
Abstract
Significance:
Prostate and ovarian cancers are major contributors to cancer-related deaths worldwide. Recently, inflammation and nitrosative stress have been implicated in carcinogenesis, with the overexpression of NOS2 and concomitant release of nitric oxide (NO) associated with cancer initiation and progression.
Recent Advances:
An increasing body of evidence indicates an association between NOS2 expression and aggressive ovarian cancer. Research also indicates a role for NO in prostate disease pathology and prostate cancer. A therapeutic role for NOS2 inhibition and/or NO drugs exists for the treatment of both ovarian and prostate tumors.
Critical Issues:
Herein, we review the key molecular effects associated with NOS2 in ovarian and prostate cancer. NOS2 increases angiogenesis and tumor proliferation and correlates with aggressive type II ovarian tumors. NOS2 expressing tumors are sensitive to cisplatin chemotherapy, and NO may be used to sensitize cisplatin-resistant tumors to chemotherapy. NOS2 is highly expressed in prostate tumors compared to non-neoplastic prostate pathologies. NO may play a role in the development of androgen-independent prostate cancer via s-nitrosylation of the androgen receptor. Moreover, NOS2 inhibitors and NO donor drugs show therapeutic potential in ovarian and prostate cancer as single agents or dual drugs, by either inhibiting the effects of NOS2 or increasing NO levels to induce cytotoxic effects.
Future Directions:
NOS2 and NO present new targets for the treatment of ovarian and prostate tumors. Furthermore, understanding NO-related tumor biology in these cancers presents a new means for improved patient stratification to the appropriate treatment regimen. Antioxid. Redox Signal. 26, 1078–1090.
Introduction
T
Several cellular processes are shared between inflammation and cancer, such as an increased proliferation rate, migration, invasion, angiogenesis, and the generation of reactive oxygen species/reactive nitrogen species (ROS/RNS) (23). Sources of inflammation-related carcinogenesis may arise due to infections, lifestyle, or chronic inflammation in the body. For example, cervical cancer is predominantly as a result of infection with the human papilloma virus (HPV), and prevention strategies for cervical cancer now include vaccination against HPV16 and HPV18 (64). Increasingly there is a body of evidence showing a role for inflammation and in particular the overexpression of inducible nitric oxide synthase (NOS2) and concomitant nitric oxide (NO) release associated with cancer initiation and progression (87).
Before we can consider the impact of NO on tumor biology and metastasis, one needs to understand the concentration effects of NO on the cell. It is well established that NO operates in a bimodal manner. The dichotomous effects of NO on cancer arise from its ability to positively or negatively regulate crucial processes, including proliferation, migration, invasion, survival, angiogenesis, and metastasis, depending on the concentration of NO involved (16, 101). An additional layer of complexity arises when one includes the impact of NO flux and the duration of NO exposure (75, 82, 103). In this review, we discuss the potential role of NOS2 and NO in two cancers arising from female and male reproductive organs that have a major impact globally on cancer incidence and mortality—ovarian and prostate cancer.
Approximately 22,000 cases of ovarian cancer are diagnosed in the United States (11.9 per 100,000) and 44,000 cases in the European Union (EU) (12.6 per 100,000) each year [Source: SEER Program (2) and EUCAN (1)]. Each year, ∼14,000 deaths from ovarian cancer occur in the United States (7.5 per 100,000) and 29,000 deaths in the EU (7.4 per 100,000) (Sources: SEER Program and EUCAN). The 5-year survival rate for women with ovarian cancer is 46%. Regarding prostate cancer, ∼180,000 cases of prostate cancer are diagnosed in the United States (129.4 per 100,000) and 343,000 cases in the EU each year (105.5 per 100,000). While survival rates are better for prostate cancer than ovarian cancer, each year ∼26,000 deaths from prostate cancer occur in the United States (20.7 per 100,000) and 71,000 deaths in EU (Source: SEER Program and EUCAN) (18.9 per 100,000). The 5-year survival rate for men with prostate cancer is 98.9%. For prostate cancer, lethal disease arises when patients develop distant metastatic disease, usually to the bone, generally 5–10 years postinitial diagnosis.
The extent to which NO plays a role in these diseases is not completely understood and represents a gap in the field. It is therefore important to determine if NOS play a role in the progression of these diseases as they may represent new therapeutic targets for their treatment.
Risk Factors Associated with Epithelial Ovarian Cancer
The natural history of the development of ovarian cancer is poorly understood. Ovarian cancers are predominantly diagnosed at a late stage, therefore limiting our ability to identify precursor lesions that may contribute to its development. One hypothesis is that ovarian cancers arise from ovarian surface epithelial cell-lined inclusion cysts that are embedded in the ovarian stroma and their proliferation is promoted by stromal growth factors. Risk factors associated with increased risk of the development of ovarian cancer include nulliparity, while gravidity and contraceptive pill use decrease the risk of developing ovarian cancer (38). This may be partially due to the influence of hormones, but it is also thought that injury due to “incessant ovulation” may play a role, with increased accumulation of p53 mutations correlating with the number of lifetime ovulatory events (Fig. 1). This may, in part, explain the inverse correlation seen between contraceptive use and decreased risk of ovarian cancer (51).

Inflammation arising from sources such as incessant ovulation and endometriosis induces genetic mutations in p53, K-ras, and PTEN, loss of heterozygosity at 1p, 9p, 11q, 17p, and 22q, and altered stromal secretions in ovarian tissues. PTEN, phosphatase and tensin homolog.
Recent research in the field of epithelial ovarian cancer has suggested that ovarian tumors develop via two pathways: type I tumors, which include low-grade serous and endometrioid ovarian tumors, mucinous, clear cell, and malignant Brenner tumors, are less aggressive and are thought to develop from defined precursor lesions, whereas type II tumors, which include primarily high-grade serous or endometrioid tumors, are more aggressive, rapidly metastasize, and have no well-defined precursor lesion within the ovary. Fortner et al. reported an inverse association between contraceptive use and risk of both type I and type II tumors. Full-term pregnancy was only inversely associated with type I tumors (32).
Clearly other factors may be involved in the development of this disease. Inflammatory conditions, including endometriosis and pelvic inflammatory disease, are associated with increased risk of ovarian cancer (93). Evidence suggests that some ovarian cancers arise from endometriosis lesions. Molecular alterations that occur in endometriosis include phosphatase and tensin homolog (PTEN) mutation, loss of heterozygosity of chromosomes 1p, 9p, 11q, 17p, and 22q, and amplification of 17q indicating clonality (Fig. 1). Predominantly clear cell and endometrioid carcinomas arise from endometriosis, and also some serous carcinomas. Major molecular changes associated with the emergence of endometriosis-associated ovarian cancer include p53 mutations (40–50%), PTEN loss of function, and v-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog (K-ras) mutations (70).
Interestingly, single-nucleotide polymorphisms in the nuclear factor kappa-light-chain enhancer of activated in B cell (NFκB) pathway genes IL-1A and TNF-related apoptosis-inducing ligand (TRAIL) have been shown to be associated with the risk of developing clear cell ovarian cancer (18). Circulating levels of the inflammation markers IL-2, IL-4, IL-6, IL-12, and IL-13 have also shown to be associated with increased risk of ovarian cancer (20). In addition in the PLCO screening trial, C-reactive protein, IL-1α, IL-8, and TNF-α are associated with increased risk of subsequently developing ovarian cancer (98). Given the link between ovarian cancer and inflammation, this review explores whether there is a potential role for the inflammatory mediator NO in ovarian cancer.
Biological Role of NOS2 and NO in Ovarian Cancer Progression
NOS2 expression has been shown to be elevated in ovarian tumors (88%) compared to normal ovarian tissue (33%) or benign lesions in the ovary (28%) (77). Trinh et al. reported that the distal-less homeobox 4 (DLX4) protein is associated with increased angiogenesis in ovarian cancer (44). Further investigation showed that DLX4 correlates with high levels of NOS2 gene expression in ovarian cancer (Fig. 2). Overexpression of DLX4 in ES2 and A2780 ovarian cancer cells induced NOS2 gene expression in a signal transducer and activator of transcription 1 (STAT1)-dependent manner, leading to enhanced vascular endothelial growth factor (VEGF) secretion (99). Recent studies have proposed that ovarian tumors can be divided into two groups, type I and type II.
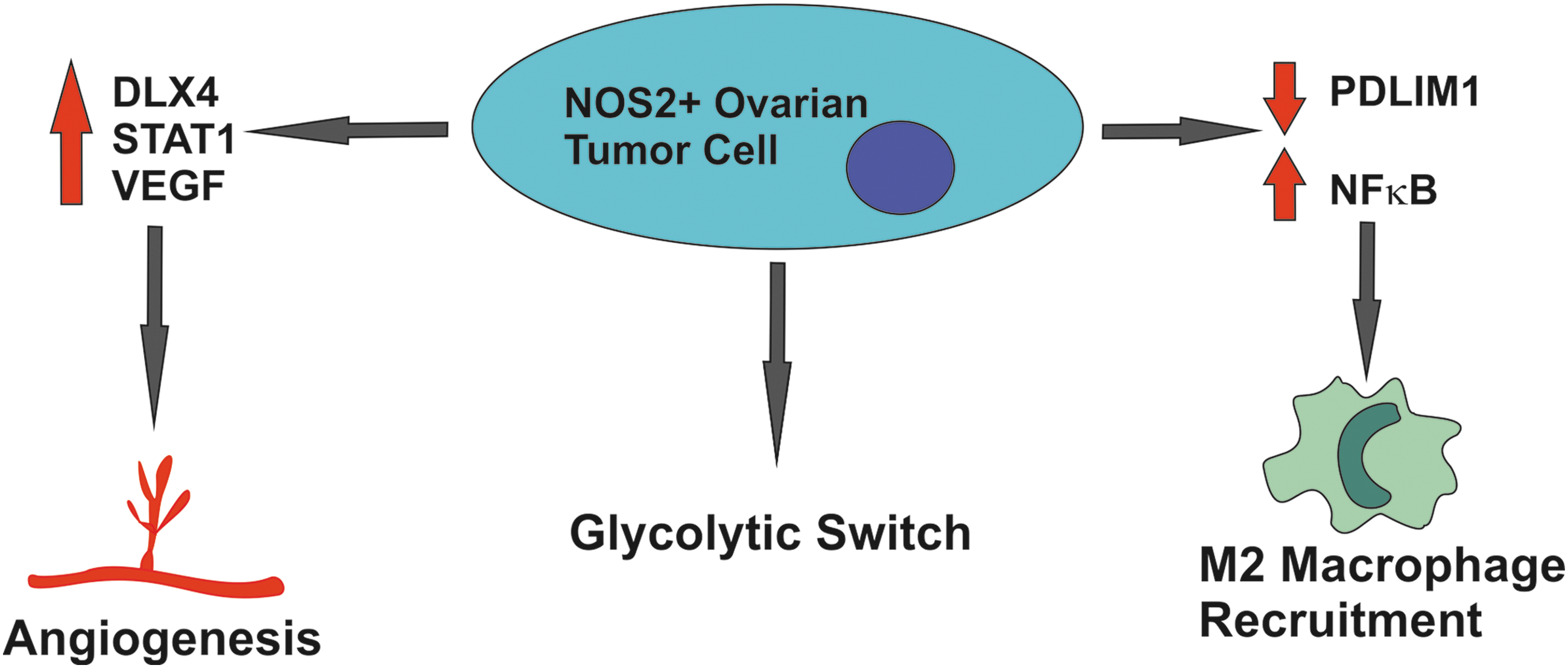
Increased NOS2 expression in ovarian tumors promotes angiogenesis via STAT1 and VEGF, increased rates of glycolysis, and recruitment of M2 polarized macrophages enhancing tumor immunosuppressive capacity. STAT1, signal transducer and activator of transcription 1; VEGF, vascular endothelial growth factor.
Type I are slowly growing tumors that include serous borderline tumors with micropapillary pattern, low-grade serous carcinomas, and mucinous, endometrioid, and clear cell carcinomas. They are genetically stable and driven by mutations in KRAS, BRAF, PTEN, and β-catenin. A subset of type I tumors can exhibit aggressive behavior. In contrast, type II tumors rapidly proliferate and include high-grade serous carcinomas, malignant mixed mesodermal tumors (carcinosarcomas), and undifferentiated carcinomas. They are characterized by accumulation of TP53 mutations and exhibit genetic instability (62).
Ali-Fehmi et al. showed that the inflammatory proteins cyclooxygenase (COX)-1, COX-2, NOS2, and glucose transporter 1 were significantly higher expressed in the aggressive type II compared to type I ovarian tumors, indicating a potential role in type II aggressive tumor biology (5). Nitrate and TGF-β levels correlate in ovarian tissues (56). Elevated nitrate levels predicted ovarian cancer over normal or benign ovarian tissues, and correlated with advancing TNM stage. High levels of TGF-β were predictive of poor outcome (56). In contrast to these findings, Martins Filho et al. failed to find an association between NOS2 levels and advanced ovarian cancer (71).
Mechanisms by which NO may promote ovarian cancer progression include induction of the Warburg effect in ovarian cancer cells. Caneba et al. showed that NOS2 induces cellular proliferation and inhibits mitochondrial respiration in ovarian cancer cells, resulting in a metabolic switch to glycolysis to maintain adenosine triphosphate (ATP) production (Fig. 2). This was accompanied by an increase in tricarboxylic acid (TCA) cycle flux and glutaminolysis (17).
Zhao et al. demonstrated that NOS2 plays a role in the recruitment of M2 macrophages in ovarian cancer. They showed that PDZ and LIM domain protein 2 (PDLIM2) expression is repressed in ovarian tumors leading to hyperactivation of NFκB, due to a decrease in ubiquitination of the p65 subunit (function of PDLIM1) (Fig. 2). This resulted in induction of NOS2 in the tumor epithelia, leading to NOS2-dependent recruitment of M2 macrophages to the tumor (106). siRNA targeting of tumor NOS2 expression resulted in significantly decreased percentage of M2 macrophages in the OVCAR-3 and Caov-3 ovarian cancer cell xenografts (106). The exact mechanism of action of M2 macrophage recruitment or potentially polarization to an M2 phenotype was not explored during this study. However, the study by Ryan et al. in colorectal carcinoma tumors may provide insight into the potential mechanism of action. Ryan et al. demonstrated that tumor epithelial NFκB expression is a key factor in determining the polarization of macrophages within the tumor microenvironment. Using an IκBα “super-repressor (SR)”' loss-of-function vector, NFκB-deficient CT26/IκBα SR xenografts displayed reduced tumor proliferation and reduced metastasis. This was accompanied by polarization of macrophages in the tumor microenvironment from an M2 to M1 NOS2 expression phenotype. Polarization could be recapitulated in vitro using NFκB-deficient CT26/IκBα SR-conditioned media, indicating that an NFκB-dependent secreted factor must mediate this effect (86).
Going forward, further research into the role of NOS2 in ovarian cancer should focus on its association with specific histological subtypes of ovarian cancer, and where possible, its role in precancerous lesions. Given the heterogeneity of ovarian cancer, we may benefit from exploring the ability of NOS2 to predict therapeutic responses or risk of metastatic progression on a histology by histology basis, in addition to comparing type I versus type II tumors.
Therapeutic Effects of Targeting Nitric Oxide in Ovarian Cancer
Inhibition of NOS2 in SKOV and MDAH2774 ovarian cancer cell lines using the NOS2 inhibitor L-NAME reduces the expression of VEGF and reduces the ability of the SKOV and MDAH2774 cell lines to induce angiogenesis (Fig. 3) (69). This indicates that targeting NOS2 in ovarian tumors may inhibit tumor angiogenesis. Ovarian cancer may also be targeted using NO donors at cytotoxic levels. Garban and Bonavida demonstrated that interferon (IFN)-γ sensitizes ovarian carcinoma cell lines to Fas-mediated apoptosis (Fig. 3) (36). Further investigation revealed that IFN-γ treatment induced NOS2 and led to increased nitrite production, and increased Fas-ligand receptor expression. Exogenous NO in the form of S-nitroso-N-acetylpenicillamine (SNAP) was able to recapitulate sensitization to Fas-ligand-mediated apoptosis. Blocking NOS2 activity in IFN-γ-treated cells reduced the levels of apoptosis (36).
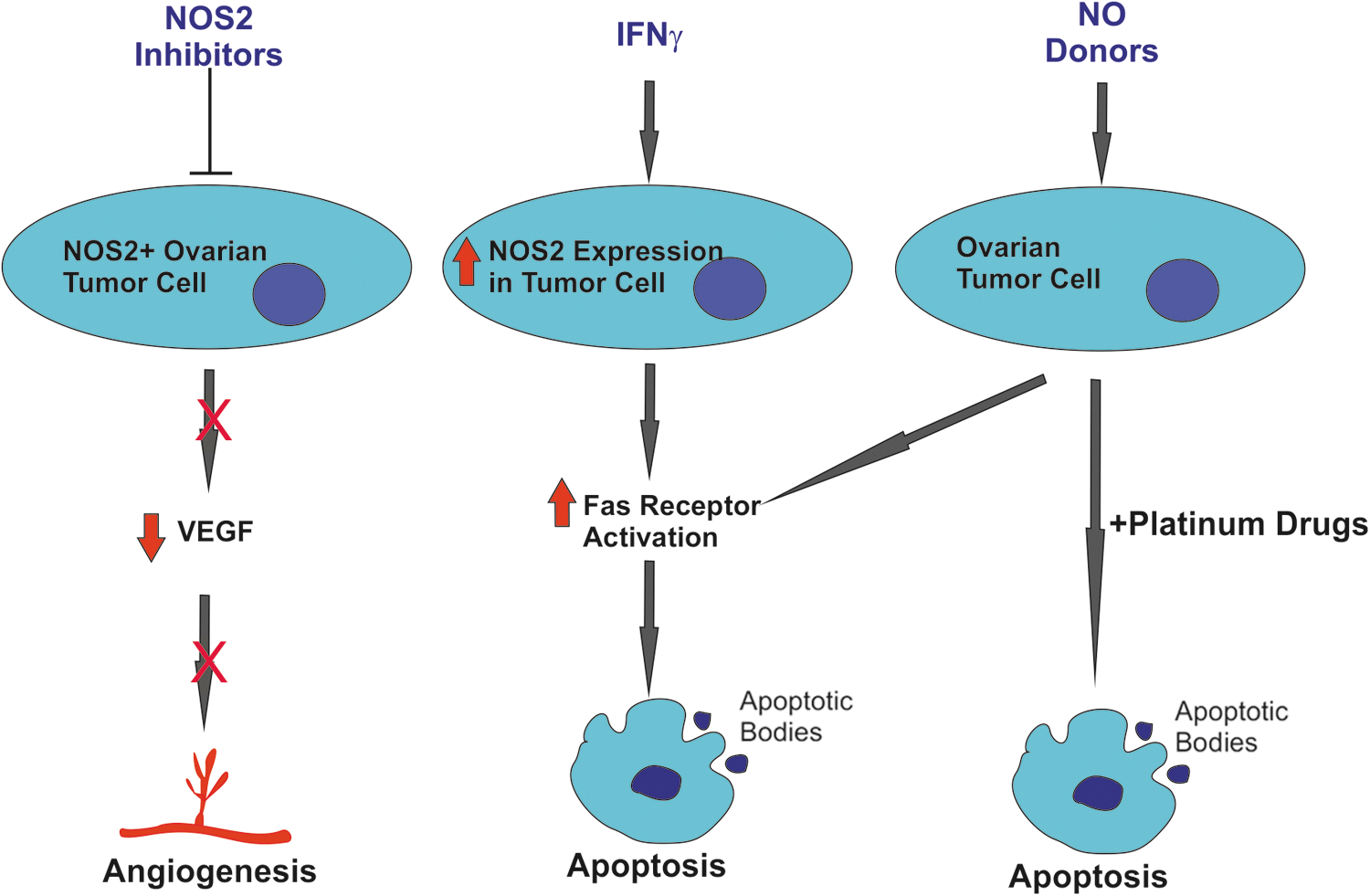
Both NOS2 inhibitors and NO donors can be used to sensitize ovarian tumors to chemotherapy, inhibit angiogenesis, and to induce Fas-ligand-mediated apoptosis.
However, the role of IFN-γ in ovarian cancer is complex. For example, IFN-γ secretion by CD8-positive lymphocytes has been shown to upregulate the immune checkpoint inhibitor PD-L1 on ovarian cancer cells and to promote tumor growth (4). Conversely, loss of the IFN-γ receptor in ovarian cancer has also been shown to be associated with poor outcome in ovarian cancer (27). Further research into NO and Fas-mediated apoptosis demonstrated that NO inhibited the transcription repressor Yin-Yang 1 (YY1) binding activity at the silencer region of the Fas receptor promoter, demonstrating a mechanism for NO induction of the Fas receptor (35). Kielbik et al. showed that the NO donors, spermine nitric oxide complex hydrate (SPER/NO, t 1/2 = 39 min) and diethylenetriamine/nitric oxide adduct (DETA/NO, t 1/2 = 20 h) induced apoptosis in SK-OV-3 and OVCAR-3 ovarian carcinoma cell lines at high concentrations, particularly in DETA/NO with its longer half-life (57). The NO donors induced apoptosis through mitochondrial membrane depolarization and caspase-3 activation (57).
El-Sehemy et al. demonstrated that Notch3 increases the expression of GUCY1B3 in noncancerous immortalized ovarian surface epithelial cell lines. GUCY1B3 is the β subunit of soluble guanylyl cyclase, which is known to mediate the effects of NO. Increased expression of GUCY1B3 in the immortalized ovarian surface epithelial cells resulted in increased NO-induced cyclic guanosine 3′,5′-monophosphate (cGMP) production resulting in phosphorylation of the vasodilator-stimulated phosphoprotein. Examining OVCA429, OVCAR-3, and SKOV3 ovarian cancer cell lines, a correlation was seen between high levels of NOTCH3 and GUCY1B3 expression (28).
Findlay et al. designed an NO donor drug, O(2)-[2,4-dinitro-5-(N-methyl-N-4-carboxyphenylamino) phenyl] 1-N,N-dimethylamino)diazen-1-ium-1,2-diolate (PABA/NO), which is dependent on expression of glutathione S-transferase (GST)-π for its activation. PABA/NO induced a significant growth arrest in A2780 human ovarian tumors in SCID mice, comparable to treatment with the chemotherapeutic agent cisplatin (Fig. 3) (31). Interestingly, Leung et al. (2008) subsequently showed that cisplatin-sensitive ovarian cancer cell lines have higher basal levels of NOS2, which was further increased with cisplatin treatment, which cisplatin-resistant cell lines did not. The NO donor SNAP increased p53 protein levels and enhanced cisplatin-induced apoptosis in the resistant C13(*) cells in a p53-dependent manner, while the NOS2 inhibitor 1400 W partially blocked cisplatin-induced apoptosis in OV2008 cells (65). This would suggest that ovarian tumors with high NOS2 expression may be a good target for platinum drugs, and that tumors with low NOS2 expression could be sensitized to cisplatin-based therapies by including NO donating drugs in the treatment regimen.
Given that type II ovarian cancer is found to have increased expression of NOS2 but is known to acquire platinum resistance, research exploring whether NOS2 expression is predictive of initial response to platinum is needed (intrinsic sensitivity). It would be of interest to determine whether acquisition of acquired resistance to platinum is associated with loss of NOS2 expression. One avenue would be to measure the levels of NOS2 in pre-neoadjuvant biopsies and in postsurgical specimens. Where residual disease remains, is there a loss of NOS2 expression and could NO donors be used to resensitize tumors to chemotherapy? As of yet this is unknown.
Prostatitis, Inflammation, and Risk of Prostate Cancer
Prostate cancer is a leading cause of cancer-related death in men worldwide. Inflammation has been implicated in prostate carcinogenesis, with the overexpression of NOS2 and concomitant NO release associated with cancer initiation and progression (87). Inflammatory conditions of the prostate include benign prostatic hyperplasia (BPH), proliferative inflammatory atrophy, and prostatic intraepithelial neoplasia (PIN), all of which have been linked to the development of prostate cancer (24, 72).
Prostatitis is an inflammatory condition of the prostate gland, which is classified into four categories, three symptomatic and one nonsymptomatic (60, 79). Acute bacterial prostatitis (category I) occurs as a result of bacterial infection (e.g., Escherichia coli) (47, 60, 79). Category II or “Chronic Bacterial Prostatitis” occurs as a result of a chronic bacterial infection (47, 60, 79). Category III prostatitis is often referred to as “chronic nonbacterial prostatitis” or “pelvic pain syndrome” and is the most common form of prostatitis, accounting for greater than 90% of cases (47). Category III is further subcategorized into an inflammatory cell presenting group (IIIa) or a group void of leukocytes (IIIb), with both groups experiencing symptoms for weeks to possibly years (60, 79). Therapies include antibiotics, nonsteroidal anti-inflammatory drug (NSAID) medication, and 5α-reductase inhibitors, but are of limited value in improving symptoms (12, 66). Finally, Category IV is referred to as asymptomatic prostatitis and is diagnosed by histological means upon pathological investigation on the prostate tissue of nonsymptomatic men (39, 76, 79).
Murine models of chronic nonbacterial autoimmune-associated prostatitis displayed elevated levels of circulating TNF-α and elevated expression of NOS2 mRNA in prostate glands, indicating a potential role for NOS2 in chronic nonbacterial prostatitis (108). Comparing type IIIa to IIIb patients, those with type IIIa display elevated levels of NO compared to type IIIb patients, indicating a link between NO and inflammation (46). NO may also play a role in patients with chronic bacterial prostatitis, as patients were found to have higher levels of plasma NO. NO levels were found to be increased with prolonged disease (107). Indeed, prostate epithelial cells can react to bacterial stimuli such as lipopolysaccharide, activating toll-like receptor 4 (TLR4) and inducing NOS2 expression and NO secretion (37).
It has been speculated that a correlation exists between prostatitis and prostate cancer. Inflammation in the prostate was shown to accelerate prostate cancer progression by alteration of the prostatic microenvironment in a chronic prostatitis Hi-Myc mouse model (90). Another study showed that bacterial prostatitis accelerated 2-amino-1-methyl-6-phenylimidazo [4, 5-b] pyridine (PhIP)-induced preinvasive lesions in the rat prostate (88). Sutcliffe et al. found a significant association between prostatitis and prostate cancer among unscreened men, but not among screened patients in the Health Professionals Follow-up Study (95). Also, there was a significant correlation between prostatitis and prostate cancer in the younger men (<59 years of age), and an inverse association between prostatitis and prostate cancer in older men (95).
In an attempt to address the issues that have limited the outcomes of many prostatitis studies, Cheng et al., performed a sizeable prospective cohort study that included representation of various ethnicities, which included African American, Asian American, Latino, and white American men between the ages of 45 and 69, taking into account family history of prostate cancer, body mass index, education, smoking, ejaculation history, use of sildenafil, sexual orientation, diabetes, and testosterone use. This investigation found a positive correlation (1.3-fold increased risk) between prostatitis and prostate cancer regardless of prostate-specific antigen (PSA) testing, suggesting that the influence of detection bias has minimal effects on the significance between the two diseases (19). Together, this suggests a link between inflammation and increased risk of the development of prostate cancer. The extent to which inflammation-associated NO may play a role in the process is not yet known and represents a gap in the field.
BPH, Inflammation, and Risk of Prostate Cancer
BPH is an inflammatory condition characterized by enlargement of both the stromal and glandular constituents of the prostate. BPH is found in ∼70% of men at the age of 70 with prevalence increasing to ∼100% by the age of 80, particularly in the western world (83). Several risk factors suggest a link between BPH and prostate cancer, for example, their increasing prevalence with age, dependence on androgens for their growth and development, and response to antiandrogen therapy (14).
Although previous studies have demonstrated an increased risk of prostate cancer and prostate cancer-related mortality in men with BPH, the nature of this relationship remains unclear (78). Nitric oxide plays several key roles in BPH and its side effects. Neuronal nitric oxide synthase (nNOS) localizes to the slender varicose nerve fibers of the smooth muscle bundle in the prostate and is a key mediator of prostate function (100). Two major side effects of BPH are erectile dysfunction and lower urinary tract symptoms (LUTS). Patients with obstructive BPH exhibit signs of reduced NO-mediated signaling via cGMP. This is due to increased enzymatic conversion of cGMP to its inactive form GMP via the phosphodiesterase type 5 (PDE-5) enzyme. Therapeutically, this is treated using PDE-5 inhibitors, which lead to increased cGMP, thus potentiating the effects of NO and alleviating the symptoms of erectile dysfunction and LUTS. Recently, one study showed that PDE-5 inhibitors may increase the risk of biochemical recurrence after radical prostatectomy for the treatment of prostate cancer, suggesting that increased NO levels have adverse effects on prostate cancer outcomes (73), while another study failed to find any association with biochemical recurrence (34). Whether the use of PDE-5 inhibitors in BPH increases the risk of developing prostate cancer is not currently known.
Biological Role of NOS2 in Prostate Cancer Development and Progression
While the role of nitric oxide is well understood in the carcinogenesis of breast cancer (15, 40, 97), colorectal cancer (6, 7), and melanoma (42), it is largely unexplored in prostate cancer. Early investigations of immunohistological staining patterns for NOS2, endothelial nitric oxide synthase (eNOS), and nNOS in the 1990s indicated a potential role for NOS2, but little research has been performed in the intervening years (41, 58). Baltaci et al. further explored NOS2 in BPH, PIN, and prostate carcinoma. Both high-grade PIN and prostatic carcinoma samples had more intense NOS2 immunostaining than low-grade PIN and BPH samples, indicating a potential increase in NOS2 expression during prostate carcinogenesis (10).
Mechanisms by which NO and its derivative peroxynitrite induce inflammation-associated carcinogenesis include induction of DNA damage, suppression of DNA repair enzymes, posttranslational modification of proteins, enhancement of cell proliferation, angiogenesis, metastasis, inhibition of apoptosis, and antitumor immunity (52, 61). Peroxynitrite can form DNA damaging 8-nitroguanine, a biomarker of inflammation-associated cancers (61). During inflammation, RNS and ROS are released by activated inflammatory cells, attacking neighboring epithelial and stromal cells, altering function, and initiating carcinogenesis (105). NOS with nicotinamide adenine dinucleotide phosphate oxidase and COX mediate hormone-induced oxidative/nitrosative stress in the rat, which leads to transformation of prostate epithelia into dysplasia (96). Another postulated mechanism is NO-mediated activation of COX-2 and the induction of CXC chemokine receptor 4 (CXCR4) (Fig. 4) (8). Inflammation-derived nitric oxide also plays a role in tumor progression, for example, through facilitating vascular permeability, thereby supporting rapid tumor growth (67). Prostate cancer PC-3 cells were shown to acquire increased survival, proliferation, migration, and invasion on receiving photodynamic therapy, through the upregulation of NOS2 and subsequent release of NO (13, 29).
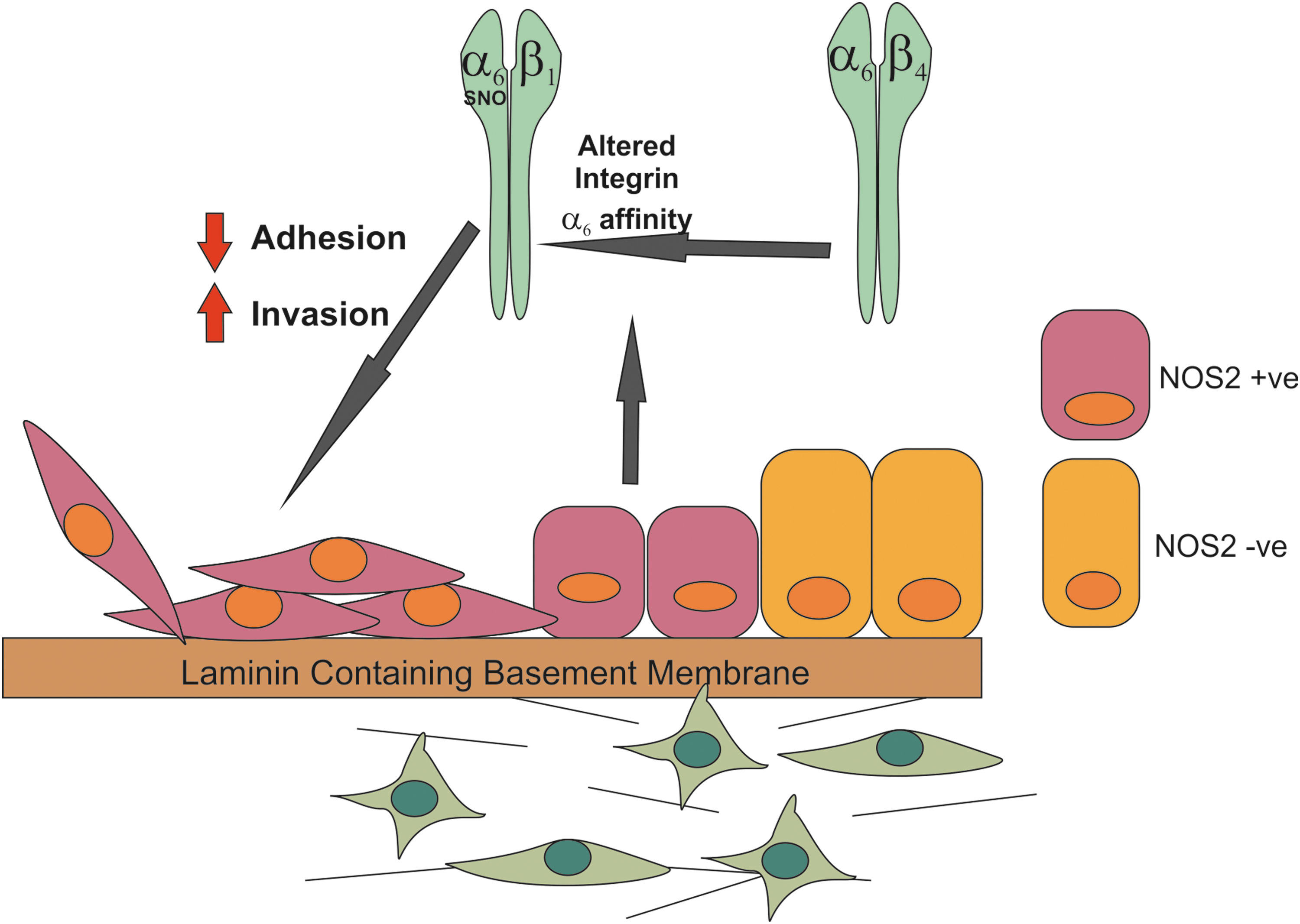
Increased NOS2 expression in prostate tumors promotes increased migratory capacity via s-nitrosylation of integrin-α6.
As mentioned previously, NOS2 is expressed in prostate tumors (58). Baltaci et al. found it was elevated in prostate cancer specimens, compared to PIN or BPH. However, no association with tumor grade was found in the 20 samples tested (10). In a larger study by Aaltoma et al., which included 198 prostate cancer specimens, an association of NOS2 was found with tumor stage, high Gleason score, DNA aneuploidy, and perineural infiltration (3). Expression of NOS2 in prostate cancer may be implicated in the development of androgen-independent prostate cancer, as NO inhibits the androgen receptor (AR) activation and proliferation of AR-positive prostate cancer cells, but does not impact on AR-negative prostate cancer proliferation (21). Intriguingly, in breast cancer, we have shown that NOS2 is predictive of poor outcome in estrogen receptor (ER)-negative tumors but not in ER-positive tumors (40). Whether there is an interaction between AR status and NOS2 expression in prostate cancer and its influence on patient outcomes has not yet been investigated. Increased intracellular NO in AR-positive prostate cancer cells leads to s-nitrosylation and inactivation of AR at the C601 residue present in the DNA-binding domain of the AR, thus inhibiting its ability to respond to androgen (Fig. 5) (80).

S-nitrosylation of the androgen receptor inhibits its activity and may promote the development of androgen-independent disease.
NO may also promote cell migration in prostate cancer cells by the s-nitrosylation of integrin-α6 at Cys86, which enhances its heterodimerization with integrin-α1. In addition the s-nitrosylation of integrin-α6 resulted in decreased ability to adhere to laminin-1 (Fig. 4) (53). This posttranslational modification results in a switch in the interaction of integrin-α6 with integrin-β4 to an interaction with integrin-β1 (53). This is frequently observed in the transition to prostate cancer and results in the disruption of the adherence of prostate cells to laminin via highly adhesive hemidesmosome structures. This allows the cells to migrate on laminin, which is key in the context of promoting prostate cancer metastasis in the laminin-rich bone microenvironment, which is the primary target site of prostate cancer distant metastasis (91). Further research is required to determine whether NOS2 is associated with the development of metastatic prostate cancer. In particular, if it is predictive of early biochemical recurrence, perineural invasion, and therapeutic responses. Ideally, an appropriately designed molecular epidemiological study would greatly add to our knowledge.
Therapeutic Potential of NO-Related Therapies in Prostate Cancer
NO has been exploited as an anticancer therapy for some time. The role of NO in cancer biology is extremely complex and it has been shown to elicit both pro- and antitumorigenic effects, the outcome dependent on the concentration, source, and cellular context of NO involved (16). In general, low levels of NO (<100 nM) are considered to be pro-oncogenic, medium NO levels prometastatic, and high NO levels (>500 nM) tumoricidal (8). This has led to the use of NOS2 inhibitors and exogenous NO delivery therapeutically, with the latter resulting in chemo- and radiosensitization in addition to overcoming drug resistance by tumor cells (11).
In a phase II study, low doses of NO releasing glyceryl trinitrate (GTN) reduced hypoxia-induced progression in prostate cancer patients following primary treatment failure, as measured by PSA doubling time (Fig. 6) (89). The NO donor DETA/NO was shown to sensitize prostate cancer cells to TRAIL-mediated cellular apoptosis at concentrations of 500–1000 μM through the inhibition of NFκB signaling and Bcl-XL expression (Fig. 6) (49). Another NO donor sodium nitroprusside was shown to have a synergistic effect with the selective epidermal growth factor receptor tyrosine kinase inhibitor ZD1839 on the inhibition of cell proliferation in LNCaP, DU145, and PC-3 prostate cancer cells (74).

High-dose NO therapy reverses P-glycoprotein-mediated drug resistance, sensitizes tumors to platinum-based chemotherapy, and sensitizes tumors to TRAIL-mediated apoptosis. Low-dose NO therapy enhanced tumor permeability for chemotherapy and inhibits HIF1α. TRAIL = TNF-related apoptosis-inducing ligand.
NO-donating NSAIDs (NO-NSAIDs) are traditional NSAIDs, which have been modified to include an NO-releasing moiety via a linking spacer (59). NO-ibuprofen and NO-aspirin have previously been shown to inhibit proliferation and induce apoptosis in the androgen-dependent (LNCaP) and androgen-independent (PC-3) prostate cancer epithelial cell lines (85). The NO-NSAID, NO-sulindac, also displayed similar properties in prostate cancer cells, while sulindac alone failed to elicit any cytotoxic effect (50). Stewart et al. went on to demonstrate that the cytotoxicity of NO-sulindac was partially attributed to the NO releasing moiety, and that the drug also inhibited the hypoxic response and invasiveness of the cells via PI3K/Akt signaling (92).
NO has shown promising results in potentiating the effects of chemotherapeutics and targeted therapies. NO releasing GTN has been used extensively as an anticancer agent, primarily due to its vasodilatory effects, which allow targeted delivery of anticancer drugs to tumor tissues, due to enhanced permeability and retention in solid tumors (94). However, additional mechanisms of actions have also been demonstrated. GTN attenuated hypoxia-mediated resistance to chemotherapeutic agents doxorubicin and paclitaxel in DU145 prostate cancer cells, as well as in a nude mouse xenograft model of PC-3 cells, where the combination of doxorubicin and GTN inhibited tumor growth to a greater extent than either alone (33). Combination treatment of the NO donor DETA/NO with cisplatin-diamine-dichloro platinum resulted in tumor growth inhibition in mice bearing human prostate cancer xenografts (Fig. 6) (48). Jia et al. demonstrated that the NO donor, S-nitroso captopril, increased the transmembrane permeability of LNCaP and PC-3 cells to paclitaxel, enhancing its cytotoxicity (55).
NO donors can also modify the activity of drug efflux pumps. Saquinavir-NO has been shown to sensitize PC-3 prostate cancer cells to P-glycoprotein substrates vincristine, actinomycin D, and paclitaxel but most likely by competing with the drugs for efflux through P-glycoprotein (Fig. 6) (84). Also, the NO donor sodium nitroprusside has been shown to inhibit P-glycoprotein efflux of vincristine mouse brain blood vessel endothelial cell clone 4 cells, indicating NO can also effect p-glycoprotein pump activity (68). In cases where tumors exploit and benefit from NO present in the microenvironment, NOS inhibitors can be used to block NO production, thereby slowing tumor growth (45).
Induction of NOS2 in response to 5-aminolevulinic acid-based photodynamic therapy (PDT) may act as a mechanism of resistance to PDT. The surviving fraction of PC-3 and DU145 cells post-PDT displayed increased expression of NOS2 and NO, which increased their capacity for migration, invasion, and proliferation (13, 29). Likewise, NO released by repeated suboptimal PDT treatments induced epithelial to mesenchymal transition in PC-3 cells via increased NFκB and YY1 and decreased Raf-1 kinase inhibitory protein (RKIP) (26). These side effects of PDT were abrogated with NOS2 inhibitors (26, 29). The use of NOS inhibitors in combination with more conventional treatments such as radiation has also been investigated. Tumor growth delay was observed when L-NAME was administered following radiation treatment in a syngeneic squamous cell carcinoma mouse model, with this effect being attributed to Th1 polarization within the tumor microenvironment (81).
Conclusion
Evidence suggests a role for NOS2 in the development of aggressive ovarian cancer. However, further research is required to elucidate its association with specific histological subtypes and more complex epidemiological analysis is needed to assess the strength of its association, ability to predict metastatic potential, and how it correlates with different clinical pathological features of ovarian cancer. Intriguingly, NOS2 expression may predict response to platinum-based therapies. This could have positive consequences for ovarian cancer patients with BRCA mutations and also triple negative breast cancer patients with basal-like phenotypes [which can display impaired BRCA function (25)] or BRCA mutations. BRCA mutations have been shown to sensitize tumors to platinum-based therapies both in ovarian (22, 102) and triple negative breast cancer (54). Cotreatments with NO donors and platinum drugs may further sensitize these tumors to platinum-containing regimens, by enhancing the effects of platinum drugs. Regarding prostate cancer, substantial further research is needed to elucidate the exact role of NOS2 in patient progression. We do not currently know whether NOS2 predicts the development of metastatic prostate cancer or the development of clinically relevant castrate-resistant metastatic prostate cancer, which are refractory to androgen deprivation therapy.
NO has been shown to have biphasic effects in many cancer-related processes, such as cellular proliferation, apoptosis, angiogenesis, migration, and invasion. The effects elicited by NO are concentration, source, and cellular context dependent and can both promote and inhibit carcinogenesis (104). In summary, while NO has shown much promise as an anticancer therapy, other studies have demonstrated that NO may also play an important role in prostate cancer development and progression. Therefore, careful consideration must be used in the use of NO donor drugs as chemotherapeutics in the treatment of cancer. The cytotoxic nature of NO is beneficial in this context; however, cells surviving NO-based treatment, but having been exposed to NO, may harbor protumorigenic characteristics and further promote tumor progression and/or chemoresistance. Therein lies the dual face of NO in cancer biology.
Footnotes
Acknowledgments
This work was supported by the Irish Cancer Society. Sharon Glynn is also funded by Breast Cancer Now and the Galway University Foundation.