
Abstract
Aims:
Changes in the composition of the gut microbiota contribute to the development of obesity and subsequent complications that are associated with metabolic syndrome. However, the role of increased numbers of certain bacterial species during the progress of obesity and factor(s) controlling the community structure of gut microbiota remain unclear. Here, we demonstrate the inter-relationship between Drosophila melanogaster and their resident gut microbiota under chronic high-sugar diet (HSD) conditions.
Results:
Chronic feeding of an HSD to Drosophila resulted in a predominance of resident uracil-secreting bacteria in the gut. Axenic insects mono-associated with uracil-secreting bacteria or supplemented with uracil under HSD conditions promoted larval development. Redox signaling induced by bacterial uracil promoted larval growth by regulating sugar and lipid metabolism via activation of p38a mitogen-activated protein kinase.
Innovation:
The present study identified a new redox-dependent mechanism by which uracil-secreting bacteria (previously regarded as opportunistic pathobionts) protect the host from metabolic perturbation under chronic HSD conditions.
Conclusion:
These results illustrate how Drosophila and gut microbes form a symbiotic relationship under stress conditions, and changes in the gut microbiota play an important role in alleviating deleterious diet-derived effects such as hyperglycemia. Antioxid. Redox Signal. 27, 1361–1380.
Introduction
T
It is assumed that the increasing abundance of bacterial species during disease states is responsible for disease progression. However, factor(s) controlling the community structure of gut microbiota and the exact roles of the abundant microbes in disease progression are scarcely known. Here, we show that larval growth was promoted by the abundant autochthonous uracil-secreting bacteria in the gut of high-sugar diet-fed flies. Redox signaling induced by bacterial uracil promotes larval growth by regulating sugar and lipid metabolism via activation of p38a mitogen-activated protein kinase (MAPK). These results highlight a novel redox-dependent mechanism through which uracil-secreting bacteria protect the host from metabolic perturbation.
Reactive oxygen species (ROS) are short-lived and highly reactive molecules produced from incomplete reduction of oxygen metabolites (28). Oxidative stress due to overproduction of ROS or depletion of antioxidants causes oxidative damage to various cellular components, ultimately leading to neurodegenerative disease, cancer, and aging (1, 73). However, recent evidence suggests that physiological levels of ROS act as signaling molecules that play important roles in regulating various cellular pathways. In nematodes, for example, aging is not caused by oxidative damage; rather, increased mitochondrial ROS levels activate intrinsic apoptosis pathways and promote longevity (79). Several recent studies demonstrate the beneficial effects of redox-dependent signaling pathways in various model organisms: for example, increased host fitness and anti-parasite resistance due to increased ROS production in the mosquito midgut (18); ROS-induced intestinal stem cell proliferation and autophagy induction in fruit fly and mammalian cells (70); and reduced disease activity in diabetic mice and humans on increased superoxide production (19). Interestingly, the beneficial effects of ROS are more obvious when organisms are placed under conditions that induce physiological stress (41, 52).
ROS production is induced not only by mitochondrial oxidative phosphorylation but also in professional phagocytes via nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) and in non-phagocytic tissues (such as mucosal epithelia) by NOX and dual oxidase (DUOX) (3, 39, 77). Recently, Lee et al. (42) showed that the bacteria-derived ligand uracil elicits DUOX-dependent microbicidal ROS generation in the Drosophila gut. Uracil is mainly secreted by non-commensal “allochthonous” bacteria introduced from external environments. Accordingly, the uracil-mediated immune response is of critical importance for efficient elimination of invading bacteria and host survival during infection. Importantly, some evolutionarily adapted “autochthonous” bacteria (e.g., Gluconobacter morbifer or Lactobacillus brevis) also release uracil into the Drosophila gut lumen. These bacterial species are regarded as pathobionts because they are normally benign, but are conditionally pathogenic on artificial dysregulation of antimicrobial activity (e.g., knockout of the homeobox gene Caudal) within a host (57). This raises an important question: Why have flies maintained these conditionally pathogenic bacteria in the gut during evolution?
The present study aimed at identifying the potential roles of these bacteria in gut microbial symbiosis by examining the inter-relationship between flies and their resident gut microbiota under chronic high-sugar diet (HSD) conditions. We found that larval growth was promoted by ROS production via both DUOX and mitochondria as a consequence of the predominant abundance of autochthonous uracil-secreting bacteria in the gut of HSD-fed flies. We also showed that redox signaling induced by bacterial uracil increased the levels of trehalose in the hemolymph and induced fat body lipogenesis. The level of glucose in the hemolymph was also reduced. Alterations in these three parameters may be due to activation of p38a mitogen-activated protein kinase (MAPK), although it is unclear as to which is affected to the greatest extent. These results illustrate how Drosophila and uracil-secreting bacteria form a symbiotic relationship under stress conditions.
Results
Bacteria resident in HSD-fed flies promote host development
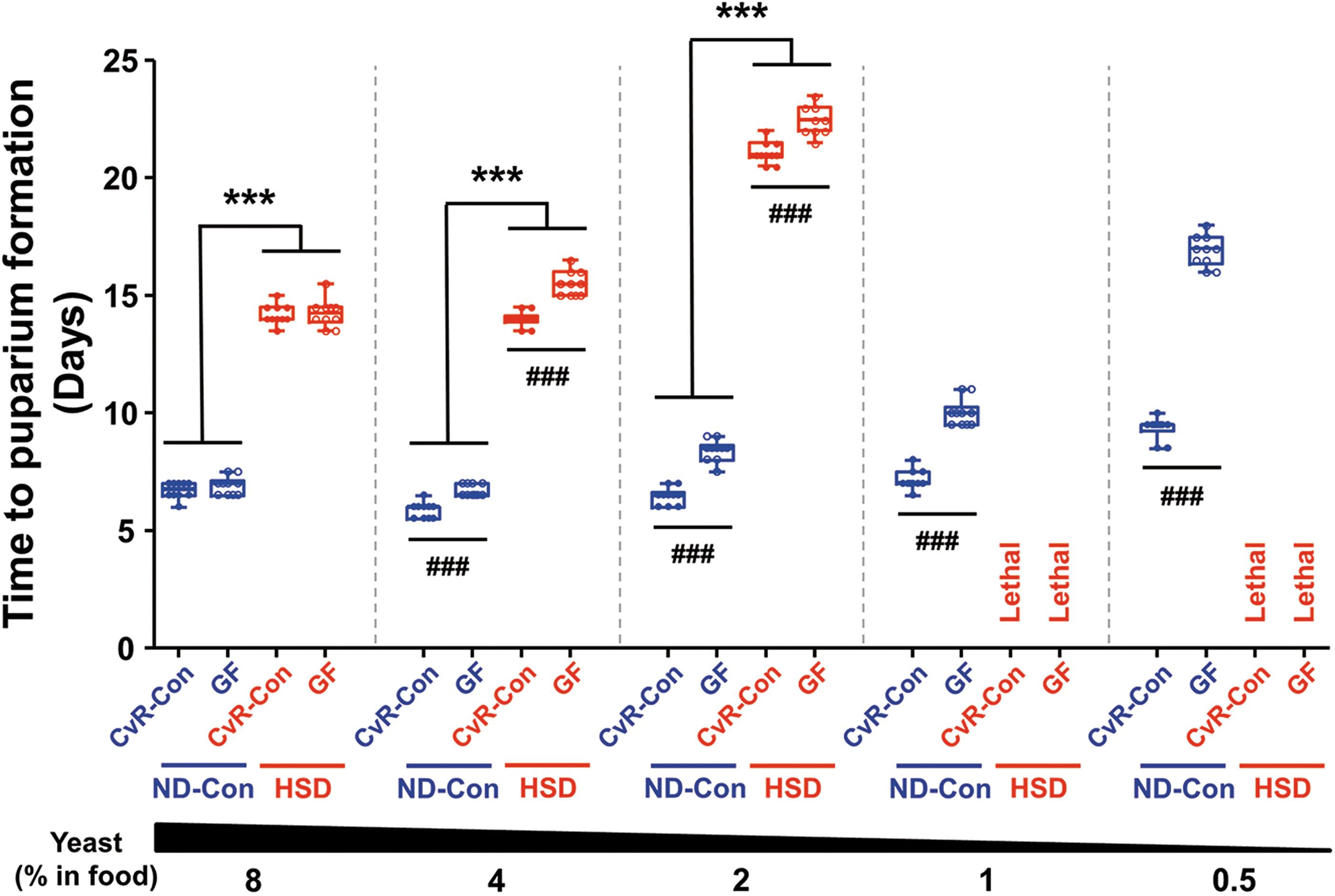
High levels of sugar in the diet increase the risk of metabolic syndrome, which includes obesity and insulin resistance, in Drosophila (49). Accordingly, we evaluated the effect of resident bacteria on the developmental period of insect larvae, which is a critical estimate of growth rates under a given set of nutrient conditions (17). When we compared the time to larval puparium formation (LPF) after egg laying in control normal diet (ND-Con)- and HSD-fed flies, we found that the LPF for HSD-fed larvae was significantly longer (Fig. 1). Feeding an HSD containing low concentrations of yeast (<1%) resulted in a lethal host phenotype. Interestingly, we observed a statistically significant difference in the time to LPF between conventionally reared control (CvR-Con) and germ-free (GF) flies. Developmental delay was observed in both the ND-Con- and HSD-fed groups of GF larvae, and the effect of resident bacteria on larval development was more pronounced as the yeast concentration in the diet was reduced. The beneficial effects of commensal bacteria and the mechanisms underlying host-microbe interactions in normal diet-fed flies have been described elsewhere (65, 68). Therefore, our subsequent analyses focused on the beneficial role of resident bacteria in HSD-fed flies.
A chronic HSD leads to a predominance of G. morbifer in the gut microbiota
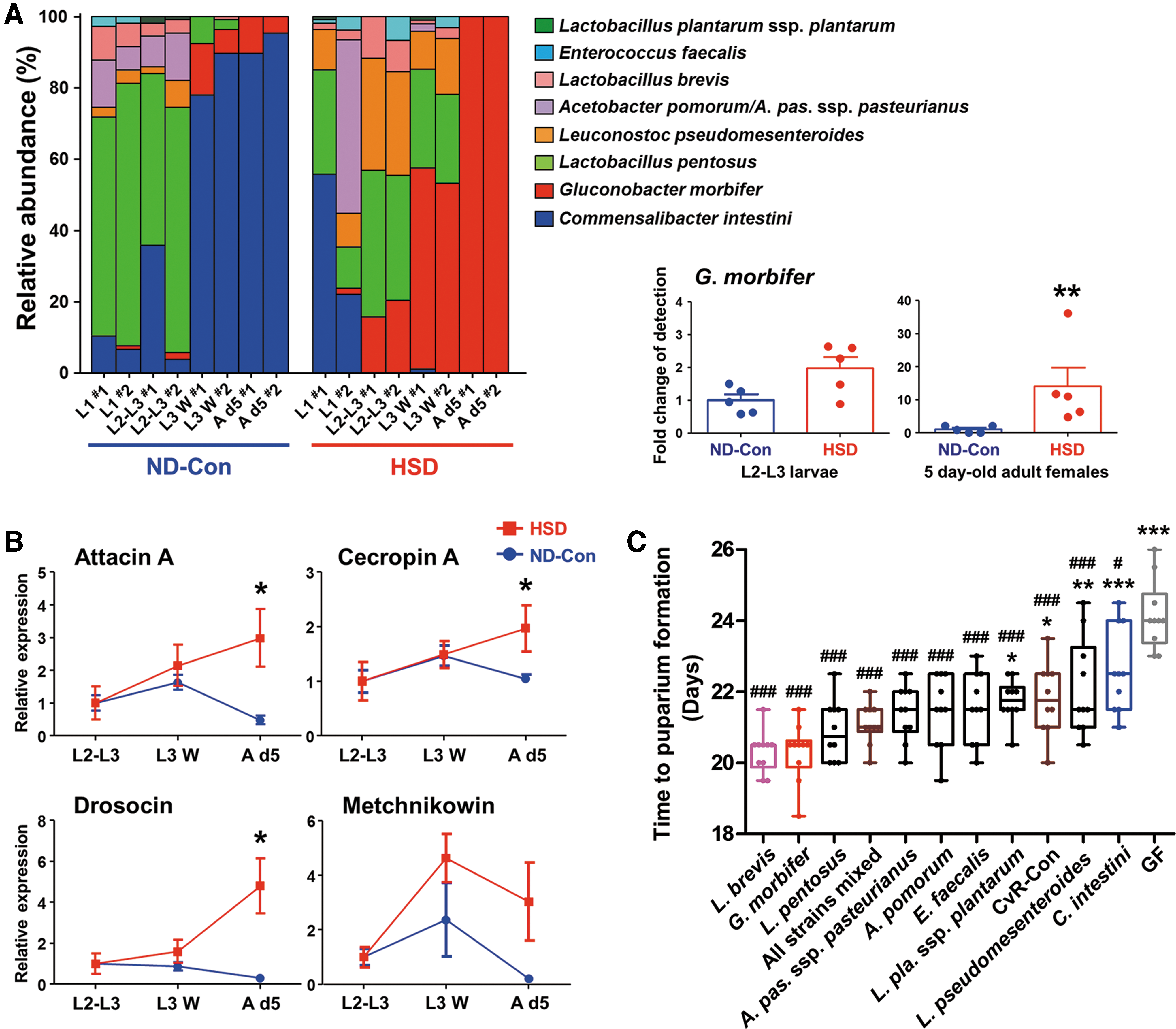
Based on metagenomic analysis of the bacterial 16S ribosomal RNA (rRNA) gene, we obtained taxonomic information about the bacterial communities that were resident in the gut of CvR-Con flies (Supplementary Fig. S1A; Supplementary Data are available online at
Larval development is greatest in HSD-fed flies mono-associated with L. brevis or G. morbifer
We next examined the effect of individual resident bacterial species on larval development under HSD conditions by introducing each of the cultured bacteria into GF larvae. Among the candidate bacterial species examined, only Acinetobacter ursingii failed to survive and colonize the host under the experimental conditions; thus, this species was excluded from further experiments (Supplementary Fig. S3). Overall, the developmental time of gnotobiotic insects harboring each of the nine autochthonous bacteria was shorter than that of the GF controls (Fig. 2C). Specifically, when exposed to an HSD, larvae mono-associated with L. brevis or G. morbifer showed the greatest decrease in LPF, whereas C. intestini or Leuconostoc pseudomesenteroides mono-associated larvae showed relatively poor growth. We reasoned that bacteria that consume high levels of sugar may reduce the absolute amount of sugar available to the host, and would, therefore, have a direct effect on host fitness by alleviating sugar toxicity. However, we found that only L. pseudomesenteroides was capable of utilizing sucrose to promote growth (Supplementary Tables S2 and S3). Insect midgut is capable of digesting sucrose into its constituent monosaccharides (i.e., glucose and fructose) by using midgut sucrose hydrolase(s). For example, a novel sucrose hydrolase has been found in Bombyx mori larval midgut and its homologues are also identified in Drosophila (75). The larval growth less-promoting bacteria (e.g., Lactobacillus plantarum ssp. plantarum, Lactobacillus pentosus, L. pseudomesenteroides, and Enterococcus faecalis) along with the larval growth-promoting bacteria (e.g., G. morbifer) are capable of utilizing both glucose and fructose (Supplementary Tables S2, S3). Thus, we did not consider the effects of bacterial sugar consumption on the physiology of HSD-fed insects.
Bacterial uracil promotes larval development under HSD conditions by inducing the production of intestinal ROS
A common feature of L. brevis and G. morbifer is the induction of DUOX-dependent ROS production in the intestinal mucosal epithelium via uracil secretion (42). Because redox-dependent signaling pathways are capable of enhancing host fitness and survival under stress conditions (52), we examined whether the HSD-boosted growth of larvae mono-associated with L. brevis or G. morbifer is attributable to intestinal ROS production induced by bacterial uracil. To this end, we fed GF larvae with an HSD containing uracil and measured the time to LPF. Dose-dependent analysis revealed that at least 10 nM (optimum, 20–80 nM) of uracil reduced the time of larval development in response to an HSD (Fig. 3A). The larval growth of Drosophila strains such as Yellow White and CantonS in response to uracil supplementation (40 nM) was more rapid than that of GF controls (Fig. 3B). To confirm that constitutive uracil secretion by conditionally pathogenic bacteria is a genuine factor that is responsible for increased larval development on an HSD, GF larvae were mono-associated with a uracil-auxotroph harboring a mutation in the carbamoyl phosphate synthase (CarA) gene (G. morbifer-carA::Tn5). The larval developmental period for gnotobiotic insects harboring the uracil-auxotroph G. morbifer was significantly longer than that for larvae mono-associated with the parental G. morbifer strain (Fig. 3C).

We next examined the effect of intestinal ROS production on the growth of HSD-fed insects. GF flies were fed an HSD containing different doses of hydrogen peroxide (H2O2) or paraquat (a potent ROS inducer). Larvae that ingested low doses of oxidant or paraquat had a shorter developmental period (Fig. 3D). Moreover, mono-association of uracil-secreting bacteria with transgenic flies ubiquitously expressing catalase (Da-Gal4; UAS-Catalase) led to a significant delay in the time to LPF when compared with that of their counterpart controls (Da-Gal4; +). Increased expression of catalase abolished the boosted growth observed in GF flies supplemented with uracil (Fig. 3E), implying that redox-dependent signaling mediated by bacterial uracil increases the fitness of host flies fed an HSD.
Conditional symbiosis is dependent on both commensalism and dietary sugar concentration
Since Erwinia carotovora ssp. carotovora 15 (Ecc15) in the Drosophila gut induces a local immune response by secreting uracil (7, 42), we next examined whether host development under HSD conditions is enhanced by non-resident uracil-secreting bacteria such as Ecc15. In addition, we included bacterial species frequently used to induce experimental sepsis as candidate taxa for mono-association in Drosophila (Supplementary Table S1). As shown in Figure 4A, gnotobiotic insects harboring each of the allochthonous bacterial species showed developmental times comparable with those of GF controls, suggesting that the beneficial effects of uracil secretion under HSD conditions might be masked by potential virulence factors that are intrinsic to non-resident bacteria. Interestingly, larvae mono-associated with Providencia rettgeri (isolated from Popillia quadriguttata) showed a shorter time to LPF than GF controls. The genus Providencia is frequently isolated from the gut of laboratory-cultured Drosophila species, and it is predominant when the flies are fed a sugar-only diet (12).
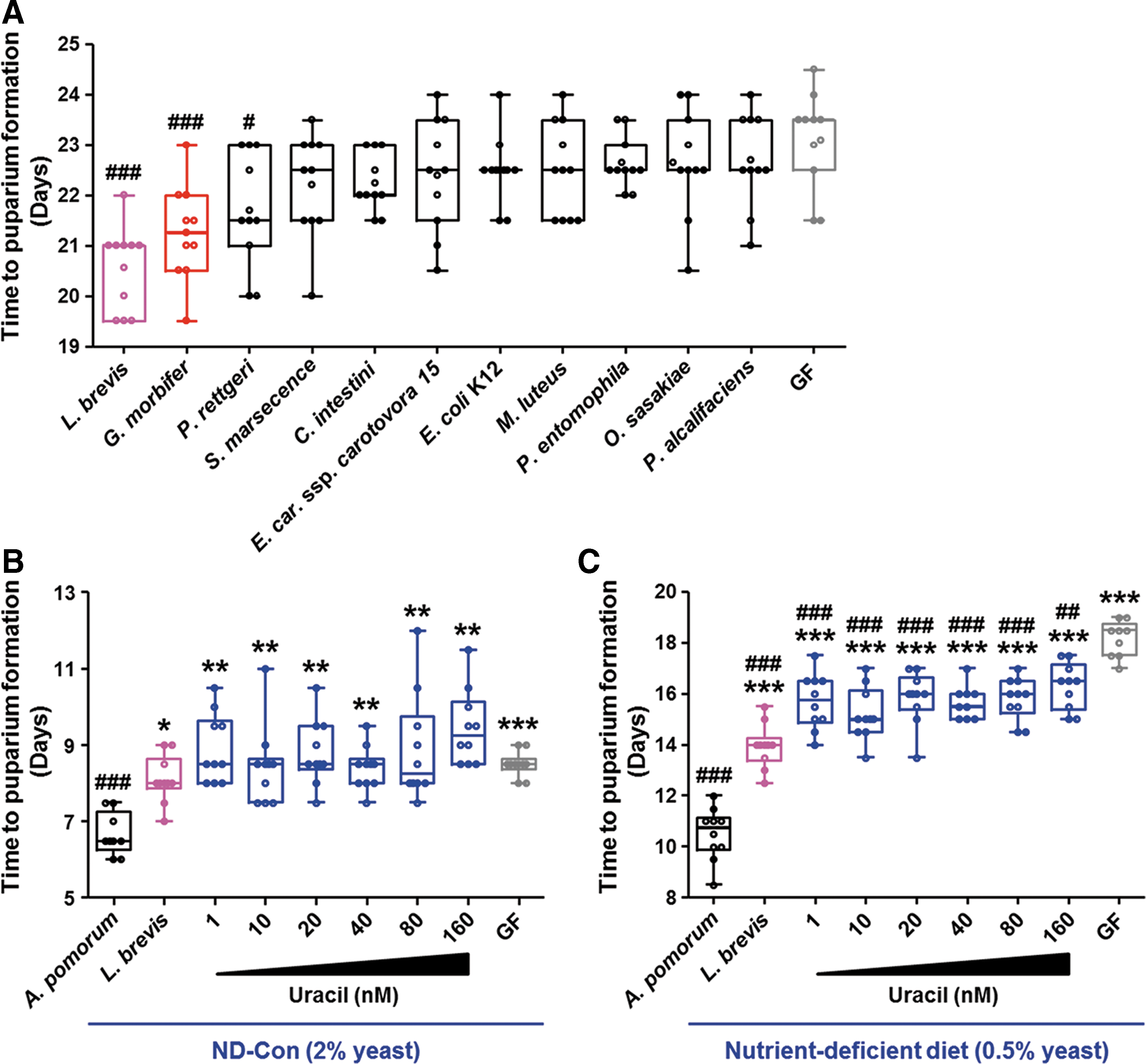
Both L. brevis and G. morbifer are minor members of the resident gut microbiota in Drosophila, and an overgrowth of these bacteria (as in the case of mono-association) is lethal to host flies under normal diet conditions (42, 55, 57). In accordance with these studies, we found that the advantageous effects of uracil ingestion were abolished in GF larvae under ND-Con conditions (Fig. 4B). In this case, larvae mono-associated with Acetobacter pomorum showed a shorter time to LPF than larvae harboring L. brevis, suggesting that the effect of these conditionally pathogenic bacteria on host physiology is dependent on the host diet, particularly the sugar concentration in the diet. We found a statistically significant difference in the host developmental period between uracil-fed GF larvae and GF controls under conditions of nutrient scarcity; this difference was caused by the reduced yeast concentration in the diet (Fig. 4C).
Uracil-secreting bacteria promote both increased levels of trehalose in the hemolymph and fat body lipogenesis under HSD conditions
We speculated that the metabolic profile of flies carrying conditionally pathogenic bacteria might differ from that of flies that do not carry these bacteria. If this was indeed the case, understanding the disparity between these conditions may be the key to the mechanism underlying conditional host-microbe symbiosis. Thus, we examined sugar and lipid homeostasis in the presence or absence of uracil-secreting bacteria. Mono-association of L. brevis or G. morbifer and uracil supplementation resulted in a significant reduction in glucose levels, but they markedly increased levels of trehalose, triglycerides (the major form of stored fat in flies), and glycogen (the major form of stored carbohydrate in flies) in HSD-fed larvae (Fig. 5A). It is noteworthy that the metabolic phenotype of the HSD-fed larvae harboring C. intestini and the GF controls was reminiscent of that in insulin pathway-impaired insects, in that it is characterized by high levels of free sugar and low levels of glycogen and lipids (66). Lipophilic staining of dissected fat bodies revealed that lipid droplets in larvae mono-associated with L. brevis or G. morbifer or in larvae supplemented with uracil were larger than those in larvae mono-associated with C. intestini or those in GF controls (Fig. 5B and Supplementary Fig. S4). Collectively, these results suggest that the uracil secreted from conditionally pathogenic bacteria in the gut of HSD-fed insects alters host metabolic status by increasing the level of trehalose in the hemolymph and by promoting fat body lipogenesis.
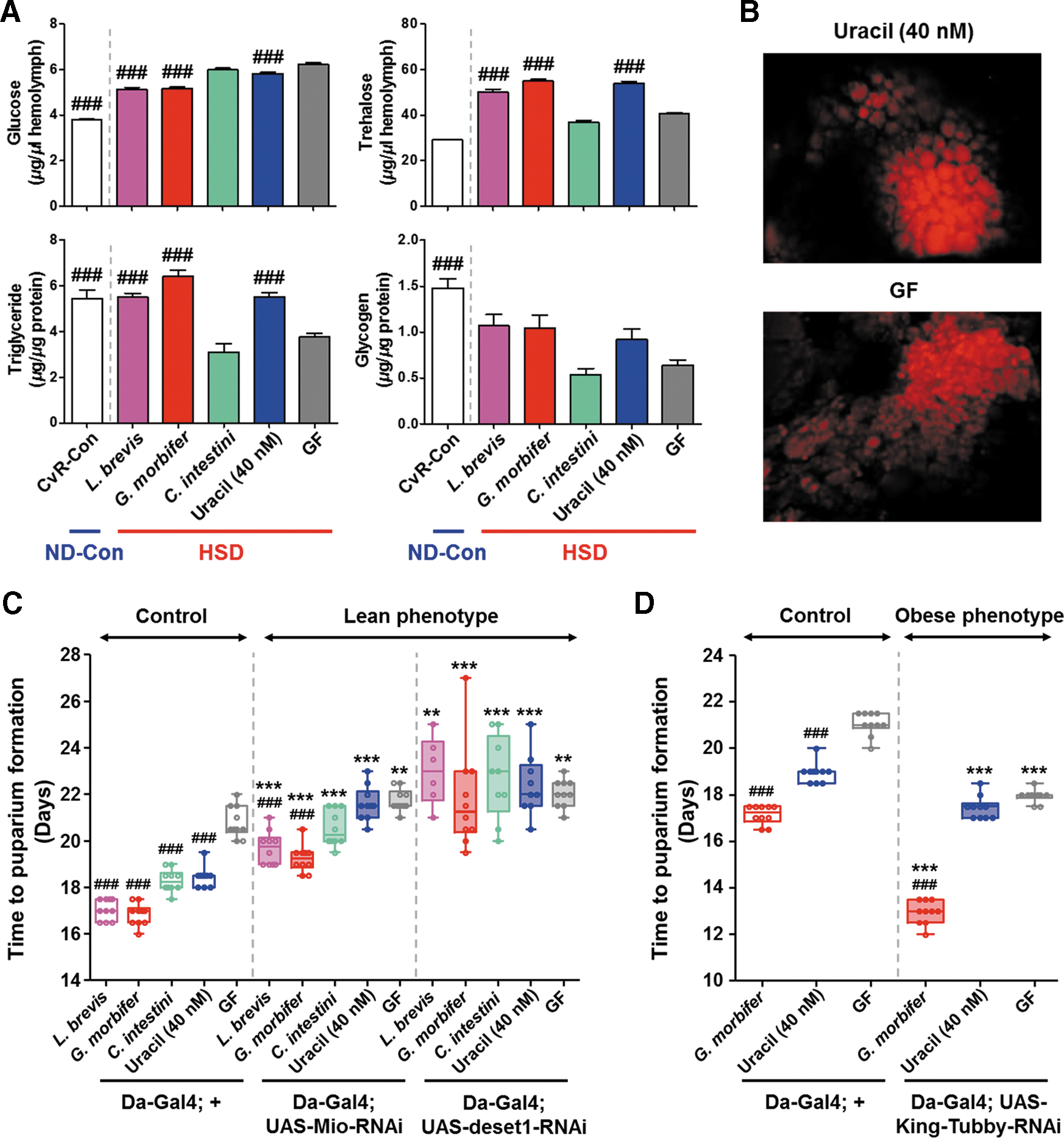
We next examined the relevance of metabolic status (e.g., the capacity for storing fat) to the fitness of HSD-fed larvae. Drosophila Max-like protein X (Mlx) interactor (Mio) and desat1 are putative lipogenic genes that encode proteins that are homologous to mammalian carbohydrate responsive transcription factor (mChREBP) and stearoyl-CoA desaturase 1, respectively (40, 58). The expression of these two genes is increased in HSD-fed larvae (49), and feeding an HSD to flies in which Mio or desat1 have been knocked down in the fat body results in flies showing both a lean phenotype and increased levels of glucose in the hemolymph (50). In accordance with these findings, we observed that Mio (Da-Gal4; UAS-Mio-RNAi) and desat1 (Da-Gal4; UAS-desat1-RNAi) knock-down flies showed a significantly longer time to LPF than their counterpart controls (Da-Gal4; +) (Fig. 5C). Also, the increased larval growth mediated by uracil was abolished. King-Tubby knock-down flies (Da-Gal4; UAS-King-Tubby-RNAi) exhibiting an obese phenotype also showed a reduced time to LPF (Fig. 5D), suggesting that lipogenesis protects against the deleterious effects of an HSD.
Activation of p38a MAPK, but not JNK, Nrf-2, or dFOXO, is required to protect the host from the deleterious effects of an HSD
Since signaling via both p38 MAPK and c-Jun N-terminal kinase (JNK) is involved in responses to cellular stressors, including oxidative stress (14, 76), we next explored whether activation of p38 MAPK and JNK is required for larval growth under HSD conditions. An HSD plus a p38 inhibitor (SB203580), but not a JNK inhibitor (SP600125), resulted in a longer time to LPF, with decreased host survival in larvae mono-associated with L. brevis (Fig. 6A and Supplementary Fig. S5A). The time to LPF of dominant negative JNK mutants (Da-Gal4; UAS-BSKDN ) and Puckered-overexpressing (Da-Gal4; UAS-Puckered) flies were comparable with those of their counterpart controls under HSD conditions (Supplementary Fig. S5B, C). Accordingly, we focused on the regulatory role of p38 MAPK under HSD conditions. Western blot analysis with an anti-phospho-p38 MAPK antibody revealed that both mono-association of uracil-secreting bacteria and uracil supplementation increased the level of p38 MAPK phosphorylation in HSD-fed second-third instar (L2–L3) larvae (Fig. 6B and Supplementary Fig. S6). We next examined the metabolic phenotypes of p38a knockout (p38a13 ) larvae under HSD conditions. In contrast to uracil-fed w1118 flies, which were resistant to an HSD (Fig. 5A), uracil-fed p38a13 flies showed significantly higher levels of glucose, and lower levels of trehalose and triglycerides, in the hemolymph (Fig. 6C). Moreover, p38a13 flies showed a significantly longer time to LPF, and p38a-overexpressing flies (heat shock [HS]-Gal4; UAS-p38a) showed a significantly shorter time to LPF, than their counterpart controls under HSD conditions (Fig. 6D, E). Collectively, these results suggest that redox signaling mediated by bacterial uracil activates p38a MAPK to protect the host from the deleterious effects of an HSD.
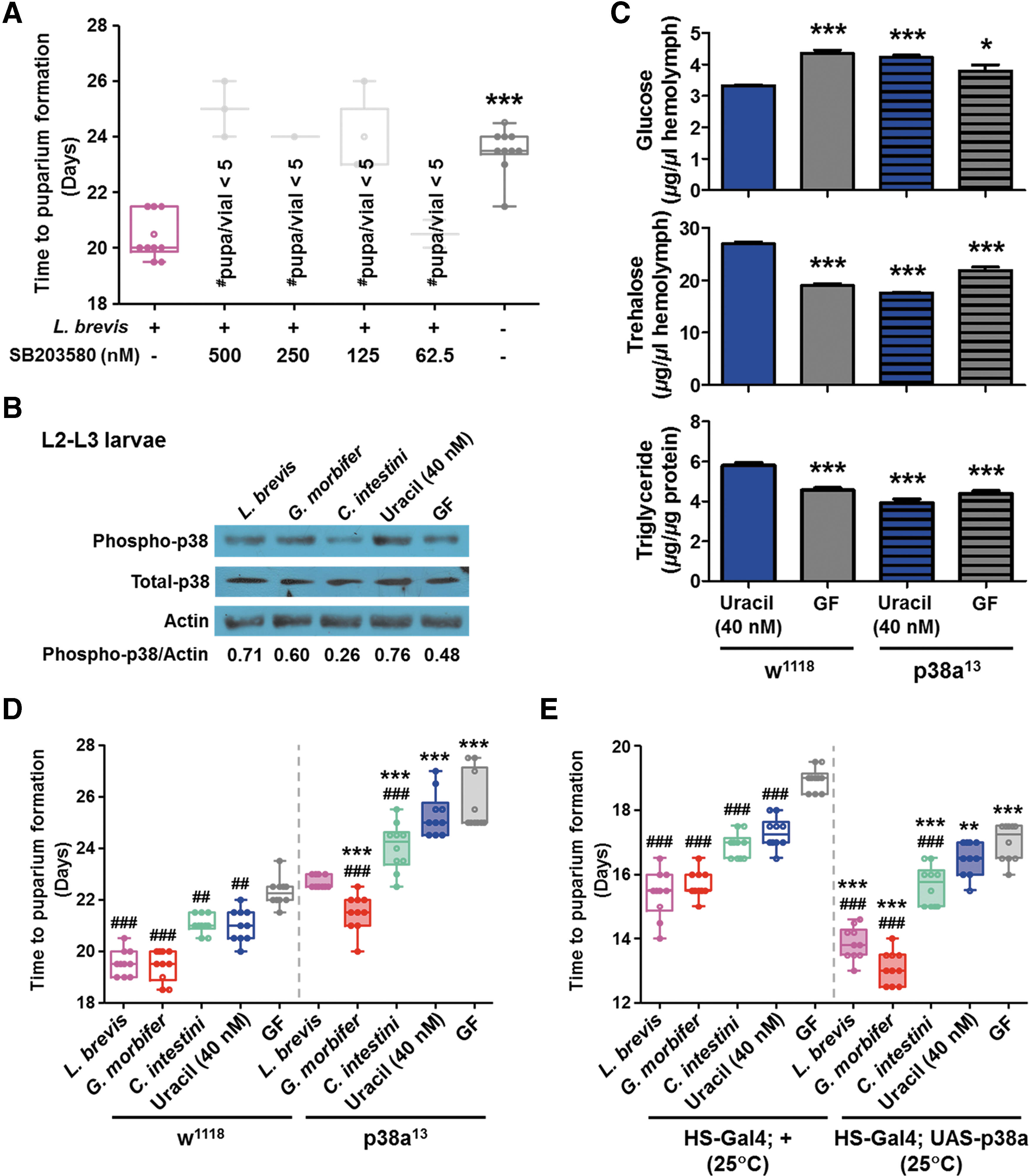
Because nuclear factor erythroid 2-related facter-2 (Nrf-2) and the Drosophila Forkhead transcription factor (dFOXO) are the main mediators of redox signaling (34, 69), we examined whether these genes are involved in the mechanism underlying the redox-dependent increases in host growth. There was no difference in the time to LPF between cncC (the Drosophila counterpart of Nrf-2)-overexpressing (Da-Gal4; UAS-cncC) and knock-down flies (Da-Gal4; UAS-cncC-RNAi) under ND-Con conditions. In contrast with the cncC-overexpressing flies (which represented a lethal phenotype under HSD conditions), the cncC knock-down flies showed a significantly shorter time to LPF than their counterpart controls (Supplementary Fig. S7A). Similarly, dFOXO-overexpressing flies (Da-Gal4; UAS-dFOXO) represented a lethal phenotype under HSD conditions (Supplementary Fig. S7B), whereas dFOXO-deficient flies (dFOXO21 ) showed a significantly shorter time to LPF than control flies (w1118 ) (Supplementary Fig. S7C). These results, when taken together with the weak fitness of the catalase-overexpressing flies under HSD conditions (Fig. 3E), suggest that activation of the antioxidant defense systems in response to ROS production may not be required for the bacterial uracil-mediated increase in the fitness of host flies on an HSD.
Bacterial uracil induces the production of both DUOX-specific and mitochondrial ROS
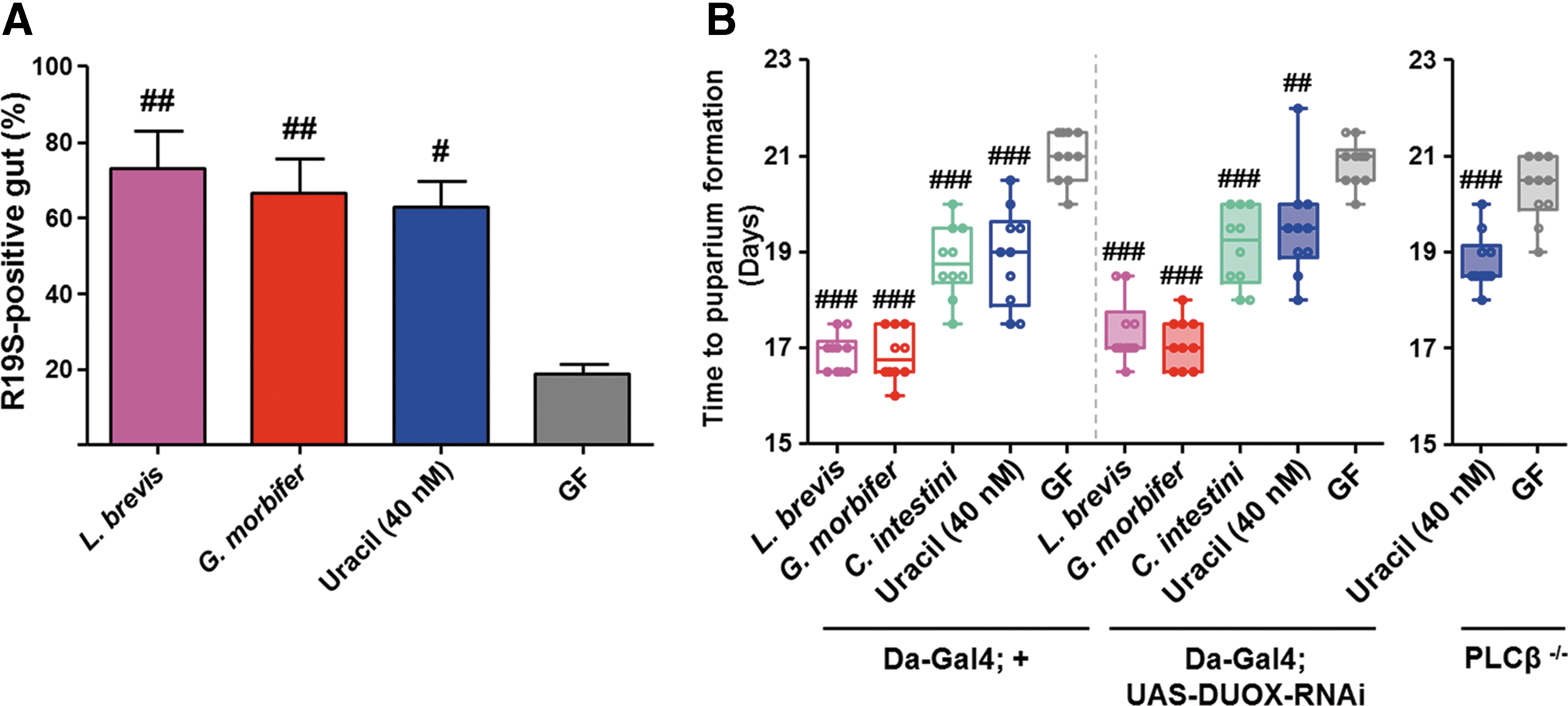
Under normal diet conditions, bacterial uracil elicits DUOX-dependent ROS generation in the Drosophila gut (42). Given that the increased growth of HSD-fed animals is dependent on redox signaling, and the fact that DUOX is the only known enzyme that regulates uracil-mediated ROS generation, we next measured in vivo ROS production in the gut of HSD-fed adult flies by using R19S (13) (a highly specific fluorescent dye that binds to hypochlorous acid [the production of which is DUOX dependent]). As expected, flies either mono-associated with uracil-secreting bacteria or supplemented with uracil generated significantly higher levels of DUOX-dependent ROS than GF controls (Fig. 7A), suggesting chronic activation of DUOX in the gut of HSD-fed animals. We hypothesized that feeding an HSD to DUOX function-impaired flies would abolish the redox-dependent increases in host growth. However, we observed no meaningful differences in the larval developmental period between DUOX-impaired flies (Da-Gal4; UAS-DUOX-RNAi) and their counterpart controls (Da-Gal4; +) (Fig. 7B). Likewise, feeding uracil to DUOX function-impaired flies of a different genotype (phospholipase C-β knockout, PLCβ−/− ) (25) resulted in increased larval growth, implying that there might be an alternative mechanism that is responsible for generation and/or transfer of the redox signal beyond DUOX-specific ROS production in the presence of bacterial uracil.
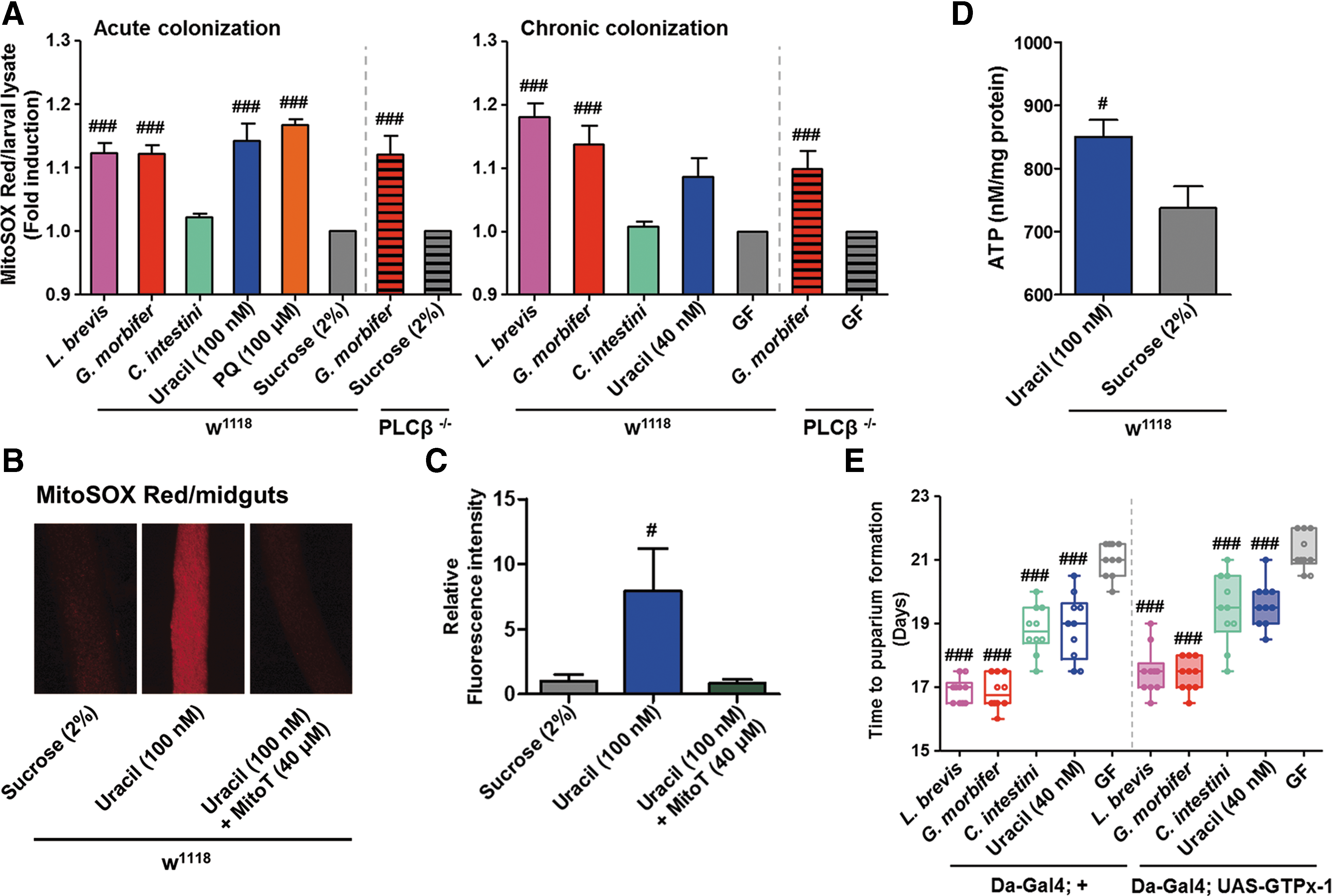
Although mitochondrial ROS production is tightly regulated, various stimuli (e.g., increased cytosolic calcium concentrations) can increase the levels and affect numerous cellular processes, including metabolic adaptation and immunity (63, 77). Accordingly, we examined whether levels of mitochondrial ROS are influenced by uracil-secreting bacteria in HSD-fed flies by using MitoSOX Red (41) (a selective fluorogenic dye that specifically reacts with mitochondria-generated superoxide) to measure mitochondrial superoxide levels in L3 larvae either acutely (i.e., CvR-Con L3 larvae infected with bacteria) or chronically (i.e., GF embryos infected with bacteria) colonized by uracil-secreting bacteria. Under conditions of acute colonization, superoxide levels in HSD-fed CvR-Con larvae after oral ingestion of L. brevis, G. morbifer, uracil, or paraquat were significantly higher than those in larvae fed a sucrose solution alone. Under chronic colonization conditions, the superoxide levels in HSD-fed larvae mono-associated with L. brevis or G. morbifer were significantly higher than those in the GF control (Fig. 8A). Interestingly, superoxide levels in PLCβ−/− flies after both acute and chronic colonization by G. morbifer were significantly higher than those in GF controls. Similar patterns were observed in acutely colonized ND-Con-fed insects (Supplementary Fig. S8), suggesting that bacterial uracil increases production of mitochondrial superoxide regardless of PLCβ−/− function [i.e., regulation of DUOX activity (25)] and sugar concentration in the diet. In vivo imaging of MitoSOX Red-treated midguts revealed significantly higher levels of superoxide in uracil-fed larvae than in larvae fed a sucrose solution alone or larvae fed uracil plus Mito-TEMPO (MitoT) (Fig. 8B, C). Because mitochondrial ROS (as a byproduct of oxidative metabolism) are generated during ATP production (24), we also asked whether bacterial uracil affects mitochondrial energetics. As expected, significantly more ATP was observed in uracil-fed larvae than in larvae fed a sucrose solution alone (Fig. 8D). Glutathione peroxidase I (GTPx-1) is an intracellular antioxidant enzyme that reduces H2O2 (mostly generated by mitochondria) (44, 48). Therefore, we next used transgenic flies ubiquitously expressing GTPx-1 (Da-Gal4; UAS-GTPx-1) and compared the time to LPF with that of their counterpart controls (Da-Gal4; +). We found no meaningful difference between the two different genotypes (Fig. 8E). Both uracil-secreting bacteria and supplementation with bacterial uracil provided larvae of GTPx-1-overexpressing flies with a growth advantage.
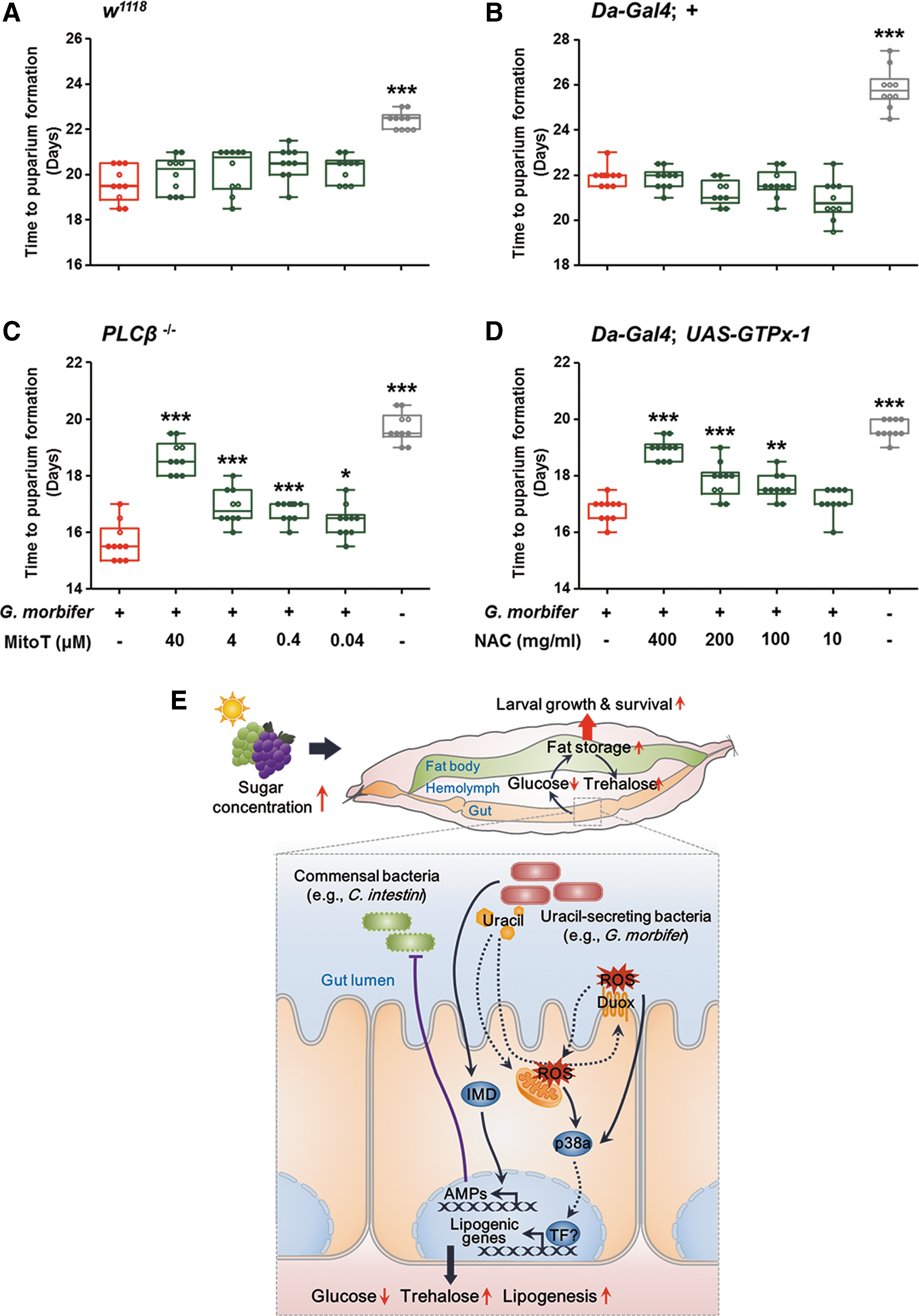
Finally, we generated gnotobiotic PLCβ−/−
flies harboring G. morbifer and treated them with different doses of MitoT to reduce mitochondrial ROS. In contrast to w1118
gnotobiotic controls (Fig. 9A), the PLCβ−/−
flies represented a significant dose-dependent delay in larval development by MitoT treatment (Fig. 9C). We then examined the growth of larvae from G. morbifer mono-associated GTPx-1-overexpressing flies fed different doses of N-acetyl-
The lifespan of adult flies is not affected by bacterial uracil under HSD conditions
We next examined the effects of bacterial uracil on the lifespan of adult flies under HSD conditions. To this end, we cultured GF larvae under ND-Con conditions and switched them to an HSD after the adults emerged (Supplementary Fig. S9A). Neither bacterial mono-association nor bacterial uracil affected the lifespan of the adult flies. However, uracil-fed adult flies showed higher survival rates than gnotobiotic or GF flies during the early days post-emergence, although the rate fell rapidly during the late days post-emergence (Supplementary Fig. S9B). Similarly, the survival of CvR-Con flies fed an HSD containing uracil (Supplementary Fig. S9A) was greater than that of HSD-fed CvR-Con flies in the early days post-emergence (Supplementary Fig. S9C).
Discussion
Obesity has long been regarded as the root cause of metabolic syndrome, which includes dyslipidemia, hypertension, heart disease, and diabetes mellitus (35, 45). Recent evidence shows that reducing the level of sugar in the blood by increasing fat storage is the key to regulating sugar homeostasis and, therefore, to protecting animals with excessive caloric intake from the onset of the diabetic phenotype (22, 38, 50). In this context, the present study demonstrates that flies mono-associated with conditionally pathogenic gut bacteria showed lower levels of glucose in the hemolymph and accumulated more stored fat than GF controls, resulting in increased larval growth under HSD conditions (Fig. 9E). Moreover, obese-prone mutants were more resistant to HSD-induced developmental arrest than lean mutants, suggesting that obesity per se is a systemic mechanism for alleviating the deleterious effects of an HSD (e.g., hyperglycemia). Another important metabolic feature in larvae harboring conditionally pathogenic bacteria was hypertrehalosemia. Interestingly, increased levels of trehalose increase the resistance of mosquitoes to desiccation or heat (43), and of cockroaches to oxidative stress (30). Therefore, it is easy to speculate that reducing the trehalose concentration in the hemolymph may render flies more susceptible to the effects of a chronic HSD. In the Drosophila gut, bacterial uracil is responsible for phosphorylating activating transcription factor-2 (ATF-2) via the DUOX-expression pathway, which comprises MEKK1-MKK3-p38 MAPK (39). Activation of ATF-2 positively regulates fat metabolism by inducing transcription of phosphoenolpyruvate carboxykinase (PEPCK) (51). Given that PEPCK regulates both biosynthesis of trehalose during gluconeogenesis and biosynthesis of triglycerides during glyceroneogenesis, redox signaling mediated by bacterial uracil may alter the metabolic status of the host by inducing expression of putative lipogenic genes, followed by activation of component transcription factor(s) via the DUOX-expression pathway.
We showed that redox signaling is induced by activation of DUOX and by increased levels of mitochondrial ROS. A previous study reported that chronically elevated glucose levels increase mitochondrial ROS production by increasing oxidative phosphorylation during anaerobic glycolysis (54). However, several recent studies demonstrate that high glucose does not stimulate endogenous ROS production in cultured cells (8), diabetic mice, or humans (19). In line with this, we showed that feeding an HSD to GF or CvR-Con insects did not increase mitochondrial ROS levels (Fig. 8A). Currently, we do not know whether receptors that bind bacterial uracil and increase mitochondrial ROS exist. If this is not the case, we propose a possible explanation for the increased levels of mitochondrial ROS induced by bacterial uracil. DUOX in the fly intestine is activated by bacterial uracil and, subsequently, generates extracellular H2O2. The H2O2 molecules might then cross the cellular membrane (5) and act as signaling molecules (16, 60) that stimulate mitochondrial ROS production. In addition, DUOX activation leads to the mobilization of calcium from the endoplasmic reticulum to the cytosol in a PLCβ-dependent manner (25); indeed, increased cytosolic calcium concentrations are linked to increased mitochondrial ROS levels (63). However, this explanation contains contradictions because DUOX function-impaired flies (PLCβ−/− ) may not allow calcium mobilization, and flies showing forced expression of GTPx-1 may scavenge intracellular H2O2. Therefore, the exact mechanism(s) underlying bacterial uracil-induced redox signaling remain to be elucidated.
We next manipulated the bacterial community in the fly gut by eliminating the established microbiota and introducing a defined microbiota. This approach, combined with 16S rRNA gene-based metagenomic analysis, enabled us to better define the composition of the bacterial community in the gut. It should be noted that feeding an HSD to flies harboring a defined gut microbiota resulted in the dominance of conditionally pathogenic bacteria (e.g., G. morbifer), which comprised only a small proportion of the microbiota in the gut of ND-Con-fed larvae (Fig. 2A). This abnormal community structure was, in part, a consequence of high levels of Relish-dependent AMP gene expression (Fig. 2B), suggesting that a high abundance of uracil-secreting bacteria leads to excessive production of AMPs, which may, in turn, eliminate AMP-susceptible bacteria (e.g., C. intestini). This argument is supported by the identification of distinct and specific AMP-dependent bacterial communities in polyps (21) and by fluctuations in the composition of bacterial communities that are induced by dysregulation of AMP production in the fly gut (53, 57). Moreover, predominant colonization by G. morbifer after colonization with an evenly mixed and defined microbiota is reminiscent of intestinal microbial dysbiosis, a condition whereby increased numbers of pro-inflammatory bacteria cause intestinal dysfunction and related diseases (67). Although the disruption of intestinal symbiosis is recognized as one cause of intestinal disease, the results presented here provide a new perspective, that is, that dysbiosis is likely to result from the host's attempts to protect itself against stress conditions.
The colitogenic condition occurring after DUOX-induced ROS production may happen regardless of the sugar concentration in the diet, and be equally harmful to the host. Given that the chronic oxidative stress that follows blooming of uracil-releasing bacteria reduces the host lifespan rather than causing mortality under normal diet conditions (42), it is likely that host protection mechanisms (e.g., activation of p38a MAPK) concomitant with uracil-mediated oxidative stress are vital for host survival under HSD-fed conditions. It is also worth mentioning that the extended larval developmental time caused by an HSD and mono-association with different bacterial species reflects increased susceptibility to the deleterious effects of dietary sugar; conversely, a shortened developmental time reflects increased resistance to such deleterious effects. Drosophila regulates the time of juvenile development according to dietary conditions, and when nutrients are scarce there is a pause in development until the larvae attain a critical weight (17, 32). Similar to nutrient scarcity, we found that chronic feeding of an HSD also had a marked effect on systemic growth, resulting in a delay in achieving this critical weight. Thus, the shortened period to pupation after egg deposition induced by bacterial mono-association is indicative of increased host fitness under HSD-fed conditions.
A fundamental question is: Why do flies harbor conditionally pathogenic autochthonous bacteria in the gut? Both autochthonous bacteria (e.g., L. brevis and G. morbifer) and allochthonous bacteria (e.g., Ecc15) are capable of inducing uracil-mediated ROS production in the Drosophila gut (42); however, only the former aids host development under HSD conditions (Fig. 4A). Gut-dwelling autochthonous bacteria are evolutionarily adapted to the host gut environment (59); therefore, our results are supported by the Lamarckian aspects of hologenome theory: (i) Evolution of the hologenome is regulated by the use and disuse of microbes; and (ii) changes in the hologenome are transmitted to offspring (56, 81). Accordingly, the host may carry (or easily acquire from the environment) autochthonous bacteria comprising both symbiotic and conditionally pathogenic microbes, and preferentially amplify the populations of specific bacterial groups, as appropriate. In nature, Drosophila utilizes fruits as an oviposition substrate; unsurprisingly, the sugar content of these fruits increases with ripening: for example, the sugar content of bananas ranges from 16.9 to 19.4 °Brix (47) and that in grapes ranges from 22 to 24 °Brix (46). Given that the flies preferentially oviposit on sugar-containing foods, and bacteria belonging to the family Acetobacteraceae are ubiquitous in sugar-rich environments (15, 61), larvae might often encounter sugar-rich conditions in nature and strike up a symbiotic relationship with coexisting uracil-secreting microbes such as G. morbifer. Interestingly, in mammals, an unstable microbial community often arises due to a sustained increase in abundance of the phylum Proteobacteria (64). This phylum, similar to the uracil-secreting bacteria in the fly gut, normally comprises only a minor proportion of the natural human gut flora; however, it becomes a colitogenic microbe that can trigger inflammatory responses under certain gut conditions. Indeed, a recent study by Kamdar et al. (36) reported a positive relationship between the outgrowth of an opportunistic commensal belonging to the Deltaproteobacteria family and increased ROS production by neutrophils in the intestine of Toll-like receptor 1 knockout mice. Thus, further studies of the ecological role(s) of enteric Proteobacteria are needed to determine the symbiotic and/or pathological relationships between the mammalian host and gut microbes.
Materials and Methods
Drosophila stocks and breeding
Drosophila w1118 were used as controls, unless otherwise specified. Fly stocks were maintained on cornmeal-agar medium containing 6% sucrose at 25°C/60% relative humidity. HSD was obtained by increasing the amount of sucrose to 30%. For all experiments, fly food was axenically prepared by autoclaving at 110°C for 20 min. Cornmeal-agar medium (ND-Con) contained 2% yeast (Lesaffre), 8% cornmeal (Sunglim Co.), 6% sucrose (Sigma, St. Louis, MO), 1% agar (Bacto, Mt. Pritchard, Australia), 0.032% tegocept, and 0.48% propionic acid. When necessary, uracil (Sigma), oxidants (H2O2 solution and paraquat dichloride; Sigma), antioxidants (NAC and MitoT; Sigma), or p38 and JNK inhibitors (SB203580 and SP600125, respectively; Sigma) were added to axenic media. Because the diets used for measurement of developmental time in CvR-Con flies contained tegocept and propionic acid, equivalent volumes of these reagents were added to the diet of mono-associated and GF flies in all remaining experiments. The following fly lines were used in the present study: Yellow White, CantonS , UAS-GTPx-1 (48), UAS-Catalase and UAS-DUOX-RNAi (26), UAS-cncC and UAS-cncC-RNAi (69), UAS-dFOXO (31), UAS-BSKDN (Bloomington Stock Center), UAS-Mio-RNAi (Vienna Drosophila RNAi Center [VDRC]), UAS-desat1-RNAi (VDRC), UAS-King-Tubby-RNAi (VDRC), PLCβ−/− (25), p38a13 (11), dFOXO21 (34), Da-Gal4 (23), UAS-Puckered (gift from Dr Cho KS, Konkuk University, Seoul, Republic of Korea), and UAS-p38a and HS-Gal4 (gifts from Dr. Lee W.-J., Seoul National University, Seoul, Republic of Korea). Transgenic flies in all assays were generated by crossing virgin females from the specified Gal4 strains with males from each UAS strain. This experimental design does not necessarily ensure genetic homogeneity. To verify the target transcript levels of these transgenic flies, CvR-Con transgenic larvae (L3) fed an ND-Con were subjected to quantitative PCR analysis (Supplementary Fig. S10). The primer sets are listed in Supplementary Table S4.
Bacterial strains
To obtain candidate taxa for bacterial mono-association, Acetobacter pasteurianus ssp. pasteurianus, A. ursingii, L. brevis, L. pentosus, and L. pseudomesenteroides were isolated from the gut of CvR-Con flies. Several bacterial strains isolated from the gut of Drosophila [A. pomorum (65), C. intestini, G. morbifer (55), L. plantarum ssp. plantarum (37), G. morbifer-carA::Tn5 (42), and E. faecalis (68)] were then added. The following bacterial strains were used for mono-association of non-resident allochthonous bacteria: E. carotovora ssp. carotovora 15, Psedomonas entomophila, Serratia marcescens, Escherichia coli K12, Micrococcus luteus, Orbus sasakiae (isolated from Sasakia charonda), P. rettgeri (isolated from P. quadriguttata), and Providencia alcalifaciens (isolated from Ampelophaga rubiginosa).
Mono-association of GF insects
GF insects were generated as described by Ryu et al. (57). Drosophila embryos were collected on egg-collecting agar (3.4% glucose and 3% agar) for 4 h and then dechorionated by exposure to sodium hypochlorite (2.7%) for 2 min. Embryos were subsequently washed three times in 70% ethanol, followed by one wash with sterile distilled water. The embryos were than maintained on an axenic diet. Bacterial contamination was checked by spread plating insect homogenates and food on bacterial culture media (GF controls in Supplementary Fig. S3). To generate gnotobiotic insects, A. pomorum, A. pasteurianus ssp. pasteurianus, C. intestini, and G. morbifer were cultured in mannitol broth; L. plantarum ssp. plantarum, L. brevis, L. pentosus, L. pseudomesenteroides, and E. faecalis were cultured in Lactobacilli MRS broth; A. ursingii and O. sasakiae were cultured in nutrient broth and tryptic soy broth, respectively; E. carotovora ssp. carotovora 15, S. marcescens, E. coli K12, M. luteus, P. rettgeri, and P. alcalifaciens were cultured in Luria broth; and P. entomophila was cultured in Luria broth containing rifampicin (100 μg/ml). A pellet of exponentially cultured bacterial cells (OD600 = 3; 50 μl) was washed with phosphate buffered saline (PBS), re-suspended in 5% sucrose solution, and added to food vials containing GF embryos after 1 day of embryonic dechorionation.
Measurement of time to puparium formation
Embryos (approximately 30–40 eggs) were deposited on sterile food by gentle pipetting. The developmental period of individual insects raised under different dietary conditions was measured by counting the number of pupae over time (10 biological replicates). After the emergence of the first pupation, the number of pupae was counted every 24 h. After the emergence of the last pupation, the mean puparium formation periods were calculated and compared with respect to different treatments (i.e., fly vials mono-associated with different bacteria). Vials containing <15, or >60, pupae were excluded.
Multiplex 454 pyrosequencing of bacterial 16 rRNA genes
The experimental samples comprised whole bodies of L1 (n > 30) and L2–L3 larvae (n = 20), and dissected guts (from proventriculus to rectum) from L3 wandering larvae (n = 20) and 3- to 5-day-old adult females (n = 20). Surface washing and gut dissection were conducted as described by Wong et al. (78), with minor modifications. Briefly, all samples were surface washed with 100% ethanol instead of sodium hypochlorite solution, followed by three rinses with sterile distilled water. Gut dissection was performed under a dissecting microscope, with samples placed in sterile Ringer's solution on clean glass slides and dissected by using sterilized forceps. Bacterial genomic DNA was extracted according to the Repeated Bead Beating plus column method (80). For gut microbiota isolated from ND-Con-fed CvR-Con flies (Supplementary Fig. S1A), the portion of the bacterial 16S rRNA gene spanning hypervariable regions V1–V4 was amplified by PCR by using bacterial universal primers 8F (5′-X-GAG TTT GAT CCT GGC TCA G-3′) and 800R (5′-TAC CAG GGT ATC TAA TCC-3′). For sets of defined gut microbiota (Fig. 2A) and the bacterial communities within fly food (Supplementary Fig. S1C), the portion of the 16S rRNA gene spanning hypervariable regions V1–V2 was amplified by PCR by using bacterial universal primers 8F (5′-X-GAG TTT GAT CMT GGC TCA G-3′) and 338R (5′-TGC TGC CTC CCG TAG GAG T-3′). To tag each PCR product, a 10-base sample-specific barcode (designated “X”) was added to the 5′ ends of the primer sequences (27). PCR analyses were performed in a C 1000 Thermal Cycler (Bio-Rad, Hercules, CA). The PCR conditions were as follows: initial denaturation at 96°C for 6 min, followed by 20 cycles (DNA extracted from dissected guts) or 25 cycles (DNA extracted from whole bodies) of denaturation at 94°C for 1 min, annealing at 50°C for 30 s, and extension at 72°C for 90 s. A final extension step was performed at 72°C for 10 min. Triplicate reactions were pooled and purified by using the QIAquick PCR purification kit (Qiagen, Valencia, CA). The purified amplicons were combined in a single tube at equimolar ratios (final concentration, 5 μg). The amplicons were pyrosequenced by Macrogen (Seoul, Republic of Korea) by using a Genome Sequencer FLX plus (Roche, Branford, CT) or FLX Titanium (Roche).
Sequence analysis and community comparison
Raw sequences were pre-processed by using the QIIME software package 1.5.0 (9) to exclude poor-quality sequences and/or sequencing errors. Briefly, raw sequences containing more than one ambiguous base call, those with errors in the barcode or primer regions, those with average quality scores <25, or those shorter than 200 bp in length, were removed. Subsequently, the remaining sequences were denoised and the reverse primer sequences were trimmed. Reference-based operational taxonomic units (OTUs) were clustered against the Greengenes-provided reference sequences at 99% sequence similarity by using UCLUST software. A representative sequence for each OTU was selected and aligned with the Greengenes-provided reference sequences by using PyNAST. Chimeric sequences were excluded from the aligned representative sequences by using ChimeraSlayer. A phylogenetic tree of the aligned sequences was then constructed by using Clearcut. Taxonomic assignment of representative sequences at the species level was performed by copying and pasting each query sequence into the EzTaxon program. The program then returned the type strain showing the best match with the query sequence (>98% similarity). The query sequences representing <98% sequence similarity were categorized as “unclassified.” For gut microbiota from ND-Con-fed CvR-Con flies, sequences assigned as Wolbachia were subsequently subtracted from the OTU table. A recent metagenomic study by Seedorf et al. (62) reported that colonization of GF animals by defined bacterial communities often yields bacterial communities that contain unexpected bacterial sequences. This is not the result of contamination by other bacteria. These unexpected sequences may simply represent DNA fragments derived from other sources, such as autoclaved food, rather than from viable contaminating bacteria. Indeed, a study by Hill et al. (29) reported that these unexpected bacterial communities might be derived from contamination by bacterial 16S rDNA usually present in autoclaved food. Accordingly, sequences not assigned to input bacterial communities were subsequently subtracted from the OTU table listing sets of defined gut microbiota and the bacterial species in fly food.
Wolbachia removal
Sequence analysis revealed that CvR-Con flies possessed sequences assigned to the genus Wolbachia, a maternally transmitted intracellular endosymbiont. Accordingly, the presence of Wolbachia was confirmed by PCR with Wolbachia-specific primers (10, 33). To exclude the possibility that insect endosymbionts may affect host metabolism and/or host fitness, Wolbachia were removed from CvR-Con flies by treatment with tetracycline (50 μg/ml) over three generations (6). To help the flies re-colonize their resident microbiota, a mixture of cultured bacteria previously isolated from the gut of parental CvR-Con flies was then re-introduced.
ROS measurement
Hypochlorous acid production (i.e., ROS generated by DUOX) was measured as described by Lee et al. (42) by using the hypochlorous acid-specific rhodamine-based dye, R19S (FutureChem, Seoul, Republic of Korea). GF insects were mono-associated with live bacteria (L. brevis or G. morbifer) and maintained on an HSD, whereas GF insects were fed an HSD containing uracil (40 nM). Adult female flies (5–7 days old) were fed a sucrose solution (5%) containing R19S (10 μM) for 90 min. The midguts were then dissected and fixed in paraformaldehyde (4%). Hypochlorous acid production was visualized under an epifluorescence microscope (Eclipse Ni-U equipped with a fluorescence filter; Nikon). ROS production was measured in R19S-positive guts (n = 20) in three independent experiments, and the number of R19S-positive guts per total number of gut samples was expressed as a percentage.
The mitochondrial superoxide indicator MitoSox Red (Life Technologies, Carlsbad, CA) was used to measure mitochondrial ROS production. Mitochondrial ROS was measured under two different experimental conditions (chronic and acute colonization). For chronic colonization, GF insects of different genotypes were mono-associated with live bacteria (L. brevis, G. morbifer, or C. intestini) and maintained on an HSD, whereas fed an HSD containing uracil (40 nM). For acute colonization, CvR-Con animals of different genotypes were reared on HSD, and L3 larvae were fed a sucrose solution (2%) containing live bacteria (L. brevis, G. morbifer, or C. intestini), uracil (100 nM), or paraquat (100 μM) for 90 min. Age-matched larvae (10 L3 larvae) were homogenized in 300 μl of PBS containing MitoSox Red (5 μM) by using a bead beater (FastPrep; MP Biomedicals). The homogenates were then incubated in the dark at room temperature for 30 min and measured with a spectrophotometer (Synergy MX; BioTek) at an excitation wavelength of 510 nm and an emission wavelength of 580 nm. Each bar graph shows the mean values of replicate samples (two biological plus two technical replicates). For microscopic images, L3 larvae were fed a sucrose solution (2%) containing uracil (100 nM), or uracil and MitoT (40 μM) for 6 h. After surface washing twice in PBS, the midguts were dissected and treated with PBS containing MitoSox Red (5 μM) for 20 min. The dissected guts were mounted on Marienfeld slide glass, and they were imaged under an epifluorescence microscope (Eclipse 50i equipped with fluorescence filter; Nikon) at 80 × magnification. Each bar graph shows the mean values of replicate samples (10 biological replicates).
Adult survival assays
GF larvae were cultured under ND-Con conditions. After the adults emerged, GF flies were mono-associated with L. brevis, G. morbifer, or C. intestini and fed an HSD, whereas GF flies were fed an HSD containing uracil. CvR-Con flies were fed an HSD or an HSD containing uracil after adult emergence. Flies were then transferred to fresh axenic media, and dead flies were counted every 3 days.
Measurement of sugar levels in the hemolymph
Bacterial mono-association and uracil supplementation were carried out under HSD conditions as described earlier. CvR-Con animals reared on an ND-Con were included as a control. Animals were moved to an empty plate for 1 h before hemolymph extraction. Hemolymph (2 μl) was collected from L3 larvae (5–10 larvae fed an ND-Con and 20–30 larvae fed an HSD) and diluted in 8 μl of sterile distilled water. To measure the level of glucose in the hemolymph, 1.25 μl of diluted hemolymph (eight technical replicates) was mixed with 18.75 μl of sterile distilled water and placed at 4°C until the next step. To measure the level of trehalose, 1.25 μl of diluted hemolymph (eight technical replicates) was mixed with 2 μl of porcine trehalase (Sigma) and 16.75 μl of sterile distilled water and incubated at 37°C for 2 h. The total amount of glucose and trehalose (glucose digested by trehalase) in the samples was measured simultaneously by adding 130 μl of Glucose Assay Reagent (Sigma). NADH was then measured at 340 nm, according to the manufacturer's instructions. A series of glucose standards (Sigma) was used to construct a standard curve for each experiment. Glucose levels were then calculated from these standard curves.
Measurement of systemic carbohydrate levels
Whole larvae (five L3 larvae; eight biological replicates) were homogenized in 300 μl of PBS. Next, 10 μl of the homogenate was incubated at 65°C for 15 min with an equal volume of starch assay reagent containing amyloglucosidase (Sigma). Subsequently, 7 μl of the reaction mixture was removed and added to Glucose Assay Reagent to measure glucose levels as described earlier. Homogenate incubated with an equal volume of PBS was run in parallel, and the values were subtracted to calculate free glucose levels. For normalization, protein concentrations were measured by using the BCA Protein Assay (Lamda Biotech, St. Louis, MO), according to the manufacturer's instructions. Standard curves were then constructed for each experiment.
Measurement of triglyceride levels
Whole larvae (five L3 larvae; eight biological replicates) were homogenized in 300 μl of PBS containing 0.1% Tween 20. The homogenates were then incubated at 65°C for 5 min to remove lipase activity. After centrifugation (8000 rpm at room temperature for 10 min), 30 μl of supernatant was mixed with an equal volume of Triglyceride Reagent (Sigma) and incubated at 37°C for 30 min. Half of the reaction mixture was then incubated with 100 μl of Free Glycerol Reagent (Sigma) for 5 min at room temperature and measured with a spectrophotometer at 540 nm according to the manufacturer's instructions. Protein concentrations were measured as described earlier and used to normalize the data. Standard curves were generated for each experiment.
Lipid droplet staining
Lipid droplet staining of larval fat bodies was performed as described by Musselman et al. (50). Briefly, L3 larvae were fixed in 4% paraformaldehyde for 20 min, followed by treatment with PBS containing 0.001% Nile Red (Sigma) for 30 min. After washing twice in PBS, fat bodies were dissected and mounted on Marienfeld slide glass. Samples were imaged under an epifluorescence microscope (Eclipse 50i equipped with fluorescence filter; Nikon) at 200 × magnification.
ATP measurements
ATP measurements were performed as described by Tennessen et al. (71). Five L3 larvae were fed a sucrose solution (2%) containing uracil (100 nM) for 6 h. After surface washing twice in PBS, the larvae were homogenized in homogenization buffer (6 M guanidine HCl, 100 mM Tris [pH 7.8] and 4 mM EDTA). The diluted homogenates were mixed with a luminescent solution (Molecular Probes), and luminescence was measured with a spectrophotometer. Protein concentrations were measured as described earlier and used to normalize the data. Standard curves were generated for each experiment. Each bar graph shows the mean values of replicate samples (five biological plus three technical replicates).
Real-time quantitative PCR
For the quantitative analyses of G. morbifer in the gut of ND-Con or HSD-fed flies (Fig. 2A right), DNA was prepared from the whole bodies of L2–L3 larvae and 3- to 5-day-old female adults as described earlier. Samples were analyzed in five biological plus two technical replicates. For the quantitative analyses of Relish-dependent AMP genes (Supplementary Fig. S2), RNA was prepared from the whole bodies of L2–L3 larvae and dissected guts from L3 wandering larvae and 3- to 5-day-old female adults by using TRIzol Reagent (Life Technologies). Complementary DNA (cDNA) was then synthesized by using the Maxima First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, MA). Samples were analyzed in two biological plus two technical replicates. The primer sets are listed in Supplementary Table S4. PCR was performed in a reaction volume of 25 μl containing 12.5 μl of SYBR Premix Ex Taq (Takara, Shiga, Japan), 10 pmol each of the forward and reverse primers, and 2 μl of fly cDNA (<100 ng) using a CFX96™ Real-Time PCR Detection System (Bio-Rad). Values are presented in terms of relative expression.
Western blot analysis
To detect phosphorylated p38, live L2–L3 larvae were homogenized on ice in RIPA buffer (Thermo Scientific) containing Protease and Phosphatase Inhibitor Cocktail (Thermo Scientific). Samples were loaded onto sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels, transferred to PVDF membranes, and detected by Western blotting with a goat polyclonal p38 (dN-20) antibody (Santa Cruz Biotechnology, Inc.) and a rabbit monoclonal phospho-p38 MAPK antibody (Cell Signaling Technology). An anti-actin antibody (Sigma) was used as a loading control.
Nucleotide sequence accession numbers
The bacterial sequences derived from the gut of flies and from fly food have been deposited in the European Nucleotide Archive and are available under accession number PRJEB10235.
Statistical analysis
Statistical analyses were performed by using GraphPad Prism version 5.0 for Windows (GraphPad Software). Comparisons between two samples were made by using the non-parametric Mann-Whitney U test (two-tailed) and Student's unpaired t-test (one-tailed). Comparisons between multiple samples were conducted by analysis of variance (ANOVA) with Tukey's post hoc test (*p < 0.05, **p < 0.005, and ***p < 0.001; N.S. denotes not significant). The lines, boxes, scattered dots, and whiskers in the box plot diagrams represent the median, first and third quartiles, replicates, and min-to-max distribution of replicate values, respectively. Values in bar graphs represent the mean ± standard error of the mean (SEM).
Footnotes
Acknowledgments
The authors thank Dr. W.-J. Lee and Dr. S.-H. Kim (Seoul National University) for help with in vivo experiments, and for providing the fly stocks. They also thank Dr. S. Hyun (Chung-Ang University, Seoul, Republic of Korea), Dr. K.S. Cho (Konkuk University), Dr. W. Sullivan (University of California, Santa Cruz, CA), Dr. B. Lemaitre (EPFL, Lausanne, Switzerland), Dr. E. Owusu-Ansah (Columbia University College of Physicians & Surgeons, New York, NY), Dr. D. Bohmann (University of Rochester), Dr. E. Hafen (ETH Zurich, Zurich, Switzerland), and Dr. S. Ishii (RIKEN Tsukuba Institute, Ibaraki, Japan) for providing fly stocks. This work is supported by the Mid-career Researcher Program (NRF-2016R1E1A1A02921587 to J.-W.B.) and the Collaborative Genome Program for Fostering New Post-Genome industry (2015M3C9A2054299 to J.-W.B.) through the National Research Foundation of Korea (NRF) funded by the Ministry of Science ICT and Future Planning, and the Basic Science Research Program (2015R1D1A4A01019390 to T.W.W.) through the NRF funded by the Ministry of Education.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.