
Abstract
Significance:
Atherosclerosis is a major cause for the death of human beings, and it takes place in large- and middle-sized arteries. The pathogenesis of the disease has been widely investigated, and new findings on vascular stem/progenitor cells could have an impact on vascular regeneration.
Recent Advances:
Recent studies have shown that abundant stem/progenitor cells present in the vessel wall are mainly responsible for cell accumulation in the intima during vascular remodeling. It has been demonstrated that the mobilization and recruitment of tissue-resident stem/progenitor cells give rise to endothelial and smooth muscle cells (SMCs) that participate in vascular repair and remodeling such as neointimal hyperplasia and arteriosclerosis. Interestingly, cell lineage tracing studies indicate that a large proportion of SMCs in neointimal lesions is derived from adventitial stem/progenitor cells.
Critical Issues:
The influence of stem/progenitor cell behavior on the development of atherosclerosis is crucial. An understanding of the regulatory mechanisms that control stem/progenitor cell migration and differentiation is essential for stem/progenitor cell therapy for vascular diseases and regenerative medicine.
Future Directions:
Identification of the detailed process driving the migration and differentiation of vascular stem/progenitor cells during the development of atherosclerosis, discovery of the environmental cues, and signaling pathways that control cell fate within the vasculature will facilitate the development of new preventive and therapeutic strategies to combat atherosclerosis. Antioxid. Redox Signal. 29, 219–235.
Introduction
I
The earliest discovery of adult stem cells can be traced back to the 1950s when researchers identified hematopoietic stem cells (HSCs) and bone marrow stromal stem cells (136). The most mature clinical application of adult hematopoietic stem/progenitor cells is bone marrow transplantation, which has been used for treatment of leukemia for many decades. In the 1990s, a population of stem cells that was discovered in the adult brain had been traditionally regarded as non-renewable cells; actually, recent studies demonstrated that they are capable of differentiation into astrocytes, oligodendrocytes, and neurons (128).
The specific environment in the body that harbors the adult stem/progenitor cells is called the “stem/progenitor cell niche,” which exists in many kinds of tissue and organs, such as: bone marrow, peripheral blood, skin, heart, brain, and blood vessel wall (77, 122). Usually, adult stem/progenitor cells are maintained in the quiescent state in the niche for many years. Once these cells are stimulated by adjacent injured tissues, they will be activated to participate in a cascade of repair via their proliferation, migration, and differentiation. In terms of adult stem/progenitor cells in vasculature, Hu et al. (59) reported the presence of stem/progenitor cells in the adventitia of the vessel wall expressing stem/progenitor markers, for example, Sca-1, CD34, and c-kit (61). Subsequently, reports from a number of groups showed the existence of vascular stem/progenitor cells, which could participate in both endothelial repair (61, 156) and the pathogenesis of atherosclerosis (9, 42).
Atherosclerosis, as one of the leading causes of mortality and morbidity in the world, is a multifactorial vascular disease that is influenced by genetic, gender-specific, and environmental factors (7, 46). It is the most common kind of arteriosclerosis and usually gives rise to coronary artery disease, carotid artery disease, peripheral arterial diseases, and cerebrovascular diseases (e.g., acute coronary syndrome, myocardial infarction, or stroke). The main characteristics of atherosclerosis are lipid deposition and vessel stiffness, which result in thickening of the vessel wall and narrowing of the lumen.
The underlying pathophysiology of atherosclerosis is initiated by endothelial dysfunction, which is caused by physical or chemical offence such as hypertension, disturbed blood flow in the branch of vessel, reactive oxygen species (ROS), hyperlipidemia, and hyperglycemia (69). These factors activate endothelial cells (ECs) to provoke a cascade of inflammatory responses. Subsequently, chemokines that are either derived from platelets in the blood or released from oxidized low-density lipoprotein (ox-LDL)-activated ECs attract inflammatory cells such as monocytes and T lymphocytes to migrate into sub-endothelial sites (74, 109, 126). During this process, ox-LDL also triggers defective ECs to express adhesion molecules (e.g., VCAM-1) on their surface. Over time, a continuous accumulation of lipid and production of inflammatory molecules by infiltrated inflammatory cells causes the lesion to become unstable. Macrophages will then replace smooth muscle cells (SMCs) in the fibrous cap to create an inflamed cap, which is more susceptible to rupture.
During this process, the role of vascular stem/progenitor cells might be crucial, because they can migrate from either media or adventitia to the intima where they may differentiate into different cell types, further contributing to atherosclerotic development. In this article, we will focus on recent studies on vascular stem/progenitor cells in atherosclerosis, discuss the underlying mechanisms of cell migration and differentiation, which could be potential targets for therapy of atherosclerosis in the near future.
Adult Stem/Progenitor Cells Within the Vessel Wall
For a long time, adult blood vessels were believed to be quiescent, and not to contribute to new blood vessel formation. However, based on observations of vascularization in tumor tissue, this idea was changed in the 1970s when Folkman proposed that new vessels were formed via angiogenesis—a process in which new vessels grow from pre-existing vessels (45). The second tremendous discovery came in 1997 from Asahara et al., who found that CD34+ vascular endothelial growth factor receptor-2+ (VEGFR2) endothelial progenitor cells (EPCs) circulating in peripheral blood could participate in the formation of new vessels at the sites of active angiogenesis (4). Subsequent studies indicated that the bone marrow, as a niche for EPCs, is a source from which the cells are mobilized into the circulation and directed to the place of new vessel formation (6, 107). However, recent studies indicate that “EPCs” described in many published papers, especially early “EPCs,” are monocytes/macrophages (110, 150). Thus, to date, the nature and function of “EPCs” in the circulating blood remains controversial.
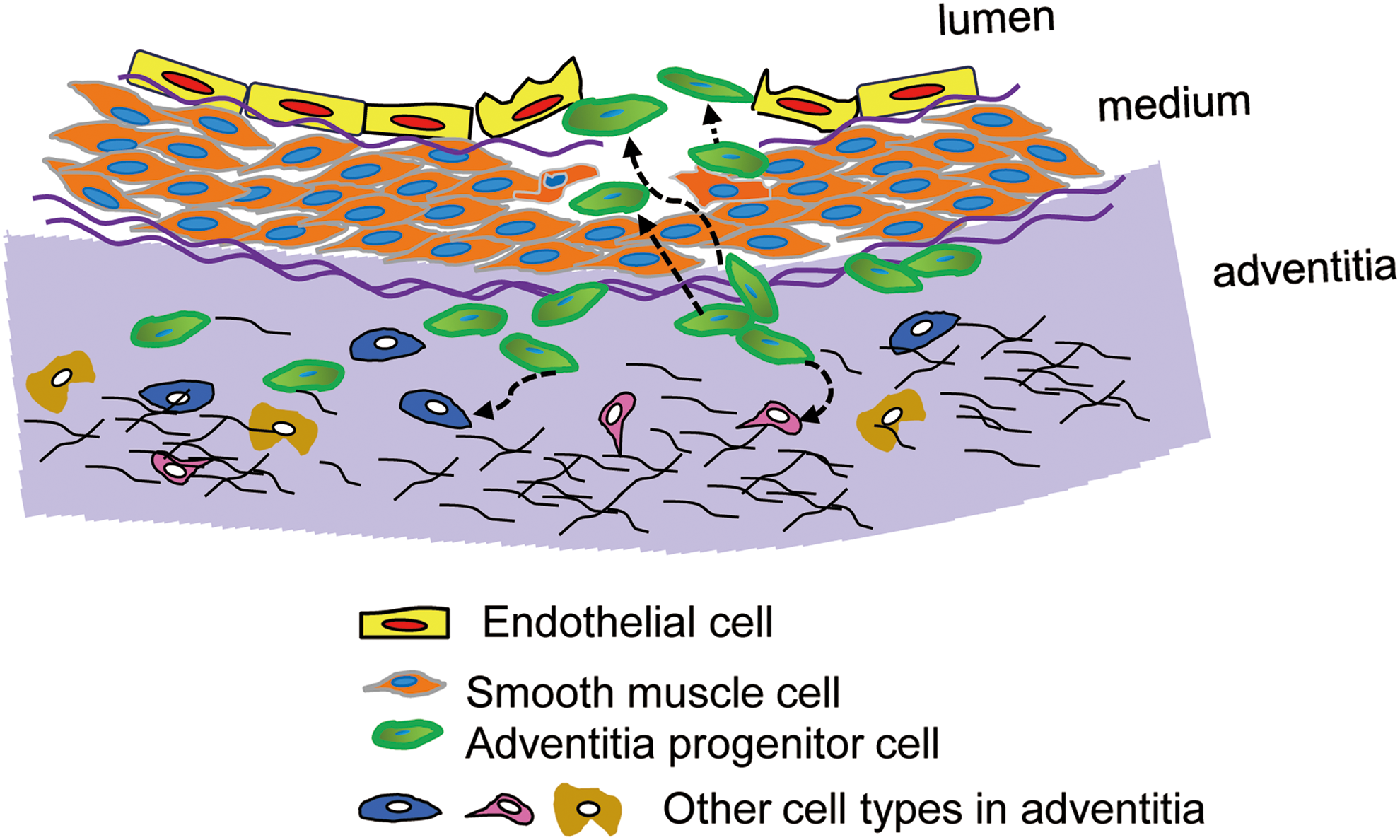
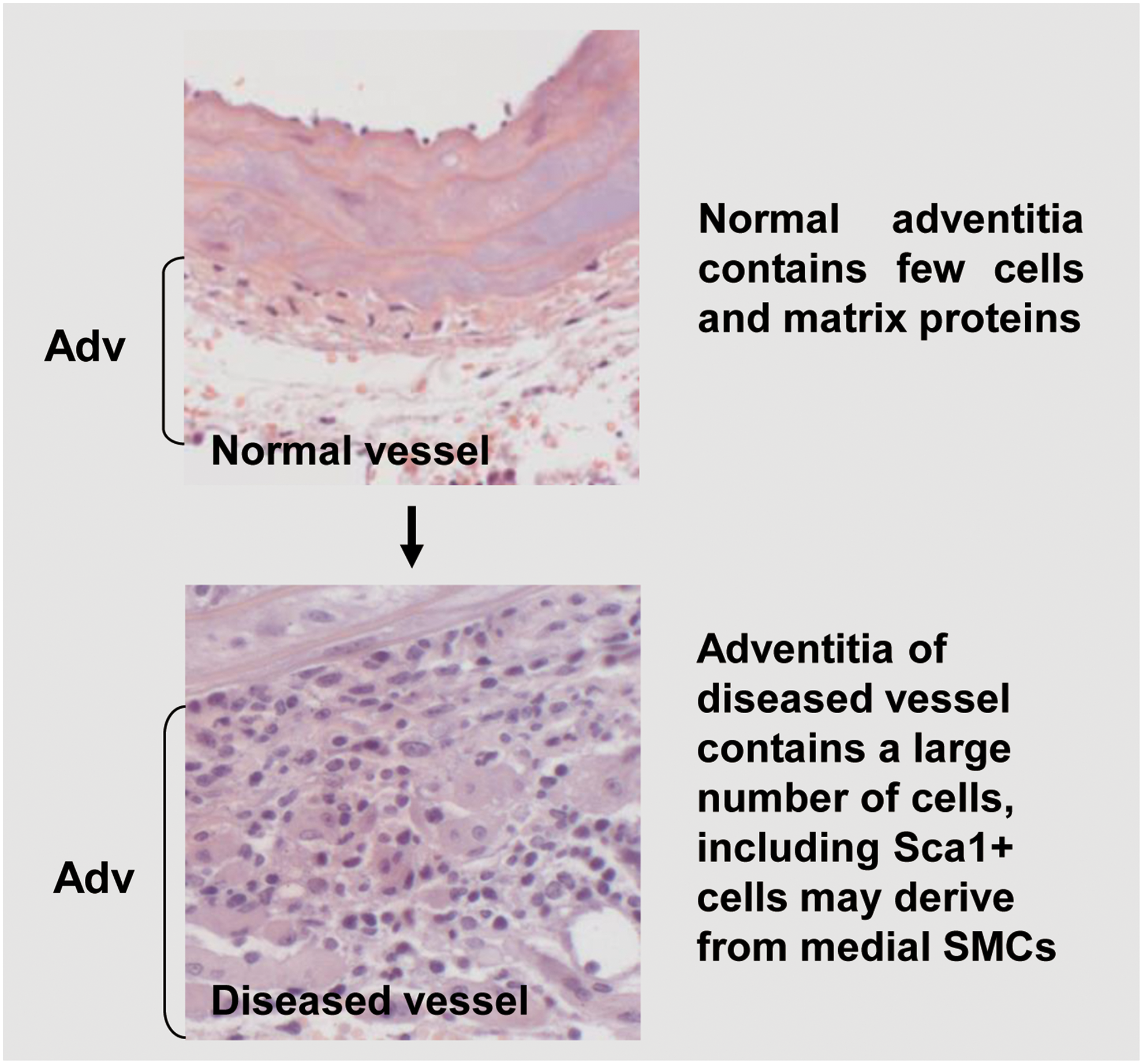
In the past decade, breakthrough experiments in the field of stem/progenitor cell research took place. Though there is still some debate, it is widely believed that stem/progenitor cells exist and reside within the vascular wall (37). Most experimental studies postulate that stem/progenitor cells reside in two main zones within the vessel wall, that is, the sub-endothelial zone and the adventitial zone (3, 40, 41, 67, 99, 102) (Table 1). The sub-endothelial zone in the micro-vessel wall corresponds to the adventitial zone in medium and large blood vessels. The adventitia is the outermost layer of the vessel wall, and it is composed of connective tissue, collagen fibers, vasa vasorum, fibroblasts, and quiescent inflammatory cells. Besides ECs, the adventitia cells also play an important role in maintaining vessel wall homeostasis, including retrieval, integration, storage, and release of regulators such as nitric oxide (NO) (53, 125). They participate in the growth and repair of the vessel and mediate cross-talk between different cell types residing in the vessel wall (53, 54, 119).
EC, endothelial cell; EPC, endothelial progenitor cell; MSC, mesenchymal stem cell; SMC, smooth muscle cell; VEGFR, vascular endothelial growth factor receptor.
ROS, as an important initiator of atherosclerosis, are usually spontaneously produced by nicotinamide adenine dinucleotide phosphate oxidase during the process of normal aerobic metabolism within healthy cells (36). However, during atherosclerosis, excess free ROS are dramatically released from either exogenetic pollutants in the circulation (e.g., smoking) (8) or activated endothelial and SMCs on the vessel wall (135). Besides their effects on oxidation of LDL (140) and reduction of NO bioavailability (15), ROS are also essential for mediating various inflammatory signaling pathways. ROS have been reported to mediate interleukin (IL)-4-induced CCL2 expression in human and mouse aortic ECs (73). Further, Nrf2 gene-transfected vascular SMCs inhibited ROS production, leading to a downregulation of CCL2 level as well as a decrease of vascular smooth muscle cell (VSMC) proliferation in rabbits (76).
In the adventitia of the aortic root, immunohistochemistry revealed the presence of progenitor cells that express a panel of progenitor cell markers such as Sca-1, c-kit, CD34, and FLK-1 (1). The adventitia niche, the region of adventitia close to the media, seems to fulfill the definition of such a niche environment and is believed to be a source of progenitors for vasculogenesis (156).
As shown in Figure 1, Sca-1+ adventitial progenitor cells (green) can differentiate into SMCs to replace lost or damaged SMCs in the media or into ECs and contribute to re-endothelialization in the intima. Further, these Sca-1+ progenitors can also respond to injury or disease of surrounding tissues and participate in the repair of damaged tissues (downward dashed arrows). (86). On the other hand, CD34+ cells are found in the vasculogenic zone, which potentially includes two subsets: CD34+VEGFR2+Tie+CD31− cells that differentiate into ECs and CD34−CD3−CD44+CD90+CD105+CD73 cells that differentiate into vascular SMCs (138). In addition, vessel wall-resident mesenchymal stem/progenitor cells have also been identified within the native adventitial niche and can similarly differentiate into SMCs. Further, adventitial Sca-1+ cells that reside within the vasculogenic zone from embryonic development to adulthood have the capacity to differentiate into SMCs in vitro under specific culture conditions (147). Finally, CD45+CD133+CD34− cells have been found to give rise to macrophages and other inflammatory cells (52).
Excessive ROS-Regulated Stem/Progenitor Cell Differentiation
Accumulating reports have demonstrated that adult stem/progenitor cells are mobilized by local signal molecules in their microenvironment; for example, in atherosclerosis, more ROS were produced by aging stromal cells, and these excessive ROS promoted HSC and EPC senescence, leading to premature exhaustion of them and impairment of their differentiation (60, 137, 143). Similarly, in transplanted mesenchymal stem cells (MSCs), either host tissues or MSCs themselves produced an excess amount of ROS that was believed to be a major factor causing MSC apoptosis by activation of MAPK signaling pathways after transplantation (113). By contrast, the enhancement of ROS level can promote embryonic stem cell (ESC) differentiation toward SMCs (149, 154). In terms of MSCs, ROS are playing different roles in MSC differentiation. For example, ROS inhibited osteogenic differentiation of murine or human MSCs (20, 90), whereas stimulation of murine and human preadipocytes with ROS promoted their adipogenic differentiation (124).
Vessel Wall MSCs
MSCs are heterogeneous multipotent stem cells that can differentiate into a variety of cells such as osteoblasts, adipocytes, chondrocytes, and myocytes and they play an important role in the development of organs and tissue derived from the mesenchyme (1). Besides the bone marrow, which is the best known storage site for MSCs, the vascular adventitia niche is also suggested to be the reservoir for MSCs in almost all organs (31). Traditionally, cells were identified as MSCs if they showed the ability to adhere to plastic in standard culture conditions and possessed a fibroblast-like morphology. Recently, immuno-selection using specific markers is more widely used to identify MSCs.
In 2006 the International Society for Cell Therapy published the minimal immunoexpression profile for defining MSCs, which is CD105+, CD73+ and CD90+, and CD45−, CD34−, CD14− or CD11b−, CD79α− or CD19− and HLA-DR− (35). However, several studies have found that CD34, which is supposed to be a negative marker of MSCs, can be expressed on uncultured bone marrow-derived MSCs and adventitial adipose-derived MSCs (80, 161). Some groups have also reported the presence of CD34+ MSCs in the vascular adventitia. In a study from Campagnolo et al. (16), CD34+CD31− cells isolated from the human saphenous vein were found to express pericyte/mesenchymal antigens together with stem cell marker Sox2 and to be endowed with clonogenic and multilineage differentiation capacities. In both in vitro and in vivo studies, they demonstrated that these cells interacted with the EC network and participated in angiogenesis (16). In another study, CD34+CD31− cells from three different human vessel walls, artery, vein, and small vessels in adipose tissue, were compared with adipose-derived stem/progenitor cells. Although they all displayed fibroblast-like morphologies, those cells exhibited different differentiation potentials, with arterial stromal cells undergoing greater osteogenic processes whereas venous and small vessel stromal cells promoted angiogenesis (152). Further, Klein et al. recently demonstrated that a population of CD44+ vascular wall multipotent stem/progenitor cells residing in the adventitial vasculogenic zone of human internal thoracic arteries are capable of differentiating into pericytes and SMCs (68) (Table 2).
ART, arterial wall; ASC, adipose-derived stem/stromal cell.
MSCs expressing a variety of surface markers have been reported by many different groups in the adventitia, so it is unclear as to how many unique MSC populations exist and how many overlap with cell populations previously defined by other groups. Through lineage tracing of Gli1+ perivascular cells, Kramann et al. recently showed that in response to injury, adventitial cells migrate, passing through the tunica media to form neointima; subsequently acquire an SMC-like profile; and can differentiate into osteoblasts (71). Thus, vascular MSCs could be another source of SMCs forming neointimal lesions.
Sca-1+ Stem/Progenitor Cells
Seminal studies by Hu et al. (59) identified the existence of a population of cells that are Sca-1+, CD34+, Flk-1+, and c-kit+ in the adventitia of aortic roots of ApoE−/− mice. Stimulated by platelet-derived growth factor (PDGF)-BB in vitro, these Sca-1+ cells can differentiate into SMCs. When Sca-1+β-gal− cells carrying the SMC-LacZ gene were seeded on the adventitial side of the vein graft in ApoE−/− mice, after 2–4 weeks, β-gal+ cells could be found in the neointima, which indicates that Sca-1+ cells can migrate from the adventitia to the neointima and differentiate into SMCs in vivo (59).
Subsequently, Passman et al. (100) reported a sonic hedgehog (Shh) signaling domain restricted to the adventitia of the artery wall, in close proximity to Sca-1+ stem/progenitor cells. In Shh−/− mice, the number of Sca-1+ cells in the aortic root is dramatically reduced. In accordance to the data shown by Hu et al. (59), the Sca-1+ cells that they characterized are also CD34+ and CD140b+ but c-kit−. They also found that in vivo, these Sca-1+ cells do not express SMC markers, but express transcription factors (e.g., serum response factor [SRF], myocardin family member) that are related to SMC differentiation. These Sca-1+ cells also express SRF-dependent transcription repressors to maintain their progenitor properties in vivo. These data suggest that Shh signaling is important to maintain the function of Sca-1+ cells as SMC progenitors (100).
Chen et al. reported a population of Sca-1+ cells, localized in the vasa vasorum in the adventitia of vein grafts, that possesses the potential to differentiate into SMCs, adipocytes, osteoblasts, or chondrocytes in vitro (23). In a bioreactor system, these Sca-1+ migrate into the decellularized vessel in response to circulating CXCL12 (37). Further, Wong et al. demonstrated that sirolimus, a commonly used drug in drug-eluting stents, can induce Sca-1+ stem/progenitor cells that are isolated from the adventitia of vein graft to migrate and differentiate into SMCs. The authors, thus, suggest a plausible mechanistic cause of restenosis after the application of sirolimus-eluting stents in patients (146).
Regarding the origin of Sca-1+ cells, Majesky et al. (87) provided the first direct in vivo evidence that a proportion of Sca-1+ progenitors can be derived from mature SMCs of the media. Using in vivo cell lineage tracing models of mice to monitor the cell fate of medial SMCs via YFP expression, the authors found that SMCs generated in situ a subpopulation of Sca-1+ progenitor cells in adventitial tissues. Two independent genetic fate-mapping systems were used and combined with the expression of an SMC-specific histone mark. The authors found that ∼8% of the total Sca1+ cell population were YFP+ SMC-derived cells within the adventitia. In contrast to the success of being found in the adventitia of blood vessels, Sca-1+YFP+ cells were not detected in peripheral blood mononuclear cells. In adult mice, it was found that a large percentage (30%–60%) of total Sca-1+ cells in the carotid artery plus aortic arch, descending aorta, and femoral artery originated from SMCs. The observation that a fraction of Myh11-expressing SMCs in the media can be reprogrammed into Sca-1+ progenitor cells in adventitia could suggest the existence of specialized cells with a higher plasticity within arterial walls. These findings highlight the complexity of the nature and origin of vascular-resident stem/progenitor cells and SMCs (Fig. 2).
Sca-1+ Stem/Progenitor Cell Migration via CCL2 and CXCL1
How can adventitial stem/progenitor cells cross the media and migrate to the intima? The hypothesis is that SMCs can be activated by the release of chemokines in response to vascular injury. Supporting this hypothesis is the fact that activated SMCs can release inflammatory cytokines and growth factors, resulting in an inflammatory microenvironment in the vascular wall after vessel injury (43, 108). Many studies have shown that cytokines and growth factors are potent molecules that can mobilize circulating stem/progenitor cells (131). However, only few studies are focused on whether these local inflammatory factors can also activate resident stem/progenitor cells in the vascular wall.
In a recent study (155), we demonstrated that proliferating SMCs can significantly induce vascular stem/progenitor cell migration. In vitro migration assays and single-cell tracking experiments in real time indicated that the cells are migrating directionally and efficiently but not randomly. In vivo, green fluorescent protein (GFP)-Sca-1+ cells were applied to the adventitial side of injured femoral arteries, and after 72 h, GFP+ cells could be easily detected on the intimal side by en face staining; whereas 2 weeks later, neointima formation was significantly increased in these treated vessels compared with injured vessels where no cells had been applied. These data indicate that when either cultured in vitro or injured in vivo, mouse SMCs are transformed into a proliferative phenotype, and in this condition, they are a potent attractant for vascular stem/progenitor migration both in vitro and in vivo.
A multiple chemokine enzyme-linked immunosorbent assay performed on supernatant from cultured SMCs demonstrated that several chemokines were upregulated in activated SMCs. Among them, CXCL1, CCL2, and CCL5 showed the highest increase in secretion. It is also noteworthy that the expression of both CCL2 and CXCL1 is much higher in SMCs compared with other cell types (Sca-1+ stem/progenitor cell, one clone of adventitia stem/progenitor cells) at both the mRNA and protein levels. Further, in vivo, immunofluorescence staining of α-SMA, CCL2, and CXCL1 on femoral arteries revealed that few chemokines are expressed on the intact vessel wall; however, after injury, there is a time-dependent increase in CCL2 and CXCL1 expression, much of which co-localizes with SMC markers. This demonstrates that, once activated, SMCs can release large amounts of CCL2 and CXCL1. Direct stimulation of Sca-1+ cells with recombinant CXCL1 and CCL2 stimulated cell migration.
CXCL1 is a chemokine that has been studied in cardiovascular disease by many research groups in recent years. Ox-LDL has been reported to trigger CXCL1 release from ECs, which recruit atherogenic monocytes and promote atherosclerosis (160). In addition, CXCL1 is expressed on human ECs and induces angiogenesis through enhancement of EC migration and proliferation (89). Microarray data revealed that the basal level of CXCL1 mRNA in untreated mouse aorta SMCs is low, but that it is significantly upregulated after tumor necrosis factor (TNF) or/and α-LTβR treatment (84). CXCL1 is expressed on human arterial SMCs and mediates EPC adhesion in a flow chamber (57). In rat aortic SMCs, the mRNA level of CXCL1 is markedly enhanced by depletion of PTEN (47).
CCL5 expression has been identified on ECs of transplant-associated accelerated atherosclerotic arteries (101). Produced by ECs in response to TNF-α and interferon-γ, it regulates macrophage and T-memory lymphocyte accumulation (88). After artery injury, CCL5 has also been shown to be released by vascular SMCs and to induce an acute vascular inflammatory response through the recruitment of inflammatory cells (70). Previous studies have reported that CCL5 mediates trafficking and homing of T cells, monocytes, basophils, and eosinophils (2, 10, 123, 132). However, the migration of progenitors after stimulation with CCL5 did not change significantly. This result may indicate that CCL5, as a versatile chemokine, is upregulated in SMCs but does not play a functional role in cell migration, suggesting that vascular stem/progenitor cells may selectively or specifically respond to certain chemokines.
CCL2 and CXCL1 and Sca-1+ Vascular Stem/Progenitor Migration In Vivo
Reports using different animal models have shown that CCL2 plays an important role in neointimal hyperplasia (37, 133). In ApoE−/− mice, the immediate release of CCL2 by SMCs in the vessel wall after wire injury was associated with increased platelet adherence to the denuded endothelium, subsequent mediation of monocyte recruitment at an early stage leading to neointima formation (79). It is noteworthy that CCL2-mediated neointimal hyperplasia not only formed in hyperlipidemic animals, for example, ApoE−/− mice, but also occurred in animals with normal blood lipids levels, for example, C57BL/6J mice (37, 48). As mentioned earlier, vascular stem/progenitor cells have been identified as playing a role in neointima formation (23, 59) and migratory progenitor cells are an important cellular component of CCL2-mediated neointima formation. Indeed, when Sca-1+ stem/progenitor cells were applied to the adventitia of wire-injured femoral arteries of CCL2 knockout or wild-type mice, a lower number of stem/progenitor cells migrated to the neointima and reduced lesion formation was observed in CCL2 knockout mice. The majority of cells contributing to neointima formation in this system expressed stem cell marker Sca-1, which was diminished in CCL2 knockout mice (155).
Several studies have demonstrated that bone marrow stromal cells are the main source of CCL2, which plays an important role in myeloma cell homing (139), monocyte mobilization (63), macrophage bone marrow homing (17), and arteriogenesis (65). On the other hand, other studies proved that CCL2 is also released from the vessel wall, by cells such as ECs (127), SMCs (22), and adventitial macrophages. Thus, the cellular source of CCL2 promoting vascular progenitor migration in vivo needs to be further investigated.
By using chimeric mouse models, it was found that after bone marrow transplantation, CCL2 could be detected in the peripheral blood of a CCL2−/− chimeric mouse (bone marrow was isolated from a WT mouse), where it was originally undetectable. This indicates that CCL2 in the blood is released from transplanted bone marrow cells, which confirms the notion that CCL2 is bone marrow derived. However, in a wild-type chimeric mouse (bone marrow was isolated from a CCL2−/− mouse), the levels of CCL2 were markedly upregulated and showed an even higher increase compared with the CCL2−/− chimeric mouse. This indicates that CCL2 from non-bone marrow tissues (e.g., SMC) contributes more to systemic CCL2 levels in the peripheral blood than CCL2 from the bone marrow. The quantitative data from neointimal lesions of each type of chimeric mice showed a consistent increase in neointima area with higher CCL2 levels in the blood, which further confirmed that CCL2 from non-bone marrow tissues induces stem/progenitor cell migration and contributes to neointima formation (155).
CXCL1 has been well established as a potent chemoattractant for leukocyte arrest and migration, which contributes to several inflammatory diseases (12, 111, 160). In cardiovascular disease, human EC-released CXCL1 induces angiogenesis and blockage of CXCL1, resulting in inhibition of EC migration and proliferation, which further abrogates angiogenesis in vivo (82, 89). The roles that CXCL1 plays in neointima formation are distinct in different studies. CXCL1 released from activated SMCs or ECs promotes neointima formation or atherosclerosis (47, 160). Further, CXCL1 is also associated with vascular inflammation, matrix degradation, and lipid deposition that lead to unstable atherosclerotic lesions (14). According to another study, CXCL1 can also mediate the recruitment of circulating EPCs, which contributes to re-endothelialization after arterial injury (57). Blockage of CXCL1 delayed endothelium recovery and increased neointimal area (79). The underlying mechanisms of CXCL1-related neointima formation are still poorly understood. In vivo data revealed that CXCL1 had a robust chemotactic effect on Sca-1+ cell migration but not on ECs and SMCs. When CXCL1 siRNA was applied to the adventitia of injured femoral arteries to create a local silencing of CXCL1, it resulted in a reduction in stem/progenitor cell migration, which indicates that CXCL1 may accelerate neointimal development through induction of stem/progenitor migration (155) (Fig. 3).

Signaling in CCL2- and CXCL1-Induced Sca-1+ Vascular Stem/Progenitor Cell Migration
At inflammatory sites, more than one chemokine exerts biological effects and they may cooperate with each other to activate subsequent downstream signaling events. Several studies have reported synergistic effects of chemokines in leukocyte recruitment and migration, which enhance inflammation both in vitro and in vivo. A single cell can also produce many different chemokines in response to a single stimulus. Since the depletion of either CCL2 or CXCL1 partially inhibits stem/progenitor cell migration, the effects of CCL2 and CXCL1 are not redundant and can act independently in the mediation of cell migration (155).
Even if most studies have reported the expression of CCR2 and CXCR2 on inflammatory cells, such as monocytes/macrophages (74, 144), neutrophils (114), and T lymphocytes (81, 126), several studies have also demonstrated that CCR2 is present on vascular wall cells. CCR2 was detected on human umbilical cord vein ECs and was found to mediate the role of CCL2 in endothelial wound repair (144) and angiogenesis (118). CCR2 was also found to be activated in response to CCL2 in rat aortic SMCs and to alter SMC function in age-related arterial remodeling (130). Yu et al. (155) found that basal levels of CCR2 and CXCR2 expression were much lower on vascular stem/progenitors in comparison to macrophages, potentially due to their quiescent status in conditions without stimuli. However, after treatment with SMC-conditioned medium, both gene and protein levels of the receptors were significantly upregulated. The ability of vascular stem/progenitor cell migration after CCR2 and CXCR2 genes were permanently silenced by shRNA was significantly reduced. Further, the application of antagonists of each chemokine markedly abrogated the influence of SMC on stem/progenitor cell migration. Thus, the data provide the first evidence of the induction of CCR2 and CXCR2 expression in vascular stem/progenitor cells in response to SMC-derived soluble factors through CCL2/CCR2 and CXCL1/CXCR2 axis (155).
Cdc42, Rac1, and RhoA are the main Rho GTPase family members that are known to regulate the formation of lamellipodia, filipodia, and focal adhesions, respectively, thus orchestrating subsequent cell movement (96). Some reports have demonstrated that chemokines induce cell migration via activation of Rho GTPase family members. RhoA, Rac1, and Cdc42 are all activated in SDF-1-induced human gastric cancer cells and T cell migration (21, 33). CXCL12 activates Rac1 as it regulates lung alveolar epithelial cell migration (49). Depletion of Cdc42 in dendritic cells resulted in a reduction of cell migratory abilities driven by CCL19, and in comparison with WT cells, their velocity decreased by 20%, but directionality dropped by 40% (72). Inhibition of RhoA by activity by using a dominant-negative mutant can significantly downregulate CCL19-elicited T cell migration (115).
It was found that in vascular stem/progenitor cells, both Cdc42 and Rac1 were activated by either CCL2 or CXCL1 via their respective receptors CCR2 and CXCR2. The inhibition of either Cdc42 or Rac1 impaired stem/progenitor cell migration in response to CCL2 or CXCL1. Addition of an RhoA inhibitor did not alter CCL2- and CXCL1-driven migration, which revealed that the RhoA signaling pathway is not involved in stem/progenitor cell migration (155).
In addition, p38 MAPK, as a stress-activated protein kinase, has been widely reported to be associated with cell migration in response to extracellular stresses. Aberrant shear stress induces human MSC migration toward wounded areas through the CXCL12/CXCR4/p38 MAPK signaling pathway (83). p38 phosphorylation is also involved in CXCL1-induced leukocyte migration in mice (19). Cytokines such as IL-4 and TNF-α can activate stress-activated pathways, leading to phosphorylation of p38 MAPK, which is dependent on Rac1 and Cdc42 (145). PAK1, as an established effector of Rac1 and Cdc42, mediates tracheal SMC migration by activation of the p38 MAPK pathway (32).
In vascular stem/progenitor cells, p38 phosphorylation was upregulated in response to mouse recombinant CCL2 or CXCL1, and it was suppressed by CCL2/CXCL1 knockdown or CCR2/CXCR2 inhibition, which indicates that CCL2 and CXCL1 activate p38 phosphorylation via CCR2 or CXCR2. These results suggest that stem/progenitor cell migration is induced by CCL2 or CXCL1 through the p38 MAPK signaling pathway. It was also found that p38 phosphorylation was markedly downregulated by Rac1 inhibition but not inhibition of Cdc42 or RhoA, suggesting that stem/progenitor migration is induced via an Rac1/p38 signaling pathway (155). Although migration assays showed the participation of Cdc42 in stem/progenitor cell migration, it may act through an as yet unidentified signaling pathway. In squamous cell carcinoma of the head and neck, for example, CCL19 activates ERK1/2 and JNK, but not p38 phosphorylation via the PI3K/Cdc42 signaling pathway during cell migration (82, 159).
FAK, paxillin, and vinculin are critical in the response to various extracellular stimuli (e.g., chemokines) regulating cell adhesion molecule and cytoskeleton structures and to process the signaling pathways for cell mobility. It has been reported that CXCL12 can induce a pro-adhesive response in progenitor B cells through FAK phosphorylation (28, 50) and that the CXCL12/CXCR4/FAK signaling pathway regulates pro-B cell development (98). In another study, CXCL8-induced FAK phosphorylation is adhesion dependent in CXCR2-expressing cells, but adhesion independent in CXCR1-expressing cells (28), and phosphorylated FAK can subsequently activate paxillin, which plays an important role in leukocyte migration (27). Further, CCL5 was reported to activate T lymphocytes via FAK, paxillin, and ZAP-70 compound (5). MCP-1-induced lymphocyte adhesion to ECs regulates vinculin and paxillin expression in lymphocytes (93). With vascular stem/progenitors, immunofluorescence staining revealed that chemokine-induced FAK phosphorylation, paxillin, and vinculin are controlling stem/progenitor migration. The signaling pathways regulating stem/progenitor cell migration and involving interactions between CCR2/CXCR2, cytoskeleton proteins, and Rho GTPase are illustrated in Figure 3.
Endothelium Regeneration by Vascular Stem/Progenitor Cells
Re-endothelialization is a self-repair process after injury that is used to maintain the protective role of endothelium in the vessel wall; it includes proliferation and migration of adjacent intact ECs, resident vascular stem/progenitor cells, and recruited stem cells from the bone marrow. Resident vascular-derived ECs have been proved to contribute to re-endothelialization much more effectively in response to damaging stimulation than stem cells originating from the bone marrow (11). The vasculogenic zone is located between the adventitia and the media, where cells identified as CD34+CD31− are described as progenitors with the ability to differentiate into ECs, to participate in blood vessel formation and progression of atherosclerosis (1, 58, 156). CD34+CD31− adventitial cells can interact with ECs and promote capillary-like structure formation and stabilization (16). When adventitial cells were injected into a hind limb ischemia mouse model, a significant pro-angiogenic effect (a full blood flow recovery) was observed as early as 7 days after injection. This indicates that adventitial cells have therapeutic potential in angiogenesis/vasculogenesis (16). Transplantation of adventitial cells can repair infarcted hearts through angiogenesis (64). Taken together, these recent data demonstrate that adventitial progenitor cells have the ability to repair vascular damage in ischemic tissue.
In response to injury or the development of atherosclerotic plaques, resident stem/progenitor cells will have different structural and functional behaviors (53). In atherosclerosis, adventitial inflammation and angiogenesis are widespread and advanced plaques are vulnerable to rupture and thrombosis formation. In normal conditions, stem/progenitor cells are involved in maintaining homeostasis of the physiological vasculature. On the other hand, stem/progenitor cells may also supply cells that contribute to neointimal lesion formation and atherosclerosis.
Vascular Stem/Progenitor Cells Contribute to Neointima Formation
SMCs are well established as the main source of cells contributing to restenosis. In recent years, many studies have demonstrated that a group of adventitial stem/progenitor cells participate in lesion formation. Some studies report that inflammatory cytokines or growth factors play a chemotactic role in stem/progenitor cell migration. EC-derived PDGF-B induces migration and proliferation of vascular stem/progenitor cells expressing PDGFRβ during embryonic vessel development (56). An ex vivo model showed that stem/progenitor cells isolated from vein graft could migrate into decellularized vessels in response to SDF-1 via CXCR4 (23). Subsequently, another study demonstrated that the migratory ability of these vascular stem/progenitor cells was significantly enhanced by sirolimus via CXCR4 activation, which elucidated a new mechanism behind sirolimus-eluting stent-induced restenosis (146). Further, metalloproteinase-8 expressed on stem/progenitor cells facilitated their migration and subsequent contribution to atherosclerosis via the MMP8/ADAM10/E-cadherin signaling pathway (151). EC-derived microparticles and non-microparticle secreted factors can induce perivascular MSC proliferation and migration by activating NF-κB signaling (85). However, the exact mechanisms of vascular stem/progenitor cell migration and subsequent neointima formation remain to be fully elucidated.
Vascular Stem/Progenitor Cells Differentiate into SMCs
SMC accumulation and alteration in response to atherogenic factors play a dominant role in atherosclerotic plaque development. The historical view of SMCs in atherosclerosis is that phenotypic switching of the pre-existing SMCs results in SMC de-differentiation and aberrant proliferation, contributing to plaque formation (51, 95). However, besides mature SMC plasticity, considerable evidence to date indicates multiple origins of SMCs within the atherosclerotic lesions, including adventitial fibroblasts trans-differentiation (30, 119), endothelial-to-mesenchymal transition (91), and a variety of stem/progenitor cell differentiation (86, 106). Studies related to vascular SMC-relevant stem/progenitor cells in atherosclerosis have been vastly expanded in recent years. In early studies, Sata et al. (120) reported that a part of α-SMA+ cells within atherosclerotic lesions originated from bone marrow, implying that circulating HSCs are capable of differentiating into SMCs during plaque formation. However, in contrast, using more specific vascular SMC markers and rigorous lineage tracing methods, subsequent studies demonstrated that SMCs of bone marrow-derived cell origin is exceedingly rare in lesions and contributing negligibly to vascular remodeling (9, 62).
To date, it is increasingly considered that local vascular stem/progenitor cells, mostly residing in vessel wall stem/progenitor cell niches, serve as the sources of SMCs in atherosclerosis. It is well established that vascular stem/progenitor cells could differentiate into SMCs in vitro (59, 117). When LacZ-labeled Sca-1+ cells were transferred to the adventitial side of vein grafts in ApoE−/− mice, SMCs with β-gal expression were found in atherosclerotic lesions and made up 30% of neointimal cells. Further, local application of adventitial Sca-1+ cells onto decellularized vascular scaffolds showed a population of stem/progenitor cells and progenitor cell-derived SMC accumulation in developing neointima (59). A lineage tracing study indicates that more than 50% of neointimal SMCs are derived from adventitial MSCs (71). Therefore, these findings suggest that adventitial resident stem/progenitor cells are capable of differentiating into SMCs and of participating in atherosclerotic progression.
In addition to adventitial stem/progenitor cells, distinct populations of mural SMC precursors were also observed in the media. Sainz et al. (117) identified and isolated progenitor cells with Sca-1+ c-kit−/low Lin− CD34−/low profile from the tunica media of adult mice aortas that had potency for SMC differentiation. Tang et al. (134) announced the discovery of multipotent vascular stem/progenitor cells in the medial layer of arteries. Such multipotent vascular stem/progenitor cells uniformly expressed stem cell markers, including Sox17, Sox10, and S100β, and could differentiate into an intermediate stage of MSC-like cells that subsequently differentiated into SMCs. Notably, in response to vascular injury in vivo, multipotent vascular stem/progenitor cells highly proliferated and differentiated into SMCs, which were responsible for neointima formation. These studies support the existence of medial stem/progenitor cells and their SMC potential in atherosclerotic diseases. Although these preliminary data implicate a role for SMCs of a vascular-resident stem/progenitor cell origin in atherosclerosis, further investigation is still needed. Importantly, SMC progenitors or specific subpopulations are still poorly defined by surface markers, which probably cause inconsistency in studies. Direct evidence precisely elucidating the functional involvement of stem/progenitor cell differentiation in pathophysiological conditions is absent and highlights a critical challenge.
Limited publications are available regarding the signal-transduction pathways that are involved in SMC differentiation of vascular-resident stem/progenitor cells. Using progenitor cells derived from ESCs, it was demonstrated that several cellular actors orchestrate cell differentiation toward an SMC lineage. Xiao et al. (150) reported that ESC-derived Sca-1+ progenitor cells cultured in a collagen IV-coated flask can be induced toward SMC differentiation, which depends on the interaction with integrin α1β1αv and the activation of FAK/paxillin-PI3K-MEK-ERK/JNK and PDGFR signaling pathways. According to this method, more than 95% of differentiated SMCs can be obtained for more than 30 days of continuous culture (150). Based on an SMC differentiation protocol established in our group, Chen et al. demonstrated that adventitial Sca-1+ progenitor cells were also capable of differentiating into SMCs through the interaction of collagen IV with integrin α4α5β1 (23). Further, Wong et al. confirmed the SMC differentiation potency of this cell type by showing the evidence that under sirolimus stimulation, vein graft adventitial Sca-1+ progenitor differentiation into SMCs is mediated by epidermal growth factor receptor and activation of extracellular signal-regulated kinase, followed by β-catenin nuclear translocation (146). Moreover, in other groups, several studies also proved that other signaling pathways play important roles in vascular wall progenitor cell differentiation into an SMC lineage. A study revealed that HOX genes are essentially involved in SMC differentiation of CD44+ vascular wall progenitor cells. Specifically, knockdown of HOXB7 and HOXC6 can induce the expression of SMC genes TAGLN and CNN1 by alteration of CpG methylation of the TAGLN promoter, leading to SMC differentiation (66).
Besides, microRNA also plays a key role in regulating the differentiation program of stem/progenitor cells. Zhao et al. showed that microRNA 22 upregulated by both PDGF-BB and transforming growth factor-β inhibited the repressive effects of Methyl CpG-binding protein 2 on SMC transcriptional factors, such as SRF and myocardin, initiating SMC differentiation in Sca-1+ adventitial progenitor cells (158). In addition, it was found that under PDGF-BB stimulation, 17β-estradiol can increase myocardin-SRF complex binding to the SM22 gene that was mediated by enhancement of SRC3 expression, which further promoted differentiation of CD34+ vascular wall stem cells into SMCs (148). The different signal transduction pathways known to be involved in the differentiation of progenitor cells into SMCs are briefly illustrated in Figure 4. As the research on the vascular wall-resident stem/progenitor cells aspect is still new, only few signaling pathways mediating SMC differentiation have been identified. In this regard, several published review articles focusing on other kinds of stem cells are available and can be used for reference (18, 103, 142, 157).

Stem/Progenitor Cells Differentiate into Monocytes/Macrophages
It is well accepted that atherosclerosis is a chronic inflammatory disease (78). The leukocytes, especially monocytes/macrophages, are of critical importance for atherogenesis, by substantially accumulating in atheroma lesions and adventitia where they regulate plaque progression and stability (92, 94). Although traditional notions assert that vascular monocytes/macrophages are solely recruited and differentiated from bone marrow-derived HSCs (92), recent evidence provides an alternative origin for macrophages during atherosclerosis. Robbins et al. (112) revisited the monocyte influx and macrophage accumulation in atherosclerosis. Intriguingly, they found that rapid local macrophage proliferation, independently of monocyte recruitment, contributes predominantly to lesional macrophage accumulation in established atherosclerotic plaques.
On the other hand, an ex vivo aortic ring model showed extensive outgrowth of CD45+CD68+ mononuclear cells with features of macrophages, leading to the inference that aortic adventitia contained immature cells that are capable of differentiating into macrophages (162). Single aortic cells disaggregated from mice presented the capacity of generating a full spectrum of multipotent and lineage-specific hematopoietic colony-forming units, especially predisposed to macrophage colonies, demonstrating the existence of rare multipotent hematopoietic stem/progenitor cells or macrophage precursors in the arterial wall (104, 105). These aortic stem/progenitor cells were adventitiously located and co-expressed Sca-1 and CD45, whereas Sca-1+CD45− cells had no hematopoietic capacity. Adaptive transfer experiments revealed that these adventitial Sca-1+CD45+ macrophage progenitor cells were not replenished or diminished by an alteration of bone marrow cells or circulation-derived monocytes/macrophages; they contributed durably to macrophage progeny in the adventitia, and, to a lesser extent, the atherosclerotic lesions of arteries (105). However, whether they are specifically converted into monocytes/macrophages in atherosclerosis requires further investigation. A combination of definitive progenitor cell marker fate-mapping and vascular or adventitia graft transposition models could provide valuable data to address the important question.
Stem/Progenitor Cell Differentiation Contributes to Calcification
Vascular calcification, which is highly prevalent in the advanced phase of atherosclerosis, is now established to be a multi-faceted pathological process that is characterized by ectopic bone formation and/or mineralized matrix deposition within all vascular layers, as well as atheroma lesions (13, 116). The presence of chondrocyte-, osteoblast-, and osteoclast-like cells in the vessels posed questions on the origin of these calcifying cells (34). Various potential sources of cells undergoing osteochondrogenic differentiation have been reported, including SMCs (129), pericytes (29), ECs (153), adventitial myofibroblasts (24), and stem/progenitor cells (13, 25, 106).
In particular, current knowledge underlines the importance of circulating and resident stem/progenitor cells in triggering vascular calcification through their differentiation into osteocytes. Eghbali-Fatourechi et al. (38, 39) discussed the presence of circulating osteoblast-lineage cells that correlated with physiological bone formation. These osteo-progenitors may seed the arteries and be involved in vessel ossification (121). The novel origin of vascular calcified cells was termed “Circulating Cell Theory,” suggesting the contribution of bone marrow-derived circulating osteo-progenitor cells to vascular calcification (44, 97). Subsequently, Cho et al. (25, 26) identified vascular Sca-1+ (Sca-1+/PDGFRα+ and Sca-1+/PDGFRα−) progenitor cells, exhibiting great osteoblastic and/or osteoclastic potentials, as calcifying progenitor cells that significantly contributed to calcified atherosclerotic plaques. In addition, circulating nestin+Sca-1+ MSCs displayed osteoblast-forming potential that they could be recruited to the arterial lesions and differentiate into osteocytes (55). Another source of calcifying progenitor cells are vascular wall-resident stem/progenitor cells that are located in vascular niches of the adventitia (106). An atherosclerotic or inflammatory plaque environment significantly increased the tendency of circulating/resident progenitor cells to differentiate into an osteogenic lineage, which is one of the mechanisms underlying the high incidence of vascular calcification in atherosclerosis (75).
Summary and Perspectives
Recently, a population of stem/progenitor cells has been identified within the vessel wall. These cells are found to express some stem cell markers, including Sca-1, c-kit, CD34, and CD90, and also some markers such as CD73 and CD45 that are expressed on MSCs. They can either play a reparative role in atherosclerosis or exacerbate neointima formation. Previous studies have shown that these cells possess high potential to differentiate into many cell lineages, including endothelial and SMCs (17, 146). The mechanism of migration and the functional roles of these cells crossing the vessel wall have been studied. It is known that these stem/progenitor cells exist in close contact with SMCs, a population of cells that have been established to play important physiological and pathological roles within the media of the vessel wall. SMCs are able to release specific chemokines to induce the migration of vascular stem/progenitor cells that further contribute to neointima formation. Meanwhile, studies from different laboratories have shown that vascular stem/progenitor cells can differentiate into various cell types such as ECs or SMCs to participate in vascular repair or neointima formation, depending on a variety of stimulations under specific circumstances.
Nevertheless, a conundrum remains that the identification and characterization of adventitia stem/progenitor cells still lacks consistency between different research groups. To date, there remains a lack of specific progenitor cell markers that can exclusively characterize and distinguish these progenitor cells from other cell types. Further, several important questions still need to be addressed: (i) What are the differentiation capacities of the vascular stem/progenitor cells into different cell types? (ii) What are their roles in vascular repair, re-endothelialization, and neointima formation? (iii) Do their responses to stimuli differ while they are in the same microenvironment? (iv) Why are the conclusions from different research groups so varied? Thus, there is still much progress to be made in the field of stem/progenitor cell research. Although some stem/progenitor cells are identified by specific markers using in situ immunostaining in the vessel wall or antibody selection from isolated cells, it is still uncertain whether they are activated after vascular injury and differentiate to play a role in vascular events. The current models of differentiation in vitro are unable to replicate the dynamic and complicated circumstances that surround and influence stem/progenitor cells in vivo.
Finally, despite the expansion of our knowledge of stem/progenitor cells in atherosclerotic calcification, some major issues still remain unknown. Are stem/progenitor cells good or bad for vascular calcification? Sca-1+/PDGFRα− cells showed bidirectional differentiation potential toward osteoblasts and osteoclasts, with the former cell type increasing ectopic bone formation, whereas the latter reversed calcified atherosclerotic plaques (141). The challenge is to comprehensively uncover the molecular mechanisms determining the differentiation direction of vascular stem/progenitor cells in pathological conditions and to translate our understanding into therapeutic strategies for atherosclerosis.
Acknowledgments
Vascular stem/progenitor cell research in the Xu lab was supported by the British Heart Foundation and Oak Foundation. This work was supported by the National Natural Science Foundation of China (91639302, 91339102, and 91539103) and the Zhejiang Provincial Natural Science Foundation (LR14H020001).