
Abstract
Significance:
The mitochondrial fraction of
Recent Advances:
Most cell types take up AA by the high-affinity sodium-dependent vitamin C transporter 2 (SVCT2) sensitive to inhibition by dehydroascorbic acid (DHA). DHA can also be taken up by glucose transporters (GLUTs) and then reduced back to AA. DHA concentrations, normally very low in biological fluids, may only become significant next to superoxide-releasing cells. Very little is known about the mechanisms mediating the mitochondrial transport of the vitamin.
Critical Issues:
Information on AA transport is largely derived from studies using cultured cells and is therefore conditioned by possible cell culture effects as overexpression of SVCT2 in the plasma membrane and mitochondria. Mitochondrial SVCT2 is susceptible to inhibition by DHA and transports AA with a low affinity as a consequence of the restrictive ionic conditions. In some cells, however, high-affinity mitochondrial transport of AA is observed. Mitochondrial uptake of DHA may take place through GLUTs, an event followed by its prompt reduction to AA in the matrix. Intracellular levels of DHA are, however, normally very low.
Future Directions:
We need to establish, or rule out, the role and significance of mitochondrial SVCT2 in vivo. The key question for mitochondrial DHA transport is instead related to its very low intracellular concentrations.
Introduction
A
This statement is based on the simple consideration that maintenance and growth of cells in culture are, under basal conditions, already associated with oxidative stress since the oxygen tension is about 10 times higher than the one experienced by the cells in vivo. Under these nonphysiological conditions, basal rates of mitochondrial superoxide anion (O2 −•) formation are significantly increased by the enhanced availability of molecular oxygen (19, 39, 40), with the ensuing triggering of prosurvival responses (5, 115). The overall scenario is then further complicated by remarkable differences of these adaptive responses, often based on the cell type, particularly when associated with the genomic instability of cultured tumor cells.
Among other considerations, not reported here for the sake of brevity, a final one refers to the composition of conventional growth media used to keep cells in culture, which on the one hand, often contain ingredients promoting oxidative stress (39
–41), but on the other hand, may not contain all essential components of antioxidant defense. Good examples are represented by vitamin E and vitamin C (
AA, a water-soluble vitamin that poorly crosses biological membranes, is normally taken up by the cells through specific transporters and it would make a lot of sense if these cells compensate for the deficiency in vitamin C with the overexpression of these transporters, in particular those with high affinity. As will be discussed later on, this is indeed what happens and the overexpression/increased activity of the transporters (73, 98, 101) introduces an additional complicating factor for the correct interpretation of results obtained in cells supplemented with AA. In addition, we should also consider that similar issues are of potential relevance when considering the distribution of AA in specific subcellular compartments. Among these, mitochondria are expected to benefit by the antioxidant function of the vitamin as these organelles produce large amounts of reactive oxygen species (ROS).
These considerations imply that the signaling revolution imposed by the cell culture conditions should always be kept in mind when translating the outcomes of cell culture experiments on oxidative stress, and/or on the effect of specific components of the antioxidant defense, to biological concepts relevant for living organisms. At the same time, however, we have learned a lot on these issues and we could possibly learn more if the limitations and flaws of the cell culture approach are correctly considered during the design of the experiments and interpretation of their results.
This review represents an attempt to critically interpret the information of the literature on the controversial issue related to AA uptake in mitochondria, in which most of the information available is largely based on results obtained in cultured cells.
Cellular Accumulation of Vitamin C
Vitamin C is synthesized in the liver of a very large majority of mammals— information that is consistent with its relevance in various physiological processes. Many comprehensive reviews published on this issue (11, 22, 25, 58, 61, 88) describe very well the importance of vitamin C not only as an antioxidant but also for its role in the regulation of critical cellular functions (e.g., syntheses of collagen, catecholamines, carnitine, cholesterol, amino acids, and certain peptide hormones and vitamin E recycling).
Other species, such as humans and other primates, as well as guinea pigs, have, however, lost their capacity to synthesize vitamin C since they fail to express functional
Scurvy is, however, rarely observed and may only take place under conditions in which plasma concentrations of the vitamin are kept below 10–11 μM (108) for a prolonged period of time. Mild vitamin C deficiency is instead frequently observed in humans. In particular, deficiencies are found in as many as 10% of adults in the industrialized world (102, 106, 113), with subgroups (e.g., smokers and families with a low socioeconomic status) displaying an even higher prevalence (31, 66, 68, 121).
An increased incidence of adverse effects associated with mild vitamin C deficiency is therefore expected in a very large population of Western countries, with potential long-term consequences associated with increased susceptibility to a variety of diseases (67). Hypoascorbemia shares several clinical symptoms with diabetes mellitus such as hypercholesterolemia, atherosclerosis, microangiopathy, capillary hyperperfusion, and hemorrhages (70). These events, blunted by vitamin C supplementation (49), are likely associated with the structural similarities of vitamin C with glucose. Indeed, the latent scurvy theory (90) postulates that hyperglycemia leads to competitive inhibition of AA uptake, thereby inducing intracellular vitamin C deficiency.
Dietary AA is readily absorbed in the gastrointestinal tract, circulates in the blood in a free form, is filtered, and may then be reabsorbed in the kidney tubule system (13).
Most of the biological effects mediated by vitamin C occur within the cells in which its concentrations are remarkably higher (often >1 mM) than those found in the plasma and other biological fluids, normally between 50 and 100 μM (59, 63), with some exception, as in the cerebrovascular fluid, in which the concentrations can reach up to 200–400 μM (96, 97). The comprehension of the mechanism(s) whereby cells regulate the uptake and accumulation of vitamin C is therefore of pivotal importance for correct interpretation of the overall effects mediated by the vitamin in vivo.
Cellular uptake of the reduced and oxidized forms of vitamin C
AA, as a consequence of its hydrophilicity, poorly penetrates plasma membranes by simple diffusion and is therefore directly taken up by most cell types via two different transporters, sodium-dependent vitamin C transporter 1 and 2 (SVCT1 and SVCT2) (18, 69, 74, 101, 114, 120, 122). SVTC2 (Fig. 1), detected in the majority of cell types of various organisms, has a high affinity (Km ∼10–20 μmol/L), and for optimal activity, requires high concentrations of Na+, Ca2+, and Mg2+ normally found in the extracellular milieu (35). The physiological importance of SVCT2 is emphasized by the observation that SVCT2 knockout mice die soon after birth (109).

SVCT1 (Fig. 1) has a more limited distribution than SVCT2 and is responsible for AA uptake into epithelial cells of the small intestine, liver, and kidney (9, 56, 65, 73, 118, 122). In addition, SVCT1 is also expressed in some epithelia of the reproductive system and brain (114). SVCT1 transports AA with a lower affinity (Km ∼100–200 μmol/L) and with a greater capacity than SVCT2, both of which are important characteristics that make it functional for intestinal and renal absorption of the vitamin. Targeted deletion of SVCT1 resulted in accelerated renal excretion and low plasma concentrations of the vitamin, as well as in ∼50% perinatal mortality (13, 109, 126). These findings emphasize the relevance of SVCT1 and, in particular, its pivotal role in the intestinal absorption and renal reabsorption of AA.
Cells employ additional strategies for vitamin C uptake and, more specifically, may take up the oxidized form of the vitamin, dehydroascorbic acid (DHA), which, as previously noted for AA, also poorly penetrates the plasma membrane by passive diffusion (63). DHA uptake is mediated by facilitative hexose transport, with the involvement of numerous glucose transporters (GLUTs) (14, 88, 99, 122). DHA uptake is followed by its rapid intracellular reduction back to AA via NADH, NADPH, and reduced glutathione (GSH)-dependent reactions (14, 55, 64, 123). Hence, the mechanism of vitamin accumulation based on DHA transport appears advantageous because of its high capacity and rapid intracellular recycling, which in concert, allow rapid accumulation of large amounts of the vitamin.
It is, however, important to keep in mind that the use of this strategy is hampered by extracellular levels of DHA, generally below 2 μM (88), with some variation associated with the release of AA-oxidizing species [e.g., O2 −•, (85, 86)], as might occur in inflamed tissues. An additional problem may arise from the possible competition for DHA transport through GLUTs by glucose itself, normally present in millimolar concentrations, which, however, is not always observed. DHA transport may not even be affected by physiological glucose concentrations, as in the case of human erythrocytes (78).
There has been much debate on the specific relevance of the Na+-dependent transport of AA versus the facilitative diffusion of DHA, and the many experiments performed in cultured cells over the years have provided relevant information for the comprehension of the overall phenomenon of vitamin C transport in various tissues (25, 63, 88, 109). On the other hand, some conflicting information has been produced when comparative studies were performed using conditions favoring an uptake system versus the other.
A typical example, as we recently discussed (4), is represented by the comparison of the amounts of vitamin C accumulated after exposure to increasing concentrations of AA and DHA. The concentrations of the two forms of the vitamin employed in some of these studies were very high, often 1 mM, that is, remarkably greater than those found in physiological conditions. Furthermore, even using 10 times lower concentrations, the comparison would nevertheless remain poorly physiological since 100 μM is still very high for DHA; 100 μM AA is instead closer to the concentrations of the vitamin detected in biological fluids, but it may actually be too elevated under conditions in which high-affinity transporters with a Km close to 10 μM are overexpressed as a consequence of cell growth in the absence of the vitamin in the culture medium (101).
What is normally observed in these experiments is that exposure to high concentrations of DHA leads to an enormous accumulation of vitamin C as a consequence of the high transport capacity, with an uptake of the reduced form of the vitamin that (in comparative terms) appears negligible since it is characterized by a low transport capacity. Clearly, these results are potentially misleading and should not be taken as an indication that in the specific cell type under investigation, the DHA uptake mechanism is preferential over the Na+-dependent one. The conditions selected are obviously ideal for the high-capacity transport of DHA, with very little advantage, if any, for the high-affinity/low-capacity transport through SVCTs.
Comparative studies performed with low concentrations of AA should instead result in significant vitamin C accumulation in SVCT2-expressing cells, with very little vitamin accumulation after treatment with the same low concentrations of DHA. We should also consider that the high concentrations of AA might be toxic for the cells since these conditions are associated with an extensive formation of ROS generated by AA auto-oxidation processes (77, 105). Uptake studies should not be performed under these conditions since active transport of the vitamin is likely hampered by the insult received by the cells. Furthermore, the process of AA autoxidation results in DHA formation, which might then be taken up by the cells, thereby complicating the interpretation of the experimental results. When using high AA concentrations, one should always use a strategy to keep extracellular vitamin C in its reduced state.
The process of AA autoxidation is a typical cell culture effect (77, 105) mediated by the elevated oxygen tension and by the presence of transition metals in the culture media (39, 40), normally not observed in the plasma or other biological fluids because of the low oxygen tension and the poor availability of free transition metals. In other words, vitamin C, at physiological levels, fails to produce a pro-oxidant effect in biological fluids for the same reasons that vitamin C is not a pro-oxidant inside the cells. Things are obviously different when very high concentrations of vitamin C are injected intravenously in an organism, either alone or with other agents, with the purpose of killing cancer cells (23, 32, 91). Under these conditions, the extensive ROS release is critically associated with the antitumor effects of the vitamin alone or in combination therapy.
The cell culture approach should therefore be carefully employed for the purpose of evaluating the relevance of AA versus DHA transport in a specific cell type, especially when using high concentrations of the vitamin. Under these conditions, the remarkably greater accumulation of the vitamin should not lead to the conclusion that the biological relevance of DHA transport through GLUTs is greater than that of AA through SVCTs. In this direction, the evidence that SVCT2 knockout mice present very little AA in their brain and other tissues (109, 126) provides a straightforward indication of the biological relevance of this transporter. Hence, SVCT-2-mediated AA transport in vivo is of critical importance and cannot be compensated by DHA transport through GLUTs.
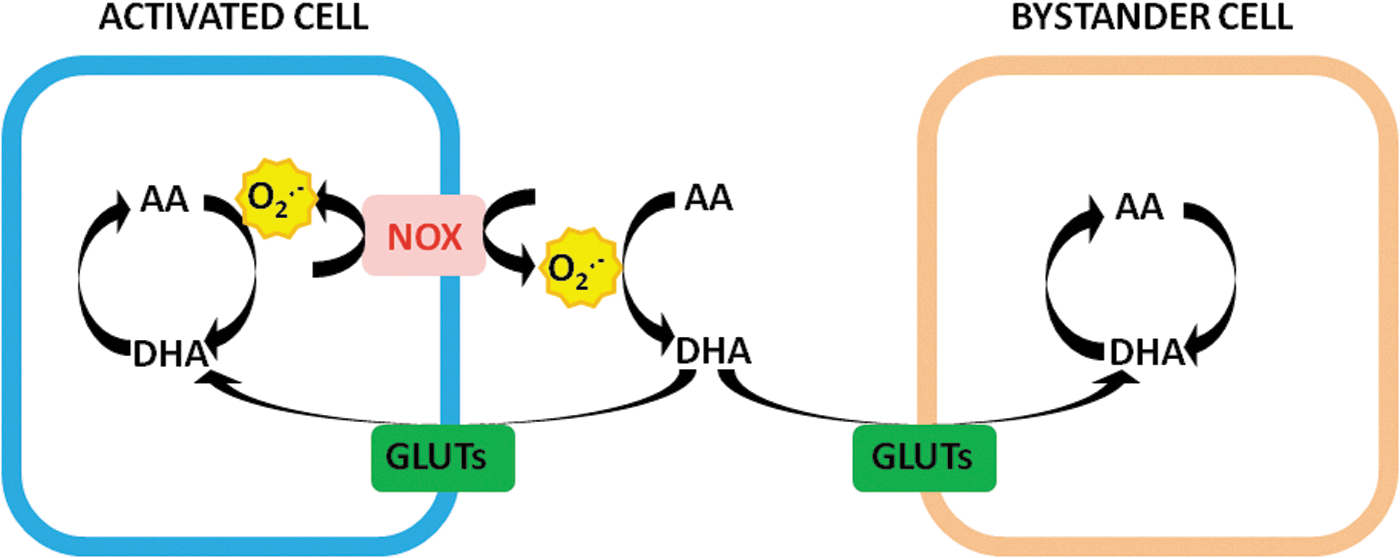
There is, however, a potential biological significance of this second transport mechanism. As mentioned above, normal DHA plasma levels are in the very low micromolar range, although some increase is expected to take place under inflammatory conditions (85, 86). Many investigators suggest that activated inflammatory cells promote the release of O2 −• and the ensuing AA oxidation, thereby enhancing the pericellular concentration of DHA and hence the possibility of its uptake through GLUTs. Furthermore, this event may take place not only in the O2 −•-releasing cells but also in the bystander cells. The cell culture approach has provided abundant information in this direction (26, 85, 86). Using different cellular systems, it was indeed observed that activation is invariably associated with an enhanced cellular accumulation of vitamin C (26, 85, 86).
While convincing, within the limits of the experimental system in which they were obtained, these results should be considered suggestive of a contribution of the bystander effect in vivo, although at least to our best knowledge, this hypothesis has not as yet received a definitive and univocal demonstration. This mechanism (Fig. 2) may fulfill the requirements for a greater availability of vitamin C under the same conditions in which it is being oxidized.
Mitochondria and Vitamin C
We previously mentioned that vitamin C is critical for an array of physiological functions and that its insufficient intake in organisms unable to produce it leads to scurvy. The well-known ability of the vitamin to mediate effects in different subcellular compartments raises the problem of the mechanism of vitamin transport in mitochondria and other organelles. At the same time, we should also be aware of the fact that scurvy may be relevant both at the cellular and subcellular levels and eventually plays a role in different pathologies (6, 70). This important issue has been addressed in several studies and the general idea is that the vitamin uptake is largely based on DHA transport. Hexose transporters were indeed detected in the Golgi apparatus (6, 82), lysosomes (2, 6, 20), nuclear envelop (6, 15, 70), and endoplasmic reticulum (6, 70, 83, 104).
Recent reviews provide detailed information on this important topic (6, 70). We would like to focus our attention on mitochondria, in which the process of vitamin C accumulation is expected be particularly important.
Mitochondria are subcellular organelles playing a critical role in a variety of functions, which include energy biogenesis by means of oxidative phosphorylation (8, 81, 117), regulation of Ca2+ homeostasis (21, 92), and cell signaling responses affecting various physiological processes (21, 92) as well as autophagic (16, 17, 54, 124) and apoptotic pathways (17, 53, 54, 94).
Mitochondria also represent the main intracellular source of ROS, in particular O2 −•, a partially reduced form of molecular oxygen generated by a one-electron reduction reaction (1, 42, 80). O2 −• formation takes place as a consequence of the electron leak from the mitochondrial respiratory chain, in particular at the level of complexes I and III (7, 12, 24, 42, 43). Although the specific contribution of each of these events is potentially affected by specific characteristics of the cell type, in particular its metabolic activity, complex III is in general more effective in mediating O2 −• formation than complex I.
Under physiological conditions, a significant fraction of molecular oxygen (about 1%, probably less) is converted to O2 −• and roughly 75% of this fraction derives from the ubiquinone cycle during electron transfer to cytochrome c in complex III (42). Importantly, O2 −• produced at the level of complex I is released in the matrix, whereas the bifacial localization of complex III leads to O2 −• release in the same compartment as well as in the intermembrane space (1, 53).
Once produced, O2 −• may rapidly react with nitric oxide, when available, to generate peroxynitrite (62, 87, 110), a highly reactive and toxic nitrogen species (87, 110). Protonation of O2 −•, with the ensuing formation of hydroperoxyl radicals, is also a mechanism potentially resulting in the induction of deleterious effects on various biomolecules (42, 53). Finally, and most commonly, O2 −• is converted to hydrogen peroxide (H2O2) either spontaneously or enzymatically (24, 43, 53), both inside and outside the mitochondria, thereby allowing its interaction in different subcellular compartments with transition metals, typically Fe2+, to generate hydroxyl radical species (1, 24, 53).
These species are highly reactive and may damage various biomolecules, in particular lipids, proteins, and DNA (24, 43, 53, 110). Under physiological conditions, mitochondrial ROS instead participate in the complex machinery regulating cell signaling (1, 42, 43, 46), and this is an important reason, among many others, explaining the lack of therapeutic effect or even the increased incidence of adverse effects demonstrated by various antioxidants in clinical studies (34).
Mitochondria are therefore continually exposed to a flux of O2 −•/H2O2 derived from the respiratory chain and other mitochondrial sources not reported here for the sake of brevity (24, 42, 43), with both physiological and pathological/toxicological implications, thereby implying the existence of an organized and efficient mitochondrial antioxidant defense. Mitochondria indeed contain a network of antioxidant enzymes such as glutathione (GSH) peroxidase, oxidized glutathione reductase, manganese-containing superoxide dismutase, catalase, and peroxiredoxins (24, 43, 50, 51, 53, 95, 100). The major nonenzymatic antioxidants are GSH, coenzyme A, ubiquinol, and vitamin E (24, 43, 70 –72, 95). Vitamin C is also expected to be beneficial for the mitochondria under both physiological conditions and during extensive ROS release.
Mitochondrial accumulation of vitamin C
Cultured cells cope very well with the elevated oxygen tension and the enhanced ROS release and indeed survive/proliferate even in the absence of vitamin C. We should therefore recognize that mitochondrial function and integrity are also well preserved under the same conditions. This would imply that vitamin C is not essential for these cells that can develop alternative/compensatory strategies allowing their survival even under harsh culture conditions. As previously discussed, however, overwhelming information derived from clinical or animal studies demonstrates a severe impact of insufficient AA intake, or cellular uptake, in living organisms (63, 70, 88, 126).
Seen in this perspective, there are very good reasons to believe that mitochondrial AA is also critical for the function and integrity of these organelles in vivo.
The available information on vitamin C uptake and accumulation in animal tissue mitochondria is surprisingly limited and some of these studies are about 35 years old. Vitamin C has been detected in rat liver mitochondria and the experimental results were in the range 0.5–1 mM (33, 60). Results compatible with these values were obtained in our laboratory, however, using mouse liver mitochondria (data not shown). Liver mitochondria isolated from guinea pig displayed an AA content three times greater than the above (47). Remarkably higher values (about 50 times) were instead reported by Ramanathan et al. (93) for rat liver mitochondria.
AA was also detected in rat brain mitochondria (119) at levels comparable with those found by most studies in liver mitochondria. Much less AA (about five times) was detected in the mitochondria from the mouse skeletal muscle (60) and these results are in keeping with our preliminary findings obtained with the mouse tibialis anterior muscle (103). We are not aware of human studies documenting the presence of vitamin C in the mitochondria.
In summary, high levels of vitamin C can be found in the mitochondria isolated from various animal tissues. We previously mentioned that the hydrophilic nature of both AA and DHA is not permissive for their passive diffusion through the plasma membrane, a consideration that holds true also for their transport through the inner mitochondrial membrane. Thus, we are back to the same problem and, at least in principle, the involvement of the same active transporters for AA, and facilitative GLUTs for DHA, should be taken into account.
There are, however, numerous differences between the context of the plasma and mitochondrial membranes. An important one is related to AA concentrations of the extramitochondrial milieu (cytosol), in general, remarkably higher than those of the extracellular milieu. Then, why should mitochondria use a high-affinity transport of the reduced form of vitamin C? In principle, it makes very little sense to express a mitochondrial transporter with a Km in the low micromolar range to transport AA from a cytosol containing millimolar levels of the vitamin. Moreover, we can think of additional critical limitations in transport efficiency based on the low cytosolic concentrations of Na+, Mg2+, and Ca2+ (35, 79). Although it would have been worth at least considering the possibility of the existence of such transporters working with a low affinity, due to the restrictive composition of the cytosol, many scientists working in the field nevertheless found this possibility unreasonable and focused their attention on the more likely involvement of DHA transport through GLUTs.
Mitochondrial transport of DHA through facilitative hexose transporters
As previously discussed, DHA is a substrate of hexose transporters and its uptake may represent a possible alternative strategy of vitamin C accumulation in specific cell types. The same transporters, however, are also expressed in mitochondria (6, 52, 57, 70, 75) despite the lack of an apparent role of glucose in mitochondrial metabolism. Furthermore, once in the cells, glucose is readily phosphorylated and glucose-6-phosphate is not a substrate for the hexose transporters. This means that DHA transport may take place in the absence of competing substrates. The second reason favoring the possibility of DHA transport through mitochondrial GLUTs is that this event is not influenced by the ionic microenvironment. Finally, high-capacity transport is expected to be advantageous for providing high levels of the vitamin under conditions of extensive mitochondrial formation of ROS. The role of GLUT1 as a DHA transporter has also been validated by in silico predictions (111).
After the initial demonstration in plant cells (112), experiments performed in cultured mammalian cells have significantly contributed to the demonstration of the physiological significance of mitochondrial DHA transport. These studies provided convincing information on the possibility of mitochondrial DHA transport, but in some circumstances, the approach employed was based on the results of comparative uptake studies performed on isolated mitochondria exposed to high concentrations of DHA and AA.
We critically considered this approach when the endpoint was cellular uptake (see above), the same is herein being proposed for studies measuring the mitochondrial uptake of the vitamin. More specifically, the use of a high concentration of AA seems appropriate in these studies as high concentrations of the vitamin are normally found in the cytosol of cells isolated from living organisms. However, the same is not true for the use of high concentrations of DHA, which will never be reached in the extramitochondrial compartments. Indeed, the DHA concentrations are low in the extracellular milieu and most likely even lower in the cytosol, in which it is indeed very rapidly reduced (14, 55, 60, 64, 76, 88).
We could then postulate that there will be more DHA to be transported under conditions of oxidative stress, as discussed above for the plasma membrane, but nevertheless an important piece of the puzzle seems to be missing. Time course studies of DHA uptake reveal its immediate intracellular reduction back to AA (14, 55, 60, 63, 64, 76, 88, 122). As this event is also observed using concentrations of DHA two orders of magnitude greater than those found in the extracellular milieu, it appears unlikely that intracellular AA can be oxidized at a rate compatible with the accumulation of cytosolic concentrations of DHA that are permissive for its mitochondrial uptake.
The scenario is therefore once again complicated by conflicting factors as the DHA transport appears likely to take place for several reasons, but at the same time, it remains unclear whether or not there will be enough DHA to be transported. In our opinion, we should not think in terms of cytosolic concentrations, but rather of selective microdomains, in which AA is oxidized to generate local concentrations of DHA. Moreover, we should consider likely a competition between reduction and transport of DHA, thereby implying that these microdomains must be located in the close proximity of DHA transporters.
The hypothesis we can make is that DHA transport in mitochondria is linked to its selective formation in the intermembrane space. As previously discussed, a fraction of the O2 −• generated at the level of complex III is released in the intermembrane space and, under these conditions, AA would be locally converted to DHA and build up local concentrations functional for significant DHA transport in the matrix (Fig. 3). The ability of AA to reach the intermembrane space has been previously documented (75). This mechanism may well represent the strategy used to transport appropriate amounts of vitamin C in the form of DHA: an enhanced rate of O2 −• release will be indeed associated with a selective increase of the DHA concentration in the intermembrane space, thereby bursting its uptake through GLUTs.

Mitochondrial transport of AA through SVCT2
While our previous considerations make an argument against the active transport of AA in mitochondria, a definitive demonstration in this direction has not yet been provided. Western blot studies for the detection of SVCTs are difficult to perform in the mitochondria isolated from animal tissues because of the problems encountered to obtain pure mitochondrial preparations (89). In addition, negative results would still leave some doubts since a very low expression of these transporters is eventually to be expected in mitochondria bathed in a milieu containing high levels of AA. Likewise, for these same reasons, transport studies performed on isolated mitochondria may fail to detect significant vitamin uptake over the short incubation time allowed by these experiments.
With these considerations in mind, we can approach this problem indirectly, once again with use of cultured cells, overexpressing active AA transporters in their plasma membrane when grown under standard conditions, that is, in the absence of the vitamin. The increased expression of these transporters in mitochondria, coupled with their functional activity, may at least prove that these transporters can indeed be expressed and can be functionally active also in these organelles.
We performed these experiments using U937 cells, which express both SVCT1 and 2 (3, 27), and were initially very skeptical about their outcomes since the cytosolic conditions in terms of cation concentrations, in particular Na+ and Ca2+, are very different from those of the extracellular milieu. To our surprise, we obtained both immunocytochemical and Western blot evidence of mitochondrial SVCT2 expression in the absence of detectable expression of SVCT1 (3, 27). The image reproduced in Figure 4 provides evidence for a virtually exclusive expression of SVCT2 in subcellular domains that can be stained by specific mitochondrial probes (27).

We reproduced these results in other cell types, for example, Raw 264.7 and C2C12 myoblasts (103, unpublished data), and other investigators detected SVCT2 in HEK-293T cells and reported a similar prevalent mitochondrial expression of SVCT2 (79). Another study (125) employed the same cell line to demonstrate that the high mitochondrial expression of the transporter detected by immunolocalization studies can be confirmed with the use of a novel fluorescent probe detecting SVCT2. These authors also used HepG2 cells, which express both SVCT1 and SVCT2. SVCT2 was mostly detected in mitochondria, with some minor expression in the plasma membrane. SVCT1 was instead uniquely detected in the plasma membrane.
The first conclusion we can draw from these studies is that SVCT2 is expressed in the mitochondria of various cell types grown in the absence of vitamin C. It is also interesting to observe that there was no evidence for the expression of mitochondrial SVCT1, even in cells expressing this transporter in their plasma membrane.
The obvious question to be asked is whether the mitochondrial expression of SVCT2 is compatible with its function and the results obtained were once again unexpected. Not only did these organelles avidly accumulate the reduced form of the vitamin, but also the Km of the transporter was very low, remarkably similar to that measured for the plasma membrane SVCT2 (27). An additional similarity was found in the pH dependence and positive cooperativity for Na+ (nH ≅2). Strikingly, however, 100-fold less Na+ was necessary to maximally support AA transport in mitochondria in comparison with transport through the plasma membrane. A final critical difference was found for the Ca2+/Mg2+ dependence; while the plasma membrane SVCT2 required millimolar concentrations of these cations, its mitochondrial counterpart was fully active in their virtual absence. We reproduced some of these results in Raw 264.7 cells and obtained remarkably similar results (unpublished data).
Other studies confirming the existence of a mitochondrial SVCT2 in human HEK-293 cells showed that under intracellular ionic conditions, SVCT2 instead behaves as a low-affinity AA transporter (79). Recently, we found that the mitochondrial SVCT2 expressed in C2C12 myoblasts also works with low affinity (103).
These results therefore lead us to a second conclusion, that is, the mitochondrial SVCT2 is functionally active, although its affinity is potentially compromised by the low intracellular concentrations of specific ions. On the other hand, probably as a consequence of the very high requirement of the vitamin in their mitochondria, some cell types, such as the U937 cells employed in our studies, have evolved a strategy to make this same transporter fully active and with a high affinity by minimizing its cation requirements (27).
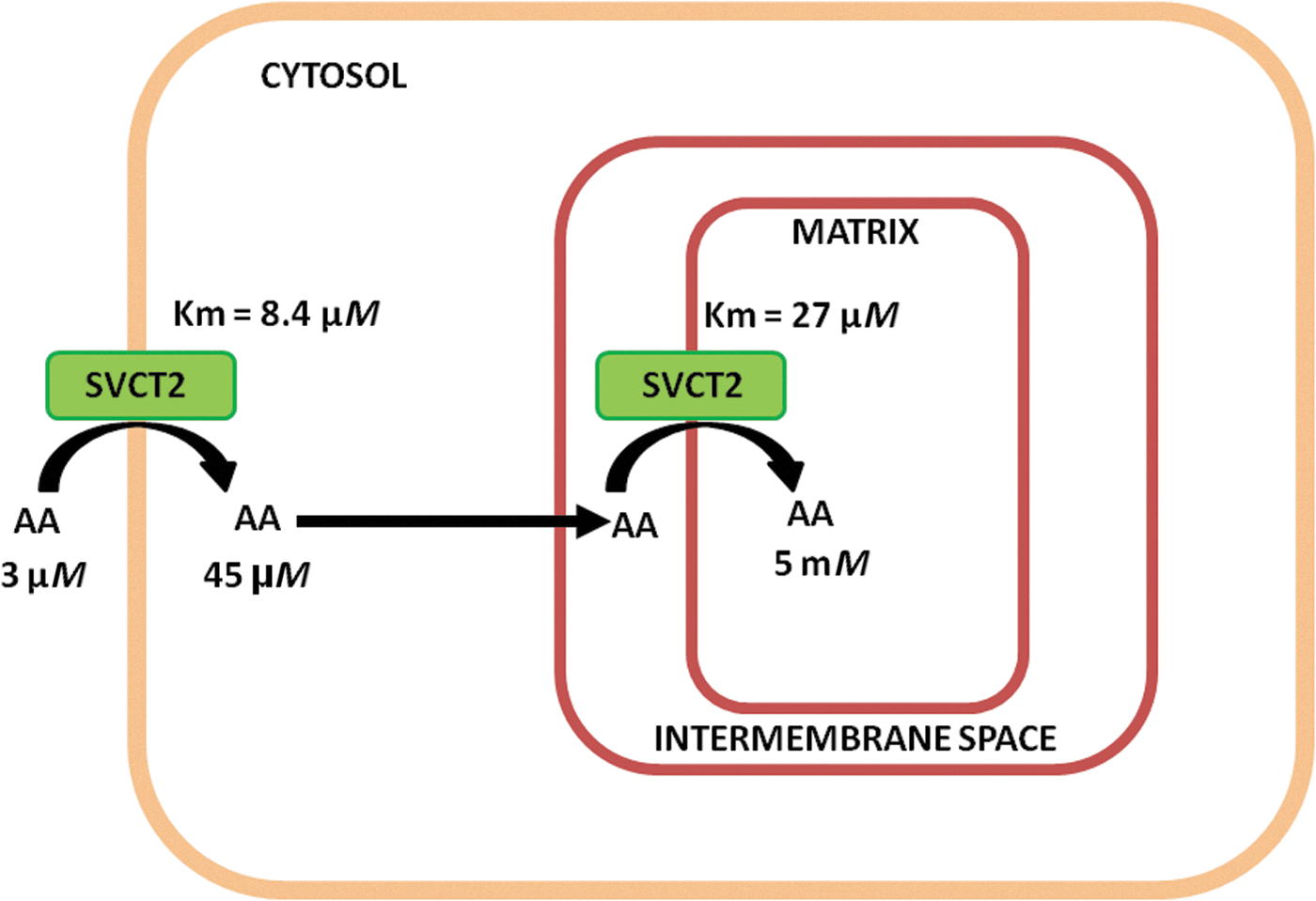
The third conclusion we can make is inspired by the combination of some of the above findings. We mentioned that the high-affinity SVCT2 of the plasma membrane is less densely expressed than its mitochondrial counterpart. We should interpret this observation as an indication that the cytosolic concentrations of the vitamin may eventually be kept low by the mitochondrial clearance processes, in particular at limiting extracellular levels of the vitamin. We found that this is indeed the case as a 15-min exposure to a very low concentration of AA, 3 μM, leads to a cellular concentration of about 50 μM and an intramitochondrial concentration of as much as 5 mM (Fig. 5). This means that during uptake, the increase in cytosolic concentration of the vitamin is continuously hampered by its more efficient mitochondrial uptake.
Based on these results, we may predict that U937 cells as well as other cultured cells expressing high-affinity mitochondrial transport of AA manifest a greater need of the vitamin in their mitochondria than in the cytosol. We take these results as indicative of a critical biological relevance of mitochondrial vitamin C, which can be easily tested against specific mitochondrial toxins.
The U937 cell model provides a very interesting system for asking questions on the relevance of mitochondrial versus cytosolic AA in the toxic response evoked by specific agents. We recently demonstrated that in the same U937 cell clone used in the above studies, arsenite promotes toxicity via a mechanism mediated by the mitochondrial formation of O2 −•/H2O2, which triggers downstream mitochondrial permeability transition-dependent apoptosis (36 –38).
All these events were sensitive to either rotenone, a complex I inhibitor, or the respiration-deficient phenotype. A 15-min pre-exposure to 10 μM AA, while producing a very high mitochondrial accumulation of AA/very low increase in the cytosolic concentration of the vitamin, generated the same protective effects (36, 37). This experimental system enabled us to link cytoprotection with the mitochondrial fraction of AA, but we could do even more using DHA in the place of AA. DHA, for the reasons that will be detailed below, can be used to enhance the cytosolic fraction of the vitamin with no apparent increase in its mitochondrial content. Under these conditions, the significant elevation of cytosolic vitamin C associated with DHA exposure failed to promote cytoprotection (37).
DHA as an inhibitor of cellular and mitochondrial uptake of AA through SVCT2
We recently investigated the existence of possible interactions between AA and DHA transport with an initial experiment that is very simple. The cells were exposed to a single concentration of either AA or DHA or the two forms of the vitamin in combination (28). The results were surprising since the uptake observed with the combined treatment was identical to that observed with DHA alone. In addition, the uptake was (in both circumstances) sensitive to cytochalasin B, an inhibitor of DHA transport, and insensitive to Na+ omission, which inhibits AA transport. This was the first evidence of an inhibitory effect of DHA on AA transport. This notion was then established using different approaches and led us to formulate the hypothesis that extracellular oxidation of AA may lead to transient and reversible negative modulation of transport of the reduced form of the vitamin with a parallel switch to the high-capacity facilitative transport of DHA.
We are currently investigating the mechanism(s) involved in this inhibitory response and more specifically challenging the possibility that DHA elicits its effects via interaction with critical thiols of SVCT2. Consistent with this notion are our previous results indicating that the plasma membrane and mitochondrial SVCT2 are both susceptible to inhibition by thiol-reactive agents (29). Furthermore, DHA is known to react with –SH groups and to inhibit the activity of enzymes containing critical cysteines (30).
At the moment, we do not have enough information to determine the physiological relevance of this event, but we can imagine that conditions associated with the bystander effect discussed above will likely result in inhibition of AA transport in SVCT2-expressing cells.
More research is needed to understand the extracellular cross-talk between the two forms of the vitamin, but probably even more interesting is the comprehension of these same interactions taking place intracellularly, with a potential impact on vitamin C accumulation in specific subcellular compartments—mitochondria in the first place.
We very well know that exposure of U937 cells to low concentrations of AA (e.g., 3 or 10 μM) is associated with prompt cellular uptake and further mitochondrial accumulation of the vitamin. Under the same conditions, however, the uptake of DHA—even when resulting in greater cellular levels of AA—was not associated with significant mitochondrial uptake (29). This event was only observed after exposure to very high concentrations of DHA, 50 μM or more. We further investigated this issue in both intact cells and isolated mitochondria and concluded that low intracellular concentrations of DHA, because of their rapid reduction back to AA, are a poor substrate for direct mitochondrial uptake. DHA concentrations, however, were sufficiently high to mediate inhibition of mitochondrial transport of DHA-derived AA (29). The increased accumulation of vitamin C observed in cells treated with the very high concentrations of DHA was instead due to its direct mitochondrial uptake.
Conclusions
AA mediates numerous effects in various subcellular compartments and its hydrophilic nature implies the expression of an active transporter, typically SVCT2, which mediates high-affinity/low-capacity uptake through the plasma membrane and in subcellular compartments in which it is eventually expressed. An alternative pathway is dependent on the oxidation of AA to DHA and its uptake by facilitative hexose transport, once again through the plasma membrane and in different compartments. In this review, we focused on the plasma membrane and mitochondrial transport of the vitamin and made an attempt to critically interpret the available information largely based on results obtained in cultured cells. A summary of the mechanisms discussed and proposed is provided in Figure 6.
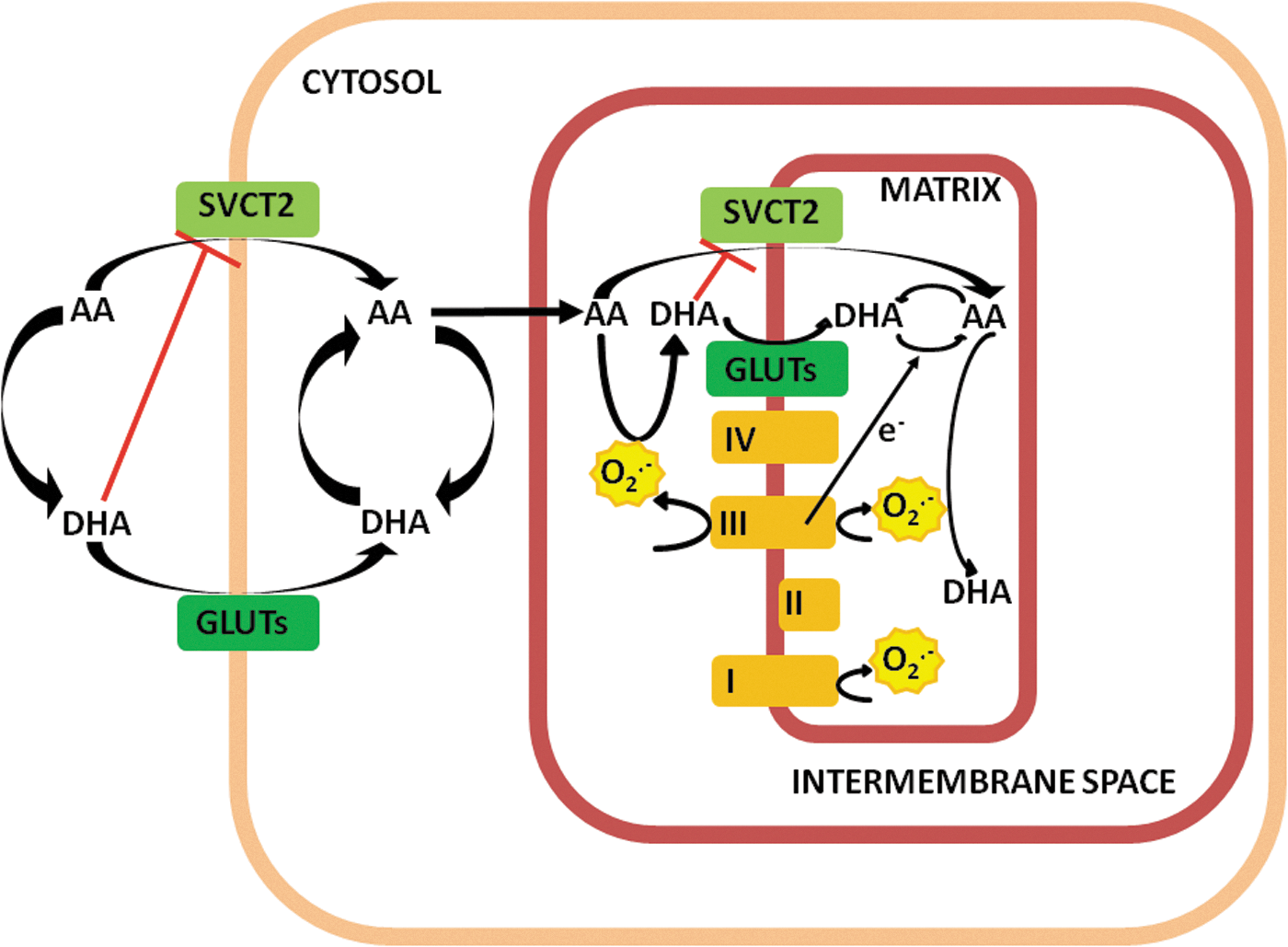
Based on the results obtained in our laboratory and on the interpretation of available information, cellular uptake of vitamin C appears to be mostly dependent on SVCT2-mediated transport of AA, provided that this transporter is expressed in the plasma membrane.
Extracellular oxidation of AA to DHA may lead to inhibition of SVCT2-mediated transport of AA and will be permissive for some uptake through GLUTs, especially in cells in which these transporters are highly expressed. The prompt reduction of DHA back to AA likely prevents the possibility of a direct mitochondrial uptake of DHA, which might eventually inhibit mitochondrial SVCT2, when expressed in these organelles. DHA, however, can also be produced by AA oxidation in the intermembrane space mediated by the O2 −• generated at the level of complex III and released in this compartment. This event promotes an antioxidant response associated with the scavenging of O2 −• and allows the DHA-dependent inhibition of SVCT2-mediated AA uptake.
Most importantly, this mechanism links the rate of DHA uptake with the rate of O2 −• release. Under these conditions, the switch in the uptake mechanism is also advantageous since intramitochondrial DHA is readily recycled back to AA by various mechanisms dependent on GSH and/or other reductants or through the acceptance of electrons released by complex III (or complex I) of the respiratory chain. This last event clearly prevents O2 −• formation in the matrix.
The scheme depicted in Figure 6, which obviously needs to be validated by further experimental evidence, nevertheless appears attractive since it provides a likely strategy for the mitochondrial accumulation of vitamin C based on a mechanism associated with an effective antioxidant action exerted in the intermembrane space as well as in the matrix. More work is also necessary to understand the complex relationships of these events with other key steps of vitamin C recycling, for example, associated with the formation of semidehydroascorbic acid and/or conversion of the latter to either AA or DHA in the intermembrane space. At least in principle, the NADH-semidehydroascorbic acid reductase activity identified in the outer mitochondrial membrane in rat hepatocytes (48) may significantly contribute to events associated with the transport of the reduced form of the vitamin.
Footnotes
Acknowledgment
This research was supported by Ministero dell'Università e della Ricerca Scientifica e Tecnologica, Programmi di Ricerca Scientifica di Rilevante Interesse Nazionale, 2015 (Grant number: 2015MJBEM2-003).