
Abstract
Significance:
“Nitroproteomic” is under active development, as 3-nitrotyrosine in proteins constitutes a footprint left by the reactions of nitric oxide-derived oxidants that are usually associated to oxidative stress conditions. Moreover, protein tyrosine nitration can cause structural and functional changes, which may be of pathophysiological relevance for human disease conditions. Biological protein tyrosine nitration is a free radical process involving the intermediacy of tyrosyl radicals; in spite of being a nonenzymatic process, nitration is selectively directed toward a limited subset of tyrosine residues. Precise identification and quantitation of 3-nitrotyrosine in proteins has represented a “tour de force” for researchers.
Recent Advances:
A small number of proteins are preferential targets of nitration (usually less than 100 proteins per proteome), contrasting with the large number of proteins modified by other post-translational modifications such as phosphorylation, acetylation, and, notably, S-nitrosation. Proteomic approaches have revealed key features of tyrosine nitration both in vivo and in vitro, including selectivity, site specificity, and effects in protein structure and function.
Critical Issues:
Identification of 3-nitrotyrosine-containing proteins and mapping nitrated residues is challenging, due to low abundance of this oxidative modification in biological samples and its unfriendly behavior in mass spectrometry (MS)-based technologies, that is, MALDI, electrospray ionization, and collision-induced dissociation.
Future Directions:
The use of (i) classical two-dimensional electrophoresis with immunochemical detection of nitrated proteins followed by protein ID by regular MS/MS in combination with (ii) immuno-enrichment of tyrosine-nitrated peptides and (iii) identification of nitrated peptides by a MIDAS™ experiment is arising as a potent methodology to unambiguously map and quantitate tyrosine-nitrated proteins in vivo. Antioxid. Redox Signal. 26, 313–328.
Introduction
I
One of the molecular footprints left by the reactions of reactive nitrogen species with biomolecules is the nitration (i.e., substitution of a hydrogen atom by a nitro group, -NO2) of protein tyrosine residues in the ortho position to the phenolic hydroxyl group to form 3-nitrotyrosine (Fig. 1). Target proteins can be nitrated by different biochemical pathways that result in the modification of specific tyrosine residues (4). Overall, biological protein tyrosine nitration is an oxidative post-translational modification (PTM) that involves a free radical process with the intermediacy of tyrosyl radicals (82); notably, it has been shown that in spite of being a nonenzymatic process, it is selectively directed under biologically relevant conditions toward a limited subset of tyrosine residues [i.e., typically only one or two tyrosine residues are targets in a nitrated protein under biologically relevant conditions, (84) and vide infra].
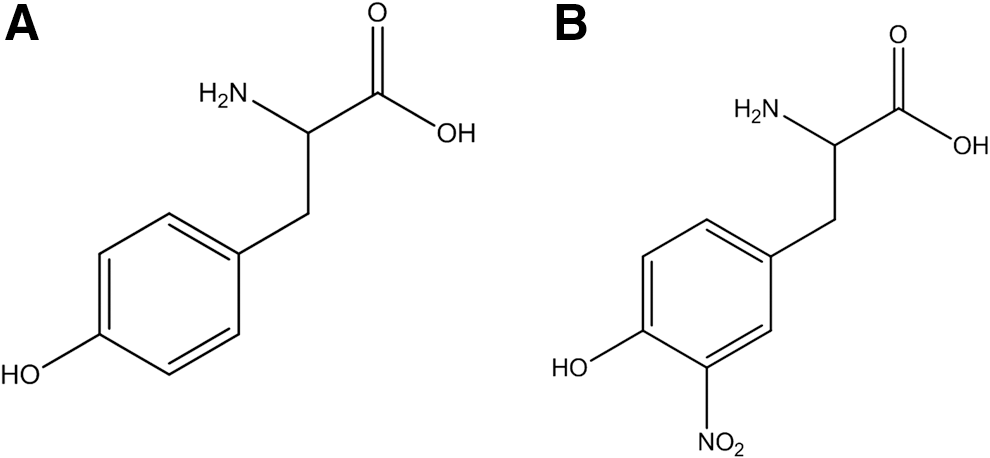
In addition to nitration, inflammatory conditions promote other oxidative modifications in tyrosine, such as chlorination, bromination, hydroxylation, and dimerization to 3-chloro-, 3-bromo-, 3-hydroxytyrosine, and 3,3′-dityrosine (82). Moreover, in proteins, another amino acid can be nitrated, namely tryptophan (5, 9, 29, 54, 55, 88), although the existing information and relevance of this modification remains preliminary and scarcely defined yet. The identification of these oxidative modifications is important not only because they may help in identifying preferential nitration pathways (82) but also because of technical difficulties that they may cause in the identification of tyrosine-nitrated proteins (87, 114). For a long time, tyrosine nitration was considered a stable PTM of proteins, although putative evidence on in vivo denitration processes has been provided (4, 43, 106), which requires further confirmation.
Physicochemical Properties and Biological Consequences of Tyrosine Nitration
Tyrosine nitration affects different physicochemical properties of the amino acid residue and of the corresponding protein: (i) causes a decrease in the pKa of the phenolic hydroxyl group, from 10.2 to ∼7.2 for the free amino acids in aqueous solution; this decrease will be affected by the protein environment (i.e., nearby presence of basic vs. acidic amino acid residues) and/or the properties of the milieu (i.e., less polarity will increase the pKa value) (91, 107, 122); (ii) changes the amino acid size by the incorporation of a substituent (30 Å3 larger) (124); (iii) changes light absorption with the appearance of a band centered at 360 nm at acidic pH and 420 nm at alkaline pH (107) (i.e., ionization of the phenol group in 3-nitrotyrosine causes a large red shift in absorptivity); (iv) influences the ionization process of 3-nitrotyrosine under certain mass spectrometry circumstances (4); and (v) increases the hydrophobicity of the nitrated peptide (14, 68). Even when the addition of a nitro group to a tyrosine residue confers particular physicochemical properties to the modified amino acid residue (and the corresponding protein), which may have important functional consequences (4, 82), the small fraction of nitrated protein has questioned its possible biological relevance (82). Protein tyrosine nitration is a process that does not occur randomly but clearly, as a selective process, which means that a relatively limited number of proteins are preferential targets of nitration (less than 110 nitrated proteins, and usually much less, were found in any of all the papers that aimed at identifying nitrated proteins by proteomics methods) (4). This “limited” number of nitrated proteins contrasts with other common PTMs of proteins, such as phosphorylation and acetylation. Enzymatic acetylation of the N-terminus of proteins occurs in 80–90% of higher eukaryotic proteins and acetylation of other lysine residues, most notably in histones and transcription, typically involves several hundreds of different proteins (45, 48, 49). In the same way, it is estimated that about 30% of the ∼30,000 proteins that were identified in the human proteome are substrates for phosphorylation (25). To date, acetylation is a very common and important PTM that regulates many important cell-signaling pathways. Actually, due to its frequency and important role in regulating the metabolism, acetylation has been proposed as a PTM that rivals phosphorylation (58). In the context of
Within the small number of nitrated proteins, only one or a few specific tyrosine residues were shown to be nitrated (14, 109, 110). Because the species participating in nitration reactions have short diffusion distances and particular formation sites, nitration reactions can be concentrated on proteins of either intra- or extracellular compartments (82). For example, under both physiological and pathological conditions, mitochondria typically contain larger amounts of nitrated proteins with respect to other cellular compartments, underscoring their role as a continuous source of peroxynitrite (63, 85, 95). On the other hand, in plasma, apolipoprotein A1 (ApoA1) in high-density lipoprotein has a much larger extent of tyrosine nitration than apolipoprotein B in low-density lipoprotein (34), mainly due to the fact that apoA1 binds one of the major extracellular sources of vascular nitrating species myeloperoxidase (MPO) (99, 100); the association of ApoA1-MPO under conditions of excess vascular
Thus, although there is reportedly a small fraction of tyrosine-nitrated proteins, nitration can be focused on specific tyrosine/proteins, resulting in loss or gain of important protein functions (4, 82, 110). As mentioned earlier, incorporation of -NO2 group to a tyrosine lowers the pKa of its phenolic -OH by ∼3 pH units and adds a bulky substituent. If placed in a relevant tyrosine residue, nitration can alter protein function and conformation, modify the sensitivity for proteolytic degradation, impose steric restrictions, and inhibit tyrosine phosphorylation. However, to have biological significance, a loss-of-function modification requires a large fraction of protein to become nitrated at specific critical tyrosine residues and it is doubtful that many proteins will undergo such an extent of nitration, with one of the few already well-demonstrated examples being the case of Mn-SOD (16, 63, 64, 84). On the other hand, sometimes, tyrosine nitration results in a gain of function (82). In that case, only a small fraction of a particular nitrated protein can elicit a substantive, “new” biological signal. This initial attractive concept has been already shown in a few proteins such as cytochrome c, which acquires a strong peroxidase activity after nitration (14, 22, 23, 108, 110) and translocates into the cytosol without initiating apoptosis (41). In the same way, nitrated fibrinogen accelerates clot formation (116); protein kinase Cɛ becomes activated and translocates on nitration (8); peroxynitrite transforms nerve growth factor into an apoptotic factor for motor neurons (72); and α-synuclein gains activity due to nitro-oxidative modifications (110).
A remarkable example of loss of enzyme activity linked to nitration in vivo is the mitochondrial enzyme manganese superoxide dismutase (MnSOD). This protein is nitrated by peroxynitrite in Tyr 34 by an Mn-catalyzed process, which leads to enzyme inactivation (63, 64, 82, 119). MnSOD usually circumvents peroxynitrite formation by dismutating superoxide anion radical (O2
•−). However, due to kinetic factors, such as the reaction of
Even when tyrosine nitration has been implicated in the loss or gain of function of proteins, it should be noted that not all studies have shown unequivocally that tyrosine nitration is the main/unique cause for the gain or loss of function of the protein, since the nitrating agent and associated oxidants also modify other amino acid residues (i.e., cysteine, methionine, and tryptophan) (4). α-Synuclein is a particular example of the more general situation of an important protein that gains activity due to oxidative modifications (110).
Another important consequence of tyrosine nitration is the modification of the immunogenicity of the modified protein. It is well known that PTM of self-proteins may lead to the generation of new epitopes, triggering an immune response, which may be the cause of an autoimmune disease (4, 70). In apoptotic or inflamed tissues, a variety of PTMs of proteins (including nitrated) have been shown to accumulate (70). The accumulation of nitrotyrosine-containing autologous proteins in inflamed tissues has been reported; these appear as foreign proteins to the immune system and might induce an autoimmune response (4, 17). Elevated levels of anti-nitrotyrosine antibodies have, indeed, been determined in synovial fluid of patients with rheumatoid arthritis and osteoarthritis (57), as well as in the serum of patients with systemic lupus erythematosus (56). Moreover, it has been shown that intratumoral reactive nitrogen species production results in the nitration of the chemokine CCL2, a process associated with the prevention of the infiltration of antigen-specific T-cells to the inner core of the tumor (66).
Mechanisms and Selectivity in Protein Tyrosine Nitration
The nitration of protein tyrosine residues in biology is a free radical process. The best well-known nitration mechanism involves the initial one-electron oxidation of tyrosine to yield tyrosyl radical followed by a diffusion-controlled reaction with nitrogen dioxide (
Considering the free radical nature of the nitration process, some caveats have to be indicated when correlating proteomic data obtained from exposure of proteins to chemical- versus biologically generated nitrating species. For example, exposure of proteins to a “bolus” of peroxynitrite [half-life of 0.8 s at pH 7,4 and 37°C (83)] can generate large initial concentrations of radical species in the bulk phase, which lead to a high initial level of tyrosyl radicals and, subsequently, 3-nitrotyrosine in solvent-exposed moieties. It is recommended that some of the experiments with peroxynitrite should be performed using a low flux of peroxynitrite and even those with the peroxynitrite donor SIN-1 should do so (14, 96), which better recapitulate biological conditions and provide a way to (i) minimize nonrelevant radical reactions and (ii) allow intramolecular electron transfer processes to modulate sites and extents of tyrosine nitration (e.g., tyrosyl radical repair by adjacent cysteine residue) (11 –13, 65).
Tyrosine is usually an abundant amino acid in proteins, and its content typically ranges from about 3% to 4% (10). However, under biologically relevant levels of nitrating species, very few tyrosine residues become nitrated, making nitration a selective free radical process. Factors influencing regio-selectivity include the protein structure, the nature of the proximal nitrating species or mechanisms, the redox environment, and the physicochemical properties of the reaction milieu, all of which will impact the nitrated protein residue (4, 14) (Table 1). The influence of the protein structure on the regio-selectivity of Tyr nitration and of some predictive models on protein nitration sites have been communicated recently elsewhere (4, 12, 15, 84). The overall conclusion is that although there are some factors that favor effective and selective tyrosine nitration, the complexity of the process requires, from a proteomic perspective, a case-by-case analysis, which also assists in revealing the nature of the proximal nitrating species.
Analytical Methods for the Detection of 3-Nitrotyrosine
The chemical structure of the analyzed species is very important for the development of any analytical method. In that sense, as stated earlier (114), 3-nitrotyrosine is very unusual, because even though it is a small molecule (molecular mass 226.19 g/mol), it has four functional groups: (i) an alpha acid carboxylic group; (ii) an alpha amino group; (iii) a phenolic group; and (iv) a nitro group at the ortho position of the aromatic group. These structural features have been used to develop different methods for the qualitative and quantitative analysis of 3-nitrotyrosine (114). Some approaches are based on chemical modification of one or more of these functionalities to improve analytical performance (e.g., detector response, volatility, chromatographic properties), whereas others target the unmodified form (114). 3-nitrotyrosine in different forms (i.e., native/unlabeled or stable-isotope labeled) has been extensively analyzed by liquid chromatography–mass spectrometry (LC-MS) and liquid chromatography–tandem mass spectrometry (LC-MS/MS) in both the positive and negative electrospray ionization (ESI) modes, and it has been recently reviewed elsewhere (114) (Table 2). In ESI+, protonated native 3-nitrotyrosine ([M+H]+ = m/z 227) produces few significant product ions when fragmented by collision-induced dissociation (CID). Some of these product ions have been used for quantitative analyses, and the use of the immonium ion of nitroTyr (m/z 181.06) is remarkable as it requires a relatively low collision energy to be produced and gives a very good signal (114). In ESI- mode, CID of the deprotonated unlabeled 3-nitrotyrosine ([M-H]− = m/z 225) gives a product ion at m/z 163 and this has been used in quantitative analyses (81). Protein-bound nitro-tyrosine has been related with the risk of coronary artery disease (CAD) (101 –103). The authors reported that protein-bound nitrotyrosine levels in plasma were significantly higher among patients with CAD (median 9.1 μmol nitro-tyrosine/mol tyrosine [interquartile range, 4.8–13.8 μmol/mol] vs. 5.2 μmol nitro-tyrosine/mol tyrosine [interquartile range, 2.2–8.4 μmol/mol]; p < 0.001) (101) (Table 2).
Adapted and updated from Tsikas and Duncan (114).
CAD, coronary artery disease; ND, not detectable; NR, not reported.
In our group, 3-nitrotyrosine determinations have been regularly performed according to the method described (65, 69). Basically, the quantification of nitroTyr in biological matrices was performed using an LC-MS approach (triple quad-ion trap MS/high-performance liquid chromatography) with the use of stable isotopically labeled precursors as synthetic internal standards, thus considering the intra-preparative sample losses and monitoring potential artifactual generation of nitroTyr during sample preparation (69). The described method allows rapid and reproducible quantification of nitroTyr in biological and clinical samples at the 100 fmol on column detection limit (69).
Proteomic Methods to Study Tyrosine-Nitrated Proteins
There are at least two main situations where researchers may need to identify tyrosine-nitrated proteins and, moreover, the tyrosine residue(s) that is (are) being nitrated; we will discuss the general strategies we have used to resolve both of them.
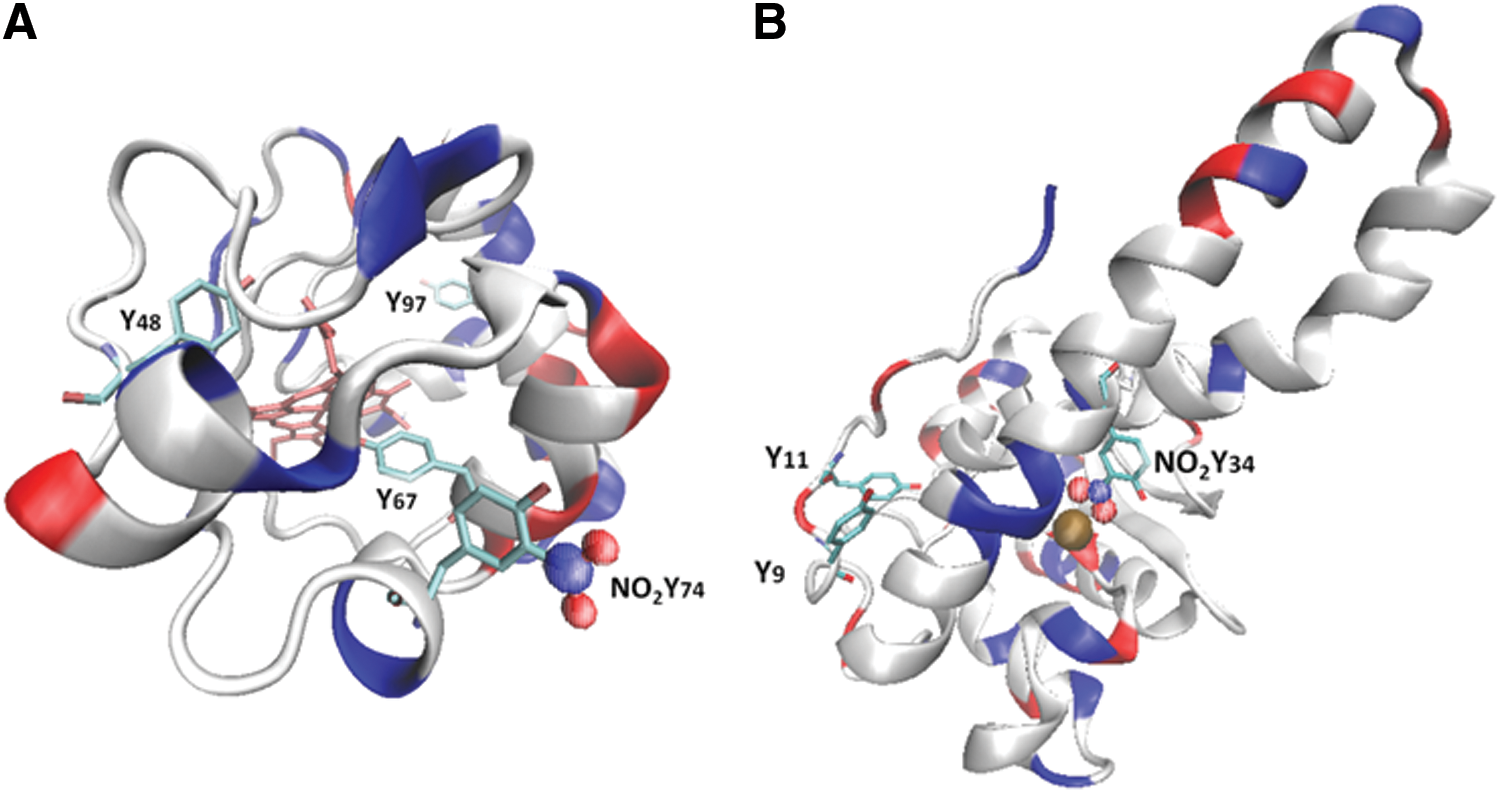
In the first case, probably the “simplest” situation, one can imagine the case of a pure protein that is being treated “in vitro” by one or more different nitrating agents. That was the case when we first analyzed the nitration of cytochrome c by tetranitromethane (TNM) and peroxynitrite in the presence or absence of bicarbonate (14). Cytochrome c contains four highly conserved tyrosine residues, three of which are highly exposed to the solvent (Tyr 48, Tyr 74, and Tyr 97). To analyze the feasibility of each tyrosine to become nitrated, we analyzed the time course and site(s) of tyrosine nitration in horse cytochrome c by fluxes of peroxynitrite or TNM (14). In that case, we developed a cation-exchange high-performance liquid chromatography (HPLC) method for purifying each nitrated cytochrome c product. Then, each purified nitrated protein was digested and a peptide map was performed by reverse-phase HPLC with double detection of peptides at 220 and 360 nm. It should be noted that 3-nitrotyrosine could form an internal hydrogen bond between the nitro and the phenolic OH-group, causing its absorption properties to be strongly pH dependent (4, 28). At acidic pH (pH <6), as is usually the case for reverse-phase chromatography of peptides, 3-nitrotyrosine has an absorption maximum at 360 nm wavelength (25, 91), which has been used for the detection of 3-nitrotyrosine-containing peptides by HPLC with ultraviolet-visible spectroscopy under acidic conditions (4, 26, 53, 109).
In our study, a flux of peroxynitrite caused a time-dependent formation of different nitrated species (Fig. 2). At low doses of peroxynitrite, the main products were two mono-nitrated cytochrome c at Tyr 97 and Tyr 74, as shown by peptide mapping and matrix-assisted laser desorption/ionization–time of flight (MALDI-TOF) mass spectrometry (MS) analysis (Fig. 2; peak B and C). At higher doses, almost all tyrosine residues in cytochrome c were nitrated, including dinitrated (i.e., Tyr 97 and Tyr 67 or Tyr 74 and Tyr 67) and trinitrated (i.e., Tyr 97, Tyr 74, and Tyr 67) forms of the protein. Interestingly, all mono-, di-, and trinitrated cytochrome c species displayed an increased peroxidase activity and were unable to restore the respiratory function of cytochrome c-depleted mitochondria (14). The nitration pattern of cytochrome c in the presence of TNM was comparable to that obtained with peroxynitrite, but with a relatively increased nitration yield at the buried Tyr 67. Interestingly, cytochrome c nitration by a “peroxidatic” mechanism involving the participation of its heme center (i.e., the initial reaction of H2O2 with the ferric heme) in the presence of either NO2
− or
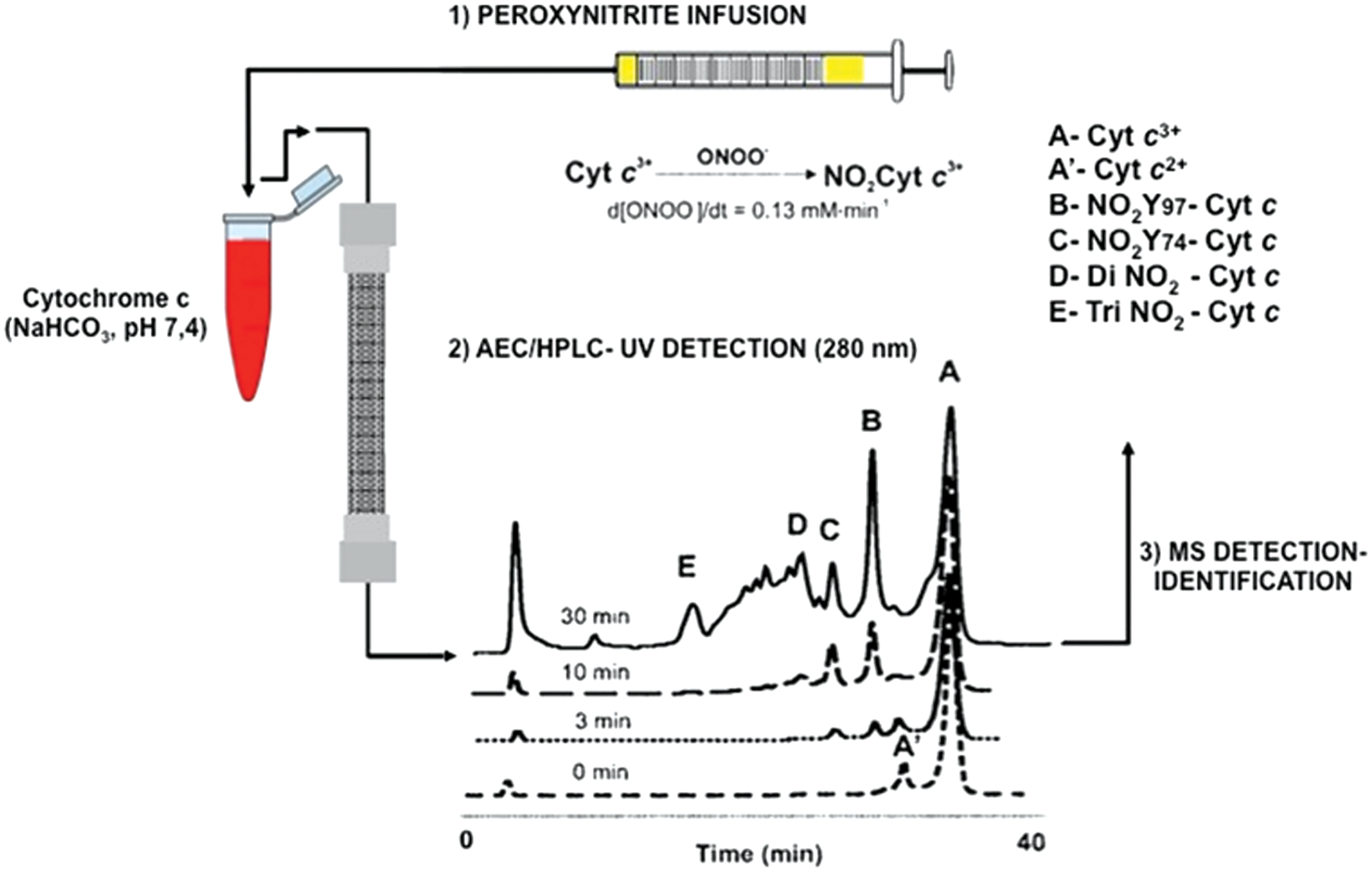
Thus, the purification of mono-, di-, and tri-nitrated species by cation-exchange HPLC coupled with reverse-phase HPLC with peptide detection at 220/360 nm allowed us to better define the sensitivity of each Tyr residue to become nitrated (14, 22). The MALDI-TOF machine we used was equipped with a laser light that emits at 337 nm. Nitrotyrosine-containing peptides are sensitive to this wavelength light and undergo a decomposition process, yielding a unique triplet signal consisting of the expected mass increase of 45 amu for the addition of a nitro group (NO2-Tyr; [M + H+ + 45]) as well as other major peaks corresponding to nitrosotyrosine (NO-Tyr; [M + H+ + 29]) and aminotyrosine (NH2-Tyr; [M + H+ + 15]) (14, 97). In our study, the expected fragmentation pattern of nitrated tyrosine-containing peptides on the ionizations induced by the nitrogen laser of the MALDI-TOF mass spectrometer was also observed and allows us to further identify the nitrated peptides (14). This is due to the fact that the absorbance maximum of 3-nitrotyrosine under acidic conditions is ∼360 nm, which is in the vicinity of the emission wavelength of the nitrogen ion laser (337 nm) (4, 97).
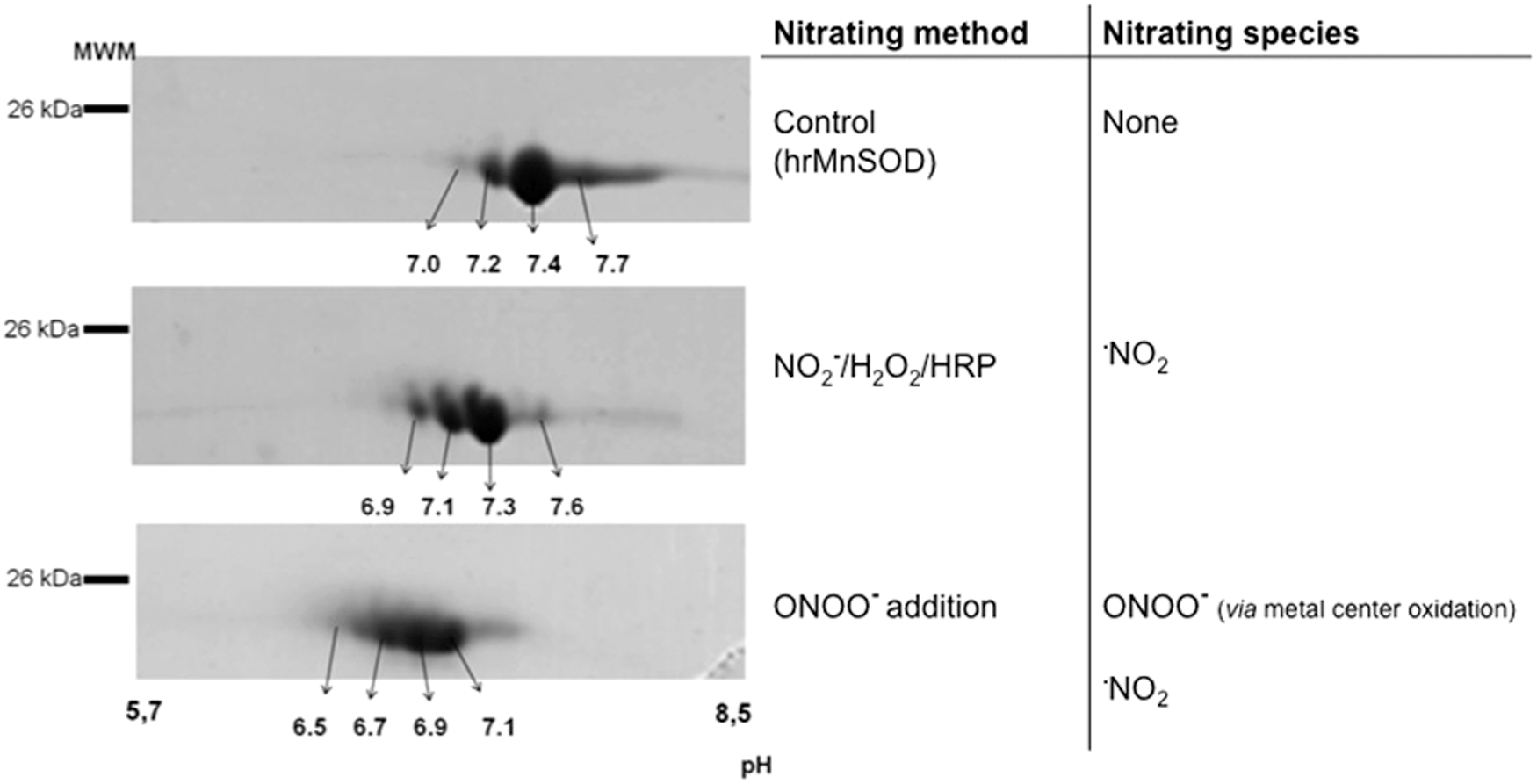
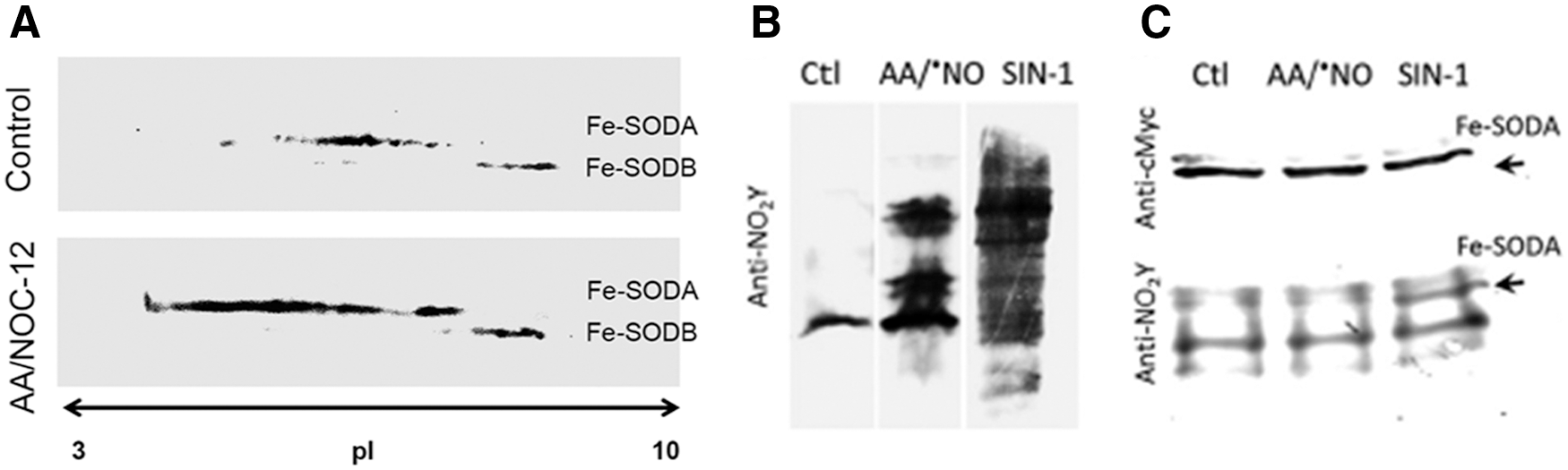
On the other hand, the analysis of the nitroproteome of tyrosine-nitrated proteins in more complex samples requires more complex analytical strategies. Up to now, the study of the “nitroproteome” by two dimensional-electrophoresis (2DE) has resulted in most of the identifications of known endogenously nitrated proteins (4). In our lab, the general strategy based on 2DE fractionation, followed by immunoblotting, in-gel digestion, and MS identification, has been widely used for nitroproteome studies of both single nitrated proteins (e.g., human MnSOD [hMnSOD], Fig. 4) (30) and more complex samples (19, 61, 65). Mn-SOD, an essential mitochondrial antioxidant enzyme, is nitrated and inactivated in vivo under a variety of conditions, leading to mitochondrial dysfunction and/or inflammation (30, 31, 63, 80, 119). The nitration of MnSOD in a critical tyrosine residue (Tyr 34) represents a prime example of an oxidative PTM in vivo that is significant and directly associated with a loss of function (63, 82, 119). hMnSOD contains a total of nine residues and depending on the nitrating agent and mechanism of nitration, one or more of them could be nitrated with a different probability (30, 112). In this regard, observed changes in isoelectric point of tyrosine-nitrated samples (Fig. 4) reflect the decrease in the pKa of each nitrated tyrosine and one should expect the generation of a new spot, more acidic, for each nitrated tyrosine in the protein. In MnSOD, four tyrosine residues were found to be nitrated (Tyr 2, Tyr 9, Tyr 11, and Tyr 34), and the distribution of the nitrated residues depends on the nitrating agent used. If peroxynitrite is the main nitrating agent, Tyr 34 is the primarily nitrated residue at low concentrations of peroxyntrite, with a prevalence of more than 80%, with functional consequences in the enzyme (30, 112). On the other hand, when nitrogen dioxide (
Even when in our experience this has been a very powerful strategy, we are aware that 2DE has several limitations that explain why the identified nitrated proteins are generally abundant and soluble ones. Moreover, sometimes, the major problem with this strategy is the failure to actually detect the nitrated tyrosine residue and this may be more likely due to: (i) limited protein load capability of the technique combined with the low levels of nitro-tyrosine on a given protein and the low abundance of the tyrosine-nitrated protein in a given sample, (ii) the recovery of nitrotyrosine-containing peptides from the gels and/or HPLC nanocolumns during subsequent nLC-MS/MS analysis (4, 19, 61, 65). It is also important to note that another important limitation of this strategy is its difficulty to analyze hydrophobic membrane proteins; this limitation becomes of particular relevance when studying tyrosine nitration in biological samples, since this PTM can be present in protein transmembrane domains and apolipoproteins (4, 10 –13, 118, 120, 126, 127). Finally, since a spot may contain more than one protein (depending on the quality of the 2D-gel and the MS being used for the analysis, this number may vary between one and more than 10 proteins/spot), it is sometimes tricky to assign an immuno-positive spot on the blotting membrane to the identified protein. It is, therefore, mandatory to confirm the nitration site of the protein by MS. As an example of this concept, recently, it was found that in 57 immuno-positive anti-nitrotyrosine spots, more than 800 proteins were identified, but only 18 were actually nitrated (76). Possible explanations regarding this unexpected observation in which an excess of “nonnitrated” proteins were detected on immune-positive spots for protein 3-nitrotyrosine will be analyzed in the next section in the context of possible technical limitations.
By using the 2DE-MS general strategy, using both an MALDI-TOF/TOF (4800 MALDI TOF/TOF Analyzer, Abi Sciex) and a nano-LC/nano spray ionization with a linear ion trap mass spectrometer (Easy nLC, Thermo; LTQ Velos, Thermo), we have also analyzed the nitration of Trypanosoma cruzi iron superoxide dismutases (Fe-SODs). The reason for studying this process is that, as was previously observed for other T. cruzi antioxidant enzymes (2), T. cruzi Fe-SODs may function as virulence factors in Chagas Disease (American trypanosomiasis), a hypothesis currently under investigation in our laboratory. It is important to note that during T. cruzi invasion to immunostimulated macrophages, T. cruzi proteins become nitrated inside the phagosome by cytotoxic levels of host cell-derived peroxynitrite (6). In this context, the nitration of parasite Fe-SOD in key tyrosine residues during host cell-T. cruzi interactions by
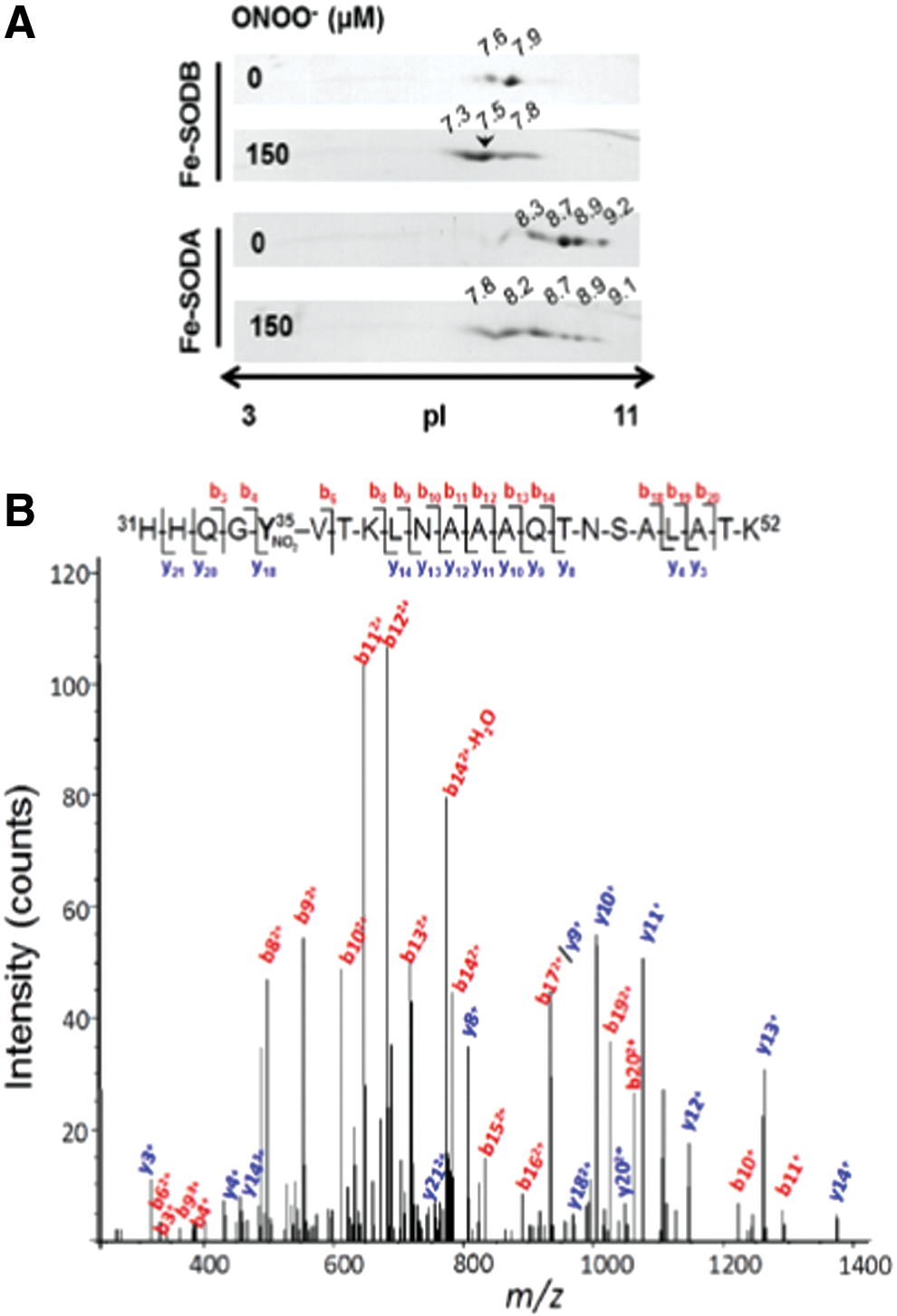
Proposal of an Alternative Method to Improve the Sensitivity and Selectivity for the MS-Analysis of Tyrosine-Nitrated Proteins
Precise quantitation and identification of nitrated tyrosine residues in proteins can be a “tour de force” for researchers working in the redox biology and chemistry area: The difficulties probably arise from the relatively small number of nitrated tyrosine versus total tyrosine residues (meaning the presence of one modified peptide per a large number of native ones), hydrophobicity of tyrosine-nitrated peptides, instability of the modification when analyzed by MALDI ionization, and other still unknown factors. To partially solve these problems, we consider that a good analytical approach should be the combination of the best orthogonal methods to separate proteins and peptides in tandem, with the more sensitive MS-based method to detect nitrated tyrosine peptides. In that sense, the “ideal” method should separate total proteins by 2D electrophoresis followed by immunodetection of Tyr-nitrated spots. Regarding this approach, one should carefully analyze the specificity of the anti-nitrotyrosine antibody and also remember that the identification of the spot is carried out on a membrane after transferring the protein from the gel, and the in-gel digestion is performed on the gel. Thus, there are at least two technical issues (i.e., antibody specificity and mistake when selecting the superimpose place to cut) that may affect the quality and robustness of the experimental result (76).
Proteins present in those spots (it is always important to remember that one single, well-resolved spot in a 2D gel typically contains more than one protein) should be identified by traditional MS methods (61, 76, 114). Once all the proteins present in that spot have been identified, one should be able to predict, in silico (software assisted) (114), all the possible Tyr-containing tryptic peptides (114) and apply a targeted mass spectrometric workflow to the sensitive identification of nitrated Tyr residues (115). The MIDAS™ (multiple reaction monitoring [MRM] initiated detection and sequencing or MRM-triggered MS/MS method) MS strategy employs MRM to search for all putative peptides that are specifically modified in a target protein. Positive MRMs above a preset threshold trigger an MS/MS (or an enhanced product information [EPI]) experiment to confirm the nature and site of the modification (2, 115). The selection of which fragment ions are the best option to use for both Q1 and Q3 for the MRM experiment is either based on previously acquired MS/MS identifications or predicted in silico based on the peptide sequence (1, 3). With these theoretical data, researchers may use a triple quadrupole MS or a hybrid triple quadrupole-ion trap MS to run the MIDAS experiment method to detect any possibly Tyr-nitrated peptide in that spot (2), obtaining the best sensitivity for both ID and quantitation of the peptide (Fig. 7). To further confirm the nitration of the peptide/protein, it may be wise to perform an EPI of the detected peptide to obtain all the MS/MS data to unequivocally confirm the nitration site. In this sense, the use of an ESI source-based MS method to identify nitrotyrosine-containing peptides has several advantages compared with an MALDI source: (i) ESI does not induce the characteristic photochemical decomposition of nitrotyrosine produced under MALDI conditions; (ii) under ESI conditions, nitrotyrosine fragmentation produces an immonium ion (m/z 181.06) that is a very useful footprint of a nitroTyr-containing peptide. Thus, the presence of this footprint in the EPI experiment will increase the confidence of the identification.

Also, the formation of this immonium ion allows the design of another alternative method to increase the sensitivity of the identification/quantitation of nitrated tyrosine-containing peptides. In this case, researchers may use the precursor ion scanning mode to search for the immonium ion for nitrotyrosine, which will specifically identify a 3-nitrotyrosine-containing peptide/protein (77). Unfortunately, as stated elsewhere (125), a huge amount of non-nitrated tryptic peptides will negatively affect the characterization and identification of nitropeptides. For that reason, it is highly recommended to associate an immunoaffinity enrichment method for nitropeptides before the MS analysis.
Conclusions and Perspectives
The area of “nitroproteomics” continues to be under active development and has revealed to be quite challenging in technical terms. Proteomic approaches have revealed key features of protein tyrosine nitration both in vivo and in vitro, including its selectivity in terms of nitrated proteins and regio-specificity of nitrated residues. Of note is the relatively limited number of nitrated proteins detected under both normal and oxidative stress conditions, in comparison to other PTMs, including the NO-dependent process of S-nitrosylation, even when nitration is a more stable PTM than S-nitrosylation. However, it is challenging to identify, map, and quantitate nitrotyrosine-containing proteins due to the low abundance of this oxidative modification in biological samples and its unfriendly behavior in MS-based technologies, that is, MALDI, ESI, and CID (125).
Tyrosine-nitrated proteins are detected in cells and tissues under normal physiological conditions, revealing a basal continuous flux of nitrating species; the levels of nitrated proteins are strongly increased under conditions of enhanced formation of
The combined use of 2D gel electrophoresis with immunochemical detection of 3-nitrotyrosine followed by identification of proteins by regular MS/MS approaches plus the immuno enrichment of Tyr-nitrated peptides and the ID of those nitrated peptides by a MIDAS experiment (see before) is arising as a potent methodology to undoubtedly characterize and quantitate tyrosine-nitrated proteins in vivo. For this approach, the use of a highly specific and sensitive antibody, well characterized, is a key element in the quality and confidence of the results (19). Finally, MS-based proteomics, independently of all the other factors that may affect the quality of the results (i.e., sample preparation, mass spectrometer use, general methodological strategy used, etc.), always depends on the software that will be used to analyze and interpret the data; this is also of particular relevance when analyzing tyrosine-nitrated peptides/proteins.
Proteomic approaches to study protein tyrosine nitration alone or in combination with immunochemical and/or bioanalytical methods represent potent tools for (i) identification of protein targets and molecular footprints in a variety of pathological conditions associated to oxidative stress, (ii) identification of the proximal nitrating species in vivo via the peptide mapping data, (iii) revealing structural and functional effects of nitration of specific protein residues in proteins, and (iv) obtaining pure modified protein species nitrated on specific tyrosine residues only to be used in further studies. In parallel to tyrosine nitration, proteomic studies can also assist in assessing parallel oxidative and related PTMs (e.g., addition of electrophiles) that may simultaneously contribute to changes in protein structure and function and have an impact in biological outcome. New methodological developments and research in proteomics of nitrated proteins open opportunities for identification of specific nitrated proteins and peptides as biomarkers and predictors of specific disease conditions, their possible participation in signaling cascades, and their role in inflammatory, immune, and degenerative diseases (66, 70, 76).
Footnotes
Acknowledgments
This work was supported by grants of Agencia Nacional de Investigación e Innovación (FCE_2014_104233), Universidad de la Republica (CSIC), and the National Institutes of Health (RO1 AI095173) to R.R. and Universidad de la República (CSIC) and Institut Pasteur de Montevideo to CB. Additional support to the investigators was obtained from Programa de Desarrollo de Ciencias Básicas (PEDECIBA), Universidad de la República (CEINBIO, Espacio Interdisciplinario), Centro de Biología Estructural del Mercosur (CeBEM), and Ridaline and Tecniplast through Fundación Manuel Perez.