
Abstract
Significance:
Peroxiredoxins (Prxs) are thiol peroxidases with multiple functions in the antioxidant defense and redox signaling network of the cell. Our progressing understanding assigns both local and global significance to plant Prxs, which are grouped in four Prx types. In plants they are localized to the cytosol, mitochondrion, plastid, and nucleus. Antioxidant defense is fundamentally connected to redox signaling, cellular communication, and acclimation. The thiol–disulfide network is central part of the stress sensing and processing response and integrates information input with redox regulation.
Recent Advances:
Prxs function both as redox sensory system within the network and redox-dependent interactors. The processes directly or indirectly targeted by Prxs include gene expression, post-transcriptional reactions, including translation, post-translational regulation, and switching or tuning of metabolic pathways, and other cell activities. The most advanced knowledge is available for the chloroplast 2-CysPrx wherein recently a solid interactome has been defined. An in silico analysis of protein structure and coexpression reinforces new insights into the 2-CysPrx functionality.
Critical Issues:
Up to now, Prxs often have been investigated for local properties of enzyme activity. In vitro and ex vivo work with mutants will reveal the ability of Prxs to interfere with multiple cellular components, including crosstalk with Ca2+-linked signaling pathways, hormone signaling, and protein homeostasis.
Future Directions:
Complementation of the Prxs knockout lines with variants that mimic specific states, namely devoid of peroxidase activity, lacking the oligomerization ability, resembling the hyperoxidized decamer, or with truncated C-terminus, should allow dissecting the roles as thiol peroxidase, oxidant, interaction partner, and chaperone. Antioxid. Redox Signal. 28, 609–624.
Biochemical Features and Dynamics of Peroxiredoxins
T
Circumstantial evidence indicates that Prxs are ancient high-affinity enzymes for peroxide detoxification when atmospheric oxygen still was very low (31). After formation of the sulfenic acid derivative by reaction of CysP with the peroxide substrate, the CysP thiol is subjected to various redox modifications ranging from disulfide, sulfinic to sulfonic acid derivatives. The site of the resolving cysteine (CysR) and the quaternary interaction interfaces define the distinct forms of Prxs that have emerged during evolution and exhibit differential specificities toward peroxide substrates, distinct subcellular localization, and regulatory capabilities. The often highly abundant Prxs stabilize redox homeostasis, transmit redox information, and control cell metabolism, in particular, photosynthesis.
Recent research focused on the spatial and temporal occurrence of post-translational modifications (PTMs) of Prxs and their mechanistic coupling to redox reaction chains. Oxidations on thiols occur with increasing oxidation number starting with disulfide (S–S), sulfenic (–SOH), sulfinic (–SO2H) to sulfonic (–SO3H) acid. Except for the sulfonic acid, each of these modifications bears functional significance. Reduction is needed for regeneration (26, 106).
Disulfide formation in 2-CysPrx causes a major conformational switch because the C-terminus carrying the CysR residue has to locally unfold and move to the sulfenylated peroxidatic cysteine. This mechanism destabilizes the dimer–dimer interface resulting in loss of decamer integrity (121). This effect can be analyzed by size exclusion chromatography (SEC) wherein disulfide-bridged oxidized Arabidopsis 2-CysPrx separates as dimer, whereas the reduced form separates as decamer and dimer (62). The oxidized dimers may ease rereduction by NADPH-dependent thioredoxin reductase C (NTRC) or thioredoxins (Trxs) such as Trx-x, CDSP, or Trx-m (26, 96).
Hyperoxidation of the CysP to sulfinic acid in the H2O2 detoxification cycle occurs with a certain frequency of about 1 out of 250 turnovers for Arabidopsis thaliana 2-CysPrx when exposed to a constant H2O2 generation rate of 1 μM H2O2 min−1 from a glucose oxidase system (62). Hyperoxidation opens up additional possibilities for signaling because the hyperoxidized decamer adopts a more stable conformation than the reduced decamer and at least in vitro acts as chaperone preventing precipitation of citrate synthase and insulin at high temperature and reducing conditions, respectively (59). Moreover, S-nitrosylation (36, 104), phosphorylation (see Table 2), Cys-glutathionylation (84), and Lys-acetylation (83) are reported PTMs of Prxs, which, in turn, alter Prx properties. Thus activity and function of Prxs are integrated into multiple signaling pathways as discussed hereunder.
Basically, there exist two common but in fact highly related classifications of Prxs. Prxs are assigned to six distinct groups based on their amino acid sequence, cysteine localization, interaction interface, and N-/C-terminal extensions. Classification and naming of Prxs changed along with novel discoveries, improved analytical methods, and also depending on the organism. Table 1 provides an overview on the Prx nomenclature. The basic classification follows that of (49). Additional information was retrieved from the Prx classification index (PREX) and concerned sequence and structural features (111). Types A, B, C, and D Prxs are present in plants, whereas types E and F are only found in some bacteria. Prxs are often classified in typical 2-cysteine (type A), typical 1-cysteine (type B), and atypical 2-cysteine Prxs, which cover most Prxs. The latter are separated into PrxQ (type C) and type II Prx (type D). Additional features distinguishing Prx types concern the regeneration and subunit composition and interaction mechanisms. The assignment considers the position of the peroxidatic nucleophilic cysteine (CysP) and the presence, position, or absence of a CysR capable of reacting with oxidized CysP (sulfenic acid). The minimal unit can be monomer or dimer. In type A Prx, dimers form intermolecular disulfide bridges (+CysR). Dimers of type B Prx lack the CysR and are linked noncovalently. Type C monomers carry a CysR and generate intramolecular disulfide bonds. Type D Prxs form intramolecular disulfide bonds with CysR as well and noncovalently linked dimers. Trxs, NTRC, glutaredoxin (Grx)/glutathione (GSH), or other electron donors mentioned in the text restore the catalytically active CysP in Prxs belonging to types A, C, and D. In the CysR-free type B Prx, the sulfenic acid immediately reacts with a thiol or another reductant such as ascorbate. The review by Dietz (31) covers many aspects of the Prx family in plants. A genome-wide search allowed us to define the minimum set of six Prxs expressed in all higher plants [31], namely one 1-CysPrx related to embryogenesis, one chloroplast PrxQ, one plastid 2-CysPrx, and one each of cytosolic, mitochondrial, and plastidic type II Prxs. Variation of this minimum set of Prxs frequently occurs with multiple isoforms such as two 2-CysPrxs and three cytosolic PrxIIB-D in A. thaliana, but two Prx II in poplar (PrxIIB, PrxIIC) and only one in rice (PrxIIB).
Based on Hofmann et al. (49), Prxs were sorted with regard to their respective biochemical property of peroxidative cysteines and structure. In addition to the two plant model organisms, the newly sequenced genome of Beta vulgaris was included as additional example (51). For further information concerning subcellular localization, function, biochemical properties, and crystallographic data of annotated proteins, see the respective Uniprot entry (1) or reference sequence for B. vulgaris (2). Subcellular localization is given in parentheses as follows: nucleus = n, cytosol = ct, chloroplast = cl, mitochondria = mt, endoplasmatic reticulum = er, peroxisome = pr, not known = —. An * indicates manual assertion inferred from sequence similarity.
All Prxs and glutathione peroxidases (Gpxs) have a highly conserved structure based on the typical Trx-like fold. Based on this observation, the hypothesis emerged that all Prxs have a common Trx ancestor. The proper Trx-like fold is built from a βαβββα structure (97). The expanded structure of Prxs consists of seven-stranded β-sheets surrounded by five α-helices in β1-β2-α1-β3-α2-β4-α3-β5-β6-α4-β7-α5 topology. The properties of the catalytic center composed of the CysP in its conserved environment were analyzed by crystallography, bioinformatics, and activity assays. These amino acids are arranged in a reaction pocket between sheets β3 and β6, helix α2, and the loop connecting α2 and β3 (81). This position is conserved whereas the resolving Cys is variably positioned in helices α2, α3, and α5 or in the C-terminal region for intermolecular resolving reaction (91). The physicochemical properties of the specified amino acid residues combined with a nucleophilic displacement reaction (SN2) provide a framework for the enzymatic mechanism (Fig. 1). The Arg and Thr/Ser together with the Cys thiolate support the binding of peroxides. The oxygen–thiolate interaction polarizes the terminal oxygen to facilitate the Cys–S–O bond formation. The hydrophobic Pro protects the highly active cysteine site from exposure to the solvent and promotes the high peroxide specificity. Amino acid sequence comparison between At2-CysPrx, PsPrxIIF, and BtPrx3 underlines a conserved positioning for flanking amino acids and peroxidative cysteine in different Prxs and across species such as BtPrx3 102PXXXTXXCX…XR185, PsPrxIIF 80PXXXTXXCX…XR165, and At2-CysPrx 162PXXXTXXCX…XR195. Around 83% of all CysP display a pK a between 5.1 and 6.3 and thus are present in the thiolate form (Cys-S−) at physiological pH of, for example, 7.2 in the cytosol (93). The nucleophilic characteristics of the catalytic center originating from deprotonation at low-pH value (low pK a) generates a rather efficient peroxide detoxification microenvironment with high substrate affinity in the low micromolar range (39, 93). However, the turnover number is low with 10–250 reactions/min. The catalytic efficiency of Prxs is in the range of 105–108 M −1 s−1 and depends strongly on the thiol regeneration mechanism (96). The variation by three orders of magnitudes points to diversification of Prxs across all kingdoms. As shown for AhpC in Salmonella (86) and Prdx2 in human systems (66), an efficiency of 107 M −1 s−1 for H2O2 is similar to catalase (107 M −1 s−1) or selenocysteine-based Gpx (108 M −1 s−1) (122).
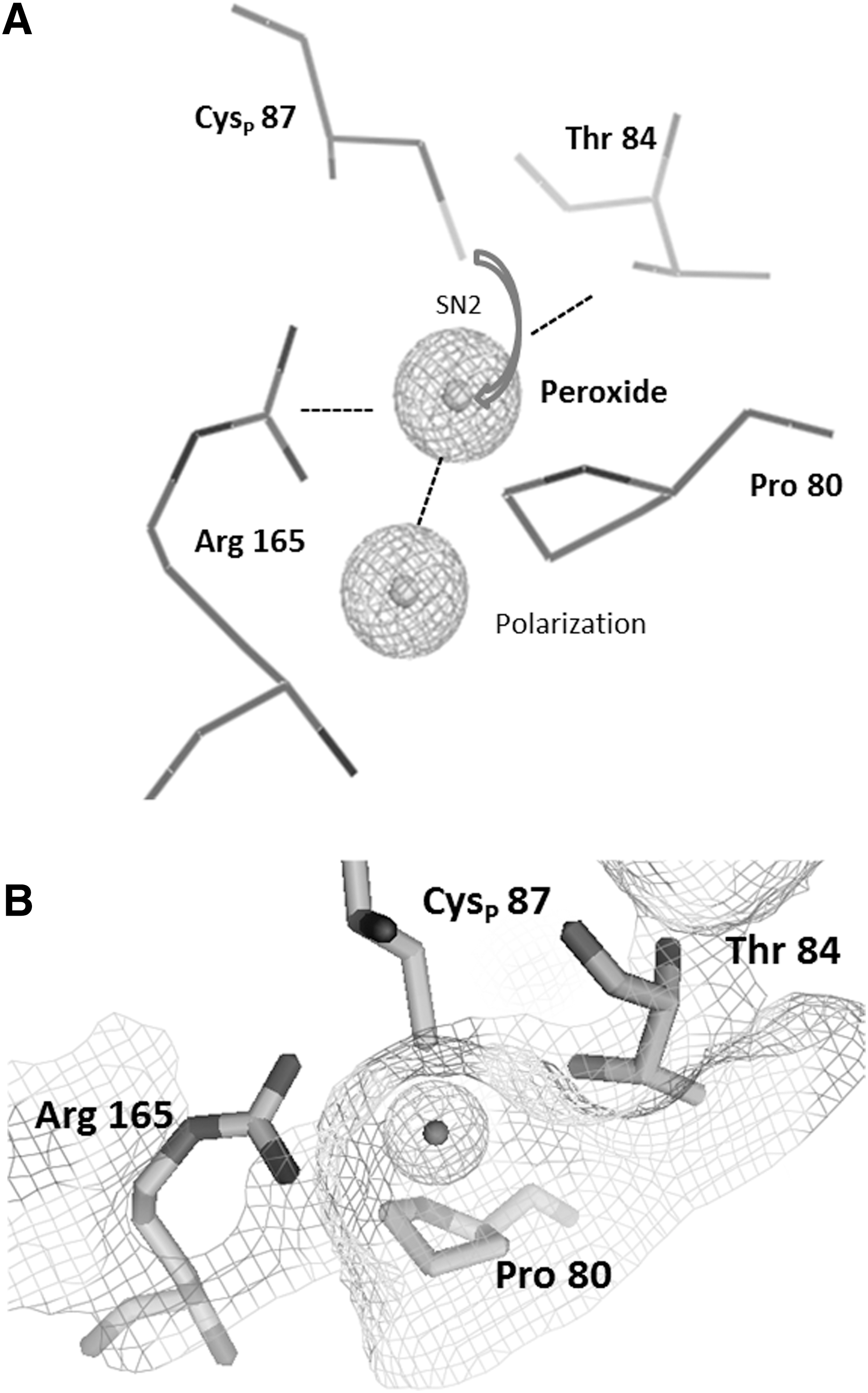
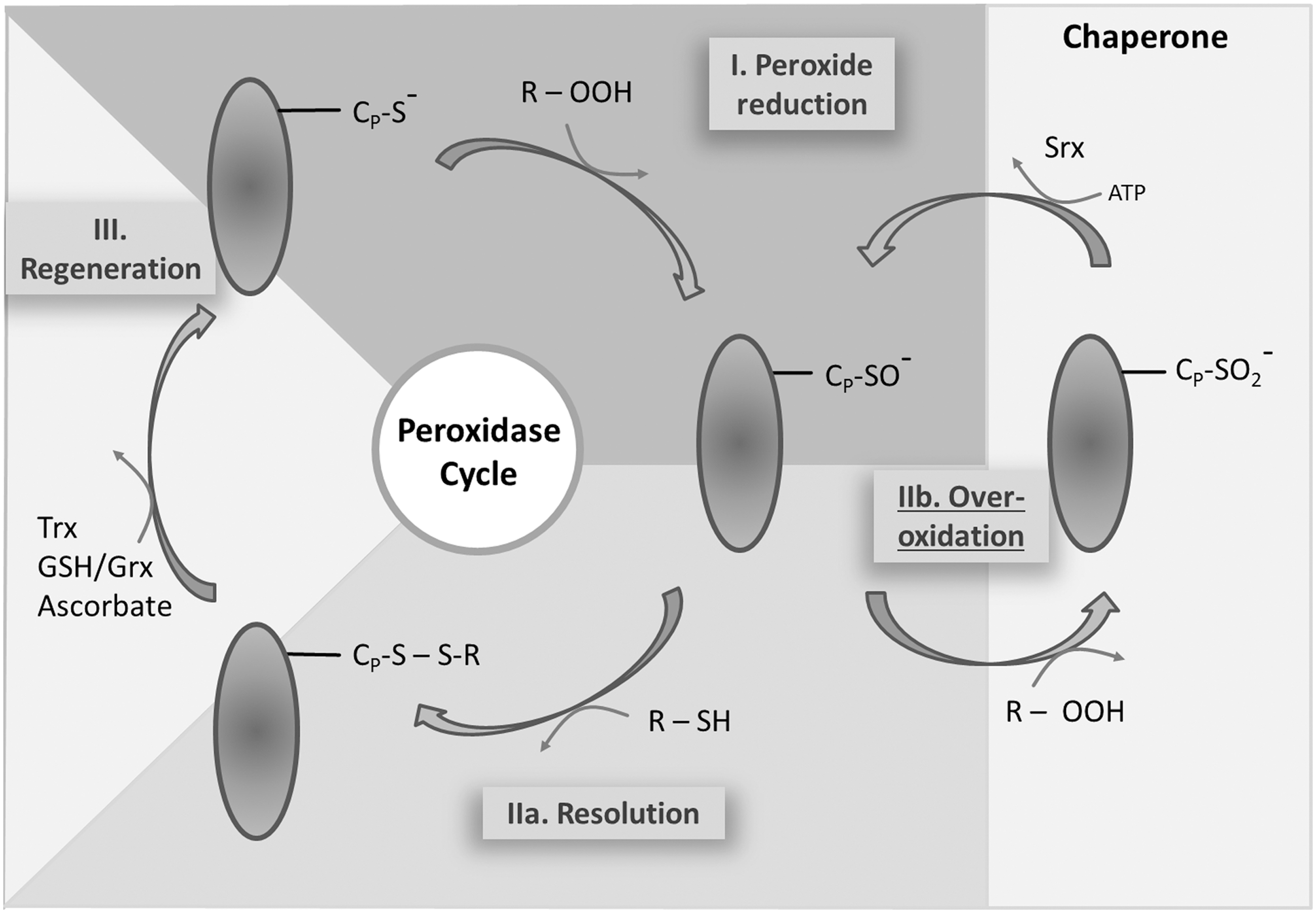
The thiol peroxidase cycle consists of three steps: the reduction of the peroxide substrate and generation of the sulfenic acid form of CysP, the resolution of the sulfenic acid to release water, and the regeneration of the thiol by electron transmitters (Fig. 2). (I) After binding of the peroxide to CysP, the oxygen–oxygen bond of the peroxide is disrupted and the corresponding alcohol from alkylhydroperoxide, water from H2O2, and nitrite from peroxinitrite are released. The active thiol group of the CysP is converted to a sulfenic acid (–SOH) group. (IIa) The CysR on the same or a different molecule attacks the sulfenic acid and forms an inter- or intramolecular disulfide bond (Cys–S–S–Cys). The disulfide bonded Prx adopts the inactive form. (IIb) Peroxide competes with the CysR for the sulfenic acid group. A second oxidation can take place before the resolving step occurs. This is considered as a regulatory mechanism that allows for inactivation and local peroxide accumulation in line with the flood gate model. The sulfinic acid is described as hyperoxidized form of Prxs. Its conversion to the active thiol peroxidase requires an additional regeneration step by sulfiredoxin (Srx) (94). (III) In the final step, the disulfide is reduced by specific redox transmitters such as Trxs, NTRC, Grx in combination with GSH, and ascorbate (reviewed in Ref. 31).
Prx Properties of Typical 1-CysPrx and 2-CysPrx
1-cysteine peroxiredoxin
Typical 1-cysteine peroxiredoxins (1-CysPrxs) belong to the B-type class and have a homodimer interface like A-type Prxs. 1-CysPrxs carry the CysP but lack the CysR. The sulfenic acid is resolved by other interaction partners. Information on plant 1-CysPrxs is still scarce. They are preferentially expressed in seeds wherein, for example, in barley they are found in the nucleus of aleurone and scutellum cells (113). Owing to its mostly confined expression in seeds, 1-CysPrx has mostly moved out of focus of current redox research. In 2009, Pulido and coworkers described the NTR/Trx system as electron donor for regeneration of 1-CysPrx (95). Enzyme assays with heterologously expressed wheat 1-CysPrx showed NTR-dependent peroxidase activity as well as localization in the nucleus as shown by immunolabeling. With 70 kDa, the molecular mass of NTR dimers is too big to diffuse through nuclear pores and thus the translocation mechanism to the nucleus remains elusive in the light of the missing nuclear localization sequence (95). It may be speculated that 1-CysPrx is taken piggyback by other cargo. Oxidized 1-CysPrx can be regenerated with ascorbate as electron donor. The efficiency appears to be low compared with that with the Trx system both for yeast mTpx and rat Prdx6. The antioxidant role of erythroascorbate remains a matter of debate despite the fact that the kcat/KM values for erythroascorbate with 104 M −1s−1 were one order of magnitude below that of Trx with 105 M −1s−1 (72). A recent report compared kinetic reactivity of the 1-CysPrx sulfenic acid with ascorbate across many species (3). The catalytic efficiencies kcat/KM for H2O2 decomposition were similar for yeast, bacteria, plant, and human 1-CysPrx with values of around 103 M −1s−1. The significance of the ascorbate-dependent regeneration in plants is still difficult to judge. Its contribution likely depends on the relative concentration of the various reductants, because in addition to Trx also GSH and glutathione-S-transferase and glutaredoxin, respectively (GST/GSH, Grx/GSH), regenerate 1-CysPrx at least in mammalia (64) and yeast (87). The link of 1-CysPrx to various regeneration systems may increase the viability of seeds facing harsh environmental conditions. Indeed Arabidopsis seeds overexpressing the PER1 homologue of Nelumbo nucifera (24) germinated better by 25%–50% than wild type (WT) in controlled deterioration treatments. The seedlings accumulated fivefold more biomass when grown on ABA for 10 days. These observations and increased germination at 50°C underpin the crucial role of 1-CysPrx in the redox network of seeds.
2-cysteine peroxiredoxin
Typical 2-CysPrxs interact through an obligate monomer–monomer interface orientated in parallel to the β-sheet bundles (B-type interface) in the Trx fold. In addition, intermolecular thiol–disulfide bonds are formed in the homodimer (46). 2-CysPrx is a highly abundant protein in the chloroplast stroma (100–120 μM in total for both 2-CysPrx A and B in A. thaliana) and likely involved in redox regulation in both the stroma and near the thylakoids (58, 89). Immunolabeling of 2-CysPrx with gold nanoparticles-conjugated antibodies revealed a location vicinal to thylakoid membranes of barley chloroplasts for 72% of all counted particles. Control analyses of etioplasts resulted in less (32%) label in proximity to the prolammelar body. Preimmune serum gave no such label in chloroplasts (58). Muthuramalingam et al. (77) detected 2-CysPrx in isolated photosystem II-enriched particles. Cerveau et al. (21) analyzed carefully washed membranes obtained from control or water-stressed A. thaliana WT and srx mutants and failed to detect 2-CysPrx in the thylakoid fraction. These controversial observations indicate that 2-CysPrx possibly associates with the thylakoids in a dynamic and conformational state-dependent manner and that this association is lost during extensive washing.
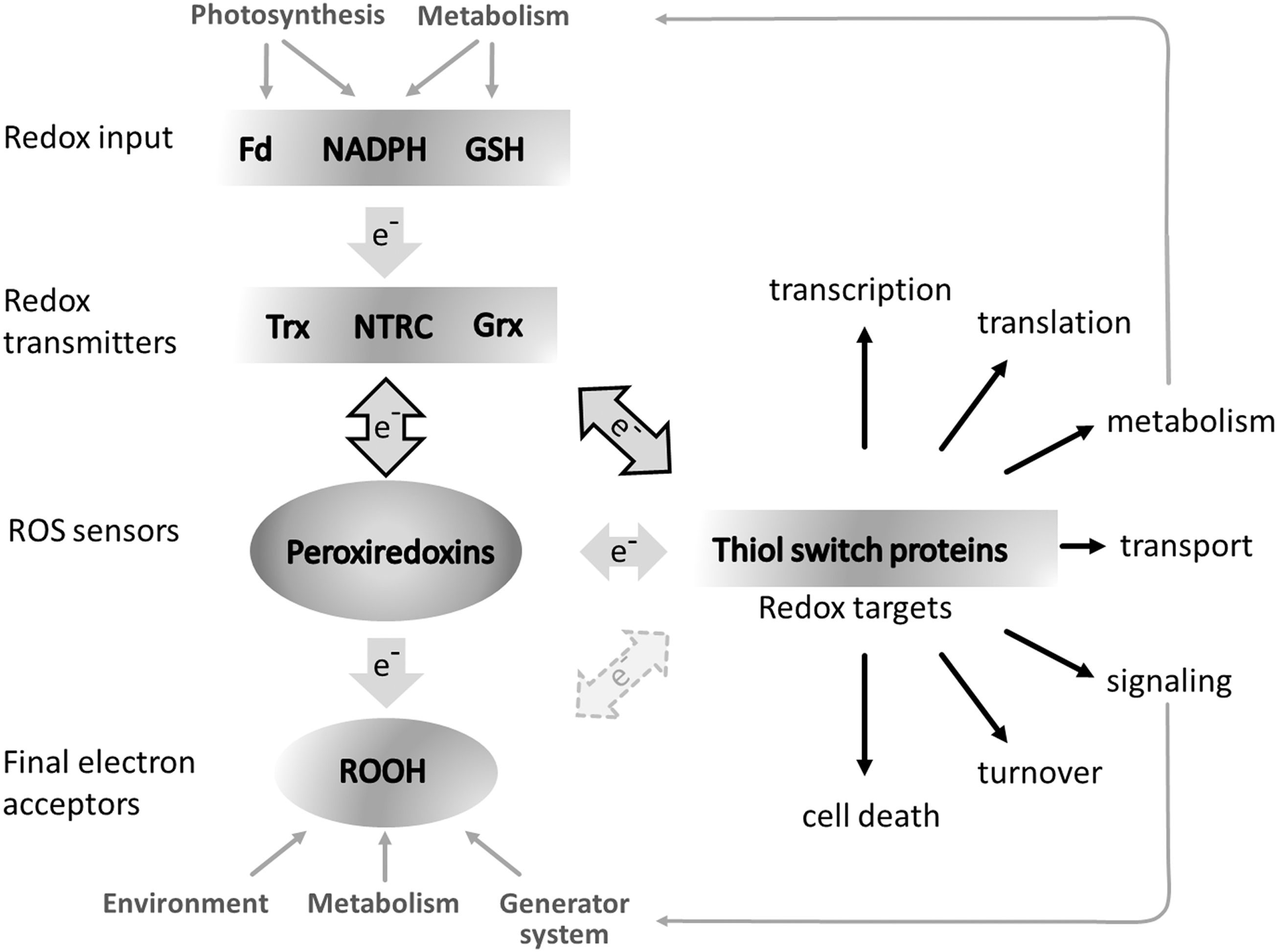
2-CysPrx adopts five different quaternary conformations. The reduced dimer, the reduced decamer, and the oxidized dimer take part in the peroxidase cycle already described. The hyperoxidized decamer and the hyperoxidized hyperaggregate realize the chaperone function of 2-CysPrx (60). Site-directed mutated variants of Arabidopsis 2-CysPrx revealed variable chaperone activity with lowest activity of a C-terminally truncated version and a disulfide-bond mimicking variant and highest activity for the hyperoxidation-state-mimicking variant C54D (59). 2-CysPrx, together with PrxIIE and PrxQ, is part of the complex thiol–disulfide network of the chloroplast, functions as redox sensors, and participates in regulation and maintenance of metabolic homeostasis (Fig. 3) (30). Owing to its high affinity and reactivity, 2-CysPrx may be considered as primary reactive oxygen species (ROS) sensor. In line with this hypothesis, 2-CysPrx was among the most sensitive targets of oxidation by H2O2 in vitro and methylviologen treatment in vivo (76). Regeneration is achieved by, for example, NTRC and Trx-x, but also by Trx-f1 and thereby is connected to the electron input elements ferredoxin (Fd) and NADPH (26, 90). The peroxidase activity is limited by the slow regeneration reaction. Increasing ROS turnover may disturb this process by hyperoxidizing the catalytic cysteines after ∼250 peroxidase cycles (56, 62). Then the alternative ATP-dependent regeneration system of Srx regenerates the active 2-CysPrx in a very slow reaction with about 1 min−1. Initially, Srx was thought to be specific for 2-CysPrx in the chloroplast until Iglesias-Baena et al. (53) proved the dual targeting of Srx also to the mitochondrium. The data revealed that Srx reduces hyperoxidized PrxIIF in the mitochondrion.
The occurrence and functional link between oligomerization, hyperoxidation, and chaperoning still need to be further investigated for structural and functional features. Noichri and coworkers (82) suggested that stacked reduced decamers act as precursors for stacked hyperoxidized aggregates in Saccharomyces cerevisiae. Two different high molecular weight (HMW) fractions were identified by SEC and native polyacrylaminde gel electrophoresis (PAGE) or SDS-PAGE, depending on the redox state. The occurrence of hyperoxidized 2-CysPrx follows a circadian rhythm in eukaryotic cells. The circadian hyperoxidation does not depend on transcriptional circuits and correlates with release of H2O2 from mitochondria in yeast (101). It should be noted that the fraction of hyperoxidation is rather small with an estimated share of up to 1.6% of total 2-CysPrx in erythrocytes of WT mice (25). The circadian oscillation of 2-CysPrx hyperoxidation also occurs in A. thaliana and other plants (34). But again the amount subjected to hyperoxidation is low (21). For the time being, the circadian appearance of hyperoxidized 2-CysPrx is considered as indicator for daily alterations in cell redox milieu possibly linked to specific metabolic activities such as FeS center assembly. A major accumulation of hyperoxidized 2-CysPrx was also not seen under temperature or water stress (21). The absence of HMW complexes was interpreted as indicator that the chaperone function might be less important than previously proposed. In line with this suggestion, hyperoxidized 2-CysPrx could neither be detected in nonreducing gels nor in Western blots using sulfinic/sulfonic antibody. Srx and NTRC transcript levels coding for the main regulators of 2-CysPrx redox state were unaffected under stress (94).
Apparently the system consisting of 2-CysPrxs, NTRC, and Srx guarantees a rather stable redox state of 2-CysPrx under most conditions. Interestingly, hyperoxidized 2-CysPrx was found in plants lacking NTRC. 2-CysPrx can turn hyperoxidized only if the peroxidase cycle is running. Therefore, the hyperoxidation in ntrc background proves that 2-CysPrx receives sufficient electrons by NTRC-independent components such as Trx-x, CDSP32, Trx-f, and Trx-m to sustain the peroxidase cycle. While questioning the significance of circadian oscillations in hyperoxidation, the authors suggest a light-dependent regulation of 2-CysPrx hyperoxidation dependent on photosynthetic electron transport (PET) and the Fd/FTR/Trx system (21). Alternatively, the small fraction of hyperoxidized 2-CysPrx might realize a gain of function by specific interaction with targets apart from general chaperoning.
A peculiar feature of reduced 2-CysPrx is its dynamic transition from dimer to decamer. Several physicochemical parameters influence this transition and the stability of the reduced decamer, in particular pH, ionic strength, redox potential, and its own concentration (12, 62, 75) (Fig. 4A). Below the critical transition concentration (CTC), reduced 2-CysPrx exclusively adopts the dimeric conformation. With extremely high cooperativity, reduced dimers assemble to decamers above the CTC similar to the abrupt transition of free lipids to lipid micelles above the critical micelle concentration. SEC, isothermal titration microcalorimetry (ITC), and dynamic light scattering defined the CTC with about 2.2 μM at pH 8 and 4.2 μM at pH 7.2. The pH dependency might possibly be linked to a histidine in the dimer–dimer interface, which is protonated at lower pH. His protonation may decrease the ability to form decamers.
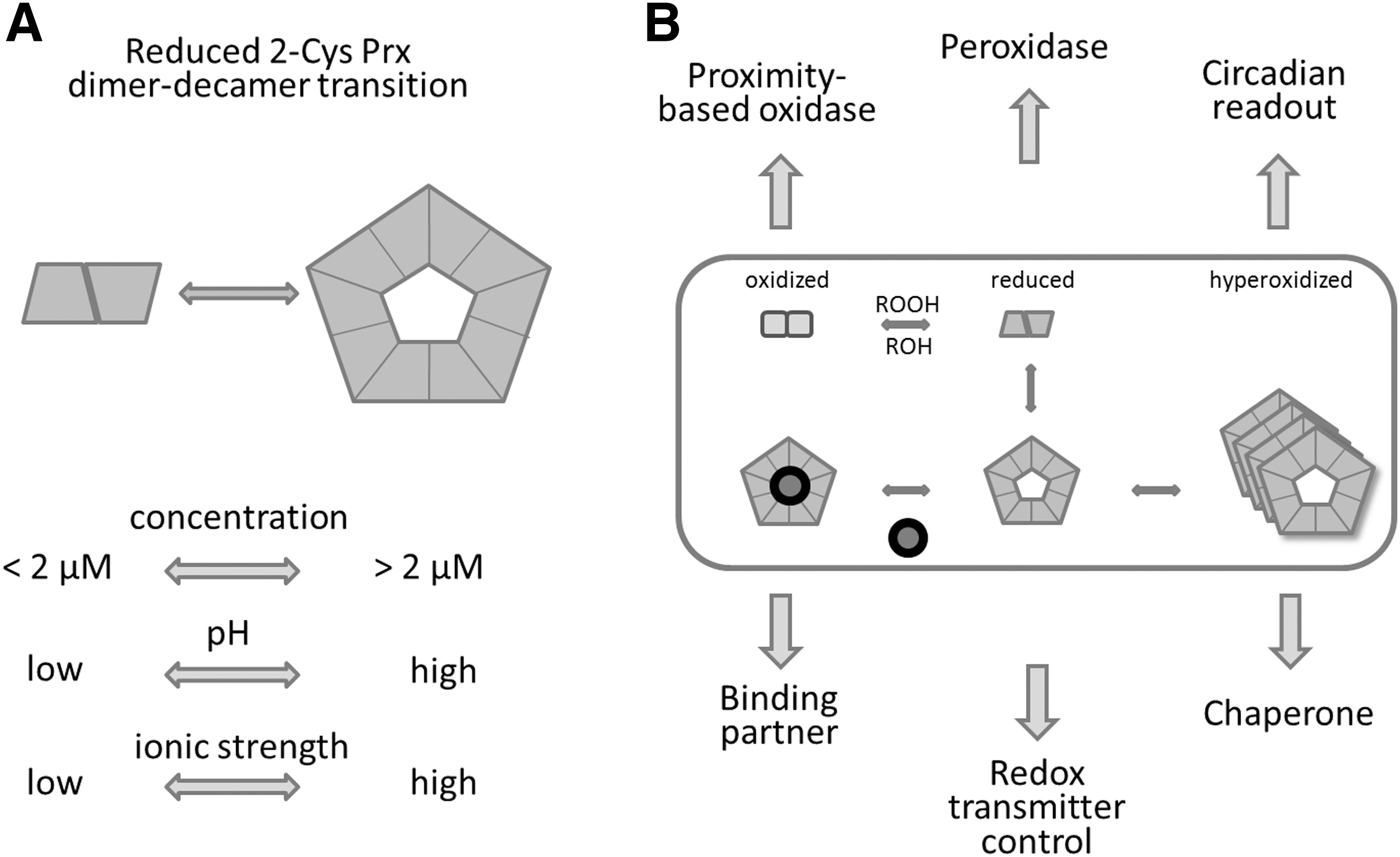
Stromal proteins interfere with the CTC, for example, the cyclophilin 20-3 (Cyp20-3), a peptidyl-prolyl-cis/trans isomerase that regulates cysteine synthesis (85). In contrast, titration of 2-CysPrx in 200 μg/ml stromal protein extract significantly decreased the CTC to 0.6 μM (62). Apparently complex and dynamic mechanisms regulate the dimer–decamer equilibrium, wherein a relatively small portion of dimers interact with several partners, or the binding partners of the decamer affect the equilibrium between dimer and decamer. The schematic depicted in Figure 4B summarizes the conformational states and their proposed or proven functions.
The Prx Interactome
2-CysPrxs, such as PrxIIE and PrxQ, are part of the chloroplast redox-regulatory network and are targeted by ROS as oxidants and electron transmitters as reductants (Fig. 3). Chloroplast Prxs and Gpx are preferentially reduced by some members of the whole chloroplast Trx family, consisting of Trx-f1, -f2, m1-4, -x, -y1, -y2,-z, 9 Trx-like proteins (NTRC, CDSP32, ACHT 1–4 [atypical cysteine histidine-rich Trx], and APR-1–3 [protein disulfide isomerase-like proteins]). The thiol peroxidases and two Asc-dependent peroxidases (thylakoid Apx and stromal Apx) reveal distinct suborganellar localization in the plastids (14, 61, 71).
The stromal 2-CysPrx is part of an intricate protein–protein interaction network. NTRC and Trx-x, together with CDSP32, are described as the predominant reductants of oxidized 2-CysPrx in vitro and in vivo and regenerate the active thiol peroxidase (Fig. 2) (14, 17). Reduced Cyp20-3 interacts with reduced 2-CysPrx as shown by fluorescence resonance energy transfer in protoplasts, ITC, and in vitro binding assays (62, 77).
Cyp20-3 binds 12-oxophytodienoic acid (OPDA), the precursor of jasmonates, with high affinity (Kd = 196 nM) (85). OPDA-liganded Cyp20-3 activates the chloroplast cysteine synthase complex consisting of serine acetyltransferase 2.1 (Serat) and O-acetylserine thiol lyase-B (OASTL-B). However, Cyp20-3 may not be necessary for chloroplast cysteine synthesis because formation of the cysteine synthase complexes occurs spontaneously (ΔG = −33 kcal mol−1) (112). In addition, only about 20% of cellular GSH synthesis derives from chloroplast Serat 2.1/OASTL-B, whereas 80% originate from mitochondrial and cytosolic cysteine synthase (112). Nonetheless, Cyp20-3 is regarded as a mediator of stress-dependent redox responses. The interaction between 2-CysPrx and Cyp20-3 occurs under reducing conditions and is suggested to compete with the activation of the cysteine synthase depending on the binding of OPDA and a potential conformational shift. This mechanism would allow for a negative regulatory feedback, slowing down stress-induced cysteine synthesis after re-establishing redox homeostasis.
Dangoor et al. (27) described an interaction of 2-CysPrx with ACHT1 as part of the chloroplast redox-regulatory network. 2-CysPrx is regarded as oxidant of ACHT1, which is suggested to regulate metabolism and acclimation processes, for example, during light–dark transitions of photosynthesis. Furthermore, ACHT4 also interacts with 2-CysPrx using a similar mechanism like ACHT1. The ADP-glucose pyrophosphorylase is an essential enzyme of starch synthesis. Its small subunit is implicated in the redox network as downstream target of ACHT4 (35). During dark–light transitions, the fast reoxidation of ACHT4 depends on 2-CysPrx. 2-CysPrx likely functions as oxidant to switch off starch synthesis in the dark similar to the proposed hypothesis that 2-CysPrx acts as a local redox sink transferring electrons to ROS to shut down photosynthesis. There is a need to kinetically resolve the redox state of components of the thiol–disulfide redox regulatory network in WT and mutants to better define the role of 2-CysPrx in rapid redox regulation (45).
Recently, an ex vivo approach based on coimmunoprecipitation (CoIP) and mass spectrometry allowed for identifying a set of 2-CysPrx interactors (20). The strength of this study rested on the comparison of CoIP proteomes of WT and 2-CysPrxA/B-deficient A. thaliana. Polypeptides recovered with anti-2CysPrx antibody from 2cysprxA/B-CoIPs were considered as false positives and subtracted from the polypeptides detected in WT CoIPs. This approach identified 67 interacting polypeptides from the chloroplast. The 91 interactions with polypeptides from extraplastidic compartments may originate from lost compartmentation during extraction. Fructose-1,6-bisphosphatase, ACHT2, and CDSP32 were among the known interactors. Surprisingly, Trx, NTRC, and Cyp20-3 were not found possibly because of the weak nature of interactions.
Interestingly, binding to other cyclophilins such as Cyp37/38 and CLP protease supports a role of 2-CysPrx protein in protein folding or regulation. The interactions with, for example, protochlorophyllide reductase B, 5-adenylylsulfate reductase (APR), GSH reductase, and three ferredoxin (Fd)-dependent proteins, namely Fd-sulfite reductase, Fd-nitrite reductase, and Fd-glutamate synthase reinforce the suggested function of 2-CysPrx in the redox-regulatory network of the chloroplast (20). Future work will have to address the significance of the interactions for interactor function and chloroplast metabolism.
A prerequisite for protein interactions are partner-specific surface-exposed residues, which form the interface either permanently or become surface exposed after conformational changes induced by interaction or variation of the milieu. A 2-CysPrx site with binding characteristics is the flexible C-terminus that is involved in binding the Ser protease Omi/HtrA2 and embraces Srx1 during the retroreduction of the hyperoxidized sulfinylated 2-CysPrx (50, 56). As shown for the interaction of At2-CysPrx with NTRC, the deletion of the C-terminus decreases the affinity for NTRC and alters the peroxidase and chaperone activities (59). The best studied and closest sequence homologues of At2-CysPrx likely are human Prdx1 and Prdx2. Analysis of their interactomes allowed the identification of the sequences CXXC, PXXP, and LXXL as motifs found in either Prdx1- or Prdx2-binding partners (15).
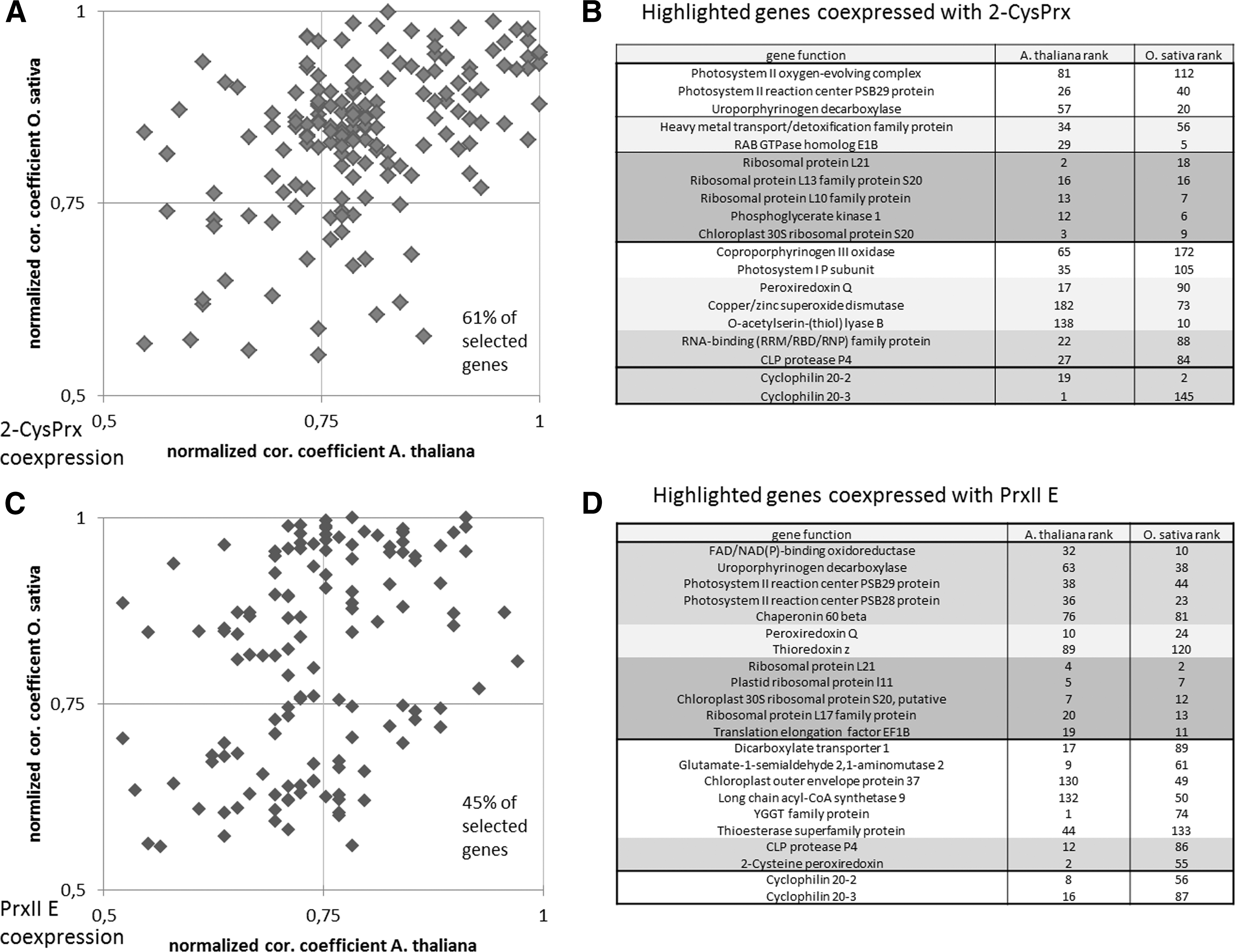
Coexpression frequently allows pinpointing to functional relationships. Transcriptional coexpression with Prxs has been reported by Muthuramalingam et al. (77) and Dietz (31) and identified Cyp20-3 as best hit for coexpression with values of 0.97–0.99. Here we extended this analysis to include the transcriptome of Oryza sativa cv. japonica (Rice Oligonucleotide Array Database [
Post-Translational Effects
The cysteine oxidation is the prime PTM on Prxs. The robust Trx-fold, conserved motifs, and flexible structures mediate conformational changes along with redox-dependent thiol modification. Apart from their thiol peroxidase function, Prxs can be considered as a prototypic thiol switch protein that is based on present-day knowledge evolved early during evolution. The dithiol–disulfide transition modifies conformation and function. Nowadays, the thiol switch mechanism is recognized for its broad significance in plants and other organisms with oxygenic metabolism (110). Such thiol switch proteins play fundamental roles in regulation and signaling of eukaryotes and prokaryotes, including mitochondria (4, 5, 40, 44, 79). ROS are generated at low rates by metabolism and, for example, by NADPH oxidases under normal conditions. They are detoxified by antioxidant enzymes, including thiol peroxidases (32). Thiol peroxidases are regenerated by redox transmitters, which then may oxidize other thiol switch proteins. Accumulating ROS under stress in addition may target sensitive thiols of other proteins (76).
Reversibility of disulfide formation, sulfenylation, and possibly sulfinylation contrasts sulfonylation and oxidative modifications of other amino acid residues, which often mark proteins for degradation (10, 28). Thus it was a very important finding that the hyperoxidized sulfinic acid forms of 2-CysPrx and PrxIIF can be regenerated by Srx (16, 53, 118). The specificity or promiscuity of Srx in reduction of sulfinylated proteins still needs to be scrutinized because hyperoxidation of 1-CysPrx may be irreversible (118). These authors state that hyperoxidized 1-CysPrx but also glyceraldehyde-3-phosphate dehydrogenase might be degraded because neither of them is a target of Srx.
Redox-dependent inhibition of Prx peroxidase activity triggers alternative functions and signaling pathways. Hyperoxidized 2-CysPrx prevents heat-induced denaturation of citrate synthase in vitro (54, 59). Overexpression of 2-CysPrx suppresses aggregation of cyclophilin 20-3 coexpressed in protoplasts in vivo (77). Combining redox-dependent peroxidase and chaperone function in a highly abundant protein provides the opportunity to operate a molecular redox switch in response to oxidative stress. The functional switch from peroxidase to chaperone is part of the floodgate model. Hyperoxidation of Prxs might shut down the detoxification in the thiol peroxidase branch and temporally and spatially enhance the ROS-dependent signaling (121). Analysis of 2-cysprx A/B knockout mutants revealed enhanced photo-oxidative damage and higher H2O2 levels in high light. Considering its high abundance in chloroplasts and its association with the thylakoids, the 2-CysPrx might act as a local redox-controlled switch to allow for the release of ROS signals from the photosynthetic electron transport chain. In turn, ROS release may change chloroplast processes but also initiate retrograde control of extrachloroplast processes. However, regulation of ROS-responsive marker genes such as ZAT12, BAP1, HSFA2, or OXI1 was unaltered in 2cysprxA/B compared with WT and only responded after introducing the additional knockout in the thylakoid ascorbate peroxidase (tApx) gene. This might indicate a synergistic floodgate control by thiol and ascorbate peroxidases and mutual compensation of the lost other enzyme (9).
PTMs of Prxs by glutathionylation, nitrosylation, acetylation, or phosphorylation partly mimic cysteine hyperoxidation and thus oligomerization, and realize oxidation-independent regulation and stress responses. The tripeptide GSH is present at millimolar concentrations in plasmatic compartments and functions as redox–thiol buffer, electron donor, substrate for phytochelatin synthesis, and in S-glutathionylation (80). S-glutathionylation is necessary for redox regeneration of 1-CysPrx and type II Prxs, which then depends on GST or Grx-coupled cysteine reduction.
Under conditions of GSSG accumulation, the mass of PsTrx-h2 increases to 14,361.1 Da corresponding to addition of 305.5 Da of GSH to 14,056.9 Da (42). Another example of reversible glutathionylation occurs with chloroplast glyceraldehyde-3-phosphate dehydrogenase and NADP-dependent malate dehydrogenase (13, 69). The GSH/GSSG pool participates in redox homeostasis and controls various signaling pathways. Reversible S-glutathionylation introduces a negative charge and a bulky tripeptide and protects thiols from hyperoxidation.
Peroxynitrite (ONOO−) serves as substrate for 2-CysPrx and PrxIIE (8, 107). Biotic and abiotic stress conditions provoke nitrosoglutathione (GSNO) accumulation, which is a storage pool for NO. NO is a radical and participates in cell signal transduction (38). A proteome analysis identified >1000 S-nitrosylated targets in A. thaliana and likewise in Kalanchoe pinnata, Brassica juncea, and Citrus aurantium. These proteins are involved in major cellular activities such as primary and secondary metabolism, photosynthesis, genetic information processing, cellular architecture, and response to biotic and abiotic stresses (8). As part of detoxification networks, PrxIIE and PrxIIB (cytosol), PrxIIF (mitochondria), and GR and Gpx are targets of S-nitrosylation. These and other nitrosylated polypeptides were inventoried by MS/MS analysis after addition of GSNO to Arabidopsis cell cultures or exposing leaves to 1 ppm NO via the gaseous phase (63). Proof of physiological significance is scarce. S-nitrosylation of PrxIIE is suggested to ease ONOO−-dependent tyrosine nitration as signaling event in pathogen defense (104). Plant Prxs are Lys acetylated as reported for Os1-CysPrx (47) and for Os2-CysPrx (124). Acetylation sites are KS
More is known about the physiological role of plant protein phosphorylation (98, 102, 115, 126). A recent analysis of the phosphoproteome of grapevine (67) confirmed phosphorylation of Vv-Apx (Vitis vinifera) in addition to the cytoplasmic VvPrxIIB in response to phytoplasma infection. A similar pattern was found in A. thaliana after wounding and in Medicago truncatula after rhizobial infection (52, 105). To address the consequences of kinase inhibition, Xu et al. (125) observed reduction in the drought-induced H2O2 production and a decrease in the drought-enhanced activities of antioxidant enzymes such as SOD, Apx, and Gpx in maize. Queries to the online databases IPTM net (
Phosphorylation sites (p-site) with phosphorylated serine, threonine, and tyrosine (small letters s, t, y in bold) of peroxiredoxins, thioredoxins, ascorbate peroxidases, glutathione peroxidases, CDSP32, and ACHT1 revealed by employing databases mentioned on the next page (48, 127). The sequence containing the modified aminoacid (aa) was arbitrarily set 5–6 aa long. Aminoacid positions refer to primary sequence of the proteins as counted from N to C terminus with transit peptide included. Following proteins were queried: superoxide dismutase, PrxIIE, PrxIIF, PrxQ, 2-CysPrx A, 2-CysPrx B, Apx, ascorbate peroxidase, tApx, thylakoid ascorbate peroxidase, sApx, stromal ascorbate peroxidase; chloroplast Gpx, chloroplast; Trx members (Trx-f1, -f2, m1-4, -x, -y1, -y2,-z), Trx-like proteins (NTRC, CDSP32, ACHT 1–4 [atypical cysteine histidine-rich Trx], and APR 1–3 [protein disulfide isomerase-like proteins]), Fd, ferredoxin 1; FNR, ferredoxin-dependent NADPH reductase 1.
P-sites were found in the cited references.
Interestingly, At2-CysPrx and the mammalian 2-CysPrx type Prdx1 have the C-terminal SKEYF site in common, which is phosphorylated at the Ser in A. thaliana and at the Tyr in mouse, rat, and human (119). In vitro phosphorylation of Tyr in the respective site of human Prdx1 decreases the peroxidase activity by 80% (119). According to their proposed model, a local rise in peroxide concentration leads to MAPK activation, and phosphorylation of peroxiredoxin, in turn, allows for further signaling. Whether this model is applicable to plant 2-CysPrxs and other redox proteins is not known. Altogether, these results indicate a relationship between redox regulation, PTMs, and signaling. The PTM of redox proteins and their implications in planta still needs to be explored.
Membrane Interaction
The C-terminus of 2-CysPrx bears the motif SKEYF, which is conserved among plant 2-CysPrx but absent in its prokaryotic homologue, the alkyl hydroperoxide reductase C (AhpC). In addition to the feature of SKEYF phosphorylation mentioned in the previous section, the SKEYF motif is also present in the C-terminal sequence 186NVDDSKEYFSKHN198 of the human 2-Cys Prx, Prdx2, where it mediates its membrane attachment (22). Therefore, as shown for barley 2-CysPrx, the C-terminus that is essential for binding to NTRC (59) likely plays a role in 2-CysPrx membrane association, in regulation of its peroxidase activity (see previous section), or both. The SKEYF-containing sequence 184EKSMKPDPKLSKEYFSAI201 is computationally predicted to interact with membranes (Table 3). A different approach to investigate the possibility of peripheral membrane association of 2-CysPrx considers electrostatic interactions between the decamer and the membrane (68). Here it is striking that the outer pentagon sites of 2-CysPrx bear Lys-rich C-termini and the inner doughnut sites are enriched in positively charged residues. Yet, it is unknown whether these features play a role in realizing the putative amphitropic character of 2-CysPrx, wherein barley 2-CysPrx is found both in the soluble plastid proteome and associated with the thylakoid (58). Amphitropic proteins reversibly interact with the membrane by lipid-binding regions containing positively charged and hydrophobic amino acids residing in coil-helical regions (18, 55). Hydrophobic faces are also often involved in membrane interactions (41) and the analysis shown in Table 3 revealed a hydrophobic face sequence 162LQALQYIQENPDEVCPAG179. The hydrophobic face has negatively charged residues opposed to their hydrophobic domains, which could play a role in At-2CysPrx membrane association as proposed for the human Prdx2 membrane interaction model (92). Protons or Ca2+ ions may neutralize the negatively charged hydrophobic faces to imbed 2-CysPrx in the membrane.
The analysis with the HELIQUEST program used the default settings. Regions of At2-CysPrx were selected with minimum 50% α-helical character, but no β-strand content and no dimer–dimer interaction site (120). Secondary structure features of At2-CysPrx are based on the Phyre 2 (
The Aramemnon online tool (

The Ca2+-associated protein calpromotin nowadays called Prdx2 in mammals interacts with integral membrane proteins such as the gardos channel (Uniprot entry O15554), stomatin (Uniprot entry P27105), and band 3 (Uniprot entry P02730) (68, 73, 74). In contrast to Cha et al. (22), Matte et al. (68) proposed that the N-terminus of cytosolic band 3 (Cdb3) is crucial for Prx2 membrane association, whereas Plishker et al. (92) assume that the redox milieu and Ca2+ are essential for the switch from the soluble to the membrane-bound state. Alternatively, binding proteins may mediate thylakoid attachment of 2-CysPrx. Muthuramalingam et al. (77) detected 2-CysPrx in photosystem II-enriched fractions and were able to coimmunoprecipitate D1 protein and 2-CysPrx. Prdx2 is highly similar to At2-CysPrx. Therefore, one may ask whether the binding epitopes of the mentioned proteins can be found in At2-CysPrx. A protein Blast (
Final Remarks
Research on plant Prxs began with the identification of 2-CysPrx in the chloroplast and 1-CysPrx in the nucleus 20 years ago (11, 113). Since then, the biochemical features, cell biological mechanisms, and physiological implications of different plant Prxs have been elucidated from various perspectives, but the most significant advancement has been achieved for the chloroplast 2-CysPrx either by targeted investigation or by coincidental findings, for example, by identifying NTRC as most efficient but not exclusive electron donor of 2-CysPrx. Since the last comprehensive review (31), significant progress has been achieved in identifying the interactome as selectively displayed in Figure 7 for 2-CysPrx (20). Key components of the major metabolic pathways that consume reductive power have been proven or are suggested to interact with 2-CysPrx. This concerns photosynthetic light reactions, carbon metabolism, nitrogen and sulfur metabolism, antioxidant defense, and secondary compound synthesis. In this context, it is intriguing that ferredoxin-dependent proteins constitute a major fraction of interacting partners. Important interactions with proteins or membranes need to be confirmed and explored with respect to binding selectivity for redox-dependent conformational states. The successful proteomics analysis indicates that there is a need to conduct metabolomics and transcriptomics analyses to reach the global assessment of the physiological significance of the different Prxs. This includes the interference with Ca2+ and hormone signaling. Most importantly we need complementation of the 2-CysPrx knockout line with variants that mimic specific states of 2-CysPrx, namely devoid of peroxidase activity, lacking the oligomerization ability, resembling the hyperoxidized decamer, or with truncated C-terminus. It will be interesting to find out which features of the knockout phenotype can be complemented by each of the variants and this approach should allow dissecting the roles as thiol peroxidase, oxidant, interaction partner, and chaperone.

Footnotes
Acknowledgment
The work in this article was supported by the Deutsche Forschungsgemeinschaft DI346/14 and SPP1710.