
Abstract
Significance:
Ascorbate is an essential antioxidant in plants. Total contents and its redox state in organelles are crucial to fight and signal oxidative stress.
Recent Advances:
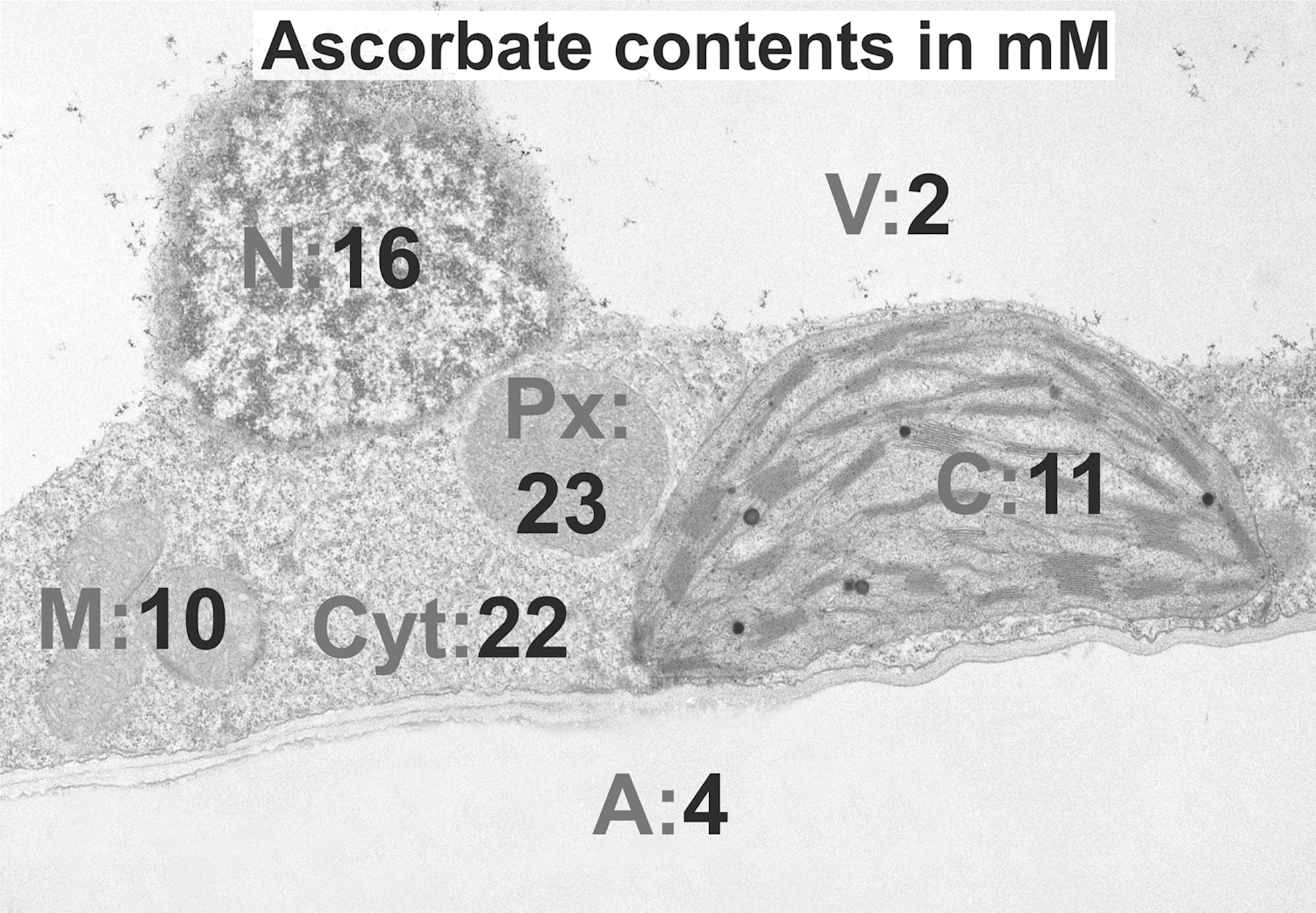
With quantitative immunoelectron microscopy and biochemical methods, highest ascorbate contents have recently been measured in peroxisomes (23 mM) and the cytosol (22 mM), lowest ones in vacuoles (2 mM), and intermediate concentrations (4–16 mM) in all other organelles.
Critical Issues:
The accumulation of ascorbate in chloroplasts and peroxisomes is crucial for plant defense. Its depletion in chloroplasts, peroxisomes, and mitochondria during biotic stress leads to the accumulation of reactive oxygen species (ROS) and the development of chlorosis and necrosis. In the apoplast and vacuoles, ascorbate is the most important antioxidant for the detoxification of ROS. The cytosol acts as a hub for ascorbate metabolism as it reduces its oxidized forms that are produced in the cytosol or imported from other cell compartments. It is a sink for ascorbate that is produced in mitochondria, distributes ascorbate to all organelles, and uses ascorbate to detoxify ROS. As ascorbate and its redox state are involved in protein synthesis and modifications, it can be concluded that ascorbate in the cytosol senses oxidative stress and regulates plant growth, development, and defense.
Future Directions:
Future research should focus on (i) dissecting roles of ascorbate in vacuoles and the lumen of the endoplasmic reticulum, (ii) identifying the physiological relevance of ascorbate transporters, and (iii) correlating current data with changes in the subcellular distribution of related enzymes, ROS, and gene expression patterns.
Introduction
A
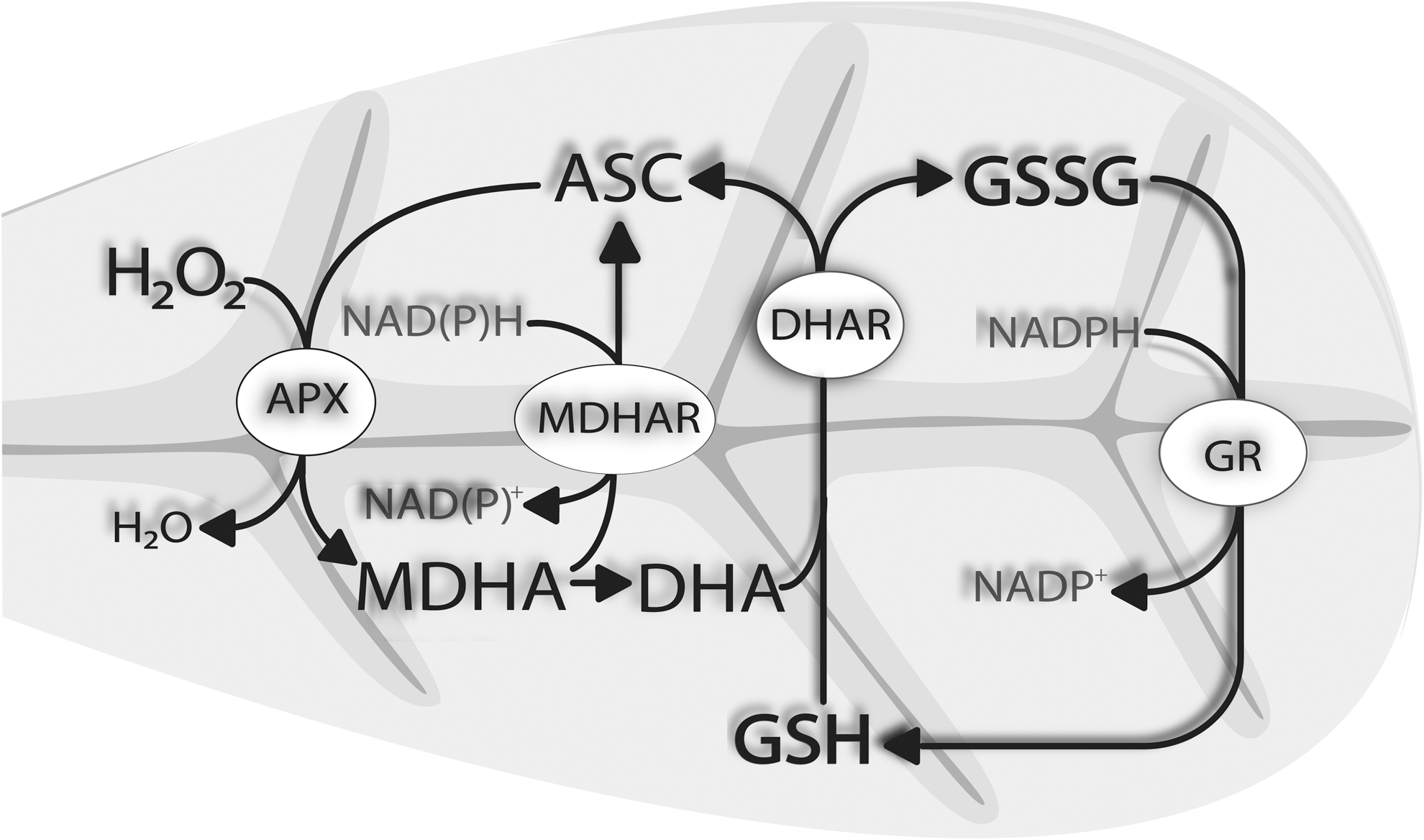
During the detoxification of ROS, the reduced form of ascorbate (ASC) gets oxidized to monodehydroascorbate (MDHA), which either gets reduced by monodehydroascorbate reductase (MDHAR) to ASC or, as very unstable, forms dehydroascorbate (DHA). DHA is reduced by dehydroascorbate reductase (DHAR) to ASC. Oxidized glutathione, which is formed during this reaction, is reduced by glutathione reductase (Fig. 2).
Inside cells of nonstressed leaves, more than 90% of ascorbate occurs in its reduced state whereas only between 10% and 50% occurs in its reduced state inside the apoplast (89). During environmental stress conditions, this ratio and total ascorbate contents are subject to change and large amounts of (oxidized) ascorbate can accumulate within certain cell compartments such as vacuoles, plastids (42, 50, 59, 60, 67, 68), and the apoplast (63, 118 –120). Thus, the ratio of ASC to DHA/MDHA within and between organelles (e.g., redox state of the apoplast vs. the cytosol) is essential for the ability of the plant to fight and signal oxidative stress.
Ascorbate plays essential roles in the protection of plants against abiotic stress, such as ozone (O3; 17, 22, 104), exposure to high light (8, 38, 50), drought (59), and exposure to heavy metals (21, 60, 100), and biotic stress such as infections with bacteria (42, 68) and fungal pathogens (67, 108, 118 –120). As all of these stress conditions induce highly compartment-specific stress (e.g., high light affects chloroplasts first, heavy metals get detoxified mainly in the cytosol, and pathogens and O3 target the apoplast before other cell compartments are affected), small shifts in the subcellular distribution of ascorbate can have major impacts on the plant.
Changes in the compartment-specific distribution of ascorbate, especially during environmental stress conditions, can reveal essential information about defense reactions and the occurrence of oxidative stress in certain organelles. In addition, subcellular changes in ascorbate contents and its redox state can be used to characterize the protective functions of ascorbate in individual cell compartments and can serve as a subcellular stress marker during environmental stress situations.
Even though different pathways of ascorbate synthesis have been proposed in plants, the
It has been proposed recently that a side branch of this pathway facilitated through GDP-
Another pathway of ascorbate synthesis in higher plants is facilitated through galacturonate and glucuronate (71). Nevertheless, this pathway has so far been found only in kiwi and not all genes and enzymes have been identified. In addition, the compartmentation of this pathway within plant cells is still completely unclear. Thus, the
Although total ascorbate contents in leaves can be as high as 10 mM (18, 121), they can reach values as high as 23 mM in peroxisomes and as low as 2 mM in vacuoles (131). Ascorbate has also been detected in other cell compartments such as mitochondria, plastids, nuclei, the cytosol, and the apoplast (118 –121, 131). As the last step of ascorbate biosynthesis in higher plants takes place in mitochondria, and as ascorbate occurs in all cell compartments, specific transporters must be located in membranes of all organelles and the plasma membrane to facilitate the transport of ascorbate.
The nucleobase ascorbate transporter (NAT) family has been identified in Arabidopsis, rice, and tomato (16, 79). Although the localization of OsNAT10 and 11 in rice was predicted in the envelope of plastids, AtNAT12 seems to be present in the plasma membrane (79). In tomato, 12 members of the NAT family named SINATs have been identified and it has been predicted by sequence analysis that they are localized in the plasma membrane, chloroplast thylakoid membrane, Golgi body, and the membrane of the endoplasmic reticulum (ER; 16). In Arabidopsis, a member of the phosphate transporter 4 family (AtPHT4; 4) has been recently identified to transport ascorbate through the envelope membrane of the chloroplast (82).
Ascorbate concentrations are most commonly measured with biochemical methods in individual plant organs and tissues. Thus, the obtained values in whole organs or tissues represent a combination of ascorbate from all individual cell compartments and cannot be used to interpret the situation in single cells or organelles. Such data can even lead to misinterpretation of stress responses if, for example, an accumulation of ascorbate in the apoplast is accompanied with a similar decrease in ascorbate contents in plastids. In such situations, total ascorbate contents might remain unchanged when measured in leaf extracts, although a stress response on the subcellular level occurs. Thus, to gain unambiguous data of defense responses in plants, it is essential to measure ascorbate contents on the subcellular level.
Methods for the Compartment-Specific Analysis of Ascorbate
The subcellular detection of ascorbate is technically challenging as it is water soluble and can, therefore, be easily washed out and/or redistributed during sample preparation. In addition, sample preparation itself can be seen as a stress to the plant, which can also contribute to a redistribution, accumulation, or reduction of ascorbate on the subcellular level. Currently, there are two major approaches that have been used to study the compartment-specific distribution of ascorbate in plants: (i) biochemical analysis after the extraction of the apoplastic fluid, and isolation of individual organelles (52, 53, 63, 111, 118 –121) and (ii) light and electron microscopical investigations after histochemical labeling with silver (73) or ascorbate-specific antibodies (131).
Biochemical methods have been used to measure ascorbate contents in isolated chloroplasts (67), mitochondria, peroxisomes (52, 53, 68 –70), and vacuoles (111, 112). Ascorbate was also detected with biochemical methods in the apoplast of barley, birch, and poplar leaves (63, 118 –121). Data for the cytosol and nuclei are not available. With biochemical methods, it is possible to differentiate between ASC, MDHA, and DHA and to measure their concentrations. As this methodology also enabled the simultaneous analysis of glutathione and related antioxidative enzymes, they delivered valuable insights into the redox state and modifications of the ascorbate-glutathione cycle on the subcellular level during environmental stress situations.
However, when preparing samples for biochemical detection of ascorbate, the apoplastic fluid has to be extracted or individual cell compartments have to be isolated from a large amount of samples before they can be analyzed with high-performance liquid chromatography or photo-spectrometry (52, 53, 63, 66 –70, 111, 118 –121). This involves long procedures with separation and centrifugation steps of up to 20,000 g (52, 68), which can result in the loss or re-distribution of ascorbate between organelles, oxidation of ascorbate, and the contamination of nonorganelle-specific ascorbate (and glutathione) and related enzymes (20, 65, 68, 88, 121). The application of compartment-specific markers for the isolated organelles is, therefore, crucial to obtain clear results as it helps to evaluate whether and to what extent a re-distribution of ascorbate occurs between organelles during sample preparation (121).
Light microscopy and histochemical staining of ascorbate has been used to detect ascorbate at the nuclear membrane, in nucleoli, and along the plasma membrane in roots of Cucurbita plants (73). In the same study, ascorbate oxidase (AO) was localized in cell walls and vacuoles. AO is a blue copper protein and is considered one of the main enzymes facilitating and controlling the oxidation of ASC to DHA in the apoplast (73). The major limitations of these methods are the resolution of the light microscope (about 200 nm) and the ability of the stain to infiltrate the samples and to react with ascorbate. As this method relies on the reduction of silver ions by ASC that form metallic silver precipitates, it can only be used to detect ASC. In addition, there is a significant risk that other reducing agents in the cell form silver metal precipitates, which can lead to unspecific staining (73). As staining takes 5 days at 4°C, it remains unclear as to what extent the obtained distribution of silver precipitates reflects the in vivo situation of ascorbate in the different organelles. Ascorbate could be re-distributed during the lengthy procedure and washed out of the apoplast. In addition, it is difficult, if not impossible, with this method to localize ascorbate in small organelles (e.g., mitochondria, peroxisomes), the ER, intrathylakoidal spaces, and within cristae.
An alternative to the earlier described methods is the localization and quantitative analysis of ascorbate after immunogold labeling and computer-supported transmission electron microscopy (TEM; 131). This method is based on primary antibodies that bind specifically to ascorbate on ultrathin tissue sections and gold conjugated secondary antibodies that bind to the primary antibody. In the TEM, gold particles can be identified and quantified with image analysis in different cell compartments (gold particles per μm2) on a high level of resolution (Fig. 3). In combination with biochemical measurements of ascorbate contents, these methods even allow the calculation of subcellular ascorbate concentrations in millimolar (Fig. 1). Highest ascorbate contents were calculated in peroxisomes (23 mM), followed by the cytosol (22 mM), nuclei (16 mM), chloroplasts (11 mM), mitochondria (10 mM), and vacuoles (2 mM).
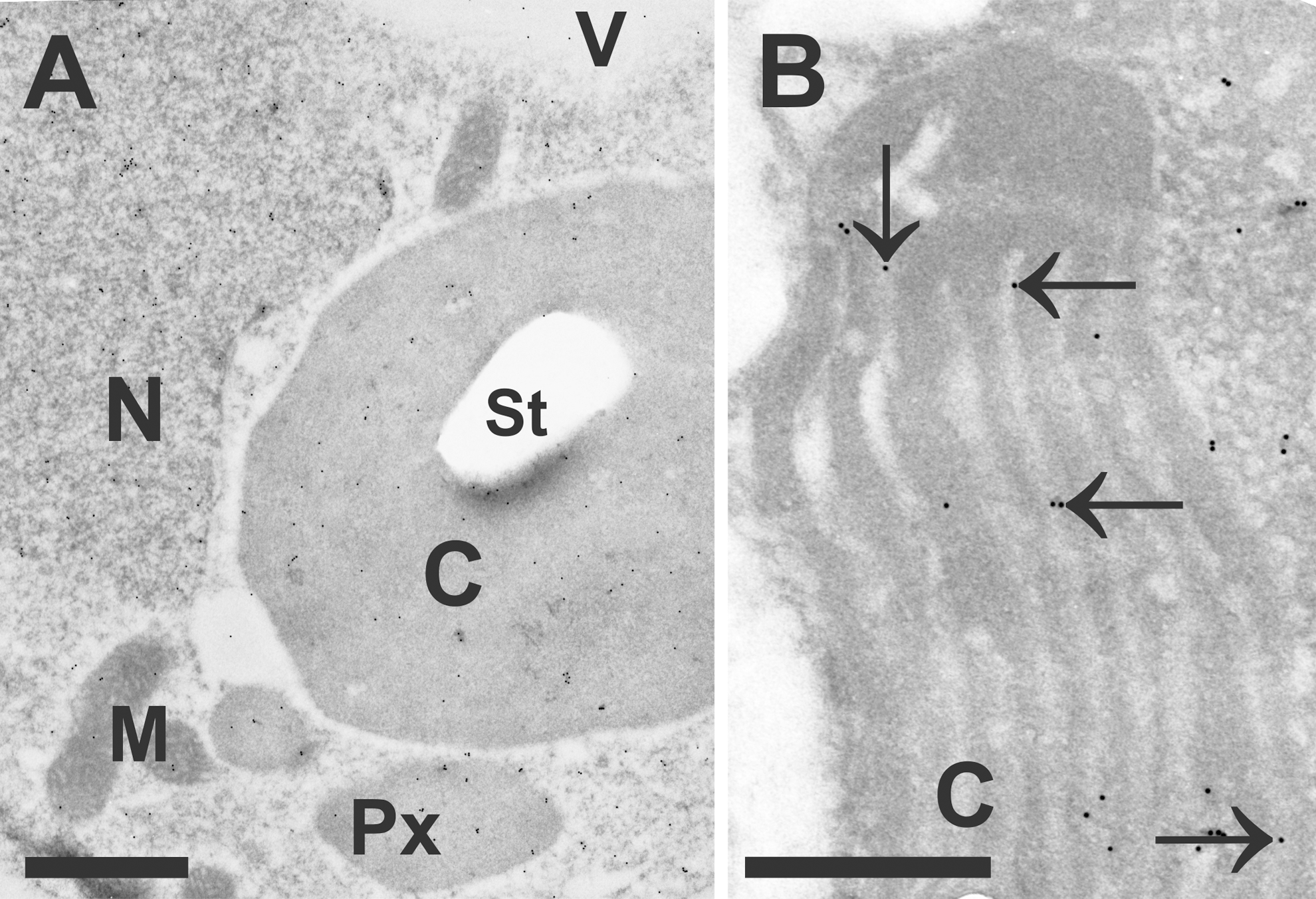
Inside mitochondria and chloroplasts, ascorbate was detected in nonstressed cells within the matrix and stroma, respectively, but it was absent within the lumen of thylakoids or cristae (131). This distribution changed after the application of high light and drought stress when ascorbate was also detected inside the lumen of thylakoids (Fig. 3B). In contrast to biochemical investigations, ascorbate could not be detected in the apoplast of mesophyll and epidermis cells with microscopical methods (73, 131). On the other hand, it could be detected in the apoplast of vascular bundle cells with TEM after the application of cytohistochemical immunogold labeling and in the plasmodesmata of mesophyll cells (131). Thus, either ascorbate contents are below the level of detection for the electron microscopical approach in the apoplast of mesophyll cells or biochemical measurements of apoplastic ascorbate contents only reflect the situation of vascular bundle cells and/or also include contamination from the cytoplasm.
Due to the high resolution of TEM (around 0.2 nm), it was also possible to detect ascorbate in very small cell structures such as the ER and to investigate their localization at membranes such as the plasma membrane, thylakoids (Fig. 3B), and cristae (131). In addition, this method enabled simultaneous investigations of different leaf tissues (e.g., upper and lower epidermis, mesophyll, and vascular tissue; 131) where a similar compartment-specific distribution of ascorbate was found. Ascorbate was further localized in plasmodesmata, companion cells, sieve elements, and also in xylem vessels (131), supporting the observation that it is transported within the plant and exchanged between different organs through vascular bundle cells (35). The section thickness (50–120 nm) for quantitative immunoelectron microscopy results in a very high spatial resolution that can be of benefit if local events are of interest, such as the penetration site of fungi (108), or aphids (130), or if one wants to investigate single cells such as pollen grains (131) or trichomes (62).
The main limitations of this method are that samples have to go through sample preparation, which includes fixation, dehydration, and embedding in resin (131). Similar to biochemical methods, it is, therefore, impossible to investigate the compartment-specific distribution of ascorbate in vivo. As this method employs the use of primary and secondary antibodies, the specificity and accuracy of the distribution of ascorbate has to be carefully and intensively evaluated (131). This was achieved by using different negative controls (omission of secondary antibody, use of pre-immune serum instead of the primary antibody, saturation of the primary antibody with ASC or DHA), using different methods of sample preparation, and comparing the subcellular distribution of ascorbate between plants with different ascorbate contents (131). From these studies, it was concluded that a re-distribution or loss of ascorbate through diffusion during fixation can be excluded.
In addition, subcellular ascorbate contents in the ascorbate-deficient vtc1-2 and vtc2-1 were found to be up to 54% and 76% lower, respectively, when compared with wildtype plants. This correlated well with biochemical methods that measured up to 40% and 90% less ascorbate in leaf extracts from vtc1 and vtc2 when compared with the wildtype (6, 22, 131). These mutants are characterized by a mutation on the VTC1 locus that controls the activity of GDP-mannose pyrophosphorylase, which is required for ascorbate synthesis (6, 22, 87). Another limitation of this method is that the antibodies against ascorbate are currently not able to differentiate between ASC, DHA, and MDHA. Thus, only total ascorbate contents can be analyzed on the compartment-specific level (131). Summing up, despite the limitations of the earlier described methods, they have all contributed toward a better understanding of the compartment-specific distribution of ascorbate and helped to dissect the subcellular roles of ascorbate during abiotic and biotic stress.
Compartment-Specific Response of Plants to Environmental Stress
Changes of ascorbate contents in plants induced by environmental stress situations such as excess light, high O3 levels, heavy metals, UV radiation, and pathogens are well documented (4, 8, 17, 23, 37, 38, 46, 66, 68, 84, 86, 108). However, the majority of these studies has investigated the situation in whole organs or tissues, rather than in single cells and organelles. Therefore, very little data are available about changes of compartment-specific ascorbate contents during environmental stress conditions. Considering that most of these conditions induce highly compartment-specific stress before the whole plant is affected, such information is essential to understand the protective functions of ascorbate on the subcellular level and inside the plant as a whole.
Chloroplasts and Peroxisomes
The importance of ascorbate in chloroplasts and peroxisomes has been extensively studied with both methods mentioned earlier during abiotic (50, 59 –61) and biotic stress (42, 52, 67, 69, 70, 108). Among all cell compartments, peroxisomes were found to contain the highest levels of ascorbate (23 mM), which were about twice as high as in chloroplasts (Fig. 1). Chloroplasts are the prime target of abiotic stress conditions such as high salinity, drought, and excess light. Metabolic changes can be observed in this cell compartment before other cell compartments are affected (3, 56, 95).
Drought, high salinity, and excess light stress induce the closure of stomata, which negatively interferes with gas exchange between leaves and the atmosphere (10, 49). Under these circumstances, low levels of CO2 inside the leaves favor the transfer of electrons to O2, which results in the generation of ROS in chloroplasts (3, 39, 47, 64, 98, 105, 113). This situation activates photorespiration, which induces the formation of phosphoglycolate, which gets degraded in peroxisomes where it leads to the generation of hydrogen peroxide (H2O2; 10, 33, 49, 81). The exposure of cadmium (Cd) leads to similar disturbances described earlier, such as stomata closure and, subsequently, CO2 deficiency, the inhibition of the activity of some enzymes of the Calvin cycle, and disturbs the production of chlorophyll and carotenoids (11, 24, 28, 31, 40, 41, 48).
The importance of ascorbate in chloroplasts and peroxisomes for the detoxification of ROS produced during excess light, drought, high salinity, and the exposure to Cd was reflected by large changes of ascorbate in these cell compartments observed during these stress situations (50, 59, 60, 61). A strong accumulation of ascorbate within 4 h could be observed in chloroplasts of Arabidopsis wildtype plants and the ascorbate-deficient mutant vtc2-1 exposed to excess light stress (50). In addition, the vtc2-1 mutant showed a 166% increase in ascorbate contents in peroxisomes than the wildtype when exposed to high light stress for 14 days (50). Despite lower ascorbate contents in all other cell compartments, the vtc2–1 mutant displayed similar or even lower sensitivity to high light stress, indicating an important role of ascorbate in peroxisomes for the protection against high light stress (50).
Another interesting observation was the appearance of ascorbate within the lumen of thylakoids of plants exposed to high light conditions (Fig. 3). Under these conditions, nonphotochemical quenching counteracts the formation of ROS by the dissipation of absorbed light as heat. Nonphotochemical quenching involves the formation of zeaxanthin to violaxanthin, which is catalyzed by violaxanthin de-epoxidase. This enzyme uses ASC as a reductant and is located inside the thylakoid lumen (43 –45, 85). Alternatively, ascorbate can also function as an alternative electron donor for photosystem II and I, which is important in situations of environmental stress when the linear electron transport is impaired (76, 116). Thus, the accumulation of ascorbate inside the thylakoid lumen during high light conditions and the increase of ascorbate contents inside chloroplasts and peroxisomes demonstrate the importance of ascorbate in the defense against excess light conditions (50).
A similar situation was found during high salinity where ascorbate contents accumulated in chloroplasts and peroxisomes and during drought stress where ascorbate contents remained unchanged in chloroplasts while dropping drastically (up to 81% and 65% in the cytosol during salt stress and drought, respectively) in most other cell compartments (59, 61). During the exposure of plants to 100 μM of Cd, ascorbate contents accumulated in chloroplasts and peroxisomes during the resistant phase at the expense of ascorbate contents of all other cell compartments (60). This response was especially pronounced in the ascorbate-deficient vtc2-1 mutant. In addition, the accumulation of ascorbate contents in chloroplasts during salt stress seemed to be an important defense strategy of the ascorbate- and glutathione-deficient mutants vtc2-1 and pad2-1, respectively (61). Ascorbate contents decreased or remained unchanged in most cell compartments of these mutants during salt stress, whereas ascorbate accumulated in chloroplasts throughout the experiment (61). Thus, it can be concluded that during abiotic stress conditions that primarily target chloroplasts, high ascorbate contents are maintained in this cell compartment at the expense of ascorbate from all other cell compartments. These results correlate well with the absence of ROS in chloroplasts during excess light and drought stress (50, 59), indicating that the primary job of ascorbate in chloroplasts is to counteract oxidative stress.
Interestingly, in all of these stress situations, ascorbate contents decreased in peroxisomes of Arabidopsis wildtype plants (50, 59, 60, 61). As ascorbate counteracts the accumulation of H2O2 formed inside peroxisomes due to photorespiration, these results indicate a diminished ability of ascorbate to counteract oxidative stress. But since ROS accumulated in the cytosol, peroxisomes, and vacuoles during these stress situations (50, 59), it seems that the diminished antioxidative capacity of the peroxisomes led to the diffusion of H2O2 into the cytosol and the vacuoles that represents an important pathway for its detoxification, as described later (Figs. 4, 5).

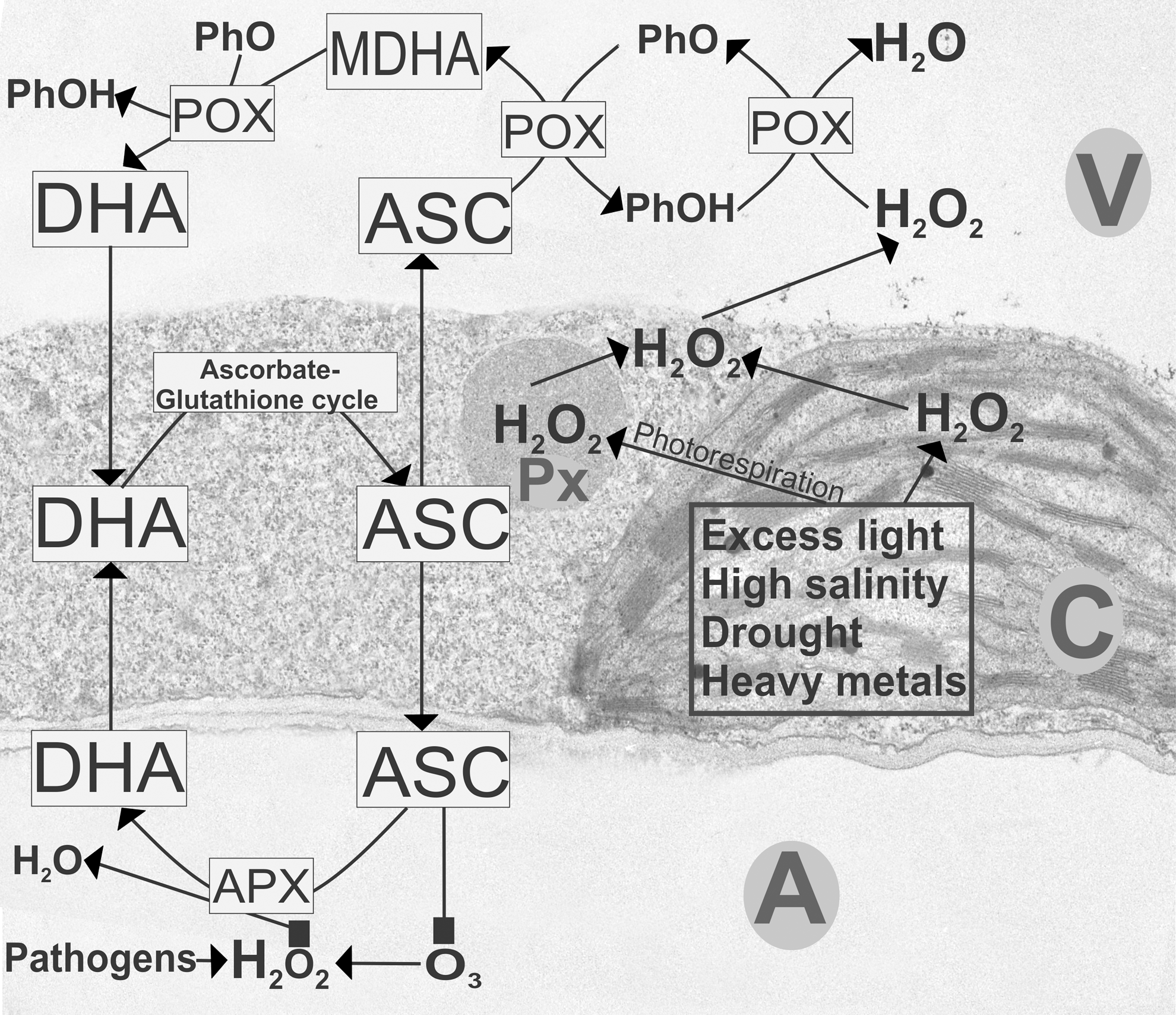
The roles of ascorbate in chloroplast are not only restricted to simply detoxifying ROS but also have important functions for signaling stress response through changes in its redox state. The treatment of Arabidopsis cell cultures with ascorbate and glutathione decreased the generation of ROS during heat treatment, which inhibited necrosis (29). On the other hand, it was found in this study that the treatment with ascorbate and glutathione increased apoptosis events such as programmed cell death, indicating that the interplay between ascorbate, glutathione, and ROS in chloroplasts is not only limited to the detoxification of ROS but also has important signaling functions in the activation of cell death (29). Similar conclusions were drawn from experiments with Arabidopsis flu mutants that contain a highly conserved protein called EXECUTER 1 inside plastids (91, 124). Changes of growth and development of this mutant were not caused by toxic effects of singlet oxygen that accumulated within the first minute of illumination in plastids but rather reflected the role of ROS as a signal initiator that activated several stress-response pathways (91, 105, 124).
The importance of ascorbate in chloroplasts and peroxisomes for plant defense against biotic stress situations is highlighted by several studies that report that the breakdown of the antioxidative system led to advanced symptom development during compatible fungal and bacterial infection (42, 67, 69, 70, 74, 94, 108). In these studies, decreased ascorbate contents and the accumulation of DHA, in chloroplasts and peroxisomes could be correlated with the accumulation of ROS in the tissue, progress of disease symptoms, and pathogen-induced leaf senescence. Ascorbate (and glutathione) contents in chloroplasts and peroxisomes were lower in the avirulent strain during Pseudomonas syringae infection of Arabidopsis than in the virulent strain (42). Ascorbate contents decreased by 70% within 12 h of infection in peroxisomes of plants inoculated with the avirulent strain whereas it remained unchanged in plants infected with the virulent strain. Avirulence is characterized by the activation of cell death during the hypersensitive response after recognition of the pathogen by the plant to restrict its proliferation (114, 115). Thus, these results confirm that ascorbate and other antioxidants in chloroplasts and peroxisomes play important roles in plant defense during abiotic and biotic stress situations.
Mitochondria
Ascorbate contents in mitochondria have been studied extensively during abiotic and biotic stress with biochemical methods and after immunogold labeling with quantitative TEM (42, 50, 59 –61, 69, 108). Antioxidants such as ascorbate play an essential role in mitochondria as they control ROS levels, which if not detoxified accumulate and induce programmed cell death (1, 122). Thus, it is not surprising that the development of necrosis could be correlated with a depletion of ascorbate (and glutathione) contents in mitochondria of Arabidopsis plants inoculated with the fungal pathogen Botrytis cinerea (108). A similar situation was observed in tomato plants infected with the same pathogen where mitochondria (after peroxisomes) showed the strongest drop of ascorbate (and glutathione) contents (70). In addition, a general shift of the redox balance toward the oxidative state could be observed in this study.
In Arabidopsis infected with the avirulent strain Pseudomonas syringae pv. tomato DC3000, an initial accumulation of ascorbate contents was followed by a steep decline that correlated with the development of cell death during the hypersensitive response (42). The drop of ascorbate in mitochondria could be directly correlated with the accumulation of ROS either in the whole leaf (42) or specifically in mitochondria (108). An accumulation of ROS in mitochondria can be correlated with the induction of cell death as ROS can lead to the damage of membrane components, resulting in the release of cytochrome C, which triggers apoptotic-like cell death (32, 101, 102).
Similar results were found when Arabidopsis plants were exposed to abiotic stress situations such as drought, Cd, and high salinity (59 –61). Salt stress decreased ascorbate contents in mitochondria (up to 67%) of wildtype plants and the ascorbate-deficient mutant vtc2-1 within 24 h and stayed within that range until the end of the experiment. Apart from this, glutathione contents decreased in mitochondria at the end of the experiment when strong symptoms and a decrease of biomass were induced in these plants (61). In addition, higher sensitivity of pad2-1 and vtc2-1 mutants to Cd treatment could be correlated with much lower ascorbate contents in mitochondria when compared with the wildtype (60).
A similar situation was found during drought stress where a depletion of ascorbate in mitochondria occurred at the very end of the experiment when chlorosis and necrosis appeared on the leaves (59). The authors concluded that, similar to the situation during biotic stress, the depletion of ascorbate (and glutathione) contents in mitochondria during abiotic stress such as drought, exposure to heavy metals, and high salinity lead to the accumulation of ROS in this cell compartment that could be responsible for the observed symptoms such as chlorosis and necrosis. Summing up, total ascorbate contents and its redox state in mitochondria are essential in controlling ROS during abiotic and biotic stress conditions. The depletion of ascorbate in mitochondria during such situations leads to the death of the plant.
Nuclei
It is well documented that the concentration and redox state of antioxidants and ROS in other cell compartments such as chloroplasts and mitochondria influence gene expression in the nucleus through retrograde signaling (57, 77, 78, 109). In addition, as most proteins in mitochondria and chloroplasts are encoded by genes in the nucleus, communication between these organelles and the nucleus regulates processes such as photosynthesis and respiration. Communication between organelles and the nucleus is, therefore, essential for proper plant growth, development, and defense. The importance of ROS and antioxidants such as ascorbate and glutathione as signaling agents and how they modify gene expression have been extensively reviewed in the past (57, 77, 78, 106) and will not be discussed here. However, little is known about the actual role of ascorbate in nuclei.
In nonstressed plants, concentrations of about 16 mM—much higher than in chloroplasts and mitochondria—have been determined in nuclei by correlating biochemical data with data obtained after cytohistochemical staining of ascorbate (131). Sufficient amounts of ascorbate play important regulatory roles during cell division and elongation. An increase of the ascorbate pool, for example, as well as increased amounts of MDHA promoted cell division (2, 25, 26, 51, 55, 72, 117), whereas depletion of ascorbate contents decreased cell division (110). The addition of DHA reduced the mitotic activity of the root meristem and inhibited cell proliferation of cells in onion (25, 92). Increased levels of reduced glutathione were not able to prevent the inhibition of cell division induced by DHA, indicating that the effects of ascorbate and glutathione on cell division are independent from each other (99). Thus, it seems that total contents of ascorbate and glutathione and their redox state play important regulatory functions for cell division and elongation (36), which becomes especially critical during environmental stress situations that result in oxidative stress, influence antioxidant levels, and shift the redox potential toward the oxidized state.
In addition, as an antioxidant, ascorbate in nuclei protects sensitive nuclear components (DNA, proteins etc.) from oxidation, which can otherwise cause mutation and result in cell death (27, 103). In this respect, it is interesting that the strongest decrease of ascorbate (58%) of all cell compartments was found in the nucleus within 12 h after infection of Arabidopsis wildtype plants with the fungal pathogen Botrytis cinerea (108). Although glutathione contents increased up to 300% in this cell compartment, a strong accumulation of ROS could be observed in the nucleus, which eventually resulted in ultrastructural alteration and cell death (108). A similar situation was found during the infection of Arabidopsis plants with the bacterial pathogen Pseudomonas syringae. An initial increase of ascorbate contents in nuclei was followed by a steep drop that correlated with the accumulation of ROS and symptom development such as chlorosis and necrosis (42). Thus, during biotic stress situations, there seems to be a direct correlation between the drop of ascorbate contents and the accumulation of ROS in nuclei, which results in ultrastructural alterations and, eventually, cell death.
Such a situation could not be clearly defined during abiotic stress conditions as ascorbate contents remained largely unchanged in nuclei of Arabidopsis Col-0 wildtype plants during salt stress, high light stress, and the exposure to Cd or decreased at the end of a drought stress experiment (50, 59 –61). Nevertheless, it is important to note here that during some stress conditions (salt stress, exposure to Cd, and, at some time points, during drought stress) ascorbate contents accumulated in nuclei relatively to the cytosol, which was depleted of ascorbate (59 –61). In addition, ascorbate contents strongly increased during these stress situations in the nuclei of the ascorbate-deficient vtc2-1 mutant or remained unchanged in nuclei during drought stress whereas ascorbate contents dropped in the wildtype. Thus, the results obtained during abiotic and biotic stress conditions indicate that the accumulation (relatively to the cytosol or absolute) of ascorbate in nuclei is an important defense mechanism against environmental stress situations to regulate cell division and elongation under these circumstances.
Vacuoles
Ascorbate could be detected in isolated vacuoles with biochemical methods (111) and with TEM after immuno-cytohistochemistry staining with gold (131). Among all cell compartments, vacuoles contained the lowest concentrations (2 mM) of ascorbate (Fig. 1). Nevertheless, a strong increase of ascorbate in vacuoles could be observed in Arabidopsis plants exposed to high light, drought, and after the inoculation with Pseudomonas syringae (42, 50, 59). Pseudomonas syringae infection induced a twofold increase in ascorbate contents in vacuoles just 12 h after the inoculation when symptoms were not visible yet on the leaves (42). In wildtype plants exposed to high light conditions of 1500 μmol m−2 s−1 an increase of ascorbate in vacuoles of 130% could be observed within 4 h of light treatment. After 14 days of high light treatment, 236% more ascorbate could be detected in vacuoles of these plants when compared with control conditions. Only chloroplasts showed an increase in ascorbate contents at these time points, whereas ascorbate was decreased in all other cell compartments (50).
In drought-stressed plants, vacuoles were the only cell compartment that showed increased levels of ascorbate (up to twofold) throughout the experiment that lasted 10 days. In addition, low ascorbate contents in vacuoles of the ascorbate-deficient mutant vtc2-1 correlated with higher sensitivity to drought (59). A general increase of O2 − and H2O2 was detected in leaves during Pseudomonas syringae infection, whereas the accumulation of H2O2 during drought and excess light stress could be specifically localized in vacuoles (50, 59). From these results, it was concluded that H2O2 produced in chloroplasts and peroxisomes leaks into vacuoles during high light and drought stress where it gets detoxified by phenols (111, 112).
The accumulation of ascorbate contents in vacuoles during these conditions can be explained by the observation that ASC in vacuoles helps to reduce phenoxyl radicals that form during these reactions. The resulting MDHA and DHA are then transported into the cytosol for reduction to ASC, which can then be imported again into vacuoles (111, 112, Fig. 5). These results demonstrate that the detoxification of H2O2 in vacuoles through ascorbate is an important mechanism in plant defense against environmental stress situations that can even contribute to higher tolerance against certain abiotic stress situations such as drought.
Apoplast
Although ascorbate could be detected in the apoplast of plants with biochemical methods (63, 118 –121), immune-electron microscopy failed to detect ascorbate in the apoplast of mesophyll cells with TEM (131). Even under severe environmental stress conditions such as the exposure to excess light, drought, Cd, and the infection with fungal or bacterial pathogens, ascorbate could not be detected in the apoplast of mesophyll cells with the latter method (42, 50, 108, 59, 60). Thus, information about changes of ascorbate contents in the apoplast in plants exposed to environmental stress is only available from biochemical methods after extraction of soluble apoplastic components (75, 118 –121). Under nonstressed conditions, ascorbate occurs in the apoplast either entirely or in up to 50% in its oxidized form (89, 118 –120). Total ascorbate contents in the apoplast were found to be about 4 mM whereas glutathione contents were either not present (e.g., oat plants) or very low (6–7 μM; 120), making ascorbate the predominant antioxidant in this cell compartment.
A decrease of DHA contents was frequently observed in the apoplast of resistant and susceptible barley and oat plants within 24 h after the infection with powdery mildew (118 –120). As the resistant strain showed a stronger decrease of DHA contents in the apoplast but unchanged ASC contents, the authors concluded that a more rapid import of DHA into the cytosol for reduction to ASC occurs in these plants (Fig. 5, 120).
The importance of ascorbate in the apoplast in plant growth, development, and defense was also confirmed during the infection of tobacco plants with the bacterial pathogen Pseudomonas syringae, and in Arabidopsis plants during drought stress (73, 128). High activity of AO in the apoplast induced the depletion of ASC and the accumulation of DHA, which mimicked the effects of ROS accumulation in this cell compartment. AO-catalyzed changes in ascorbate contents and its redox state in the apoplast could also be linked to changes in the activity of antioxidant enzymes, modifications in plant growth through changes of hormone synthesis and MAPK activity, and changes in calcium signaling (73, 123). Thus, the authors concluded that AO-catalyzed changes in ascorbate contents in the apoplast play essential roles in redox signaling that influence plant, growth, and defense in response to environmental stimuli (73).
An important role has also been assigned to ascorbate in the apoplast in the protection against O3 (4, 23, 30, 104). O3 oxidizes a variety of cell components such as double bounds of fatty acids, amino acids, sulfhydryl groups, and other antioxidants such as tocopherol (54). In contrast to other ROS, O3 is not created within the cell but enters the plant through stomata. The first reaction sites of O3 are, therefore, the apoplast and the plasmalemma, where ascorbate seems to be the most important agent to detoxify O3. The remaining O3 degrades to ROS, such as H2O2, superoxide anion radical, and hydroxyl radical after it dissolves in the apoplastic fluid or after entering the cytosol (Figs. 4 and 5). Ascorbate and other antioxidants detoxify O3 and ROS during such situations, which results in commonly observed changes in antioxidant levels, their redox status, and an increase in their biosynthesis during the exposure of plants to O3 (4, 23, 30, 104).
Several studies have found a strong increase in apoplastic ascorbate contents after the treatment of plants with O3 (19, 93, 121, 125). As the redox state of the apoplast became more reduced in poplar leaves and due to a general increase and an unchanged redox state of ascorbate inside the cells of poplar and winter wheat, the authors concluded that the successful stress response of the apoplast against O3 relies on both the export of ASC from the cytosol into the apoplast, where it forms DHA after reacting with O3, and the import of DHA into the cytosol, where it is reduced to ASC by DHAR (93, 121, 125). These results are supported by the observation that the Arabidopsis mutant dhar that lacks cytosolic DHAR showed higher O3 sensitivity and a 61.5% lower ascorbate pool in the apoplast (129). As the cytosolic DHAR was found to be localized in the plasma membrane and as DHA levels in dhar were twice as high as in the wildtype, the authors came to the conclusion that DHA formed during O3 treatment in the apoplast could not be efficiently detoxified by DHAR in the cytosol. Thus, the lack of a recycling system of apoplastic ASC (in the cytosol) leads to higher O3 sensitivity in the dhar mutant (129).
The importance of apoplastic ascorbate in plant defense, growth, and development has further been dissected with transgenic tobacco plants with different ascorbate contents and redox state in the apoplast (96, 97, 128). In these studies, the authors were able to demonstrate that decreased ascorbate contents of the apoplast led to ROS accumulation independent from ascorbate levels in the cytoplasm. In addition, low ascorbate contents in the apoplast decreased plant resistance whereas low contents inside the cell increased plant resistance to pathogens through auxin-mediated signal transduction (96, 97). Thus, the authors concluded that ascorbate contents and the redox state in the apoplast are of crucial importance in controlling signal transduction processes during environmental stress conditions that are essential for plant defense, growth, and development.
Summing up, ascorbate plays important roles in the protection of plants against external stress, such as exposure to O3 and pathogen attack that affect the apoplast first. There is common agreement that the main supply of ASC necessary to detoxify ROS in the apoplast comes from the cytosol and that DHA formed in the apoplast gets transported across the plasmalemma for reduction inside the cell (Fig. 5). Changes of ascorbate contents and its redox state were commonly found in the apoplast during O3 treatment and pathogen attack, whereas modifications of ascorbate could not always have been directly related to modification of plant defense (e.g., higher tolerance). Thus, the roles of ascorbate in the apoplast are not simply restricted to the detoxification of ROS but also involve signaling functions of external environmental stimuli to modulate plant defense, growth, and development.
Cytosol
Ascorbate contents in the cytosol (22 mM) have been determined to be among the highest within plant cells (Fig. 1; 131). These concentrations are more than double as high as those observed in mitochondria, which are exclusively responsible for ascorbate synthesis (7, 14, 80, 107, 126). This demonstrates that large amounts of ascorbate are exported from mitochondria into the cytosol and that the cytosol acts as a sink for ascorbate. Due to the essential roles of ascorbate in redox signaling, modulation of gene expression, and the regulation of enzymatic activities (33, 34, 36), the cytosol can, therefore, be considered the hub of ascorbate metabolism in plants that controls plant development, growth, and defense.
On the one side, the cytosol contains all components of the ascorbate-glutathione cycle, which enables it to directly detoxify ROS that diffuse from other cell compartments into the cytosol (Fig. 4). On the other side, it controls the redistribution of ascorbate to all other cell compartments through selective ascorbate transporters (16). It also represents the main hub for the reduction of DHA imported from the apoplast and the vacuoles (112, 118 –120). Once reduced in the cytosol, ASC gets exported into the apoplast and the vacuole where it is used to detoxify ROS (Fig. 5). In this respect, the cytosol is able to directly counteract ROS accumulation and it also acts as a reservoir and provider of ASC for other cell compartments. As the cytosol, besides peroxisomes, was found to act as a sink for ascorbate in nonstressed plants (131) a redistribution of ascorbate during environmental stress situations seems to be a key factor for successful plant defense.
During several environmental stress situations such as drought, exposure to high salinity, fungal pathogen attack, heavy metal treatment, and short- and long-term light stress, a strong depletion of ascorbate contents in the cytosol could be observed whereas other organelles showed increased levels of ascorbate (50, 59 –61, 108). This was especially evident during salt stress, drought, and long-term high light stress where ascorbate contents in the chloroplasts, which usually only contain about half as much ascorbate than the cytosol, reached values similar or higher than what were found in the cytosol (50, 59 –61). Thus, it can be concluded that under these stress situations, ascorbate is actively imported into and accumulates in chloroplasts.
Interestingly, changes in ascorbate contents observed in the cytosol mimicked changes observed in mitochondria during most of the earlier described environmental stress situations such as drought, high salinity, exposure to heavy metals, and high light (50, 59 –61), indicating that levels of ascorbate in the cytosol strongly depend on the supply of ascorbate from the mitochondria during environmental stress situations. As ascorbate contents in the cytosol are also influenced by ascorbate levels and its redox state in the apoplast, the cytosol can be seen as a central hub where signals from the outside in terms of changes in ROS, ascorbate levels, and redox state are sensed and processed. As protein synthesis and post-translational modification of proteins take place in the cytosol and are regulated by ascorbate contents and its redox state (9, 12, 83, 90), such changes will then result in a coordinated response and physiological modification of the whole cell.
Within the cytosol, ascorbate could not be found to be associated with dictyosomes but could be detected along the membranes of the ER but not inside the lumen. In mammalian and vertebrate tissue, ascorbate levels and their redox state inside the lumen play important roles for antioxidative defense, in regulating protein folding, and regulating the activity of enzymes by maintaining the redox state of their active sites (reviewed by Ref. 5). As both forms of ascorbate are needed to perform these functions, the transport of ASC and DHA across the ER membrane has been proposed in these cells. The roles of ascorbate in the ER of plants are not well investigated. As described earlier, ascorbate has only been detected at the membrane of the ER but not inside the lumen in nonstressed plants (131). Nevertheless, ascorbate transporters (SINAT) have been predicted by sequence analysis in the membrane of the ER in tomato plants, indicating that ASC (and DHA) could be transported into the lumen of the ER of plants where it could fulfill similar roles as described earlier for animal tissues (16). Further studies about their functional importance in plants and distribution in other plants besides tomato would help to clarify the role of ascorbate in the ER of plants.
It can be concluded that ascorbate in the cytosol fulfills important functions in protecting plants against ROS, and also in sensing and signaling oxidative stress. On the one side, ascorbate levels and their redox state in the cytosol are influenced by ROS that cannot be detoxified at its origin and diffuse into the cytosol. On the other side, they are influenced by DHA that is transported into the cytosol from the apoplast and vacuoles during the detoxification of ROS in these cell compartments. The cytosol reacts to these situations by modifying the transport of ASC to the individual cell compartments and by modifying proteins that can lead to changes in plant growth, development, and defense.
Conclusion and Perspective
Based on compartment-specific changes of ascorbate contents and its redox state during abiotic and biotic stress situations in plants, a clear picture can now be painted about the importance of ascorbate for the individual cell compartments and for the cell as a whole for plant defense. The accumulation of ascorbate contents in chloroplasts counteracts ROS produced during abiotic stress situations that primarily target chloroplasts such as excess light conditions, drought, high salinity, and the exposure to heavy metals (Fig. 4). The depletion of ascorbate contents in peroxisomes during abiotic stress conditions and the complete breakdown of the antioxidative system in both chloroplasts and peroxisomes during biotic stress situations lead to the development of chlorosis and necrosis.
A similar situation was found for mitochondria, where the depletion of ascorbate contents was linked with the accumulation of ROS and the development of chlorosis and necrosis during biotic and abiotic stress. There is increasing evidence that the role of ascorbate in chloroplasts and mitochondria is not only limited to control the accumulation of ROS but also that the redox status of ascorbate and ROS in these cell compartments is used as a signal to modify gene expression in the nuclei (retrograde signaling) to regulate plant defense, growth, and development.
Interestingly, vacuoles were identified as the main hotspot for H2O2 as well as ascorbate accumulation during most stress situations. These results indicate that H2O2 produced in chloroplasts and peroxisomes diffuses into vacuoles where it gets detoxified by ascorbate (Fig. 5). A similar situation was found for the apoplast where ascorbate was found to protect plants against ROS produced during stress situations such as fungal and bacterial infection and O3 treatment. In addition, as ascorbate contents in the apoplast could not always be directly related to plant defense, it seems that the roles of ascorbate and its redox state in the apoplast are restricted not simply to the detoxification of ROS but also to signal external environmental stimuli into the cell.
For both vacuoles and the apoplast, it was proposed that ASC gets transferred into these compartments from the cytosol whereas DHA, produced as a consequence of ROS detoxification, gets transported back into the cytosol for reduction. This is especially important during environmental stress situations when ASC is used in the apoplast and vacuoles to counteract oxidative stress (Fig. 5). In this respect, the cytosol can be considered a hub for ascorbate metabolism, which acts as a sink for ascorbate produced in mitochondria and selectively distributes ascorbate to the individual cell compartments as needed. In addition, ascorbate detoxifies ROS that diffuse into the cytosol from other cell compartments. As protein synthesis and modifications take place in the cytosol and as ascorbate contents and its redox state are directly involved in these processes, it is obvious that ascorbate senses and signals oxidative stress, which then regulates plant growth, development, and defense.
To achieve further insight into the compartment-specific roles of ascorbate for plant growth, development, and defense during abiotic and biotic stress situations in plants, future research should focus on: the dissection of the roles of ascorbate in vacuoles during abiotic and biotic stress situations, which should help to clarify whether ascorbate simply detoxifies ROS in this cell compartment or whether changes of its redox state in vacuoles can also be related to signaling; the clarification as to whether ascorbate also occurs in the lumen of the ER in plants and whether it has similar roles as have been proposed for mammalian and vertebrate tissues such as antioxidative defense, regulation of protein folding, and regulation of the activity of enzymes; the identification and dissection of the physiological relevance of ascorbate transporters located in the membranes of the different cell compartments and to what extent they are responsible for the import and export of ascorbate from cells, tissues, and organs during environmental stress situations; the combination and progression of current techniques that would enable the measurement of actual ascorbate concentrations and their redox state in the individual cell compartments; correlating available data on changes of ascorbate contents with changes (i) in the subcellular distribution of related enzymes of the ascorbate-glutathione cycle, (ii) in the compartment-specific distribution of ROS, (iii) in gene expression patterns of related genes, especially the ones encoding enzymes involved in the ascorbate-glutathione cycle, and (iv) in the proteome.