
Abstract
Aims:
Human immunodeficiency virus (HIV) infection induces oxidative stress and alcohol use accelerates disease progression, subsequently causing immune dysfunction. However, HIV and alcohol impact on lipid rafts-mediated immune dysfunction remains unknown. In this study, we investigate the modulation by which oxidative stress induces reactive oxygen species (ROS) affecting redox expression, lipid rafts caveiloin-1, ATP-binding cassette (ABC) transporters, and transcriptional sterol regulatory element-binding protein (SREBP) gene and protein modification and how these mechanisms are associated with arachidonic acid (AA) metabolites in HIV positive alcohol users, and how they escalate immune dysfunction.
Results:
In both alcohol using HIV-positive human subjects and in vitro studies of alcohol with HIV-1 gp120 protein in peripheral blood mononuclear cells, increased ROS production significantly affected redox expression in glutathione synthetase (GSS), super oxide dismutase (SOD), and glutathione peroxidase (GPx), and subsequently impacted lipid rafts Cav-1, ABC transporters ABCA1, ABCG1, ABCB1, and ABCG4, and SREBP transcription. The increased level of rate-limiting enzyme 3-hydroxy-3-methylglutaryl HMG-CoA reductase (HMGCR), subsequently, inhibited 7-dehydrocholesterol reductase (DHCR-7). Moreover, the expression of cyclooxygenase-2 (COX-2) and lipoxygenase-5 (5-LOX) mRNA and protein modification tentatively increased the levels of prostaglandin E2 synthases (PGE2) in plasma when compared with either HIV or alcohol alone.
Innovation:
This article suggests for the first time that the redox inhibition affects lipid rafts, ABC-transporter, and SREBP transcription and modulates AA metabolites, serving as an important intermediate signaling network during immune cell dysfunction in HIV-positive alcohol users.
Conclusion:
These findings indicate that HIV infection induces oxidative stress and redox inhibition, affecting lipid rafts and ABC transports, subsequently upregulating AA metabolites and leading to immune toxicity, and further exacerbation with alcohol use. Antioxid. Redox Signal. 28, 324–337.
Introduction
H
To the best of our knowledge, this is the first study that demonstrates both in vitro and ex vivo using peripheral blood mononuclear cells (PBMCs) from human immunodeficiency virus (HIV)-positive alcohol users, that immune dysfunction characteristic of HIV-positive alcohol user affects redox expression, subsequently affecting lipid rafts, rate-limiting enzymes, ABC transporters, and SREBP transcriptional function and activation of arachidonic acid metabolites impacting PBMCs. The present findings suggest that HIV-induced oxidative stress affects signaling mechanisms that are accelerated by alcohol, intensifying viral replication and disease progression by impairing immune function.
HIV infection and alcohol use are known to affect the immune system and cause immune dysfunctions. Increasing evidence shows that HIV infection (36, 51) and alcohol (33, 38, 53, 55) negatively impact monocytes (MC), dendritic cells (DC), and T cells both in vitro and in vivo. These effects subsequently increase HIV replication and disease progression (2, 6), which is exacerbated by alcohol. Further, these immune dysfunctions and pathogenic mechanisms may facilitate the ability of the HIV virus to enter the central nervous system, inducing neuropathogenesis. Studies have shown that HIV infection and alcohol use directly affect the periphery, causing antigen abnormalities (2, 3), and increasing susceptibility and severity of dysfunction in the immune system (5, 6, 12). Previous studies have shown that HIV-infected patients with immune impairments are synergistically potentiated in alcohol consumption (42, 47, 48).
Oxidative stress induces free radicals (H2O2 and O2 −) and reactive oxygen species (ROS), causing cellular damage in many diseases including HIV/AIDS and alcohol use (11, 14, 40), which affect redox expression and, subsequently, activate arachidonic acid (AA) metabolites such as cyclooxygenase-2 (COX-2) and lipoxygenase-5 (5-LOX). However, redox protein glutathione synthetase (GSS), super oxide dismutase (SOD), glutathione peroxidase (GPx), and catalase are the main players in intracellular antioxidant defense mechanism and are associated with maintaining immune function (27). The reduced level of GSS increases the production of free radicals in HIV-positive as well as in alcohol users (1, 28, 54), which could lead to immune dysfunction. Inhibition of redox proteins, subsequently, accelerates AA and its metabolites COX-2 and 5-LOX. These metabolites are major contributors to chronic inflammation and imbalance of immune cell response. HIV infection induces oxidative stress and affects redox expression, which may impact lipid rafts and ATP-binding cassette (ABC) transporters in the immune cells. However, lipid rafts are enriched in AA; once altered, COX-2 activation of AA metabolites, subsequently, affects lipid rafts.
Caveolin-1 (Cav-1), a specialized form of lipid raft, serves as a cellular marker and contains cholesterol (63, 65). Cholesterol plays a major role in HIV infection by budding target cells and delivering virions to the plasma membranes (9, 29). Also, HIV-induced oxidative stress and the impact of sterol deficiency could affect sterol regulatory element-binding proteins (SREBPs) that are involved in the progression of immune dysfunction (8). Lipid protein Cav-1 then targets cells activated by SREBP and SCAP pathways that modulate cholesterol biosynthesis. These SREBP transcriptional functions depend on the cholesterol biosynthesis of rate-limiting enzymes 3-hydroxy-3-methylglutaryl HMG-CoA reductase (HMGCR) and 7-dehydrocholesterol reductase (DHCR7) (7, 34, 52), and ABC transporters (59, 64). HIV modulates circulating and tissue cholesterol levels, which have major impacts on initiation, progression, and regression of HIV-associated neurocognitive disorders (HAND). It is known that ABC-transporter families play important roles in this modulation and are induced by the same sterol-sensing nuclear receptor system. Clinical observations suggested that patients with HAND may have dyslipidemia deficits causing anti-retroviral function and degradation of the protein responsible for altering lipid rafts (63, 65). However, the underlying mechanisms of how HIV and alcohol use act on accelerating immune dysfunction are not clearly understood. Despite mounting evidence that excessive alcohol use may exacerbate HIV disease, the combined role and effects of alcohol and HIV infection on the induction of oxidative stress leading to affect lipid rafts, ABC transporters, and AA metabolites in disease progression remain unknown. Therefore, we try to understand the mechanisms of alcohol-enhanced HIV infection-induced lipid rafts-mediated immune disorders.
In this study, we investigated alcohol effects when combined with HIV-1 infection and/or HIV-1 envelop glycoprotein 120 (HIV-1 gp120) in oxidative stress, lipid rafts, ABC transporters, and SREBP transcription. We showed that alcohol comorbidity with HIV infection or HIV-1 gp120 protein is involved in the enhanced changes of redox inhibition, lipid rafts, ABC transporters, SREBP transcription, and AA metabolites COX-2, 5-LOX mRNA expression and protein modification by peripheral blood mononuclear cells (PBMCs).
Results
HIV-induced oxidative stress and ROS production affect redox expression and these effects are accelerated by alcohol in PBMCs
HIV infection-induced ROS production and inhibited redox expression lead to immune dysfunction (40). However, the mechanism of how HIV infection alters ROS production and inhibits redox expression subsequently affecting lipid raft and how these mechanisms accelerate alcohol effects are not yet studied. Therefore, we were determined to study whether HIV-1 gp120 induced oxidative stress and whether alcohol accelerated ROS production and affected redox expression in PBMC. Observed results demonstrated that HIV-1 gp120 comorbidity with alcohol at 24 h significantly increased oxidative stress and ROS production, subsequently inhibiting the redox expression in PBMCs compared with either HIV-1 gp120 or alcohol alone. Figure 1 shows that HIV-1 gp120, alcohol, and HIV-1 gp120 with alcohol induce ROS production (Fig. 1A), whereas dose-response effects of gp120 (Fig. 1B) and alcohol (AL; Fig. 1C), and combined effect of GSS (Fig. 1D), SOD (Fig. 1E) and catalase (CAT; Fig. 1F) redox expression significantly decreased when compared with the control at 24 h. The effects of ROS (p < 0.01), GSS (p < 0.003), SOD (p < 0.003), and CAT (p < 0.003) confirmed that alcohol comorbidity accelerated HIV-1 gp120 inhibition of redox gene expression. Figure 2 demonstrated that PBMCs treated in vitro with alcohol and HIV-1 gp120 induced oxidative stress, which, subsequently, increased AA metabolites COX-2 (p < 0.001) (Fig. 2A) and 5-LOX (p < 0.001) (Fig. 2B) in vitro. In the case of alcohol users (p < 0.04), HIV-positive (p < 0.03) and HIV-positive alcohol users (p < 0.002), there was an increase in COX-2 expression (Fig. 2C). Similarly, the expression level of 5-LOX significantly increased in alcohol users (p < 0.03), HIV-positive (p < 0.04) and HIV-positive alcohol-using subjects (p < 0.001) (Fig. 2D) compared with normal subjects. The plasma level of prostaglandin E2 synthases (PGE2) was significantly increased in alcohol users (p < 0.01), HIV-positive subjects (p < 0.001), and HIV-positive alcohol users (p < 0.001) compared with healthy normal subjects (Fig. 2E). These in vitro results suggested that HIV-1 gp120 protein induced oxidative stress and ROS production, inhibiting redox expression and, subsequently, upregulating AA metabolites, and these effects were accelerated by alcohol. This supported the notion that at least, partially, HIV positive with alcohol users increased AA metabolites COX-2 and 5-LOX expression, which may induce ROS production and impact redox inhibition.


HIV-1 gp120 and alcohol effect on lipid rafts in PBMCs
It is known that oxidative stress-inducing H2O2 and free radical O2− scavenges are major targets of membrane lipids. However, HIV infection and alcohol comorbidity induce peroxidative damage on lipid rafts and their functions are not yet studied. We determined the effect of HIV-1 gp120 and alcohol on lipid rafts and their immune role in PBMCs. Figure 3A demonstrated that HIV-1 gp120 induced significant downregulation of Cav-1 expression in AL (p < 0.03), HIV-1 gp120 (p < 0.03), and HIV-1 gp120 with AL (p < 0.001)-exposed PBMCs. Lipid rafts contain cholesterol, which is controlled by SREBP transcription and rate-limiting enzymes HMGCR. Figure 3B, C illustrates that HIV-1 gp120 with AL significantly reduced SREBP (p < 0.001) and increased the level of HMGCR expression in HIV-1 with AL (p < 0.001). Further, the impact of Cav-1 activates rate-limiting HMGCR gene expression, subsequently inhibiting the transcriptional factor SREBP. Therefore, in this study, we examined the effect of HIV-1 gp120 and alcohol, which could be employed to assess the progression of lipid rafts, mediated immuno-pathogenic mechanisms in PBMCs. The observed results confirmed that HIV-1 gp120 affected lipid raft functions that are potentiated by alcohol comorbidity.

HIV-1 gp120 and alcohol effect on ABC transporters in PBMCs
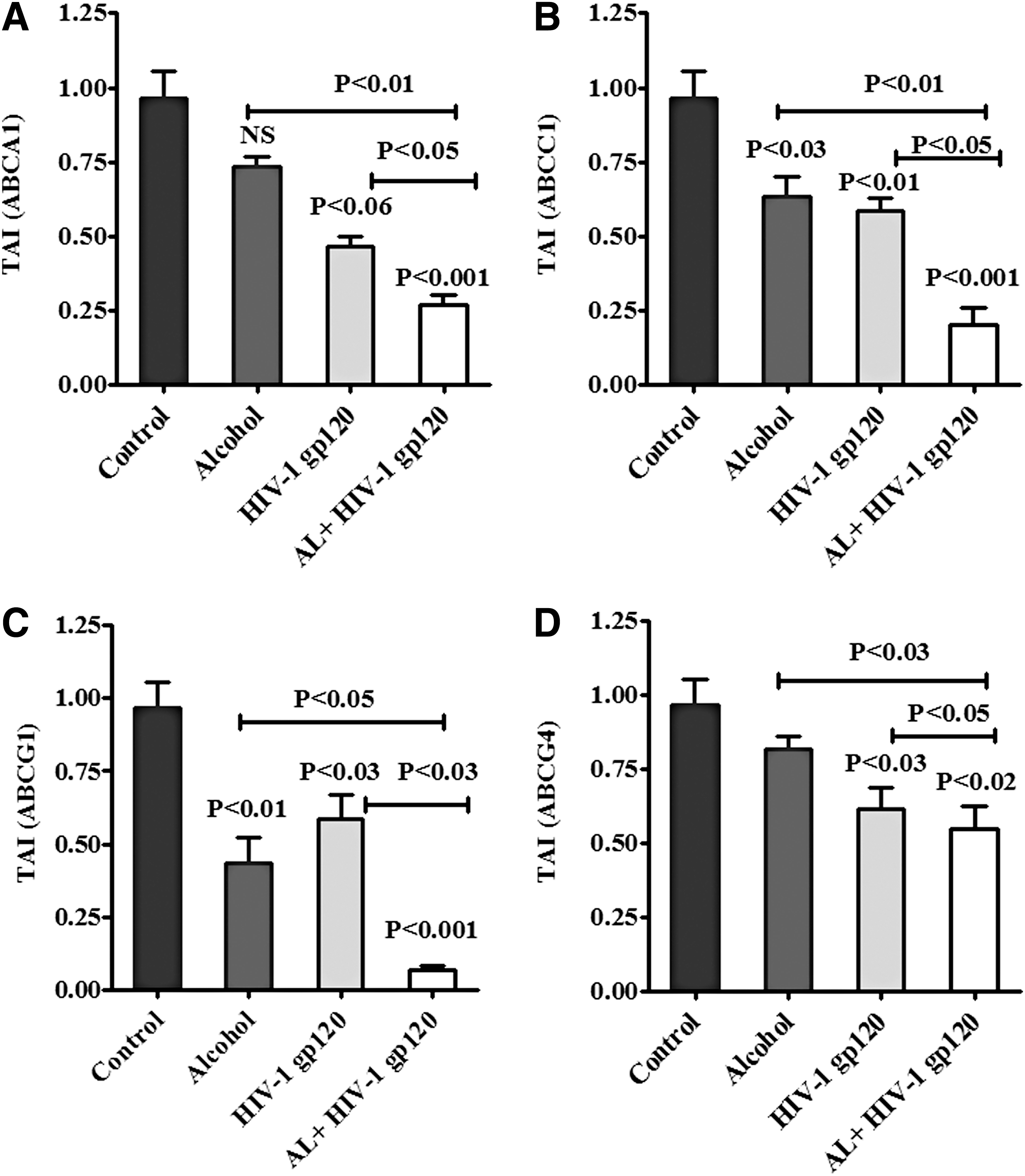
Previous studies demonstrate that ABC transporters significantly modulate immune disorders in HIV infection (64). We have investigated the role of ABC transporters in HIV infection in combination with alcohol and how they affect lipid raft dysfunction in PBMCs. Data presented in Figure 4 show that HIV-1 gp120 and AL-treated PBMC have inhibited the expression level of ABC transporters ABCA1 in HIV-1 gp120 (p < 0.06), AL-non-significant (NS), and HIV-1 gp120 with AL (p < 0.001) (Fig. 4A); ABCC1 in HIV-1 gp120 (p < 0.03), AL (p < 0.01), and HIV-1 gp120 with AL (p < 0.001) (Fig. 4B); ABCG1 in HIV-1 gp120 (p < 0.03), AL (p < 0.01), and HIV-1 gp120 with AL (p < 0.001) (Fig. 4C); and ABCG4 in HIV-1 gp120 (p < 0.03), AL (NS), and HIV-1 gp120 with AL (p < 0.02) (Fig. 4D), respectively, at 24 h. These findings demonstrate that alcohol combined with HIV-1 gp120 significantly downregulated all ABC transporters compared with either HIV-1 gp120 or AL exposure alone.
Effect of HIV-1 gp120 and alcohol on redox proteins and AA metabolites in PBMCs
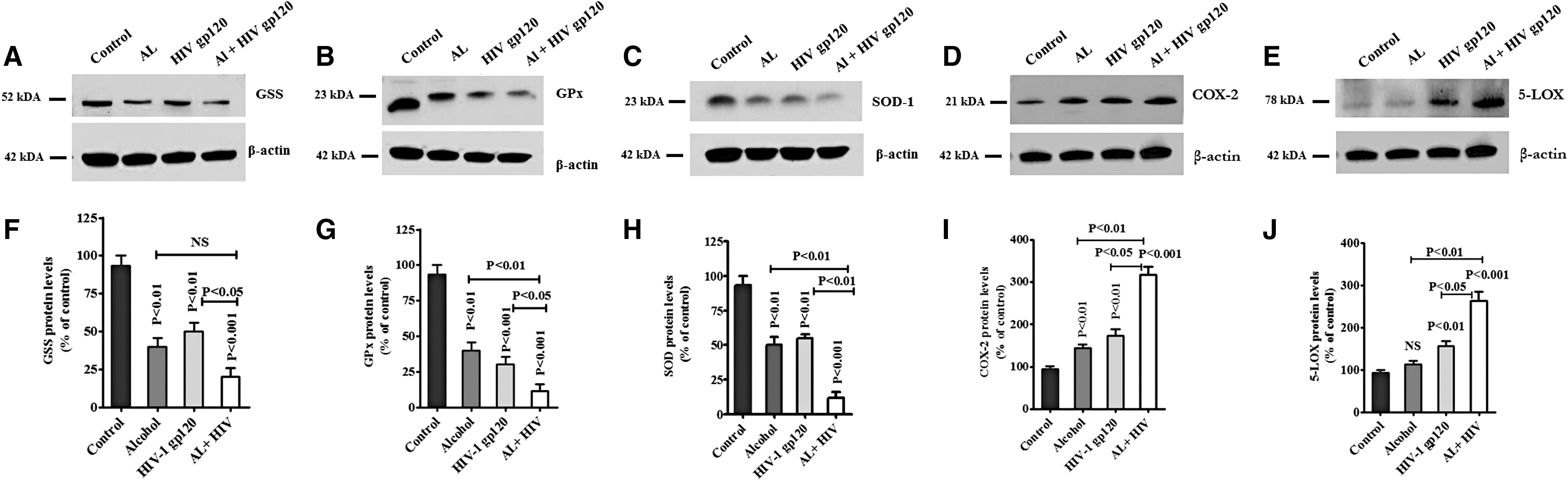
Next, we analyzed these redox proteins GSS, SOD, and GPx, and protein expression for AA metabolites COX-2 and 5-LOX in alcohol, HIV-1 gp120, and HIV-1 gp120 with AL-exposed PBMCs. Figure 5 demonstrates that GSS (Fig. 5A), GPx (Fig. 5B), and SOD (Fig. 5C) protein were significantly downregulated; whereas COX-2 (Fig. 5D) and 5-LOX (Fig. 5E) were significantly upregulated by alcohol, HIV-1 gp120 and HIV-1 gp120 with AL. These observed results were consistent with gene expression in AL with HIV-1 gp120 protein. Data presented in Figure 5F–J show the densitometry evaluation in HIV-1 gp120 with AL, respectively, in GSS (p < 0.001), SOD (p < 0.001), GPx (p < 0.001), COX-2 (p < 0.001), and 5-LOX (p < 0.001) compared with control.
Effect of HIV-1 gp120 and alcohol on lipid rafts, ABC transporter proteins, and fatty acid binding protein
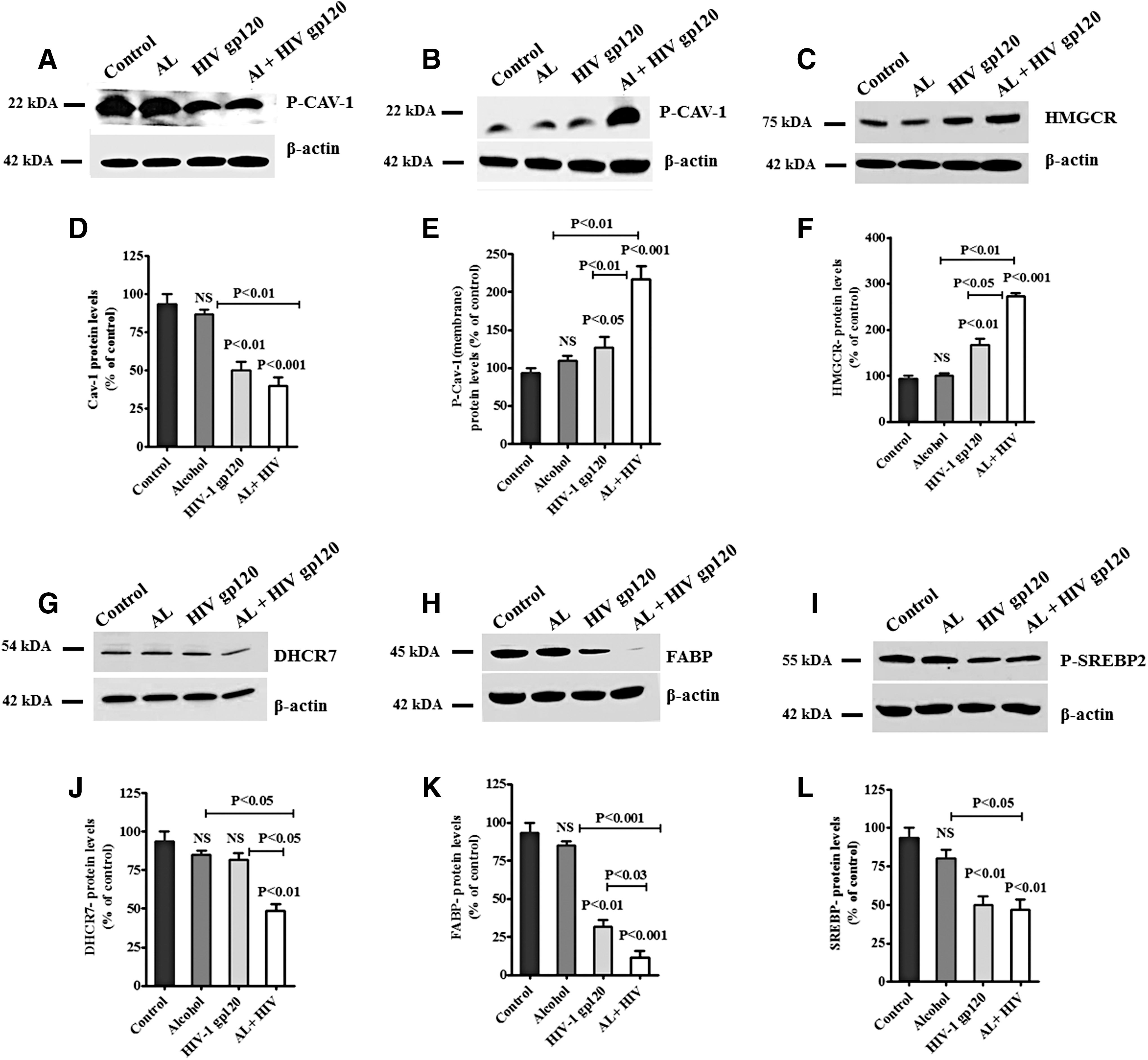
We also analyzed lipid rafts Cav-1, fatty acid protein 7 (FABP-7), and SREBP, and rate-limiting enzymes HMGCR and DHCR7 protein expression in terms of AL, HIV-1 gp120, and HIV-1 gp120 with AL exposure. Figure 6 shows the expression of cytoplasmic Cav-1 (Fig. 6A), membrane Cav-1 (Fig. 6B), HMGCR (Fig. 6C), DHCR-7 (Fig. 6G), FABP-7 (Fig. 6H), and SREBP-2 (Fig. 6I) proteins. Results demonstrated that Cav-1 (Fig. 6A), DHCR7 (Fig. 6G) FABP (Fig. 6H), and SREBP (Fig. 6I) proteins were significantly downregulated; whereas membrane Cav-1 (Fig. 6B) and HMGCR (Fig. 6C) were significantly upregulated by AL, HIV-1 gp120, and HIV-1 gp120 with AL. HIV-1 gp120 and AL induced Cav-1 in cytoplasm, subsequently translocating to the membrane (Fig. 6A, B). Figure 3H demonstrated that HIV-1 gp120 and alcohol significantly downregulated FABP-7 expression compared with either HIV-1 gp120 or alcohol alone. Data presented in Figure 6D–F and J–L show the densitometry evaluation for cytoplasm CAV-1 (p < 0.001), membrane CAV-1 (p < 0.001), HMGCR (p < 0.001), DHCR7 (p < 0.01), FABP (p < 0.001), and SREBP (p < 0.01) in HIV-1 gp120 with AL exposure.
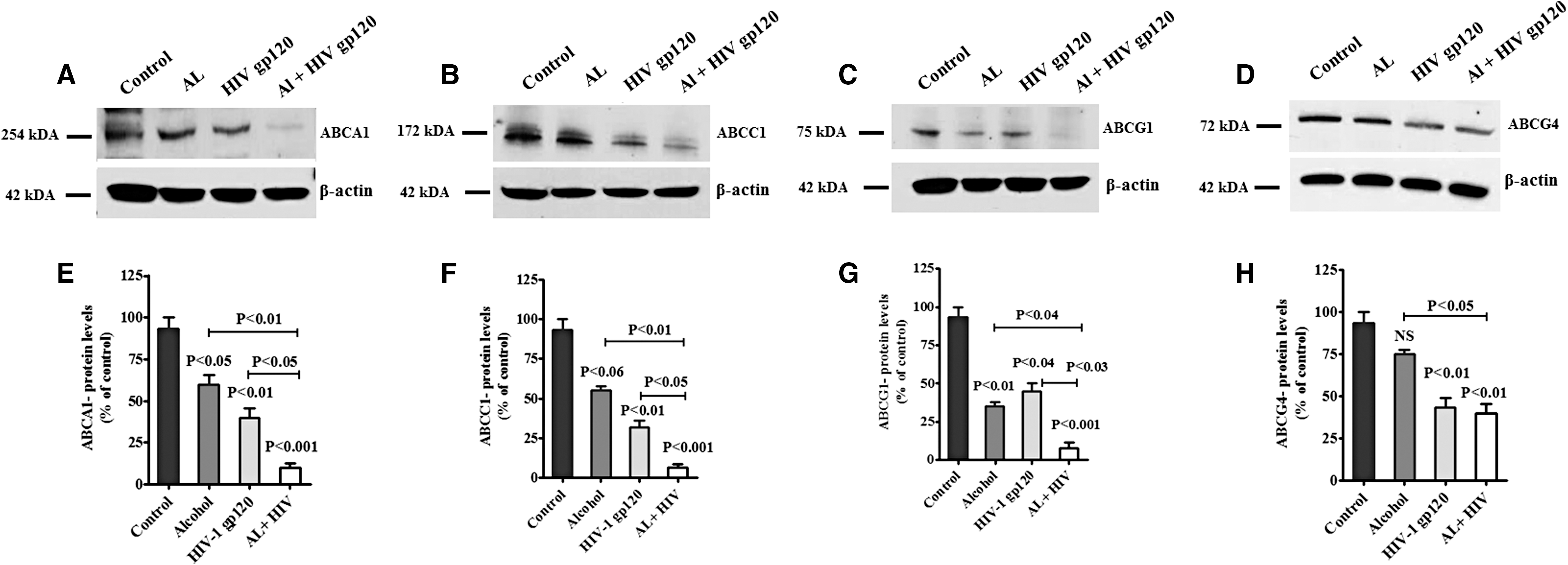
In addition, we also analyzed ABC transporters ABCA1, ABCC1, ABCG1, and ABCG4 protein expression in alcohol, HIV-1 gp120, and AL with HIV-1 gp120-treated PBMCs. Figure 7 demonstrated that ABCA1 (Fig. 7A), ABCC1 (Fig. 7B), ABCG1 (Fig. 7C), and ABCG4 (Fig. 7D) proteins were significantly downregulated in AL, HIV-1 gp120, and AL with HIV-1 gp120 when compared with control. These observed results were consistent with gene expression in AL with HIV-1 gp120. Data presented in Figure 7E–H show the densitometry analysis of ABCA1 (p < 0.001), ABCC1 (p < 0.001), ABCG1 (p < 0.001), and ABCG4 (p < 0.01) compared with control.
Discussion
Lipid rafts are cell membrane components enriched by cholesterol/phospholipids and transporters, and very sensitive indicators of immune dysfunction (23). It is well known that lipid rafts play a major role in membrane structure, functional organization, and protection from oxidative stress and AA metabolites. Lipid rafts serve as entry and exit sites for microbial pathogens and toxins, including the HIV and substances of abuse such as alcohol (15, 57). Cholesterol has been associated with immune disorders and it is important during various stages of the HIV-1 replication cycle, especially during the early stages of viral infection and disease progression (26, 31). Alcohol consumption has enhanced the deleterious effect of HIV in lymphocytes (25), neutrophils (42), MC/macrophages, phagocytes, and DC (18, 55, 61) in HIV-infected individuals (4, 48). Also, HIV-1 infection induces oxidative stress and redox modification, which is associated with enhanced viral replication and disease progression in HIV-infected patients as well as in alcohol users (47, 48). Oxidative stress inducing ROS production and redox inhibition is a major target in lipid rafts that affects immune regulatory functions (e.g., immune tolerance) (35). Increased level of ROS production, subsequently, affects immune function in HIV infection (13). Increased oxidative stress and ROS production altered cholesterol synthases affecting membrane lipids, which may play a wide role in the development of AIDS (37).
The Env glycoprotein (gp) precursor protein (gp160) undergoes proteolytic maturation to generate gp41 (membrane inserted) and gp120 (membrane inserted or shed from the cell surface). Soluble gp120 can induce apoptosis in lymphocytes via activation on chemokine receptors (44). The transmembrane region of gp41 is also important for Env activity. Rafts contribute to HIV assembly and budding and this is known as HIV-1 infection; during replication, lipid rafts can direct pathogen trafficking to specific sites for replication and/or facilitate the assembly of the enveloped viruses. Specifically, HIV-1 protein gp120, which mediates several steps in virus assembly, is anchored into T cell lipid, rafts (20). Studies have also shown that HIV-1 binding to T cells involves interactions between HIV-1 envelope proteins gp120 with raft-associated CD4 and co-receptors CCR5 and CXCR4 (44). Disruption of lipid rafts by cholesterol depletion is known to inhibit HIV entry into T cells. The results of this study provide strong evidence that caveolae translocation and localization, subsequently, have an effect on the cellular immune response by HIV-1 gp120 and alcohol. People with a history of alcohol abuse or dependence may be more prone to disease progression (45, 46). In addition, alcohol has a major effect on AA via its perturbation of enzymes that are involved in the conversion of PGE2 and COX-2 related metabolites. COX-2 is expressed in several cell types after inflammatory stimulation, including MC and macrophages (41), and it is known that the rate-limiting enzyme is involved in AA metabolism, thereby generating prostaglandins and thromboxanes, molecules that play important roles in the inflammatory response and formation of ROS (58). These results confirm that COX-2-mediated reactions, oxidized metabolites, and byproducts of these enzymatic reactions contribute to ROS within the cell. Supporting this, we have recently reported that HIV infection significantly upregulated COX-2 and 5-LOX induction and plasma level of PGE2 in HIV-positive subjects (50).
The ABC transporters are an important group of membrane-associated proteins that serve as transmembrane transporters for various cellular functions and are known to be associated with membrane lipid rafts (19). However, there are no reports on the molecular mechanisms of alcohol effects on lipid rafts modulating or ABC transporter activity by HIV-1 gp120. Studies have shown a direct causal link between the activation of the COX-2/PGE2 signaling pathway and ABC transporters (21). The ABC transporters (ABCA1, ABCG1, and ABCG4) constitute a ubiquitous superfamily of integral membrane proteins that are responsible for the ATP-powered translocation of many substrates across membranes (43), playing a major role in cholesterol transport and sterol function (7). ABCC1 and ABCG1 export excess cellular cholesterol into the HDL pathway and reduce cholesterol accumulation in PBMCs (39). Further, disruption of ABCA1 and ABCG1 promotes accumulation of excess cholesterol in PBMC. These transporters are highly regulated post-transcriptionally by diverse processes. FABP is known to be part of lipid rafts and impacts redox changes in immune as well as neuronal cells. However, FABP-7 plays a crucial role in AA metabolism controlling lipid homeostasis (17). Therefore, we have tested whether HIV- and AL-induced stress response inhibits ABC transporter and FAB proteins. The current investigation provides new insights into the functional role of oxidative stress and redox changes affecting AA metabolites that, subsequently, affect lipid rafts and ABC transporters in HIV-1 gp120 with AL-exposed PBMCs.
In this study, we have demonstrated, for the first time, that in vitro as well as ex vivo (HIV-positive and HIV-positive AL users) there is an increase in the levels of oxidative stress and redox inhibition of GSS, SOD, and GPx (Fig. 1), associated with increased AA metabolites such as COX-2 and 5-LOX mRNA expression (Fig. 2). It is known that oxidative stress is the major player in immune dysfunction (14, 40), and increased AA metabolites PGE2, COX-2, and 5-LOX may enhance viral replication and disease progression. These studies suggest that alcohol comorbidity in HIV-positive subjects may have an enhanced role in oxidative stress and AA metabolites compared with normal subjects and nonalcohol-using HIV-positive subjects. This is consistent with earlier reports of HIV-positive cases, where activation of the COX-2 and 5-LOX pathways has been observed (16, 24, 50).
Also, studies have shown that redox impairments are associated with altered lipid rafts and membrane transporters (10). It is known that lipid rafts are involved in HIV-1 entry, host cell signal transduction pathways, intracellular trafficking, and assembly of viral proteins at several stages of HIV-1 replication (9, 29). These studies further confirm that the downstream effect of lipid rafts protein Cav-1 translocates to the membrane and is accordingly reduced in the cytoplasm, which, subsequently, inhibits SREBP transcription (Fig. 3A, B). Cholesterol plays a major role in HIV infection by budding target cells and delivering virions to the plasma membranes. In addition, HIV-induced oxidative stress alters sterol deficiency and could affect lipoprotein-mediated Cav-1 and SREBPs that are involved in the progression of HIV-associated immune dysfunction. Studies have also shown that certain enveloped viruses have also evolved to use lipid rafts or cholesterol to support their replication in host cells (30). However, these SREBP transcriptional functions depend on the cholesterol biosynthesis of rate-limiting enzymes HMGCR and DHCR7. The rate-limiting step in this pathway is the conversion of HMGCR-CoA into mevalonate by HMGCR. These two enzymes oppositely regulate each other; reduced levels of DHCR7 lead to increased HMGCR and are further exacerbated by alcohol in cholesterol levels. Observed results confirm that HMGCR is significantly upregulated, whereas DCHR7 is downregulated in HIV-1 gp120 along with alcohol (Fig. 3C, D). It is known that membrane cholesterol may have a consequential effect through the translocation of viral components. Collectively, cholesterol and lipid rafts might be vulnerable to the effects of HIV infection. These results confirm that increased oxidative stress and AA metabolites exacerbate viral replication and disease progression in alcohol with HIV-1 gp120.
Further, our results show that alcohol, HIV-1 gp120, and AL along with HIV-1 gp120 induce oxidative stress and inhibit redox expression, which may induce apoptosis. Soluble gp120 is known to induce apoptosis in lymphocytes via activation of chemokine receptors (44); however, to the best of our knowledge, this is the first study to demonstrate a combined effect of ethanol and gp120 on human PBMCs. Even though we did not measure apoptosis, caveolae translocation and localization play a regulatory role in immune suppression and inflammatory cytokines induce Cav-1/β-catenin signaling in rat cell apoptosis through the p38 MAPK pathway (60), which could also be applied in the context of alcohol and HIV effects on cellular responses in PBMC. In addition, AA metabolites COX-2 and 5-LOX protein activation (Fig. 5) is associated with a concomitant inhibition of Cav-1, SREBP, DCHR7, and FABP-7 and upregulation of HMGCR (Fig. 6). The main observation in this article is that cells from HIV-positive AL users express higher levels of oxidative stress and ROS production impacting lipid rafts and ABC transporters effect due to increased COX-2 and 5-LOX activation (Fig. 7). However, HIV-1 gp120 with AL has a higher significant impact than either HIV-1 gp120 or AL alone. This suggests that there is an interactive role between AL and HIV that synergistically potentiates and increases dysfunction when compared with either AL or HIV positive or HIV-1 gp120 protein individually.
Previous studies indicate that HIV infection and HIV proteins with drugs of abuse affect immune cells such as MC and DC (36, 49, 51). In this study, using PBMCs from HIV-positive alcohol users and treating PBMCs with HIV-1 gp120 and AL in vitro, we demonstrated that an increase in oxidative stress and ROS production affects redox expression, lipid rafts, ABC transporters, and SREBP transcription when compared with the control. Also, there was an increase in oxidative stress and damage of membrane lipid rafts in PBMCs, which eventually may lead to an enhanced production of AA metabolites COX-2 and PGE2, altering immune tolerance and causing immunopathogenic dysfunction (Fig. 8). These results suggest that PBMCs play a wide role in the protective mechanisms of immune function. Once the effect of HIV infection or AL alters lipid rafts, this may, subsequently, accelerate disease progression that is mediated by ABC transporters.
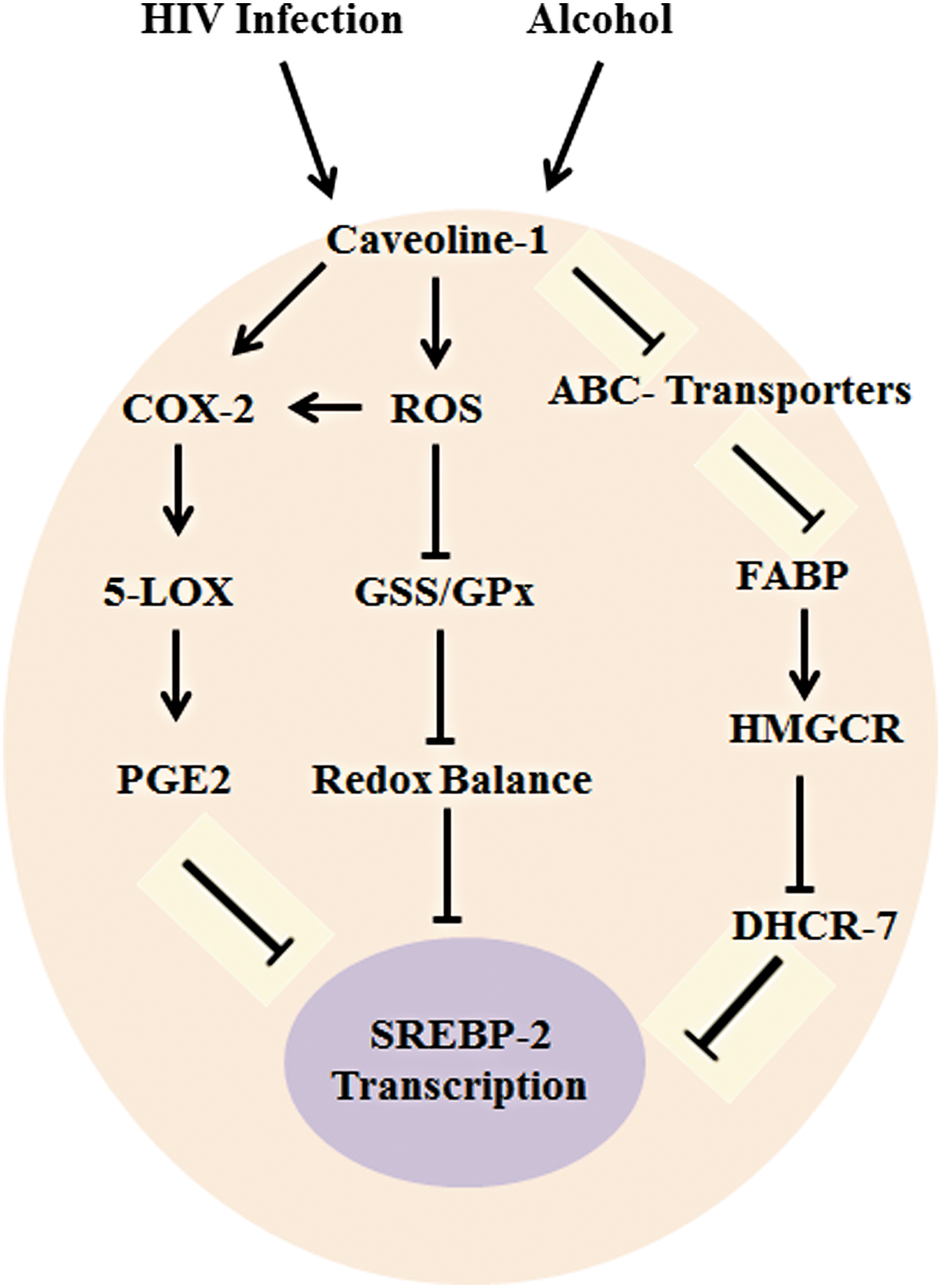
In conclusion, our data provide evidence of immune dysfunction altered by comorbidity of AL and HIV, leading to an association between redox inhibitions and increased AA metabolites, which, subsequently, affect lipid rafts and SREBP-mediated ABC transporters. Based on these results, oxidative stress and AA metabolites potentiate HIV viral replication and disease progression, which may lead to immune toxicity. Therefore, this study supports the molecular basis for increased immune toxicity in HIV-positive alcohol users (49).
Materials and Methods
Human subjects
Blood donors were recruited from the Borinquen Healthcare Center, Inc. (Miami, FL). Consent was obtained consistently with Florida International University (FIU) and National Institute of Health (NIH) policies. The protocol was approved by the institutional review board (IRB) at FIU. Exclusion criteria included polydrug use, hepatitis, other medical conditions, age <18 and >52 years old, and pregnancy. Race/ethnicity of the participants included African American, White, and Hispanic. HIV-1-positive subjects were defined as individuals with CD4+ count ≥500 and plasma viral load <10,000 copies/ml (Table 1). Alcohol users, according to the NIH protocol, were those who reported drinking an average of 7 drinks/day and 5 days/week.
HIV, human immunodeficiency virus.
HIV-1 gp120 recombinant proteins
The HIV-1 BaL gp120 protein was obtained from the NIH AIDS Research and Reference Reagent Program. The recombinant gp120 proteins purity is >95%.
Isolation of PBMCs
PBMCs were separated from patients' blood samples derived from normal, alcohol users, HIV-positive and HIV-positive alcohol users by using the density gradient method previously reported (36, 49 –51).
Treatment of PBMCs with HIV-1 gp120 and alcohol
PBMCs (3 × 106cells/3 ml) were separately treated with an optimized concentration of 50 ng/ml HIV-1 gp120 protein and alcohol 0.1% at 24 h, and untreated cells served as control.
Determination of ROS production
The HIV-1gp120 and alcohol-induced ROS production was analyzed by a fluorescence microplate reader at λex: 485 nm and λem: 530 nm. Briefly, PBMCs (5 × 105) were exposed to either HIV-1 gp120 (50 ng/ml) alone or combined with alcohol (0.1%/ml) for 24 h. At the end of the time periods, 10 μM 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA; Invitrogen, Carlsbad, CA) was added directly to the medium and incubated at 37°C for 30 min followed by washing with phosphate buffered saline (PBS); then, ROS production levels were analyzed respectively.
RNA extraction and real-time quantitative PCR
Total RNA from PBMC was extracted by using the Qiagen kit (Invitrogen Life Technologies, Carlsbad, CA) by following the manufacturer's instructions. The total RNA (3 μg) was used to synthesize the first strand of cDNA. Amplification of this cDNA was performed by using the following specific primers: GSS (Assay ID Hs00609286_m1), SOD1 (Hs00166575_m1) catalase (Hs00156308_m1), COX-2 (Assay ID, Hs00153133), 5-LOX (Assay ID Hs00386528), Cav-1 (Assay ID, Hs00971716), SREBP-2 (Assay ID, Hs01081784), HMGCR (Assay ID, Hs00168352), DCHR7 (Assay ID, Hs01023087), ABCA1 (Assay ID, Hs010159118), ABCG1 (Assay ID, Hs00245154), ABCB1 (Assay ID, Hs00184500), and ABCG4 (Assay ID, Hs00967540); β-actin (Assay ID, Hs99999903) (Applied Biosystems, Foster City, CA) was used as housekeeping gene for quantifying real-time PCR. Relative abundance of each mRNA species was assessed by using the brilliant quantitative PCR master mix from Stratagene by using the Mx3000P instrument that detects and plots the increase in fluorescence versus PCR cycle number to produce a continuous measure of PCR amplification. Relative mRNA species expression was quantitated, and the mean fold change in expression of the target gene was calculated by using the comparative CT method (Transcript Accumulation Index = 2−ΔΔCT). All data were controlled for quantity of RNA input by performing measurements on an endogenous reference gene, β-actin. In addition, results on RNA from treated samples were normalized to results obtained on RNA from the untreated control sample.
Western blot
To assess protein expression, PBMCs (3 × 106) were treated with HIV-1 gp120 (50 ng/ml) and alcohol (0.1%) for 24 h at 37°C. After incubation, cells were washed with PBS and lysed by using lysis buffer (Pierce, IL) with 1 × complete cocktail of protease inhibitors. Extracts were centrifuged at 14,000 g for 5 min at 4°C, and total cellular protein was determined by Bio-Rad protein determination assay (Bio-Rad, Hercules, CA). The equal amounts of proteins were resolved by 4%–15% polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. After blocking, the membrane was probed with primary rabbit polyclonal GSS, GPx (One world lab, CA), SREBP-2, (Cayman, CA) Cav-1, HMGCR (Santa Cruz Biotechnology, CA), DHCR-7 (GeneTex, Inc., Irvine, CA), FABP (Abnova, Taiwan), ABC transporters ABCA1, ABCC1, ABCG1, and ABCC4 (Boster Biological Technology, Pleasanton, CA), and mouse monoclonal SOD antibody (Chemicon International, CA); this was followed by secondary goat antirabbit IgG antibody and goat antimouse IgG antibody (Santa Cruz Biotechnology, CA). Immunoreactive bands were visualized by using a chemiluminescence Western blotting system according to the manufacturer's instructions (Amersham, NJ).
Quantification of PGE2 by enzyme-linked immunosorbent assay
Plasma were separated from normal, alcohol users, HIV-positive and HIV-positive alcohol-using subjects. Separated plasma was analyzed for PGE2 level (GenWay Biotech Inc. San Diego, CA) by using commercially available enzyme-linked immunosorbent assay (ELISA) kits as per the manufacturer's instructions.
Statistical analysis
Data were analyzed by using GraphPad Prism software. Comparisons between groups were performed by using paired t-test and two-way analysis of variance. Differences were considered significant at p ≤ 0.05. Data are expressed as mean ± SE. Experiments were performed at least three times in triplicate.
Footnotes
Acknowledgments
This study was supported by grants from the National Institute of Health (NIH): DA 027049, DA 025576 and MH085259 and NIAAA-R01AA018084.
Author Disclosure Statement
No competing financial interests exist.
Author Contributions
Study conception and design: T.S. and M.P.N.N; Acquisition of data: T.S. and C.T.M.; Human samples analysis: J.G.D.; Analysis and interpretation of data: T.S., C.T.M., and V.S.R.A.; Article edits: M.A. and R.R.; Authoring of the article: T.S. and M.P.N.N. All authors approved the final published version of this article.