
Abstract
Significance:
Erythrocyte hemoglobin (Hb) and myocyte myoglobin, although primarily oxygen-carrying proteins, have the capacity to do redox chemistry. Such redox activity in the wider family of globins now appears to have important associations with the mechanisms of cell stress response. In turn, an understanding of such mechanisms in vivo may have a potential in the understanding of cancer therapy resistance and neurodegenerative disorders such as Alzheimer's.
Recent Advances:
There has been an enhanced understanding of the redox chemistry of the globin superfamily in recent years, leading to advances in development of Hb-based blood substitutes and in hypotheses relating to specific disease mechanisms. Neuroglobin (Ngb) and cytoglobin (Cygb) have been linked to cell protection mechanisms against hypoxia and oxidative stress, with implications in the onset and progression of neurodegenerative diseases for Ngb and cancer for Cygb.
Critical Issues:
Despite advances in the understanding of redox chemistry of globins, the physiological roles of many of these proteins still remain ambiguous at best. Confusion over potential physiological roles may relate to multifunctional roles for globins, which may be modulated by surface-exposed cysteine pairs in some globins. Such roles may be critical in deciphering the relationships of these globins in human diseases.
Future Directions:
Further studies are required to connect the considerable knowledge on the mechanisms of globin redox chemistry in vitro with the physiological and pathological roles of globins in vivo. In doing so, new therapies for neurodegenerative disorders and cancer therapy resistance may be targeted. Antioxid. Redox Signal. 26, 763–776.
Introduction
T
The mechanisms of the redox activity of globins have relevance in bacteria, plants, and animals and the understanding of proteins from one class of globins may inform on the redox functions of others. This may be of particular relevance to the understanding between potential links between the redox activity of vertebrate globins and mechanisms of disease and cell protection. For example, there has been a realization of the importance of redox chemistry of cytoglobin (Cygb) in cancer therapy resistance and neuroglobin (Ngb) in protection against neurodegenerative diseases. However, the mechanisms that control possible redox functions of these globins remain unclear. Potential multifunctional roles of globins may provide an answer to conflicting reports concerning redox functions of globins. Mechanisms that can modulate the activity of globins in vivo, such as oxidation of cysteines, will be explored. In addition, true multifunctional globins such as dehaloperoxidase (DHP) will be examined to give insights into the potential mechanisms of protein function modulation in vivo.
Peroxidase Activity of Hb and Myoglobin in Disease
During the pioneering work on the mechanisms of peroxidase activity conducted during the 1950s and 60s, myoglobin (Mb) and Hb provided an ideal model system for study. The ease of obtaining significant quantities of the protein and the slow reaction between H2O2 and these respiratory heme proteins provided an opportunity to follow the reaction in detail (35, 36, 57). The reaction between Mb or Hb and H2O2 was termed (and frequently still is) a pseudoperoxidase activity as it was not considered that these reactions were relevant in vivo. However, many now consider globins to be true peroxidases with specific biological roles for this activity. This reaction generates a hypervalent ferryl heme iron from both ferrous and ferric oxidation states with a protein-based radical if initiated from the ferric state:
where P represents protein and P•+ a protein cationic radical. For peroxidases such as ascorbate peroxidase and catalase, this radical is stabilized on the porphyrin or nearby amino acid. For Mb and Hb, this reactivity occurs mainly under uncontrolled pathological conditions with the radical migrating throughout the protein or onto unintended substrates, with the ferryl (equivalent to compound II) also able to initiate oxidation of substrates such as lipids. For reviews on this subject, see Refs. (86, 97). The reaction between Mb or Hb and lipid peroxides also generates ferryl heme iron and radical species. However, being a one-electron acceptor, the radical products of lipid-based peroxides are the lipids themselves, leading to a potential cascade of lipid oxidation reactions and potent cell signaling molecules (49, 68, 96).
The redox chemistry of Mb or Hb is suppressed within the myocyte and erythrocyte, respectively, and generally only becomes apparent following myocyte injury and/or hemolysis. This is particularly relevant for erythrocytes due to the constant process of hemolysis in the blood stream. There are numerous systems for dealing with this undesired redox chemistry in the blood plasma, including haptoglobin (Hp) and hemopexin (24, 98). These proteins sequester free Hb and free heme, respectively, providing a defense against Hb-mediated oxidative damage during intravascular hemolysis. The binding of alpha beta dimers of Hb to Hp also appears to inhibit the damaging effects of Hb peroxidase activity, not by preventing the primary reactivity of the globin with peroxidase, but by limiting the reactivity of the high oxidation state of the protein (17). In this study, the ferryl heme iron is stabilized, increasing the steady state of ferryl heme iron. Ferryl stabilization is accompanied by an increase in the amount of the peroxide-induced tyrosine free radical on the beta chain (βTyr145). Thus, Hp may function to protect against vasculature damage by preventing oxidation of species that initiate undesirable free radical chain reactions without altering the reactivity of Hb with small exogenous ligands such as O2, nitric oxide (NO), and H2O2 (17). Extensive hemolysis can overwhelm the Hp and heme-sequestering proteins and Mb, released from muscle damage (rhabdomyolysis), cannot be sequestered by Hp, leading to kidney damage (68). The redox activity of alpha Hb can also be suppressed when bound to its chaperone protein, alpha Hb-stabilizing protein (67). The decrease in peroxidase activity results from the change in heme coordination from penta to hexa when alpha Hb is bound to the chaperone protein (30, 44). Thus, the chaperone prevents oxidation of the protein, and hence protein destabilization, before formation of α2β2 or α2γ2 tetramers.
The high oxidation state of iron (Fe4+) is difficult to detect directly in vivo due to the transient nature of hypervalent ferryl heme iron and protein-based radical generated concurrent with ferryl heme. In addition, the reactions they can potentially promote (that of free radical-mediated oxidation) can be sourced from a variety of alternative reaction systems such as Fenton chemistry or cyclooxygenase enzymes. Nonetheless, isolated ischemic rat hearts showed significant oxidation of Mb to the ferryl state by residual H2O2 (3). The ferryl heme iron and protein radical are highly oxidizing compounds, damaging proteins, DNA, and small molecules; however, lipids are particularly vulnerable to oxidation. This pro-oxidant activity of Hb is linked to a number of disease states, including atherosclerosis (84). The formation of ferryl Mb or Hb may not be preventable, with ferryl Hb-associated free radical observable in normal blood by EPR spectroscopy (110). However, the ferryl iron and the protein radical can be safely dealt with in the environment of the myocyte or erythrocyte by various electron-donating antioxidant agents such as ascorbate and urate (18). It is of note that iron chelators such as desferrioxamine and deferiprone, ameliorators of oxidative damage, can efficiently reduce the ferryl heme iron to the ferric iron in addition to their prevention of Fenton chemistry by labile iron (16, 90). Therefore, it is difficult to determine the mechanism of any ameliorative effects obtained from the use of such chelators in vivo, that is, whether the mechanism of antioxidant effects of these chelators is by donation of electrons to the high oxidation states of Hb or by preventing the Fenton chemistry of nonheme iron.
A covalent link between the heme and protein is formed from heme undergoing a ferric–ferryl redox cycle under acidic conditions (14, 88, 94). Although noted as a stable green-colored by-product of ferric–ferryl redox cycling over half a century ago, it was considered primarily as a curiosity with no obvious biological relevance (35). However, as a stable and specific product of Mb/Hb redox cycling, it had the potential to fulfill the role as a marker of their previous redox chemistry in vivo. Heme to protein cross-linked Mb has been detected following rhabdomyolysis (muscle damage) in the kidney of animal models and urine of human patients (48, 49, 68). Such covalent modifications of Hb have also been detected following subarachnoid hemorrhage in the cerebrospinal fluid (91). This is direct evidence of the involvement of these globins in redox chemistry, which appears to occur whenever the globin becomes separated from its normal reductive environment. Potent vasoactive lipid oxidation products, such as isoprostanes, are generated during acute kidney injury following rhabdomyolysis and during delayed vasospasm in the cerebrospinal fluid. Isoprostanes can be formed in vitro by the peroxidase activity of Mb/Hb on cell tissue or low-density lipoproteins (8, 68, 92). Hence, globin redox chemistry is a likely origin of these vasoactive compounds in vivo, particularly when combined with the evidence that globins undergo redox cycling by cross-linking in the same disease tissue. It should be noted that Fenton chemistry can also lead to production of vasoactive products (130, 131) and hence the significance of the finding of such heme to protein cross-links in vivo is debatable.
Other covalent modifications are noted in human Hb peroxidase redox activity. Both adult and fetal Hb forms show extensive oxidative modifications to α and beta/gamma chains, although the β/γ chain is more prone to oxidative modifications compared with α chain (55). In a variant of Hb (γ-Val67 to Met, Hb Toms River), incorporation of labeled oxygen from H2 18O2 to the methionine converts the amino acid into asparagine (106). Irreversible oxidation of β Cys93 occurs; up to two-thirds of β Cys93 is oxidized with ∼10-fold excess of H2O2 (55). This modification is even more extensive in sickle cell Hb. Other hotspots of amino acid modifications include β Trp15, Trp37, Met55, and Cys112, as well as α Tyr24, Met32, Tyr42, Met76, and Cys104 (55). These structural instabilities from such modification can lead to heme loss and oxidative damage to mitochondria.
Ferryl Hb also has important reported effects on inflammation and cell adhesion molecules. Heme released from damaged globins is a powerful pro-oxidant, qualifying it as a danger-associated molecular pattern molecule, increasing the activation of Nrf2 and HO-1 (125). However, Hb itself has reported proinflammatory effects, targeting vascular endothelial cells and inducing the expression of proinflammatory genes in endothelial cells, including activation of the NF-κB family of transcription factor cell adhesion molecules such as E-selectin, Icam-1, and Vcam-1 (104). This leads to effects on the actin cytoskeleton, disrupting the endothelial cell monolayer. Furthermore, actin filament length is also affected in red blood cells, with ferryl Hb decreasing the actin polymerization in cells infected with Plasmodium falciparum malaria in vitro (20). This effect is enhanced with sickle cell Hb S or Hb C, potentially preventing the parasite from creating its own actin cytoskeleton within the host cell cytoplasm, giving a possible mechanism of protection against the malarial parasite. The ferryl form of Hb S also induces oxidative stress in lung epithelial cells over that of HbA, attributed to the longer lived ferryl species of Hb S (55). However, both HbA and HbS induce HO-1 and superoxide production with impairment of mitochondrial oxygen consumption.
The redox chemistry of Hb is also relevant in the development of Hb-based oxygen carriers (HBOCs) used as potential blood substitutes or oxygen therapeutics. Numerous attempts have been made at generating a stable nontoxic form of cell-free Hb with limited success to date. However, these studies have been focusing on preventing kidney damage (through increasing the size and stability of the protein structure by polymerization, encapsulation, or pegylation) and preventing scavenging of NO (through site-directed mutagenesis) (2, 119). These aspects of HBOC chemistry are undoubtedly important factors in generating safer HBOCs; however, the peroxidatic activity of Hb and HBOCs has only recently been considered to be a potential factor in taming HBOCs (1, 2). One strategy to limit the redox chemistry of HBOCs in vivo is to enhance the ability of naturally occurring antioxidants in the blood to remove the cytotoxic ferryl form of the Hb by introducing electron transfer pathways to the engineered HBOC (vide infra).
Through-Protein Electron Transfer Pathways
Through-protein electron transfer pathways (TPETPs) have the ability to accelerate reduction of the ferryl oxidation state of globins, enhancing the safer removal of cytotoxic forms of Hbs by physiological concentrations of antioxidants such as ascorbate, urate, or even low concentrations (micromolar) of iron chelators such as deferiprone. We first reported these TPETPs to be naturally occurring in Mb of some species (87), noting the potential implications for therapeutic intervention in oxidative stress and for engineering more effective blood substitutes. These pathways incorporating redox-active tyrosines (Tyr103 in human/horse/sperm whale Mb) are able to pass electrons from the reductant to the heme iron more efficiently than direct access to the heme pocket (Fig. 1). Human Hb alpha subunit contains such a TPETP with a redox-active tyrosine (α-Tyr42), but the beta subunit equivalent position is occupied by a redox-inactive phenylalanine (β-Phe41) and hence has no TPETP. The positions of Tyr42 in Mb and Tyr103 in α Hb are in similar spatial positions (even though from different helices) and thus appear to perform similarly with regard to rates of electron transfer. Mutagenesis can introduce such pathways into the beta chain or remove them from the alpha chain, affecting the kinetics of electron donation of the reductant to the heme iron accordingly (89). Substitution with other redox-active residues such as tryptophan does not show TPETP activity (89). At physiological concentrations of ascorbate (∼50 μM), proteins with TPETPs can effectively enhance ferryl iron reduction up to 12-fold over those without the pathway (89). Other serum antioxidants such as uric acid as well as phenols, hydroxyurea, and iron chelators such as hydroxypyridinone CP20 (deferiprone) can also act as ferryl reductants utilizing the TPETP (18, 87, 89, 90).
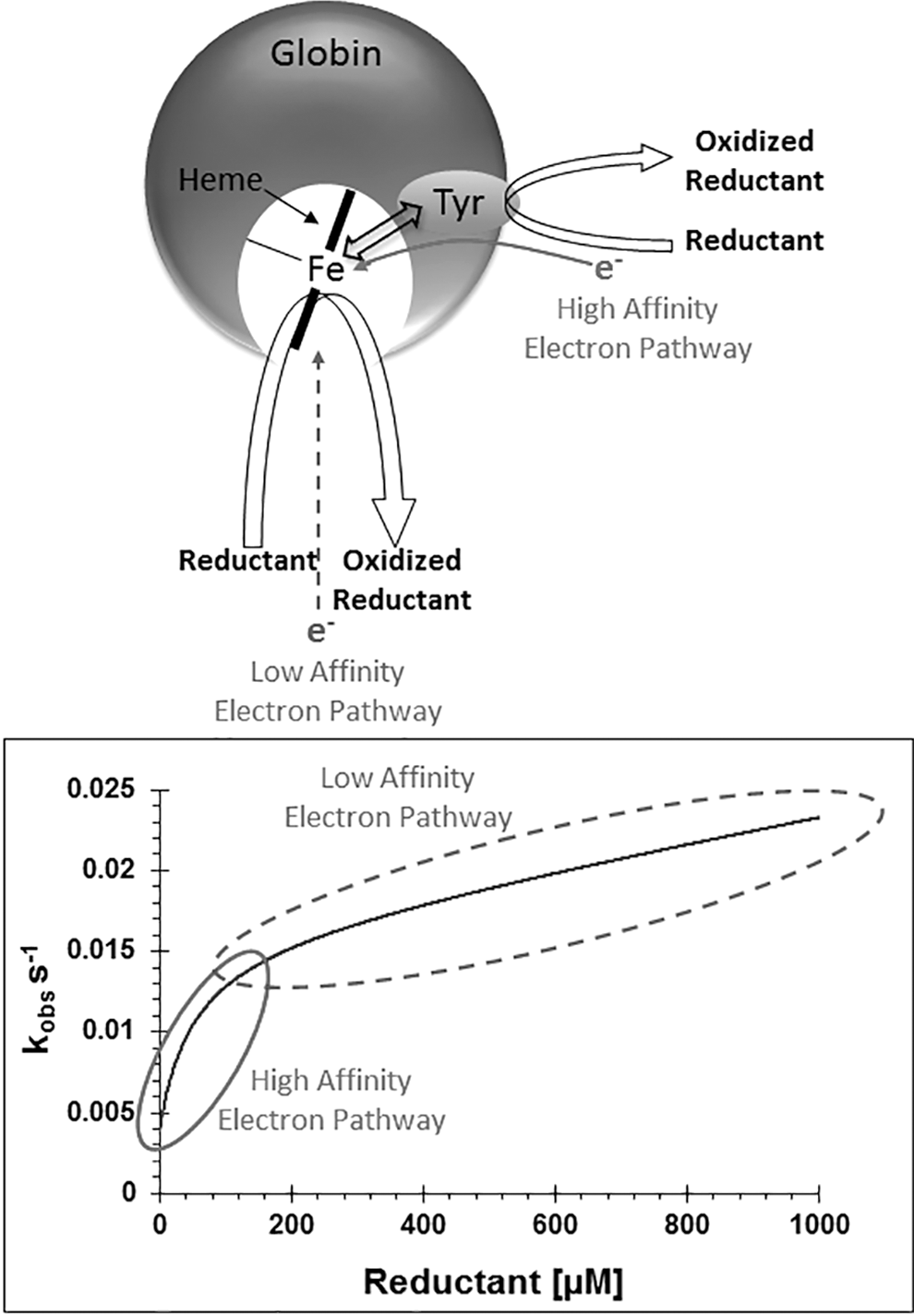
A study using a model Mb from Aplysia Fasciata (sometimes referred to as Aplysia Limacina, a junior synonym of A. Fasciata) showed that the addition of a single tyrosine residue (Phe98Tyr) to generate a TPETP interfacing the heme iron and external environment can enhance ferryl reduction up to 1200 times, two orders of magnitude faster than those found in human Hb (93). With the inclusion of a distal histidine (V63H) to more closely mimic the redox chemistry of human Hb, the half-times of ferryl reduction are altered from tens of minutes seen with the V63H Aplysia Mb variant (and human Hb) to subsecond half-times with the V63H/F98Y Aplysia Mb mutations (93). Currently, the reason for the two orders of magnitude difference of the electron transfer rates of engineered Aplysia Mb from A. Fasciata and human Hb is unknown. However, it is a current goal of research to incorporate such high rate TPETPs into recombinant human Hb as a potential HBOC (103). It is not clear whether these mechanisms of ferryl reduction relate to globins where the high oxidation state of the globin may have physiological relevance. It is noteworthy, however, some globins such as Hell's Gate globin-I, a thermophilic protein structurally similar to Ngb in Methylacidiphilum infernorum, have multiple tyrosine residues within the heme pocket (80, 112). Whether this high tyrosine content relates to thermostability or redox activity remains to be seen.
Physiological Role for Redox Chemistry of Ngb and Cygb in Health and Disease?
It is evident that the redox chemistry of vertebrate Hbs and Mbs, primarily involving the peroxidase activities of these proteins, relates to conditions of pathology where such redox activity is undesirable. However, the redox activity of globins appears to be of potential benefit, particularly to cell survival under conditions of stress. In the last two decades, the family of globins has expanded considerably, with particular interest in human globins, Ngb and Cygb, and the potential functions and relationships to cell protection in disease states. Both Ngb and Cygb are upregulated under hypoxia, suggesting potential oxygen binding/sensing, NO scavenging, or nitrite reductase functions. However, both proteins also appear to play a role in oxidative stress mechanisms and may be involved in cell signaling or cell protection. Recent research has led to the proposal of numerous potential biological functions under normal cellular conditions and in stress responses. Due to this large and diverse literature, there is currently uncertainty with regard to the primary function of these proteins (28, 72, 81, 86, 101, 123). The molecular mechanisms are still poorly understood, particularly with respect to their functionality in vivo. What follows focuses on the current opinions relating to the biological activity of Ngb and Cygb that have been advanced over the last few years.
Ngb and Neuroprotection
Discovered in 2000, Ngb is the third globin to be found in humans after Hb and Mb (12). An Hb with typical 3-on-3 α-helical fold of both Mb and Hb, Ngb differs from these proteins by way of the geometry of heme iron coordination. In the absence of exogenous ligands, the distal histidine is directly coordinated to the iron, making the ferric and deoxy ferrous heme iron low-spin hexacoordinate, coordinated by both proximal and distal histidines. The oxygen-binding proteins, Hb and Mb, are high-spin pentacoordinate forms in the absence of exogenous ligands, with the distal histidine unable to form a bond with the iron and instead forming a hydrogen bond to stabilize oxygen binding (65, 74). Nonetheless, Ngb has an affinity for oxygen of ∼1 torr, similar to that of Mb (12, 81) with a proposed function of temporary oxygen storage (25). The physiological function of Ngb may be related to its ability to react rapidly with NO in nervous tissue, functioning as an NO dioxygenase enzyme (Fig. 2). This could be important in maintaining the function of neuron mitochondria, preventing mitochondrial poisoning by NO (54, 118). However, this reactivity requires the Ngb to be in the oxyferrous form with high concentration of oxygen in neuronal cells, a supposition that is highly debated (4, 9, 12, 25, 116). An alternative view is that Ngb may function as a nitrite reductase, generating NO under conditions of oxidative or hypoxic stress (Fig. 2) (83, 114), similar to that proposed for Hb under similar conditions (19, 37, 63). Nitrite reductase activity has been proposed to underlie neuroprotective signaling under hypoxic conditions through NO production from nitrite (114), although Hb nitrite reductase activity is greatest when the protein is at its p50 for O2 saturation due to allosteric modulation (37, 38). It has also been proposed that Hb can form dinitrogen trioxide (N2O3), dissociating into NO and NO2 (37, 51). Such an application for this biochemistry of Ngb has not been elucidated, but as an uncharged molecule, N2O3 can diffuse through the cell membrane, which could potentially act as a transport for NO to the endothelium (13, 51).

Low concentrations of Ngb in brain neurons are associated with increased risk of Alzheimer's disease (111). However, Ngb has also been shown to be upregulated in Alzheimer's disease (108). It has been proposed that Ngb prevents cytochrome c from forming apopotosomes, averting neuron cell death (85). Another proposal is that Ngb functions as an oxidative stress-responsive sensor, with oxidized ferric Ngb inhibiting GDP dissociation from the alpha subunit of heterotrimeric G protein, thereby acting as a guanine nucleotide dissociation inhibitor (GDI) (122, 124). The GDI activity of Ngb correlated with its neuroprotective activity during oxidative-induced structural changes to alpha G protein (123). However, it is of note that Human Ngb, but not Zebrafish Ngb, exhibits such GDI activity (123), suggesting a more complex and diverse nature of this protein.
Whatever the biochemistry of Ngb in vivo, it is clear that the primary function of this highly conserved protein is related to the protection of neuronal activity (10) and that more research is required for a more complete understanding of the molecular mechanisms of this protein in vivo and its potential role in protection against stress-related cell damage and neurodegeneration.
Cygb, Tumor Suppression, and Cancer Therapy Resistance
Cygb is the fourth globin to be found in humans and discovered in 2001. Initially referred to as stellate cell-activating protein (56) and histoglobin (115), the heme pocket structure of Cygb is similar in nature to Ngb, possessing a hexacoordinate iron ligation in the absence of exogenous ligands (22). However, the distal histidine coordination of Cygb appears to be weaker than observed in Ngb, with partial heme pentacoordination observed in the structure of one of the Cygb subunits of the homodimer (22). This pentacoordinate-like nature of the Cygb can be further enhanced by increasing the distal histidine off-rate, observed in the monomeric form of the protein with an intramolecular disulfide bond (7). There have been many suggested functions of Cygb. Cygb has been proposed to act as an oxygen diffusion protein similar to Mb based on observations of increased expression of Cygb in heart and muscle in amphibians that lack Mb (126). Cygb has also been proposed to function as an NO scavenger, from observed high rates of NO consumption and its fast rereduction by ascorbate and cytochrome P450 reductase (64). This proposal of NO consumption has also been enhanced by increased NO levels and decreased intracellular nitrate production from partially silenced Cygb (41). However, the most intriguing property of the protein appears to relate to cell protection against oxidative stress and potential relationship with carcinomas. However, this is a double-edged sword as Cygb appears to function not only as a tumor suppressor but also protects hypoxic tumors against therapy. The potential role of Cygb in cancer suppression and hypoxic tumor protection has been recently reviewed (15).
Promotor methylation downregulates Cygb expression as observed in oral cancer (100), in head and neck squamous cell carcinoma (101), and in nonsmall cell lung carcinoma (127), indicating that the gene for Cygb functions as a tumor suppressor. Furthermore, methylation of the gene for Cygb is able to discriminate patients with and without lung cancer, using biopsies and sputum sample (102). Cygb-deficient mice have been found to promote carcinogenesis in lung and livers when exposed to carcinogens such as diethylnitrosamine (113). Cygb loss may contribute to tumor recurrence and a worse prognosis in gliomas and may serve as an independent predictive factor for prognosis of glioma patients (128).
Expression of mRNA for Cygb was found to correlate with hypoxia-inducing factor 1-alpha in squamous cell carcinoma (101), indicating that expression of Cygb is regulated by hypoxia. Increased expression of this gene correlates with clinicopathological measures of a tumor's biological aggression (101). The Cygb gene also has a protective role for proliferating keratinocytes under normoxic conditions and protects against exogenous hydrogen peroxide oxidative insults (59). This study shows that ΔNp63, a p63 transcription factor protein, targets the Cygb gene, leading to upregulation of Cygb and inhibition oxidative stress-induced apoptosis in keratinocytes. This is also present in lung and breast cancer cell lines, indicating that Cygb-mediated scavenging of reactive oxygen species may also have a role in epithelial tumors and lung cancers. Furthermore, in human lung cancer data sets, the interaction of ΔNp63 with the Cygb gene significantly predicts reduction of patient survival (59). From the studies of Cygb and cancer prevention and promotion, it is clear that Cygb has bimodal tumor suppressor and oncogene functions (73).
Although the relationship of Cygb with tumors has seen the greatest attention in recent years, other properties of Cygb also involve protection of the liver from fibrosis. Stemming from the protein's original naming in 2001 (56), Cygb is expressed in stellate cells, but not fibroblasts (70), and as such, the expression of Cygb can distinguish stellate cells from portal myofibroblasts in damaged liver. Furthermore, overexpression of Cygb in rat hepatic stellate cells inhibits their differentiation into myofibroblast-like cell phenotypes when challenged with oxidative stress (129). Similarly Cygb antifibrosis activity on rat livers decreased thioacetamide-induced liver fibrosis, which was proposed to involve apoptosis induction from activation of the caspase cascade (46).
Controlling Protein Function Through Cysteine Oxidation
As described above, the biochemistry of heme can potentially promote scavenging or formation of NO, as observed in vitro for Hb, Ngb, and Cygb (Fig. 2). In addition to this biochemistry, surface-exposed cysteine residues have the capacity to generate complexes with NO, forming S-nitrosothiol. This has been observed for human Hb in vitro binding NO to the cysteine on the beta chain of human Hb (βCys93) to form Hb-SNO (39). The relevance of Hb-SNO for hypoxia response remains a widely debated subject and only discussed in brief here (37, 39, 53, 79, 121, 132). Roles for this NO binding chemistry have been suggested to be a mechanism for the regulation of hypoxic vasodilation. This cysteine is highly conserved in the mammal and avian species (and is also present in fetal Hb as γCys93), so much so that the only other amino acids that are as highly conserved as β/γ Cys93 are a histidine and phenylalanine involved in heme stabilization (132). The relevance of Hb forming and carrying NO relates to conditions of hypoxia, with the hypothesis that hypoxic conditions trigger the release of NO to induce a vasodilation response. Indeed, mice with a βCys93Ala mutation appear to have deficient hypoxic vasodilation and enhanced mortality with transient hypoxia (132). Thus, βCys93 appears to have an antioxidant role, protecting mice against lipopolysaccharide-induced reactive oxygen species and hypertension (121).
Like Hb, Ngb and Cygb also possess Cys residues that are surface exposed and have the potential to form SNO complexes. In vitro data show that Cygb and Ngb can form SNO complexes, for example, when reacting with nitrite under anaerobic conditions (83), but the relevance of this chemistry in vivo has yet to be explored in detail. However, the cysteines of Ngb and Cygb are in pairs, promoting the view that formation of disulfide (cysteine) bonds is important in functional regulation of proteins. Such dicysteine bridges can be intermolecular, forming dimers (dimerS-S); intramolecular, forming monomers (monomerS-S); or reduced to free sulfhydryl (monomerS-H). These dicysteine bridges can alter the structure and ligand binding properties of the active site of both Ngb and Cygb and in doing so potentially regulate its function in vivo. Both Ngb and Cygb show significant dependence of their biophysical properties on the redox state of their surface-exposed cysteines (120). These cysteine pairs (Cys38/Cys83) in human Cygb (Fig. 3) are highly conserved throughout mammals, but not in other vertebrates. Cys46/Cys55 in human Ngb (Fig. 3) is more widely found in other vertebrates. Numerous studies have shown that these cysteine pairs affect both exogenous and endogenous ligand binding to the heme iron, with supposition that the redox state of the cell modulates ligand affinities of these proteins (7, 42).

Recombinant expression of Ngb primarily generates the protein with a disulfide bond between Cys46 and Cys55 (monomerS-S) located on the CD loop region of the protein (42). The protein in this monomerS-S form shows high NO dioxygenase and nitrite reductase activities, but negligible peroxidase-like activities (71, 114). This, in part, has reinforced the view that Ngb functions primarily in a cytoprotective role against NO-induced impairment of mitochondrial metabolism (105). However, the state of the cysteine pair also appears to modulate nitrite reductase activity, with enhanced formation of NO when the protein is in the monomerS-S conformation and proposed to be a stress-responsive mechanism (114). Breaking the disulfide bond leads to a large decrease in the oxygen affinity of Ngb, linked to a decrease in the histidine dissociation rate (43), Figure 4. Stabilization of the hexacoordinate form within the monomerS-H protein has also been proposed to be the mechanism of slowing the nitrite reductase activity of Ngb (114). The crystal structure of the monomerS-S form of Ngb was difficult to generate, with original published crystal structures of a double cysteine mutant (82). Molecular dynamic simulations suggested that formation of a disulfide bond changes the CD loop from strand to a helix structure, structuring the active site and acting as a redox control for Ngb function (69). The predicted structural changes of the CD loop were confirmed with a recent publication of the structure of the wild-type protein in the monomerS-S form (40). Ngb is presumed to be monomeric in vivo; however, dimers have been reported from studies using retinal neurons (77) and that such dimers are likely to be disulfide linked (60). Hence, elucidating conditions, where the protein is monomer or dimer, and the effects of these conformations on cell signaling pathways is of particular importance when considering the physiological role of the protein.

Recombinant expression of Cygb in Escherichia coli tends to lead to the formation of mixture of three forms of conformations of protein depending on the redox state of two surface-exposed cysteine residues (Cys38 and Cys83)—(i) dimerS-S with two intermolecular disulfide bridges spanning Cys38 on one protein to Cys83 on the other protein, and vice versa, (ii) monomerS-S protein with the intramolecular disulfide between the B and E helices, and (iii) monomerS-H protein with free sulfhydryl (7). Current crystallographic data on recombinant Cygb show either dimerS-S conformation [e.g., PDB structure 1UMO (22)] or monomerS-H conformation with free sulfhydryl [e.g., PDB structure 3AG0 (66)] or Cys-to-Ser mutations [e.g., PDB structure 1 V5H (107)]. To date, no structures of Cygb with intramolecular disulfide bond have been published.
MonomericS-S Cygb shows significantly different carbon monoxide and cyanide ligand affinities compared with monomer with free sulfhydryl or with cysteine mutations, showing that ligand binding in both ferrous and ferric oxidation states of the protein is affected by the redox state of cysteine residues (117). The dissociation rate for the distal histidine is 1000-fold higher with the monomerS-S protein compared with monomerS-H (6), Figure 4. Thus, the monomerS-S form of Cygb, although still primarily bis-his hexacoordinate in the absence of exogenous ligands, possesses properties that are more pentacoordinate-like properties compared with the monomerS-H protein. Furthermore, the monomericS-S form of Cygb shows higher peroxidase-like activities compared with other forms of the protein, oxidizing lipids and lipid membranes more efficiently (7). In contrast with this, the dimeric protein with intermolecular disulfides or monomer with free thiol shows poor lipid peroxidase activity, decreased ligand binding kinetics, and a ridged hexacoordinate nature (6, 7).
Crystal structure of dimerS-S Cygb places the two sulfur atoms some 12 Å distant (1 V5H) (107), suggesting a significant shift in the B/E helices to accommodate the formation of a disulfide bond between the two residues. Comparison of crystal structure of Cygb with free sulfhydryl (2DC3) (66) shows the two cysteine sulfurs to be some 7 Å closer (Fig. 3D). These effects on structure are likely to affect the heme pocket geometry on the distal side; however, no structural information of the monomerS-S form of the protein is yet available other than the ligand binding and reactivity effects described above. Nonetheless, the question remains, what is the form of Cygb in vivo and is this modulated by the redox environment of the cell?
We have previously reported that the conformer state of these cysteines also influences lipid-induced structural changes to Cygb (95). This effect is observed only in the ferric oxidation state of Cygb and only with the protein possessing an intramolecular dicysteine bond (monomerS-S) between Cys38 and Cys83. There is no effect of lipids on ferrous Cygb or ferric Cygb dimerS-S or monomerS-H (6, 7). Thus Cygb appears to have a precise response to molecules such as lipids in an oxidized cell environment. Whether this distinctive mechanism of altering protein function in vitro has relevance in vivo is currently unknown. However, the interaction between Cygb and fatty acids and lipids is similar to the finding that cytochrome c interacts with the lipid cardiolipin, a mechanism that relates to caspase-induced apoptotic response to mitochondrial damage (75, 78).
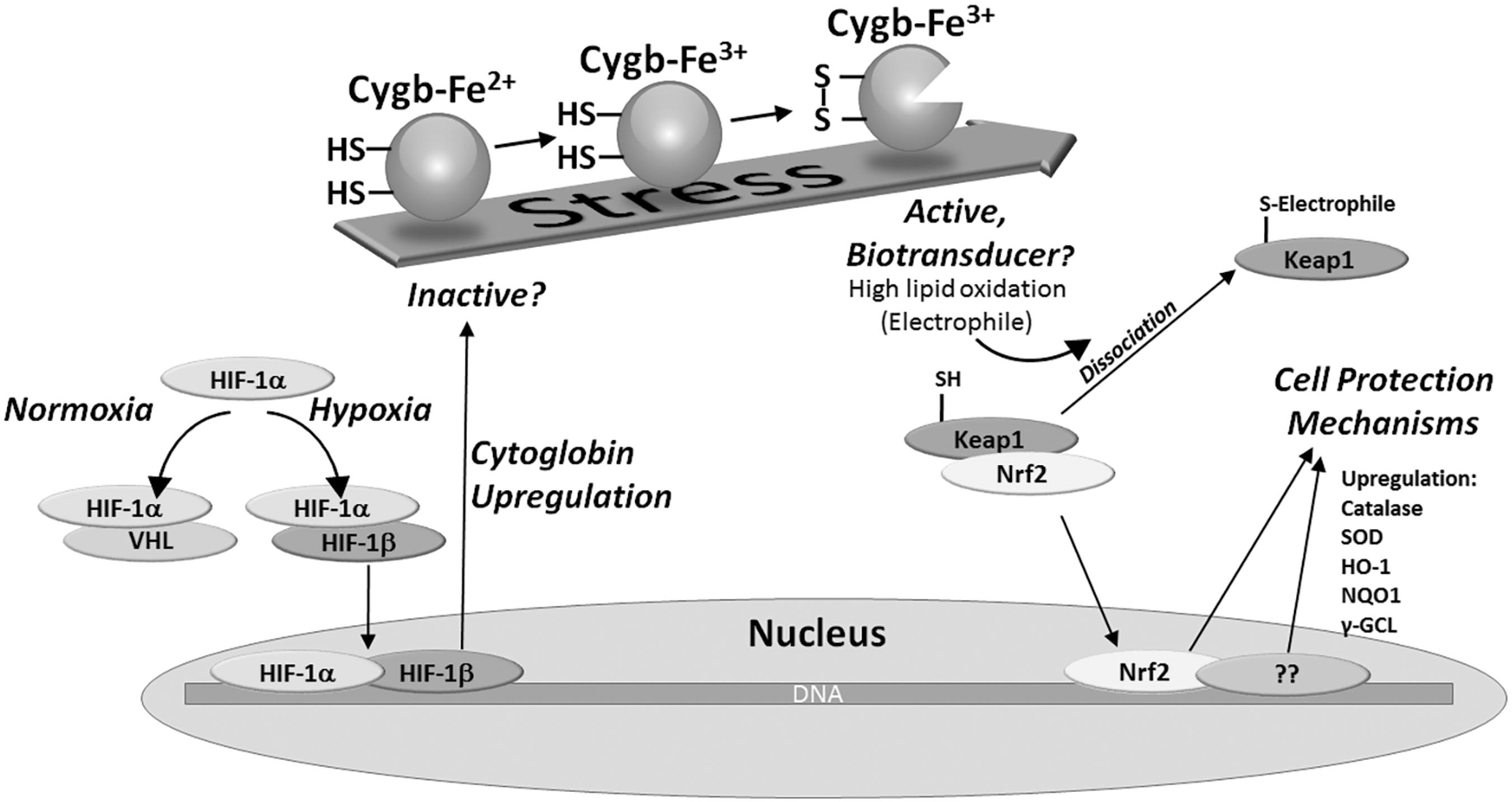
It is possible that the interaction between Cygb and lipids (or other similar molecules) has important biological and clinical roles. This together with the observations that cysteines affect the binding properties of the protein suggests a hypothesis that Cygb forms an essential signaling element between the cell's mechanism for sensing stress and its response through upregulation of antistress proteins (7, 95). This hypothesis is summarized in Figure 5. In this study, the oxidation state of the two surface cysteines is key to the protein function. In an oxidizing environment, the formation of an intramolecular disulfide bond alters the biochemical properties of the protein through changing the protein heme pocket structure and hence changes to its redox activity. One consequence of these changes in protein function, namely a high lipid peroxidase activity, would lead to production of lipid-based oxidation products. Like that of the formation of isoprostanes generated from Hb and Mb under conditions of oxidative stress, lipid oxidation would have consequences on cell signaling. This potentially links observations that hypoxia and oxidative stress upregulate Cygb production with an antioxidant response via activation of pathways such as the electrophilic-responsive element. Electrophilic lipids such as 4-hydroxynonenal, prostaglandins, and isoprostanes are known to activate antioxidant responses such as through the Nrf2/ARE (antioxidant-responsive element) pathway, inducing upregulation of antioxidant proteins, or alternative stress-responsive pathways (e.g., NF-κB) (58, 61). This mechanism of linking stress conditions to antioxidant responses could in turn be linked with pathological roles of the protein, including the observed correlation between increased Cygb gene expression and a hypoxic tumor's biological aggression (101) or the cell protection mechanism of upregulated Cygb in hypoxic cancer cells protecting hypoxic tumors from chemotherapy and radiotherapy (101). The scheme depicted in Figure 5 is based primarily from in vitro data and in vivo observations; any direct link between the two is currently unconfirmed and requires further research.
DHP: The Multifunctional Hb
When considering the potential functions of mammalian proteins such as Ngb and Cygb, information about potential physiological functions can be gained from examination of similar globins from other organisms such as bacteria and annelids. Phylogenetic studies on globins have provided insights into potential functions, particularly when the globin is localized in specific organs or is found in selected cell types. [for recent reviews, see Refs. (11, 99)]. However, most of these studies focus on vertebrate globins, and one class of globin has been largely overlooked until recently. The annelid protein, DHP, shows remarkable diversity in its capacity to have multiple functional activities (33). Despite its name, the structure of DHP has much more in common with the Hb superfamily than it has with the peroxidase superfamily, hence DHP is often referred to as DHP-Hb. DHP has a biologically relevant peroxidase activity, despite having a typical globin fold structure, converting trihalogenated phenols to quinone products using hydrogen peroxide:
DHP has the highest known peroxidase activity of any globin. Found in Amphitrite ornate, the peroxidase activity is believed to stem from evolutionary pressure for the organism to deal with toxic brominated and chlorinated compounds produced from the ecosystem in shallow coastal mudflats and marine estuaries (31 –33, 62). Two isoforms of the protein, DHP A and DHP B, differ by five amino acids (21, 23), but have considerably different peroxidase activities. The peroxidase activity of DHP A is lower by one order of magnitude than that of horseradish peroxidase, but higher by one order of magnitude than sperm whale Mb (76). DHP B is the dominant isoform and has three times the peroxidase activity compared with DHP A (33). The peroxidase mechanism involves classic peroxidase activity with the trihalogenated phenol oxidized by the radical generated with the hypervalent ferryl iron. The trihalophenol can be oxidized by compound I, taking the radical from the porphyrin, or by compound ES, with the radical believed to be associated with a series of tyrosine residues (27, 29). The second electron comes from the ferryl iron of compound II to form ferric DHP and the quinone product. Additionally, DHP has further activity revealed with peroxygenase activity, with oxygen atom transfer to substrate following ferryl formation, as well as oxidase activities, with dioxygen consumption in the absence of peroxide (5).
The oxygen affinities for DHP A and B are 4.2 and 2.0 torr, respectively (33, 109), hence DHP appears to have affinities for oxygen in the range for useful delivery of oxygen to the organism. Indeed, DHP A and B are the only Hb genes identified in A. ornata (45). Sometimes referred to as the dehaloperoxidase paradox, the function of DHP as an O2 carrier and a peroxidase depends on the oxidation state of the heme iron (34). A structural comparison of the heme pocket of DHP compared with erythrocyte Hb and Cygb (Fig. 6) shows small differences, specifically in the flexibility of the distal histidine and the number of aromatic residues in the distal pocket. While DHP has features that appear to be unique to the family of globins, both the structure and properties of the protein support the possibility, indeed a probability, of multifunctional roles of other globins, particularly those of Cygb and Ngb.

Summary and future directions
It is evident that the redox chemistry of Hbs, once considered an in vitro curiosity, has real physiological and pathological roles. DHP is a true multifunctional protein, not only binding oxygen but also performing peroxidase activities vital for the organisms' survival. Mb, although primarily an O2-binding protein, has a secondary function as an NO dioxygenase, protecting mitochondria from respiratory poisoning by NO. Some view Hb to have a role for carrying NO as SNO-Hb, but like Mb, it definitely shows redox chemistry when outside its normal antioxidant environment. These proteins have, or appear to serve, multifunctional roles. It is not a great leap in faith to propose that other heme proteins such as Ngb and Cygb may also have multifunctional properties, involving ligand binding and stress response mechanisms. However, the detailed mechanisms of these roles in vivo have yet to be fully explored, particularly in respect to linking the in vitro reactivities of the proteins to gene expression studies and biochemical studies of the properties of the proteins in vitro.
The recent discovery in 2012 of a fifth globin in humans, androglobin (Adgb), extends the interest in globin redox chemistry and function even further. With a 1667 amino acid sequence, Adgb comprises a calpain-like domain, an Hb domain, and an IQ calmodulin-binding motif (50). The discovery of a role of Adgb as an oncogene, promoting cell proliferation, has important implications in cancer research (52); however, very little is known about this multidomain protein. Phylogenetic studies have gained insights into the potential functions of some globins, particularly when appearing in specific organs and vertebrate species (11, 99). Nevertheless, many of the physiological functions of globins are yet to be identified or the observed in vitro functions verified in vivo. The current interest of the role of globins such as Ngb in neurodegenerative disease and Cygb cancer therapy resistance makes deciphering these mechanisms of globins a critical objective for developing an understanding of these diseases and potentially discovering novel targets for therapy.
Footnotes
Author Disclosure Statement
B.J.R. has patents and patent pending pertaining to the use of through-protein electron transfer pathways for use in Hb-based blood substitutes.