
Abstract
Intraorgan communication in the cardiovascular system is exerted not only by direct cell–cell contacts but also by locally released factors, which modulate neighboring cells by paracrine signals (e.g., NO, vascular endothelial growth factor, adenosine, reactive oxygen species). Moreover, cells in close proximity to the typical cardiovascular cells such as fibroblasts, red blood cells, as well as resident and invading immune cells must be considered in attempts to understand cardiovascular function in physiology and pathology. The second level of communication is the interorgan communication, which may be distinguished from intraorgan communication, since it involves signaling from remote organs to the heart and circulation. Therefore, mediators released by, for example, the kidney or skeletal muscle reach the heart and modulate its function. This is not only the case under physiological conditions, because there is increasing evidence that the organ-specific response to a primary insult may affect also the function of remote organs by the release of factors. This Forum will summarize novel mechanisms involved in intraorgan and interorgan communication of the cardiovascular system, with a special view on the remote organs, skeletal muscle and kidney. Antioxid. Redox Signal. 26, 613–615.
T
Hypoxia, hormone and nutrient levels, inflammation, and mechanical load represent multifaceted and divergent stimuli, which cause characteristic adaptive responses of heart and blood vessels. Extracellular signals, released from one cell, induce intracellular signal transduction leading to specific responses of the target cell. Thus, paracrine signaling of parenchymal cells, which affects vascular function, represents the basis for intraorgan communication. On the contrary, also, long-distance interorgan communication exists. Whereas the release of hormones from endocrine organs is an accepted and long-studied phenomenon, there is increasing evidence that novel mechanisms of interorgan communication involve the release of mostly undefined factors by nonprofessional endocrine organs. Triggered by, for example, pathological stimuli, these factors are able to affect the function of remote organs.
Intraorgan communication in the cardiovascular system may be exerted by direct cell–cell contacts involving, for example, connexins in the heart and myoendothelial junctions, and is also achieved by locally released factors that modulate neighboring cells in a paracrine manner (e.g., NO, vascular endothelial growth factor, adenosine, reactive oxygen species [ROS]). It is also evident that cells in close proximity to the “professional” cardiovascular cells such as fibroblasts as well as resident and invading immune cells must be considered in attempts to understand cardiovascular function in physiology and pathology. Cardiac remodeling after prolonged ischemia involves the destruction of contractile myocardium, triggers invasion of inflammatory cells, and activates fibroblasts, leading to formation of extracellular matrix and build-up of a scar. Moreover, altered vascularization and hypertrophy of the remote myocardium give rise to a (partial) functional compensation for the loss of viable myocardial tissue. This example shows that an extensive exchange of information between cardiovascular cells, immune cells, and resident fibroblasts has to be considered in an attempt to understand cardiovascular biology on the organ level.
The second level of communication, termed interorgan communication, may be distinguished from intraorgan communication in that it involves signaling from remote organs to the heart and circulation. Therefore, mediators released by, for example, the kidney, liver, or skeletal muscle reach remote organs, such as the heart, and modulate their function. This is not only the case under physiological conditions, which is illustrated by increasing evidence that the organ-specific response to a primary insult may influence also the function of remote organs by the release of factors such as adipokines or myokines. Signals of interorgan communication may be protective, since ischemia of, for example, skeletal muscle sends protective signals to other organs, including the heart and kidney, resulting in protection of these organs from an ischemic insult, a mechanism termed remote ischemic preconditioning. On the contrary, interorgan communication is the underlying cause of comorbidities known, among others, as the cardiorenal syndrome and cardiac cachexia. As communication routes mediating these pathological forms of organ crosstalk, activation of the sympathetic nervous system, as well as the local production and systemic distribution of proinflammatory cytokines via the circulation, appears to be involved.
Therefore, this Forum on “intra- and interorgan communication in the cardiovascular system—a special view on redox regulation,” addresses recent novel findings on short- and long-distance communication.
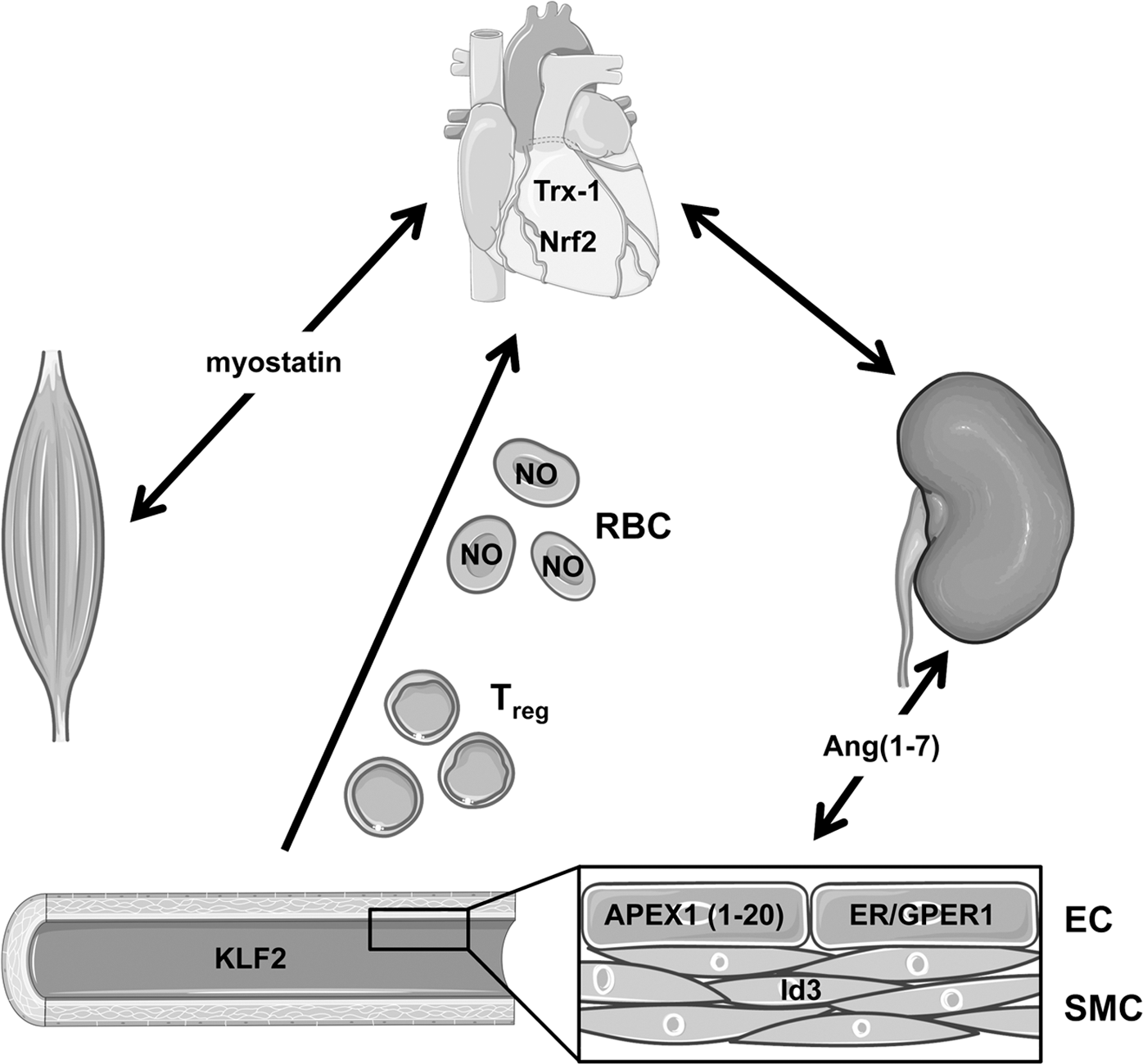
Important changes in intra- and interorgan communication have been attributed to the onset of atherosclerosis and to the molecules involved. Two contributions in this Forum highlight on one hand the role of the redox homeostasis and on the other hand the effects of redox-dependent transcription factors in atherosclerosis (3, 4). Loss of the oxidoreductase thioredoxin-1 expression in the vessels leads to dramatic progression of atherosclerosis and changes in plaque composition (3). Kohlgrüber et al. summarize examples of transcription factors intimately involved in the various processes of plaque formation and progression. Among them are Krüppel-like factor 2, which mediates the response of the endothelium to blood flow, the regulation of vascular smooth muscle cell proliferation and apoptosis by inhibitor of differentiation 3, and the different estrogen receptors and their role in atheroprotection (4).
Another important focus of this Forum is on the mechanisms leading to myocardial infarction and the role of the redox homeostasis and lymphocytes therein. Boag et al. review the important roles of specific subsets of immune cells in the progression of myocardial infarction. The effects of lymphocytes in the ischemic myocardium are time and subset specific, with some having protective effects, while others are pathogenic. A deeper understanding of the involvement of lymphocytes in myocardial ischemia/reperfusion injury may allow the development of targeted therapies, because up to now no effective treatment has been identified (1). Moreover, the protective role of the redox-dependent transcription factor nuclear factor (erythroid-derived 2)-like 2 and the oxidoreductase thioredoxin-1 in ischemia/reperfusion injury is discussed in detail by Jakobs et al., which further underscores a role for ROS in ischemia/reperfusion (3).
A proper endothelial function is required for intra- as well as interorgan communication. Thus, damage to the endothelium has to be prevented. In the original contribution of this Forum, it has been demonstrated that the APEX nuclease (multifunctional DNA repair enzyme) 1 (APEX1) protects the endothelium from ROS-induced apoptosis. This antiapoptotic effect depends on the first 20 amino acids of APEX1 and converges on thioredoxin-1 (2). Another important cell type in the blood are red blood cells, which were for a long time considered to be only transporters of oxygen and nutrients. However, recent studies indicate that red blood cells are important interorgan communication systems with functions including participation in the control of systemic NO metabolism, blood rheology, and viscosity. These noncanonical functions of red blood cells in the cardiovascular system are described by Kuhn et al. in this Forum (5).
With respect to interorgan communication, Leitner et al. summarize mechanisms leading to skeletal muscle wasting. Diseased organs such as the heart, lung, kidney, or cancer tissue secrete soluble factors, including angiotensin II, myostatin (growth differentiation factor [GDF] 8), GDF11, transforming growth factor β, and others, which travel along the vascular system and act on skeletal muscle. There, they induce a set of genes called atrogenes, which induce the ubiquitin–proteasome system, leading to protein degradation. Moreover, elevated ROS levels due to modulation of NADPH oxidases and mitochondrial dysfunction contribute to disease progression, which is characterized by loss of muscle mass, exercise resistance, and frailty (6).
It is well known that the kidney, heart, and blood vessels represent a system of intimately linked regulatory units, which integrate their functions to the setting of blood pressure, organ perfusion, and supply. This relationship is illustrated by the renin–angiotensin system (RAS), being involved in the regulation of blood pressure and volume homeostasis. However, dysregulation of kidney and organ-specific RAS is a major cause of hypertension, elevated ROS formation, and cardiovascular remodeling. Accordingly, the RAS may be considered as part of intra- and interorgan communication. Yang et al. address in their review recent findings on the role of the angiotensin-converting enzyme 2 (ACE2), angiotensin (1 –7) [Ang(1–7)], and its receptor Mas. The short cleavage product of angiotensin II, Ang(1–7), generated by ACE2 exerts protective functions in the vasculature and the kidney by binding to the Mas receptor and appears to counteract angiotensin II-mediated actions in lowering blood pressure, reducing ROS formation, and elevating NO release (7).
Taken together, this Forum will shed light on novel mechanisms relevant for intra- and interorgan communication within the cardiovascular system and to remote organs (Fig. 1).
Footnotes
Acknowledgments
The authors acknowledge the Deutsche Forschungsgemeinschaft (IRTG1902). Single elements for figures were taken from the Powerpoint image bank of Servier Medical Art (