled H2S. No significant changes were found on the level of IL-1β, TNF-α, and iNOS (Supplementary Fig. S3A–C). Next, we detected the electrocardiograph (ECG) of rats and found that neither NaHS nor inhaled H2S altered the wave of ECG in rats (Supplementary Fig. S3D). Hematoxylin and eosin (H&E) staining also showed no significant changes in cytological characteristics of hippocampus, heart, liver, spleen, lung, and kidney (Supplementary Fig. S3E). These results demonstrate that acute inhalation of H2S produces the antidepressant-like effects in rats.
Hydrogen polysulfides (H2Sn) are generated from H2S by enzyme in the brain, which produces a variety of physiological functions (21, 22). Sodium salt of polysulfide, Na2S4, was used to explore whether the effect of H2S was related to polysulfide. Na2S4 (8.7 mg/kg, i.p.) significantly reduced the immobility time of FST in normal rats at 30 min (vehicle: 30.00 ± 2.83 s; Na2S4: 14.76 ± 2.72 s, F 1,9 = 14.713, p < 0.01 vs. vehicle), 3 h (vehicle: 28.44 ± 3.29 s; Na2S4: 18.53 ± 1.67 s, F 1,9 = 6.325, p < 0.05 vs. vehicle), 24 h (vehicle: 29.46 ± 3.17 s; Na2S4: 19.83 ± 1.82 s, F 1,9 = 6.197, p < 0.05 vs. vehicle), and also lasted for 1 week (vehicle: 30.12 ± 2.81 s; Na2S4: 20.64 ± 2.21 s, F 1,9 = 6.582, p < 0.05 vs. vehicle; Fig. 1H). These results suggested that H2S-produced antidepressant effect may partially be mediated via its transformation to polysulfides.
H2S produces rapid antidepressant effect on CUMS rats
To further examine the effect of H2S on depressive-like behaviors of animal models for MDD, we used the sucrose preference test (SPT) to detect anhedonia and novelty-suppressed feeding test (NSFT) in response to anxiety, as well as FST, a behavioral despair model (25, 26). As shown in Figure 2A, CUMS is a well-validated animal model that induces depressive behaviors in rats. In this context, according to the results of SPT, NSFT, and FST, the rats can be divided into susceptible and resistant on days 32–34. The susceptible rats received the drug treatments, such as NaHS (11.2 mg/kg, i.p.), MK-801 (0.1 mg/kg, i.p.) or inhaled H2S (120 ppm, 3 h), on day 35, followed by behavioral tests successively on days 36, 37, and 38.

A single dose of NaHS reversed the decreased sucrose preference (vehicle: 88.99% ± 5.16%; CUMS: 38.13% ± 5.25%; NaHS: 80.74% ± 6.14%; two-way ANOVA, F 3,28 = 12.642, p < 0.01 vs. CUMS, Fig. 2B), the increased latency to feed (vehicle: 165.33 ± 18.02 s; CUMS: 334.63 ± 25.92 s; NaHS: 218.60 ± 24.63 s; two-way ANOVA, F 3,34 = 9.835, p < 0.01 vs. CUMS, Fig. 2C), and the immobility time in FST (vehicle: 31.63 ± 5.36 s; CUMS: 72.37 ± 7.50 s; NaHS: 40.22 ± 5.32 s; two-way ANOVA, F 3,23 = 6.902, p < 0.05 vs. CUMS, Fig. 2D), which is similar to that of MK-801 in SPT (MK-801: 77.89% ± 8.43%; two-way ANOVA, F 3,28 = 12.642, p < 0.01 vs. CUMS, Fig. 2B), NSFT (MK-801: 230.22 ± 20.00 s; two-way ANOVA, F 3,34 = 9.835, p < 0.05 vs. CUMS, Fig. 2C), and FST (MK-801: 43.72 ± 7.87 s; two-way ANOVA, F 3,23 = 6.902, p < 0.05 vs. CUMS, Fig. 2D).
Moreover, similar to that of NaHS, inhaled H2S also displayed a parallel action on SPT (76.64% ± 5.01%; two-way ANOVA, F 2,21 = 26.609, p < 0.01 vs. CUMS, Fig. 2E), NSFT (234.56 ± 20.38 s; two-way ANOVA, F 2,25 = 15.184, p < 0.05 vs. CUMS, Fig. 2F), and FST (41.57 ± 9.15 s; two-way ANOVA, F 2,17 = 7.592, p < 0.05 vs. CUMS, Fig. 2G). The above results demonstrate that H2S could restore the behavioral deficit in CUMS rats.
H2S recovered the deficit of synaptic spines in hippocampus of CUMS rats
It is well known that chronic stress may produce atrophy and loss of dendritic spines. As shown in Figure 3A, using Golgi staining, the dendritic spine density in the CA1 subfield of hippocampus of CUMS rats was measured at 24 h after NaHS injection. It was found that NaHS (11.2 mg/kg, i.p) was sufficient to restore the decreased total spine density in the CA1 region of CUMS rats (vehicle: 13.1 ± 0.43/10 μm; CUMS: 7.2 ± 0.5/10 μm; NaHS: 9.8 ± 0.4/10 μm; two-way ANOVA, F 2,27 = 43.636, p < 0.01 vs. CUMS, Fig. 3B). Meanwhile, NaHS increased both newborn thin spines (vehicle-thin: 4.6 ± 0.3/10 μm; CUMS-thin: 2.8 ± 0.2/10 μm; NaHS-thin: 4.1 ± 0.3/10 μm, two-way ANOVA, F 2,27 = 9.361, p < 0.05 vs. CUMS) and mature mushroom-like spines (vehicle-mushroom: 8.5 ± 0.3/10 μm; CUMS-mushroom: 4.4 ± 0.4/10 μm; NaHS-mushroom: 5.7 ± 0.3/10 μm, two-way ANOVA, F 2,27 = 43.900, p < 0.05 vs. CUMS), an indication of increased spine maturation and synaptic strengthening (Fig. 3C). Taken together, the results suggest that NaHS rescues the decreased dendritic spine density induced by CUMS, raising the possibility that NaHS may be beneficial in depression treatment.

Activation of mTORC1 signaling contributes to the antidepressant effect of H2S
Next, we investigated whether the antidepressant effect of H2S was related to the activation of mTORC1, a signaling cascade implicated in fast onset of antidepressant action. p70S6 kinase (p70S6k) is one of the major downstream targets of mTORC1, and activation of mTORC1 leads to phosphorylation of P70S6K (p-P70S6k). We found that NaHS (11.2 mg/kg, i.p.) significantly increased the ratio of phosphorylated P70S6k to total P70S6k (p-P70S6k/P70S6k) from 0.7350 ± 0.0485 to 1.0189 ± 0.0942 in the hippocampus of CUMS rats (two-way ANOVA, F 3, 24 = 3.133, p < 0.05 vs. CUMS; Fig. 4A and Supplementary Fig. S2). We also observed that MK-801 (0.1 mg/kg, i.p.) prevented the decreased level of p-P70S6k/P70S6k induced by CUMS from 0.7350 ± 0.0485 to 1.0001 ± 0.0768 (two-way ANOVA, F 3,24 = 3.133, p < 0.05 vs. CUMS; Fig. 4A).
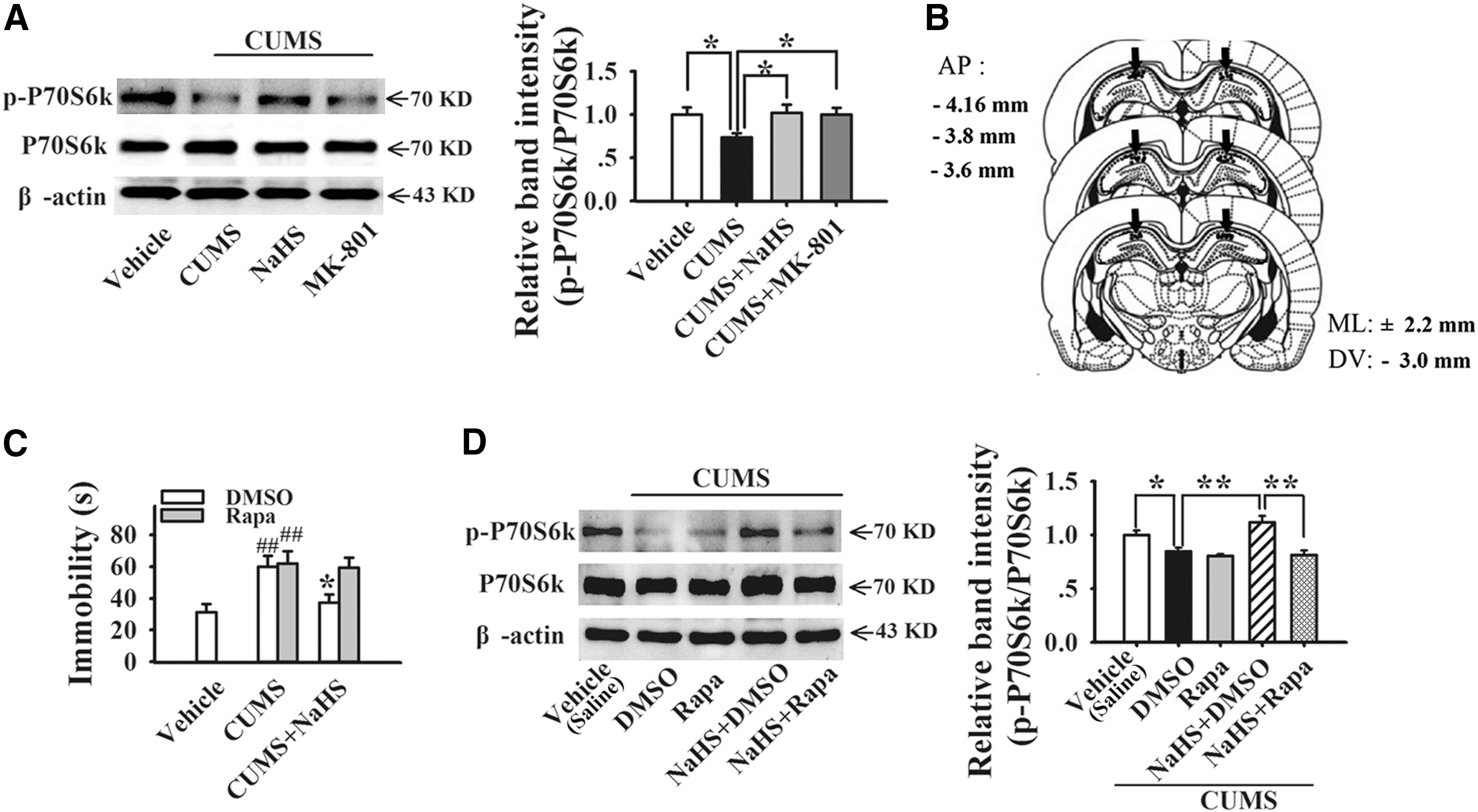
To validate the relationship between mTORC1 activation and NaHS-induced antidepressant responses, a selective mTORC1 inhibitor rapamycin (Rapa) was given by intrahippocampal infusion (Fig. 4B). We found that rapamycin completely blocked NaHS-induced antidepressant-like effect in FST (NaHS: 37.35 ± 5.31 s, Rapa+NaHS: 59.50 ± 6.32 s, two-way ANOVA, F 4,30 = 4.909, p < 0.05 vs. NaHS, Fig. 4C) as well as the elevated level of p-P70S6k/P70S6k (NaHS: 1.1171 ± 0.0621, Rapa+NaHS: 0.8139 ± 0.0395, two-way ANOVA, F 4,25 = 10.776, p < 0.01 vs. NaHS, Fig. 4D and Supplementary S2), indicating that the effect of NaHS is mTORC1 dependent.
Inhibition of mTORC1 signaling in the hippocampus antagonizes the antidepressant effect of H2S
Furthermore, we found that NaHS increased the level of regulatory-associated protein of mTOR (raptor) protein in the hippocampus of CUMS rats (CUMS: 0.7292 ± 0.0519, NaHS: 0.9240 ± 0.0706, two-way ANOVA, F 4, 20 = 4,416, p < 0.05 vs. CUMS, Fig. 5A and Supplementary Fig. S5), which was not abolished by rapamycin (NaHS+rapa: 0.8848 ± 0.1056, Fig. 5A and Supplementary Fig. S2), indicating that rapamycin-inhibited NaHS-induced mTORC1 activation may be related to the dissociation of raptor from mTOR, but not decreased raptor expression. However, NaHS had no effect on the level of rapamycin-insensitive companion of mTOR (rictor) protein (Fig. 5B and Supplementary Fig. S2).

Given that NaHS activated the mTORC1 signaling pathway, we constructed a lentivirus short hairpin RNA (shRNA) of raptor. Intrahippocampal injection of raptor shRNA into the CA1 subregion for 15 days (Supplementary Fig. S4A) inhibited NaHS-induced increase in levels of both raptor protein (control shRNA+NaHS: 1.0002 ± 0.0560, raptor shRNA+NaHS: 0.6186 ± 0.0399, two-way ANOVA, F 4, 25 = 23.260, p < 0.01 vs. control shRNA+NaHS, Fig. 5C and Supplementary Fig. S2) and p-P70S6k/P70S6k protein (control shRNA+NaHS: 1.0370 ± 0.1070, raptor shRNA+NaHS: 0.7219 ± 0.0758, two-way ANOVA, F 4, 35 = 4,585, p < 0.05 vs. control shRNA+NaHS, Fig. 5D and Supplementary Fig. S2) in the hippocampus of CUMS rats.
We also found that raptor shRNA reversed the antidepressant behavioral responses of NaHS by reducing sucrose preference (control shRNA+NaHS: 94.84% ± 1.77%, raptor shRNA+NaHS: 69.72% ± 9.28%, two-way ANOVA, F 4, 29 = 12.329, p < 0.01 vs. control shRNA+NaHS, Fig. 5E), increasing the latency to feed (control shRNA+NaHS: 236.00 ± 17.33 s, raptor shRNA+NaHS: 359.33 ± 43.01 s, two-way ANOVA, F 4,29 = 6.057, p < 0.05 vs. control shRNA+NaHS, Fig. 5F), and prolonging the immobility time (control shRNA+NaHS: 38.86 ± 3.81 s, raptor shRNA+NaHS: 63.43 ± 10.57 s, two-way ANOVA, F 4, 30 = 5.692, p < 0.05 vs. control shRNA+NaHS, Fig. 5G). Thus, our findings suggest that NaHS induces antidepressant effects through activating mTORC1 signaling cascades.
H2S rapidly rescues the deficit of synaptic protein expression caused by CUMS
Considering that mTOR activation can regulate rapid translation of some critical proteins for synaptic function, we then examined whether H2S upregulated some key synaptic proteins during CUMS, including postsynaptic density protein 95 (PSD95) and presynaptic protein synaptophysin. In this study, we found that NaHS obviously increased the fluorescence of PSD95 and synaptophysin in CA1, CA3, and DG regions of CUMS rats (Fig. 6A). Raptor shRNA completely abolished the increased expression of PSD95 protein (control shRNA+NaHS: 1.086 ± 0.079, raptor shRNA+NaHS: 0.683 ± 0.059, two-way ANOVA, F 4, 30 = 13.084, p < 0.01 vs. control shRNA+NaHS, Fig. 6B and Supplementary Fig. S2) and synaptophysin (control shRNA+NaHS: 0.946 ± 0.050, raptor shRNA+NaHS: 0.773 ± 0.022, two-way ANOVA, F 4, 20 = 7.497, p < 0.05 vs. control shRNA+NaHS, Fig. 6C and Supplementary Fig. S2) induced by NaHS, suggesting that H2S increases the expression of synaptic protein via activation of mTORC1 signaling.

H2S increases neurotrophic signaling TrkB expression in CUMS rats
Previous studies have shown that the level of TrkB protein is significantly reduced in the hippocampus of suicidal subjects and overexpression of TrkB reduces anxiety and depressive behavior in mice (8, 23, 24). To investigate the impact of H2S on TrkB expression, the level of TrkB was observed in the hippocampus of CUMS rats. As shown in Figure 7A and Supplementary Figure S2, NaHS recovered the decreased expression of TrkB protein (vehicle: 1.000 ± 0.0416, CUMS: 0.7489 ± 0.0478, NaHS: 0.9634 ± 0.0472, two-way ANOVA, F 2,12 = 6.988, p < 0.01 vs. vehicle, p < 0.05 vs. CUMS), indicating that the antidepressant effect of NaHS may be related to the upregulation of TrkB.

Considering that the activity of mTOR signaling plays an important role in synaptic protein translation, we then examined whether the increase in TrkB expression was involved in mTORC1 activation. We found that raptor shRNA canceled the upregulation of TrkB protein induced by NaHS (control shRNA+NaHS: 0.9541 ± 0.0399, raptor shRNA+NaHS: 0.7504 ± 0.0360, two-way ANOVA, F 4, 25 = 4.065, p < 0.05 vs. control shRNA+NaHS, Fig. 7B and Supplementary Fig. S2), indicating that H2S-elevated TrkB expression is dependent on mTORC1 activation.
To further assess the role of TrkB in the antidepressant effect of NaHS, we conducted SPT, NSFT, and FST after knockdown of TrkB in the hippocampus by shRNA. A similar result was found that TrkB shRNA prevented the increased sucrose preference in SPT (control shRNA+NaHS: 92.0% ± 2.85%, TrkB shRNA+NaHS: 66.0% ± 10.5%, two-way ANOVA, F 4, 33 = 9.239, p < 0.05 vs. control shRNA+NaHS, Fig. 7C), the decreased latency to feed in NSFT (control shRNA+NaHS: 236.0 ± 17.3 s, TrkB shRNA+NaHS: 358.5 ± 30.3 s, two-way ANOVA, F 4, 31 = 10.453, p < 0.01 vs. control shRNA+NaHS, Fig. 7D), and the reduced immobility in FST (control shRNA+NaHS: 38.9 ± 3.8 s, TrkB shRNA+NaHS: 69.3 ± 8.2 s, two-way ANOVA, F 4, 30 = 7.338, p < 0.01 vs. control shRNA+NaHS, Fig. 7E) induced by NaHS in CUMS rats. These results indicate that NaHS-produced antidepressant effect is also dependent on neurotrophic signaling pathways.
Activation of α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor contributes to the antidepressant effect of H2S
Previous reports have demonstrated that the rapid antidepressant actions of ketamine are dependent on glutamate transmission. We then examined whether NaHS was involved in the expression of α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) receptor by activating mTORC1 signaling cascades. The results showed that NaHS restored the deficit of GluR1 (CUMS: 0.7723 ± 0.0552, NaHS: 0.9851 ± 0.0579, two-way ANOVA, F 2, 12 = 4.917, p < 0.05 vs. CUMS, Fig. 8A and Supplementary Fig. S2) and GluR2 expression (CUMS: 0.7649 ± 0.0631, NaHS: 1.0285 ± 0.0478, two-way ANOVA, F 2, 15 = 6.589, p < 0.01 vs. CUMS, Fig. 8B and Supplementary Fig. S2) in the hippocampus induced by CUMS. This result was consistent with the findings regarding mTORC1 activation-dependent synthesis of synaptic protein.

Moreover, we also found that the level of phosphorylated GluR1 (p-GluR1) was elevated by NaHS (CUMS: 0.7134 ± 0.0562, NaHS: 0.9091 ± 0.0595, two-way ANOVA, F 4, 30 = 4.533, p < 0.05 vs. CUMS, Fig. 8D and Supplementary Fig. S2). Raptor shRNA prevented the NaHS-induced increase in the expression of GluR1 (control shRNA+NaHS: 1.0176 ± 0.1059, raptor shRNA+NaHS: 0.7105 ± 0.0751, two-way ANOVA, F 4, 30 = 3.548, p < 0.05 vs. control shRNA+NaHS, Fig. 8C and Supplementary Fig. S2) and GluR2 (control shRNA+NaHS: 1.0776 ± 0.0582, raptor shRNA+NaHS: 0.8263 ± 0.0619, two-way ANOVA, F 4, 25 = 4.392, p < 0.01 vs. control shRNA+NaHS, Fig. 8E and Supplementary Fig. S2), but had no effect on p-GluR1 level (Fig. 8D and Supplementary Fig. S2).
Additionally, TrkB shRNA prevented the NaHS-induced increase in the level of GluR1 (control shRNA+NaHS: 0.9374 ± 0.0829, TrkB shRNA+NaHS: 0.4518 ± 0.0659, two-way ANOVA, F 4, 25 = 12.496, p < 0.01 vs. control shRNA+NaHS, Fig. 8F and Supplementary Fig. S2) and p-GluR1 (control shRNA+NaHS: 0.9365 ± 0.0880, TrkB shRNA+NaHS: 0.3504 ± 0.1350, two-way ANOVA, F 4, 20 = 7.615, p < 0.01 vs. control shRNA+NaHS, Fig. 8G and Supplementary Fig. S2), but not GluR2 (Fig. 8H and Supplementary Fig. S2).
Discussion
The present study demonstrated that deficiency of endogenous H2S in the hippocampus contributed to abnormal behaviors induced by CUMS, while increasing hippocampal H2S levels by administration of NaHS or inhalation of H2S can reverse the depressive-like behaviors in rats within few hours, indicating that H2S can act as a fast-onset antidepressant. We also found that H2S could counteract the loss of dendrite spine in hippocampus induced by CUMS. As shown in Supplementary Figure S5A, H2S increased mTORC1 cascades and sequentially accelerated the translation and synthesis of synaptic proteins such as PSD95 and then upregulated TrkB expression and increased the activity of GluR1 and GluR2 subunits of AMPA receptors, thus contributing to the treatment of depressive-like behaviors.
In this study, we found that a single treatment with intraperitoneal injection of NaHS produces fast-acting antidepressant responses, which were similar to that of NMDAR antagonist MK-801, while conventional antidepressant fluoxetine showed delayed onset of action. Notably, the behavioral effect of NaHS lasted for 1 week, but MK-801 only lasted for 24 h in our system, indicating the long-term effect of NaHS. The fast-acting antidepressant effect is very necessary, especially for suicide-risk patients. Moreover, the immobility time of rats significantly decreased in a concentration-dependent manner evoked by inhalation of 120 and 160 ppm H2S for 3 h. A recent study has reported that intraperitoneal injection of 1.68 mg·kg−1 and 5.6 mg·kg−1 NaHS once daily for seven consecutive days improved the depressive -like behaviors in animal model (4).
Our present study differs from previous report with three characteristics. First, the previous observation was obtained from naive animals using FST or TST, which are only assessments for depressive-like behaviors, but not recognized as animal models that can mimic the symptoms more closely to MDD patients. We examined the effects of H2S on a well-recognized animal model, CUMS, and measured them with assessments such as SPT, NSFT, and FST to detect anhedonia, anxiety, and despair, respectively. Second, our results showed that a single dose of NaHS at 11.2 mg kg−1 produced a fast-onset antidepressant effect, suggesting that a higher dose of NaHS is required for the fast-onset antidepressant effect. Third, we found that H2S applied through gas inhalation also produced rapid antidepressant-like effects in CUMS rats.
The investigation from animal and clinical experiments indicates that NMDAR antagonists, such as ketamine and MK-801, receive much attention on the activation of mTOR signaling that can produce their antidepressant effects within hours and last for a few days (26). The mTOR complex is ubiquitously expressed in the brain, which locates at the crossroads of neuronal activity, growth factors, energy, and nutrient levels to regulate protein translation and synaptic plasticity, as well as other cell functions (14). In our study, H2S activated mTORC1 cascades and then increased the expression of some synaptic proteins that are critical to synaptic transmission. This is very similar to the antidepressant mechanisms of ketamine. Meanwhile, we also found that mTORC2 cascades were not required for the antidepressant action of H2S.
Notably, several studies have demonstrated that H2S selectively enhances NMDAR-mediated responses and facilitates hippocampal LTP (1). Our results showed that exogenous H2S produced the fast-onset antidepressant effect, which was similar to that of NMDAR antagonist ketamine. It seems not to be consistent since both NMDAR antagonist and agonist have potential to treat major depression (18). This inconsistency may be explained as follows. First, at the cellular level, both H2S and ketamine can activate mTOR signaling pathways. The activity of mTOR is regulated by a number of upstream mechanisms, which might represent a common mechanism to other putative rapid antidepressants (5, 13). Considering that the upstream of mTOR signaling is complicated, we speculated that H2S increased mTORC1 signaling cascades through some other pathways distinguished with NMDAR blockade.
Second, perturbations in the balance between synaptic and extrasynaptic NMDAR activity contribute to neuronal dysfunction in depression and other neurodegenerative diseases. A vigorous review reveals that NMDAR-induced responses depend on the receptor location: stimulation of synaptic NMDARs leads to the buildup of a neuroprotective shield, whereas stimulation of extrasynaptic NMDARs promotes cell death (12). It is likely that ketamine preferentially reduces the efficacy of excessive extrasynaptic NMDAR activity. However, H2S may preferentially improve the deficit of synaptic NMDAR activity.
Additionally, NMDARs are widely distributed in limbic–cortical circuits and adaptive functional interactions of the highly integrated network, which are responsible for homeostatic control of emotion. The neural substrates underlying depression are complex and the dysfunction of corticolimbic network is involved. It is theoretically plausible that H2S-mediated increase in NMDAR activity may be related to neuronal projections originated from the hippocampus and multiple circuits that produce the antidepressant-like behavior different from that of NMDAR antagonist.
The antidepressant actions of ketamine have been reported to require glutamate-induced activation of AMPAR, and pretreatment with a selective AMPAR inhibitor NBQX completely blocked the antidepressant effect (3). Our previous study has reported that H2S facilitated the function of AMPAR and NMDAR in entorhinal cortex neurons by using electrophysiological recording. NaHS significantly increased the amplitude of AMPA-mediated eEPSC and NMDA-mediated eEPSC, suggesting that NaHS potentiated the postsynaptic functions via AMPAR and NMDAR (27).
In line with this, we found that treatment with H2S significantly increased the expression of GluR1 and GluR2 subunits of AMPAR, indicating that the antidepressant responses of H2S are also mediated through activation of AMPAR. Therefore, the antidepressant mechanisms of H2S and ketamine may converge on activation of mTORC1 and AMPAR, suggesting a relationship between NMDAR enhancement and inhibition underlying the antidepressant actions. Further investigation is needed to compare other behavioral effects as well as benefit and risk profiles between NMDAR agonists and antagonists.
BDNF is linked to the traditional antidepressant action, and BDNF deletion in the hippocampus attenuates antidepressant behavioral responses (2, 32). In our study, the results showed that H2S reversed the reduction of TrkB receptors induced by CUMS, indicating the critical role of neurotrophic signaling in the antidepressant actions of H2S. These findings are consistent with the results that H2S produced neuroprotection against formaldehyde-induced toxicity in PC12 cells mediated by BDNF-TrkB pathways (19), proving that synaptogenesis induced by activation of glutamate receptor requires the release of BDNF to stimulate TrkB-mTOR signaling and synaptic protein synthesis.
Higher cardiovascular morbidity and mortality in a depressive patient have been demonstrated repeatedly (15, 34). It has been accepted that H2S can relax vascular smooth muscle by opening KATP channels to exert cardioprotective effects (46). Combined with our present results, H2S may have greater therapeutic advantage on the prevalence of depression comorbid with cardiovascular diseases. In addition, exogenous H2S reduces blood pressure by improving vasorelaxation. Hypertension in mice with deletion of cystathionine γ-lyase has been observed (45). However, as a small gas molecule, H2S can diffuse rapidly across cell membranes and blood–brain barrier. In our present study, whether the peripheral effect of H2S contributed to the antidepressant responses was still not excluded and the related investigation is still needed.
Together, the present study demonstrated that the acute application of H2S, through either H2S donor NaHS or H2S gas inhalation, produced a robust antidepressant effect, which was predominantly mediated through activation of the mTORC1 signaling pathway, and then increased the synthesis of synaptic proteins, including PSD95 and synaptophysin. Moreover, H2S upregulated the level of TrkB receptors and further increased the activity of GluR1 and GluR2 subunits of AMPARs. A better understanding of the roles of H2S can provide insights into potential therapeutic interventions against depression. More specifically, available data strongly suggest that H2S may become a potent preventive and therapeutic agent for depression with fast onset, and this should be addressed in future studies.
Materials and Methods
Animals
Male Sprague-Dawley rats (Animal Center of Tongji Medical College, Wuhan, China) weighing 200–250 g were individually housed and maintained in standard conditions with a 12-h dark–12-h light cycle and ad libitum access to food and water. Animals were allowed to acclimate for 4–5 days before the experiments. Animal care and use were in accordance with the guidelines approved by the Animal Welfare Committee of Huazhong University of Science and Technology.
Drugs
NaHS (11.2 mg/kg [42]; Sigma, St. Louis, MO), MK-801 (0.1 mg/kg [3]; Sigma, St. Louis, MO), fluoxetine (10 mg/kg [7]; Sigma, St. Louis, MO), and Na2S4 (8.7 mg/kg, Huaxia Chemical Reagent Ltd., Chendu, China) were dissolved in 0.9% saline and administered intraperitoneally (i.p.) in a volume of 1 ml/kg of body weight. Rapamycin (0.015 nmol/0.5 μl/side [41]; Sigma, St. Louis, MO) was dissolved in 2 μl DMSO and infused into CA1 area (16, 40). The final concentration of DMSO was less than 0.5%.
For H2S inhalation, rats were exposed to air containing 120 ppm of H2S (New radar Gas Co. Ltd., Wuhan, China) in a custom-made chamber (d 100 × w 100 × h 60 cm, plexiglas) for 3 h. The concentration of H2S was chosen based on dose-ranging studies and was continuously measured using a portable gas monitor (T40 H2S-Gas Monitor, detection limit 1 ppm, Ao Bolong Electronics and Technology Co. Ltd., Jinan, China).
CUMS model
Rats were subjected to a variable sequence of mild and unpredictable stressors for 35 days. This procedure produced depressive-like behavioral phenotype in rats. Rats were exposed to 2 stressors per day and received a total of 11 different stressors, including lights off for 3 h during the day, crowded housing, 45° tilted cages, constraint, food and water deprivation, wet bedding overnight or during the day, strobe light overnight, forced swimming, lights on overnight, placement in a 4°C ambient temperature, and electric footshock (Table 1) (3).
The stressor type, duration of stress exposure, and days of administration for the CUMS exposure period are listed here.
Behavioral assessments
All behavioral tests were conducted during the dark circadian period (19:00–23:00).
Sucrose preference test
Rats were given the choice to drink from two bottles in individual cages, one with 1% palatable sucrose solution and the other with water. All rats were habituated to the drinking for consecutive 2 days before testing, and the position of the bottles was changed every 12 h to prevent possible effects of side preference in drinking behavior. Followed by 24 h of food and water deprivation, rats were then exposed to premeasured bottles for 1 h with their position interchanged during the test. Sucrose preference was defined as the ratio of the volume of sucrose versus total volume of sucrose and water consumed during the 1-h test (26).
Novelty-suppressed feeding test
Rats were food-deprived for 24 h and placed in an open field (d 70 × w 70 × h 40 cm, Plexiglas) with a small amount of food in the center. Animals were allowed to explore the open field for 8 min. The sessions were filmed by a camcorder from above. The latency to feed was the time taken for the animal to approach and take its first bite of the food. Specifically, it was recorded offline by a stopwatch in seconds (26).
Forced swimming test
Rats were individually placed in a glass cylinder (20 cm in diameter, 40 cm in height) filled with 30 cm high water (25°C ± 1°C). The depth of water was set to prevent rats from jumping out and touching the bottom with their hind limbs and the water was exchanged after each trial. Rats were forced to swim for 6 min, and the immobility time during the final 4-min interval of the test was recorded with a video camera from the side. Immobility was defined as motionless floating in the water or making only those movements necessary to keep its head above the water. The immobility time was counted offline by a stopwatch in seconds in a single-blind study (25).
Tissue preparation and Western blot analysis
The hippocampus was prepared for the assay of protein expression. Rats were decapitated and the brain was quickly removed. Coronal brain slices (400 μm thick) containing the hippocampus were cut using a vibratome (VT 1000S; Leica, Wetzlar, Germany), and CA1 was dissected thoroughly under the microscope using a special blade. The tissues were homogenized separately on ice in extraction buffer containing 50 mM Tris-base (pH 7.4), 100 mM NaCl, 1% NP-40, 10 mM EDTA, 20 mM NaF, 1 mM PMSF, 3 mM Na3VO4, and protease inhibitors. The homogenates were centrifuged at 12,000 g for 15 min at 4°C. The supernatant was separated and protein concentration was estimated by Coomassie brilliant blue protein assay (Nanjing Jiancheng Institute of Biological Engineering, Nanjing, China).
Then, the samples were mixed with sodium dodecyl sulfate (SDS) sample buffer, boiled at 95°C for 5 min, and stored at −80°C until electrophoresis. Samples (40 μg) were separated through 10% SDS-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Schleicher and Schuell, Keene, NH). After blocking with 5% bovine serum albumin (BSA) in tris-buffered saline containing 0.1% Tween-20 (TBST) for 1 h at room temperature, transferred membranes were incubated overnight at 4°C with appropriate primary antibodies against β-actin (1:3000; Santa Cruz, CA) and synaptophysin (1:000; Abcam, CAMB, MA). p-P70S6k (1:500), P70S6k (1:500), raptor (1:1000), rictor (1:1000), p-GluR1 (1:1000), GluR1 (1:500), PSD95 (1:1000), and TrkB (1:1000) were all purchased from Cell signaling (Cell signaling, MA).
Following three washes with TBST buffer, membranes were visualized with horseradish peroxidase-conjugated goat anti-rabbit or goat anti-mouse secondary antibodies (1:3000; Santa Cruz, CA) in TBST with 1% BSA for 1 h at room temperature. Using enhanced chemiluminescence (Pierce, Rockford, IL), the bands were quantified using ImageJ software (National Institutes of Health [NIH], Bethesda, MD). The results were normalized to the quantity of β-actin and the p-P70S6k was normalized to the quantity of total P70S6k in each lane. All assays were performed at least three times.
Measurement of bound sulfane sulfur content
Bound sulfane sulfur content in hippocampal tissue was measured using a GC-MS approach. Briefly, hippocampal tissues were homogenized in phosphate-buffered saline (PBS, pH 7.4, 5 μl/mg tissues). A volume of 0.1 ml of the resulting homogenate was then pipetted and mixed with identical volume of 15 mM dithiothreitol in 0.1 M Tris-HCl buffer (pH 9.0). Following complete incubation at 37°C for 50 min under a sealed condition, 0.4 ml of 1 M sodium citrate buffer (pH 6.0) was added and an additional incubation at 37°C for 10 min under a sealed condition was performed. Thus, the generated 100 μl headspace gas was subjected to GC-MS analysis using Agilent 6890N-5975B GC-MS system (Agilent Technologies, Santa Clara, CA).
Analytes were separated with an HP-5 MS column (Agilent Technologies). The column temperature program was as follows: initially held at 40°C for 3 min, elevated to 160°C at a rate of 10°C/min, and then maintained at 160°C for 2 min. All direct injections were performed in the splitless mode (1 min). The injector temperature was held at 160°C. The carrier gas was helium at a flow rate of 1.0 ml/min. When operated in the selected ion monitoring mode, mass of 33 U was monitored and ionization was carried out in the electron ionization mode. The transfer line temperature was maintained at 280°C and the ion source temperature was 230°C. The concentrations of bound sulfane sulfur in the samples were calculated using a standard curve prepared with standard NaHS solutions.
Golgi silver staining
Rats were perfused and the brains were processed using modified Golgi staining. All rats were deeply anesthetized with pentobarbital sodium (60 mg/kg, i.p.) and perfused transcardially with 0.9% saline, followed by 4% paraformaldehyde in 0.1 M phosphate buffer, and then with the mordant containing 5% chloral hydrate, 5% potassium, and 10% formalin in ddH2O. The brains were removed and stained by postfixing in the same mordant solution for 3 days in a dark room. Thereafter, the brains were immersed in 1% silver nitrate in ddH2O for 3 days under the conditions of light protection. The interested regions in coronal sections (50 mm thickness) were obtained using a vibratome (VT 1000S; Leica, Wetzlar, Germany). Sections were then treated with 2% potassium dichromate in ddH2O for 10 min, followed by dehydrate in graded alcohols from 70% to 100%. Finally, the sections were rinsed with dimethylbenzene for 10 min and mounted with resinous medium and air-dried overnight.
Golgi-impregnated pyramidal neurons in the CA1 subfield of hippocampus were studied. Neuron that was relatively isolated from neighboring impregnated cells was allowed for identification. For each selected neuron, the size and shape were similar and the criteria were employed to analyze neurons within and between groups. The spine density (mushroom or thin) was obtained from the secondary dendrite on the basal tree by the use of an image analysis program (Image-Pro Plus).
Surgery for intrahippocampal injection
Rats were anesthetized with pentobarbital sodium (60 mg/kg, i.p.) and placed in a stereotaxic frame. Under aseptic conditions, double-guide cannulas (C235; Plastics One, Roanoke, VA) with inserted dummy cannula and dust cap were implanted into the skull using a stereotactic holder and fixed with dental cement. Bilateral cannulas were delivered into the hippocampal CA1 region according to bregma: anteroposterior, −3.6 mm; mediolateral, ±2.2 mm; and dorsoventral, −2.9 mm. The animals were allowed to recover for 7 days before the experiments were started. During injection, the bilateral stainless steel cannulas (0.28 mm o.d.) were connected to a 0.5-μl microsyringe (Hamilton, Fisher Labosi, France). To limit reflux along the injection track, the electrode was maintained in place for 10 min. Drug injections (for DMSO and rapamycin) were performed at a constant rate of 0.1 μl/min (final volume, 1 μl/side).
Lentivirus and shRNA expression
A third generation of self-inactivating lentivirus vector was purchased from Shanghai Genechem Co., Ltd. (Shanghai, China), which contains a CMV-driven enhanced green fluorescent protein (GFP) reporter and a U6 promoter upstream of cloning restriction sites (HpaI and XhoI) to allow introduction of oligonucleotides encoding shRNAs. Raptor shRNA construct and TrkB shRNA construct targeted to rat Raptor and TrkB mRNA sequence, respectively, and were subcloned into the vectors. The sequence for Raptor shRNA was 5′-GGAACACTCGAGTCAGGAA-3′, for TrkB shRNA was 5′-GCTTAAAGTTTGTGGCTTA-3′, and for control scrambled shRNA was 5′-UUCUCCGAACGUGUCACGU-3′.
Effective sequences of a Raptor shRNA, a TrkB shRNA, and a control shRNA were prepared and constructed with lentivirus-expressing GFP, respectively. GFPs together with shRNAs were stereotactically and bilaterally injected into the hippocampal CA1 regions of rats. Bilateral injection of lentiviral constructs into the CA1 was carried out using a stereotaxic instrument; 0.5 μl of lentivirus suspension containing 1 × 109 TU/ml was injected in each hemisphere at a rate of 0.05 μl/min. The needle was withdrawn over a course of 10 min. Rats were subjected to experiments on 14 days after injection.
Immunofluorescence
The rats were deeply anesthetized by pentobarbital sodium (60 mg/kg, i.p.) and sacrificed by intracardial perfusion with 0.9% saline, followed by ice-cold 4% paraformaldehyde in PBS. Brains were removed and postfixed overnight in the same 4% paraformaldehyde solution and then incubated with a 30% sucrose solution in PBS at 4°C for 3 days. Coronal brain slices (30 μm thick) containing the hippocampus were cut using a vibratome (VT 1000S; Leica, Wetzlar, Germany). Slices were immersed in PBS and stored at 4°C for further use.
Following incubation in a blocking buffer that contained 3% BSA and 0.3% Triton X-100 for 1 h, the sections were incubated with primary antibodies at 4°C overnight. The antibodies used were rabbit anti-GFP (1:300; Abcam, CAMB, MA), rabbit anti-PSD95 (1:100; Cell signaling, MA), and mouse anti-synaptophysin (1:100; Abcam, CAMB, MA). The secondary antibodies, including Alexa Fluor 488 goat anti-rabbit IgG and Alexa Fluor 594 goat anti-mouse IgG (1:200; Invitrogen, CA), were applied for 1 h at room temperature. Fluorescent image acquisition was performed with an Olympus Fluoview FV1000 confocal microscope using 10 × and 40 × objective lens. Images were analyzed using Image-Pro Plus.
Electrocardiograph
Fifteen rats were randomly divided into three groups: saline, NaHS, and H2S groups. After anesthesia with pentobarbital sodium (3%, 0.2 ml/100 g), ECG test equipment for laboratory use (BL-420E, Chengdu, China) was employed to confirm the standard in II-limb lead AVL. After recording the normal ECG, rats were injected intraperitoneally with saline (2 ml/kg), NaHS (11.2 mg/kg), and inhaled H2S (120 ppm, 3 h). One hour later, ECG was tested again.
H&E stain
After ECG measurement, rats were anesthetized with pentobarbital sodium (3%, 0.2 ml/100 g) and perfused with 100 ml saline, followed by phosphate buffer (0.1 M, pH 7.4) of 4% paraformaldehyde. Then, individual lobes of tissue biopsy material were placed in processing cassettes, dehydrated through a serial alcohol gradient, and embedded in paraffin wax blocks. Before immunostaining, 5-μm-thick tissue sections were dewaxed in xylene, rehydrated through decreasing concentrations of ethanol, and washed in PBS. The sections were selected at the same layers from each tissue, and then stained with H&E. After staining, sections were dehydrated through increasing concentrations of ethanol and xylene.
Enzyme-linked immunosorbent assay
Rats were decapitated and blood was taken with the filtrated tube of heparin. After centrifugation (3000 g, 20 min), the supernatant was retained and saved at −80°C. The enzyme-linked immunosorbent assay (ELISA) kits used to detect TNF-α, IL-1β, and iNOS were employed according to the instructions of kits (Guangrui Biological Technology Corporation, Shanghai, China).
Statistical analyses
All analyses were performed using SPSS 18.0 software (SPSS, Inc.) and data are presented as mean ± standard error of mean. Differences between mean values were evaluated using one-way ANOVA or two-way ANOVA, as appropriate. For one-way ANOVAs, post hoc tests were performed using least significant difference test. For two-way ANOVAs, Bonferroni post hoc tests were used to assess isolated comparisons. p < 0.05 was considered statistically significant.
Footnotes
Acknowledgments
This work was supported by grants from the National Basic Research Program of China (No. 2014CB744601 to F.W.; No. 2013CB531303 to J.G.C), the National Natural Scientific Foundation of China (No. 81471377 to F.W.; No. 81473198 to J.G.C), the Innovation Group of Natural Science Fund of Hubei Province (No. 2015CFA020), PCSIRT (No. IRT13016), and the Fundamental Research Funds for the Central Universities, HUST (No. 2015ZDTD045), to J.G.C.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.