
Abstract
Significance:
Diabetes is associated with markedly accelerated rates of micro- and macrovascular complications that increase morbidity and mortality. Understanding the molecular mechanisms can promote much needed therapeutics.
Recent Advances:
Long noncoding RNAs (lncRNAs) are important regulators of gene regulation and cellular function and are emerging as important players in diabetes and its complications. There are number of examples in which lncRNAs are responsive to hyperglycemia and clearly involved in regulation of genes and pathways associated with the development of diabetic complications.
Critical Issues:
As there are likely thousands of lncRNAs that are expressed in any given tissue, understanding how they are regulated and function in the normal healthy state as well as pathological states is a challenge.
Future Directions:
Further studies in how lncRNAs are involved in the development and progression of diabetic complications as well as development of methods to target dysregulated lncRNAs or evaluate them as biomarkers of early detection of organ dysfunction will be highly beneficial to treating diabetic patients.
Introduction
W
Long noncoding RNAs (lncRNAs) have garnered much attention in recent years owing to their influence on a variety of molecular pathways. In the literature, the best well-known lncRNA is Xist that was originally discovered in 1991 (10). This highly expressed regulatory lncRNA regulates X-inactivation through recruitment of polycomb repressive complex 2 (PRC2) silencing complex to the X chromosome from which it is transcribed (21, 58). This function of Xist in cis leads to spreading of the silencing complex to the rest of the inactivated X promoting heterochromatin formation and transcriptional silencing (21).
Although Xist has been studied since the early 90s, the interest in lncRNAs only gained momentum in the last decade with the advent of next-generation sequencing technology. These high-resolution sensitive methods have uncovered thousands of these noncoding RNAs, which, for the most part, are lowly transcribed in both normal and disease states (29). As a class, these nonprotein coding RNAs are RNA polymerase II-dependent transcripts that are often processed much like protein-coding RNAs: they are 5′ capped and 3′ polyadenylated (12, 23, 29).
Today, lncRNAs are generally classified as noncoding RNAs that are >200 nucleotides long and have a multitude of functions in the cell (43). Similar to Xist, other lncRNAs can also regulate chromatin modifying complexes. For example, the HOTAIR transcript, transcribed from the HOX locus can bind to both the PRC2 complexes as well as the lysine-specific demethylase-1A/REST corepressor/RE1-silencing transcription factor (LSD1/REST/CoREST) complex (50a). In addition to binding to chromatin modifiers, lncRNAs can function as scaffolds for protein complexes. Linc-p21 transcripts interact with hnRNPs in the regulation of p21, which encodes a well-known tumor suppressor, in cancer cells (25). The interaction of these lncRNAs with protein complexes highlights the importance of lncRNA–protein interaction in lncRNA function. It is likely that these lncRNA–protein interactions are dependent on the secondary structure of the lncRNA.
In contrast, a number of additional lncRNA functions are dependent on primary RNA sequence. For example, linc-MD1 lncRNA, transcribed during muscle differentiation, squelches microRNAs (miRNAs), miR-135 and miR-133, from their targets, MEF2C and MAML1 mRNAs. Therefore, expression of linc-MD1 lncRNA reduces the efficacy of miRNA-directed degradation of MEF2C and MAML1 mRNAs (12a). In addition to binding of miRNAs, lncRNAs can also directly interact with targeted mRNAs and affect their stability. lncRNAs termed half STAU1-binding site RNAs (1/2-sbsRNA) bind target 3′UTRs of mRNAs through imperfect base pairing through their Alu element sequence and the resulting RNA duplex is degraded through the Staufen 1 (STAU1)-mediated messenger RNA decay (SMD) (22). Interestingly, the terminal differentiation-induced ncRNA (TINCR) lncRNA also binds to target mRNAs through a 25-nucleotide motif sequence that is also recognized by the SMD machinery (32). These examples show how lncRNAs impact gene expression at the post-transcriptional level in a sequence-dependent manner. Finally, lncRNAs have also been described to host miRNAs. In fact, about 10% of lncRNAs are host transcripts for miRNAs (16, 26). The variety of molecular functions (Fig. 1) make lncRNAs very appealing and interesting, but also challenging to study.

Evaluation of the functional roles of lncRNAs in diabetes and diabetic complications has only begun in recent years. Indeed, studies showing a number of human single-nucleotide polymorphisms (SNPs) associated with type 1 diabetes (T1D) and type 2 diabetes (T2D) are located within lncRNA loci (51, 52), suggesting that lncRNAs may be involved in diabetes. In this review, we discuss notable lncRNAs involved in metabolic tissues (i.e., pancreas, liver, muscle, and adipose) as well as those known to be involved in diabetic complications (Fig. 2).
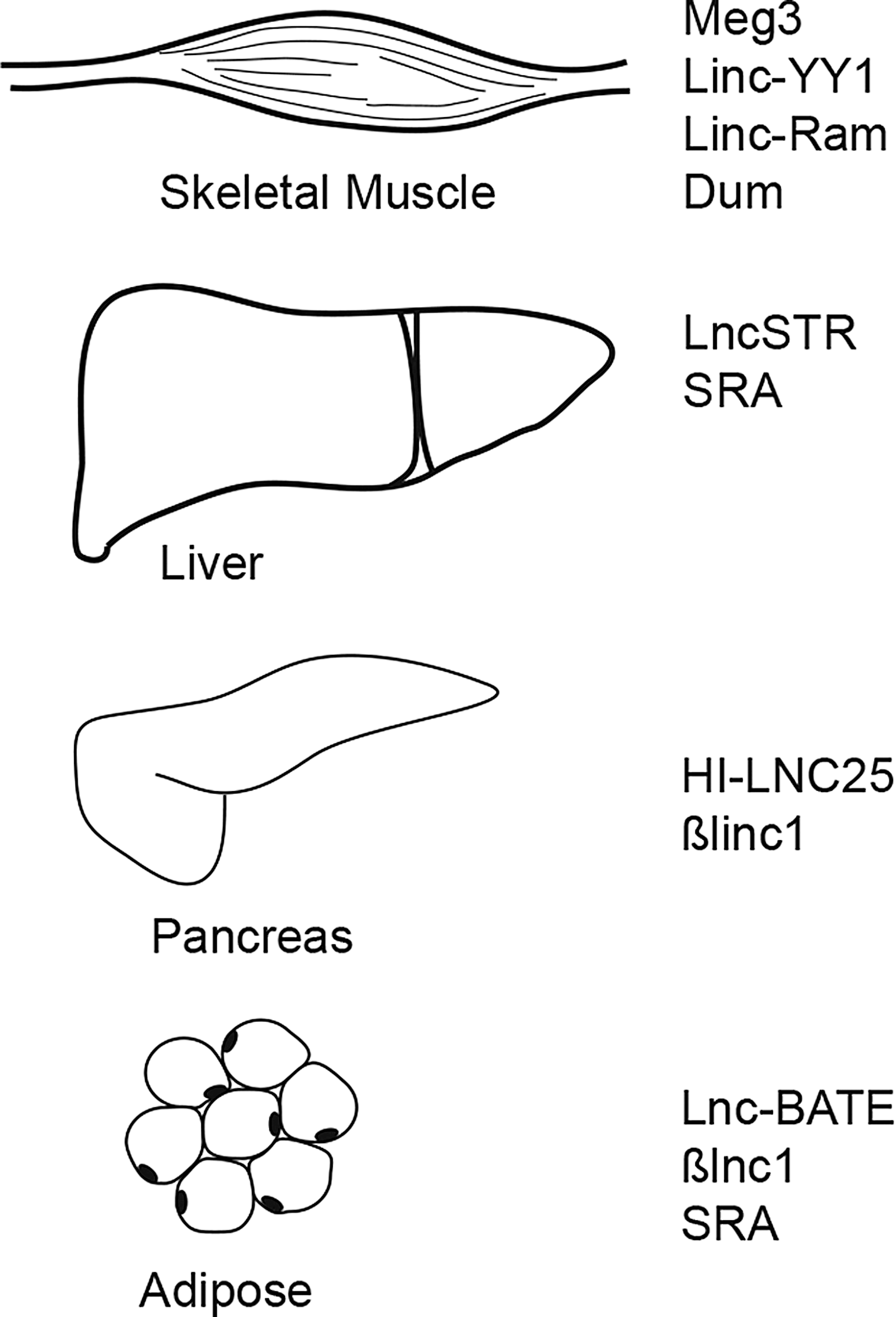
lncRNAs in Metabolic Tissues
Pancreas
The pancreas is responsible for sensing glucose levels and secreting insulin from the pancreatic beta cells (β-cell) to stimulate glucose uptake in peripheral tissues. In T2D, hyperglycemia results from β-cell dysfunction and insulin resistance, which is the inability to respond to normal insulin levels.
In T1D, hyperglycemia is due to loss of β-cells and insulin production. In recent years, it has become clear that lncRNAs are involved in development of the pancreas and its normal function. In fact, with high throughput sequencing efforts, greater than 1000 intergenic and antisense lncRNAs, were identified as transcribed in islet cells (41). Specifically, one lncRNA, HI-LNC25, is a β-cell-specific lncRNA that regulates GLIS3 mRNA (41). More recent studies have found that the role of lncRNAs in β-cell function may be pervasive. Analysis of β-cell transcriptome has shown that β-cell lncRNAs can control major regulatory networks involved in β-cell specification and formation (4). Specifically, β-cell long intergenic noncoding RNA 1 (βlinc1) is an lncRNA that regulates neighboring genes encoding transcription factors that are required for islet development. Loss of βlinc1 lncRNA results in dysregulated glucose homeostasis in mouse models (4). Comparisons of islet transcriptomes generated by RNA sequencing from islets isolated from normoglycemic and hyperglycemic donors uncovered lncRNAs that are differentially expressed in the two groups as well as those associated with hemoglobin A1c (HbA1c levels) (18). Further analysis of the functional roles of these lncRNAs may determine whether they are involved in islet dysfunction.
Importantly, cross-species analysis comparing human to mouse islet transcriptomes by RNA sequencing suggests that many lncRNAs are commonly regulated and transcribed in both species during β-cell development (8, 41). These analyses are useful to prioritize lncRNAs expressed in both humans and mice and thereby allow researchers to utilize mouse models to evaluate the function of lncRNAs in vivo.
Liver
The liver is the site of many metabolic processes including lipid synthesis, drug metabolism, and, importantly, gluconeogenesis. In normal liver function, a number of lncRNAs have been identified. Specifically, lnc liver-specific triglyceride regulator (lncLSTR) is a liver-specific lncRNA that regulates systemic lipid metabolism (36). This lncRNA binds to TDP-43 to regulate Cyp8b1 activity, altering bile acid levels that affect the farnesoid X receptor (FXR) pathway to promote triglyceride clearance in the liver (36). This initial study uncovered how lncRNAs can be involved in normal liver metabolic pathways. Similarly, the lncRNA SRA is involved in modulating lipid levels by regulating the expression of adipose triglyceride lipase (ATGL), which is critically involved in hydrolyzing hepatic triacylglycerol (13). SRA transcript inhibits forkhead box protein O 1 (FoxO1), which normally induces ATGL (13). These examples highlight the importance of lncRNAs in normal liver function. Additional studies have identified lncRNAs in human livers that are associated with liver disease such as nonalcoholic fatty liver disease and nonalcoholic steatohepatitis (5, 46, 47). Diabetic subjects often have metabolic dysfunction of the liver, and thus understanding the role of lncRNAs in liver pathology is also beneficial.
Muscle
Muscle cells are important for uptake of glucose in response to insulin secretion. Myogenesis and myogenic differentiation are critical processes that involve lncRNAs. One critical lncRNA is the MEG2 lncRNA that is located within the DLK1-MEG3 locus. This locus has been of interest for quite some time because it is an imprinted locus wherein the adjacent genes DLK1 and MEG1 are reciprocally imprinted (17). Loss of MEG3 expression in knockout mouse models results in perinatal lethality and skeletal muscle defects (62). Additional lncRNAs involved in myogenic differentiation include linc-YY1 and lncRNAs relevant to MyoD activity. In skeletal myogenesis, linc-YY1 lncRNA complexes directly with the transcription factor Yin Yang 1 (YY1) that interacts with the PRC2 complex to silence genes in trans (60). The presence of a human ortholog of linc-YY1 lncRNA suggests the function of this lncRNA is conserved between humans and mouse (60). MyoD encodes a vital transcription factor in myogenic differentiation (11). MyoD induces the transcription of at least two lncRNAs, Linc-RAM and Dum (53, 56). Linc-RAM lncRNA binds to MyoD and together can interact with additional chromatin modifiers: Baf60c and Brg1, two important nucleosome remodelers (20). The ribonucleotide complex targets myogenic genes to properly coordinate gene expression during differentiation (56). Dum lncRNA is also induced by MyoD (53). In cis, transcription of the lncRNA recruits DNA methylation proteins to silence the neighboring developmental pluripotency-associated 2 (Dppa2) gene during differentiation (53). These examples clearly show the importance of lncRNAs in regulating myogenesis and myogenic differentiation at the transcriptional level.
Adipose
Adipose is a major insulin responsive site and adipose cells make key adipokines that have critical metabolic functions. In obesity, there is increased adipose inflammation due to accumulation of macrophages and this can contribute to insulin resistance, T2D, and the metabolic syndrome (34). Transcriptome analysis of cells undergoing adipogenesis reveals that many lncRNA are expressed during adipocyte differentiation and maturation, suggesting these transcripts play an important role in the development of adipose tissue (48). Indeed, deep sequencing of transcriptomes from white adipose tissue, brown adipose tissue, and epididymal tissue reveals that many lncRNAs are adipose tissue-specific (1). One lncRNA, lnc-BATE1, functions to regulate the development of brown adipocytes through its interaction with hnRNP U (1). Brown adipocyte development also involves Blnc1, an lncRNA that interacts with the transcription factor EBF2 to regulate thermogenic gene regulatory programs (59). In addition to functions in adipose differentiation, lncRNAs can impact the physiological state of an animal. For example, in mouse models, SRA lncRNA knockouts are more insulin sensitive and resistant to diet-induced obesity (38). It is likely that there are additional adipose cell-expressed lncRNAs involved in obesity, which can impact the development of diabetes.
lncRNAs in Diabetic Complications
Retinopathy
Ocular disease (retinopathy) is one of the common complications related to diabetes. A few lncRNAs have been implicated in diabetic retinopathy (Fig. 3). Retinas derived from diabetic rodents reveal that Malat1 and Meg3 lncRNAs are dysregulated when compared with nondiabetic rodents (37, 42). Malat1 lncRNA was found to be upregulated in retinas under high glucose conditions, whereas knockdown of Malat1 lncRNA attenuated retinopathy in STZ-induced rats (37). This suggests that this lncRNA promotes retinopathy under diabetic condition. In addition, retinal endothelial cells cultured in high glucose show increased expression of linc-MIAT lncRNA, which can function as a competitive endogenous RNA of miR-150 to regulate vascular endothelial growth factor (VEGF) levels, or to suppress miR-29b in regulating apoptosis (6, 54, 57). This reveals how microvascular cells utilize lncRNAs under normal and diabetic conditions.
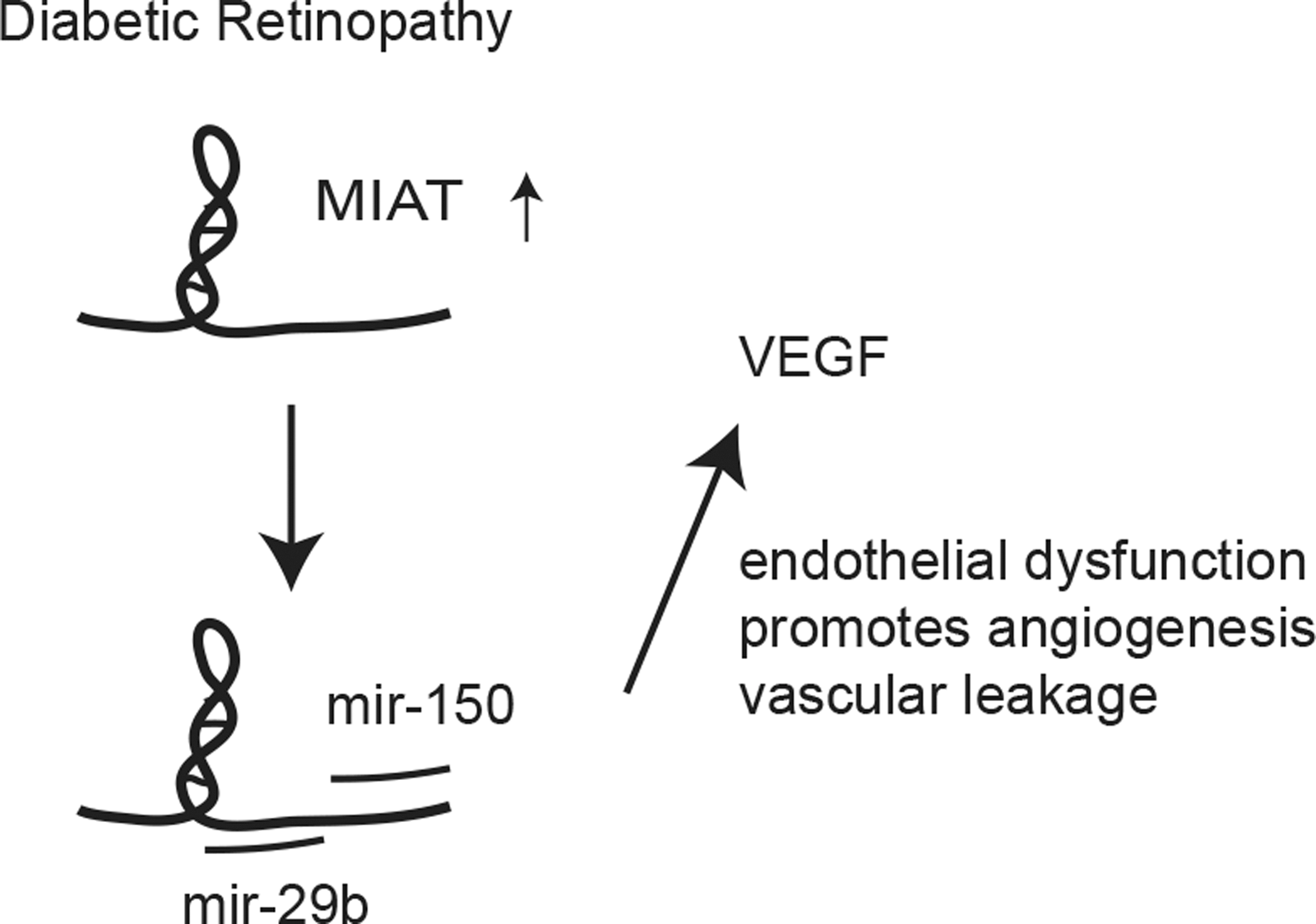
In addition, VEGF, a potent angiogenic factor, is also regulated by the lncRNA ANRIL (49). Human retinal endothelial cells treated with high glucose upregulated the VEGF and ANRIL transcripts, whereas siRNAs targeting ANRIL could abrogate high glucose-induced VEGF expression. ANRIL is an lncRNA originating from the CDKN2A-CDKN2B locus, and has been implicated in a number of vascular-related diseases (15). Further analysis of retina transcriptomes in healthy and disease states will allow for additional identification of lncRNAs involved in diabetic retinopathy. Since ocular injections with anti-VEGF antibodies are already in clinical use, combination with lncRNA-based oligonucleotide therapy can be a potential treatment in the future.
Nephropathy
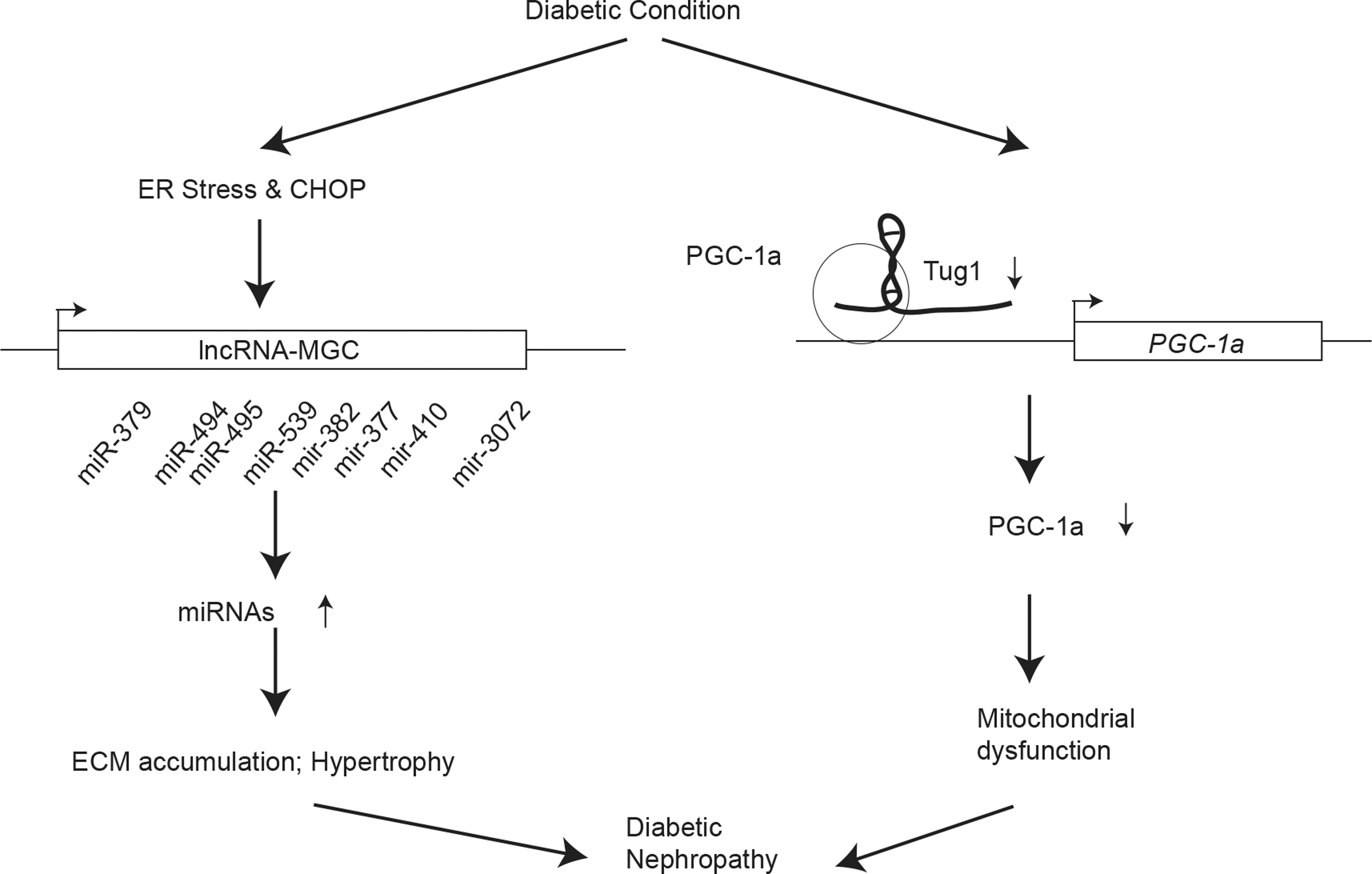
Diabetes leads to significant impairment of kidney function leading to diabetic nephropathy, one of the most common and debilitating complications of diabetes, which can result in end-stage renal disease. Evidence shows that lncRNAs also clearly play a role in the development of renal disease (Fig. 4). One of the major features of diabetic nephropathy is increased fibrosis in most renal cells due to the accumulation of extracellular matrix (ECM). One lncRNA, PVT1 lncRNA, seems to be involved in this process during diabetes. First, PVT1 transcripts are upregulated in human renal mesangial cells with high glucose treatment, and second, manipulation of PVT1 expression affects the accumulation of ECM (2). Interestingly, PVT1 lncRNA also harbors miR-1207. This miRNA functions independently of the lncRNA and regulates transforming growth factor-beta1 (TGF-b1), plasminogen activator inhibitor-1 (Pai-1), and fibronectin (FN1), all important ECM-related genes (2). A couple of other lncRNAs have also been shown to regulate growth and fibrosis in diabetic nephropathy. lnc-MGC is an lncRNA that is regulated by the endoplasmic reticulum (ER) stress-related transcripton factor, CHOP. Treatment of glomerular mesangial cells with high glucose or TGF-b1 results in an increase in lnc-MGC transcript levels, which promotes the expression of a cluster of several miRNAs encoded within the transcript (28). Levels of lnc-MGC and the cluster miRNAs are also increased in glomeruli derived from mouse models of diabetic nephropathy. Target genes of the lnc-MGC encoded miRNAs have functions related to ER stress and protein synthesis. Further analysis in CHOP knockout mice, which are protected from diabetic nephropathy, showed a decrease in glomerular lnc-MGC and cluster miRNA expression, suggesting that they play a role in early features of the disease (28). Notably, a modified antisense oligonucleotide (gapmer) targeting lnc-MGC inhibits candidate cluster miRNAs, glomerular ECM, and hypertrophy in diabetic mice, further supporting the translational value of lncRNA-based interventions for diabetic complications.
lncRNAs have also been found to be dysregulated in models of renal fibrosis, and many of them were regulated by Smad3 transcription factor, a key regulator of fibrotic genes and effector of TGF-b1 (61).
Podocytes are cells with critical functions in the renal glomerulus, and diabetes leads to podocyte effacement and dysfunction. Recent studies also show a role for lncRNAs in podocyte functions in diabetic nephropathy. taurine-upregulated 1 (Tug1) lncRNA affects mitochondrial function in podocytes by modulating the expression of peroxisome proliferator-activated receptor γ (PPARγ) coactivator α (PGC-1α), a transcription factor important for mitochondrial biogenesis. Tug1 is downregulated under diabetic conditions. Podocyte-specific overexpression of Tug1 in diabetic mice can confer improvements in mitochondrial bioenergetics and reverse several key features associated with diabetic nephropathy (39). Furthermore, lncRNAs are also involved in local inflammation in the kidney. LincRNA-GM4419 is an lncRNA that regulates the activity of NF-κB/NLRP3 inflammasome (55). This lncRNA can directly interact with the p50 subunit of NF-κB in promoting inflammation. Knockdown of LincRNA-GM4419 reduces inflammation in mouse model (55). These examples make it clear that lncRNAs are involved in diabetic nephropathy. Increasing interest in the ncRNA field has generated new initiatives and projects such as the “Kidney Precision Medicine” Project supported by the National Institutes of Health, which will likely reveal a larger number of lncRNAs involved in kidney function and in disease.
Cardiovascular disease
Diabetes promotes vascular inflammation and cardiovascular diseases, including hypertension and atherosclerosis. Key cells involved in these pathological conditions are monocyte/macrophages and vascular smooth muscle cells. Transcriptome profiling of bone marrow macrophages isolated from diabetic Leprdb/db mice and control nondiabetic Leprdb/+ mice revealed that diabetes alters several lncRNAs. The function of one of these lncRNAs, E330013P06, which is upregulated in macrophages from Leprdb/db and high fat diet-induced type 2 diabetic mice, was further characterized. It was also upregulated, along with inflammatory genes, in mouse macrophages treated with high glucose and palmitic acid, a diabetic stimulus. E330013P06 overexpression enhances the response to inflammatory signals, and foam cell formation, whereas siRNA-mediated gene silencing inhibits the response to inflammatory signals. These results reveal the first functional role for a macrophage lncRNA in diabetic conditions, and also yield new insights into mechanisms by which macrophages mediate inflammation in diabetic complications (44). These observations also support potential future lncRNA-based therapies for inflammatory diabetic complications.
In vascular smooth muscle cells, angiotensin II signaling, which is often elevated in diabetes and cardiovascular disease, promotes the expression of a number of lncRNAs (35). One specific lncRNA, lnc-Ang362, is upregulated with angiotensin II signaling in vascular smooth muscle cells and is a host RNA for two miRNAs, mir-221 and mir-222, which are involved in vascular smooth muscle cell proliferation (Fig. 5A) (35). Therefore, induction of lnc-Ang362 lncRNA leads to upregulation of the miRNAs and increase in cell proliferation. This initial study highlighted the relevance of lncRNAs in vascular cell function. Additional studies in human vascular smooth muscle cells have revealed additional lncRNAs that are involved in vascular cell function. For example, SENCR is an lncRNA involved in regulation of many contractile genes as well as genes involved in regulation of MYOCD, an important transcriptional regulator (9, 50). More recently, SMILR lncRNA was found in atherosclerotic plaques of human patients. Knockdown of SMILR results in decrease of vascular proliferation, suggesting that the lncRNA promotes cellular proliferation in disease states (Fig. 5B) (7). Furthermore, development of vascular endothelial cells relies upon at least three lncRNAs (33). Analysis of one of these, PUNISHER lncRNA, has shown that it is important for mitosis and cell division, vessel maturation, and density lipoprotein uptake (33). Altogether, these studies show the importance of lncRNAs in vascular tissues and in the development of vascular complications such as restenosis, hypertension, and atherosclerosis.

Future of lncRNAs in Diabetic Complications
lncRNAs are important regulatory RNAs that can function in many cellular processes during normal and disease states. In diabetes, recent work has shown that lncRNAs, as a class, are often dysregulated during pancreatic β-cell differentiation, as well as by hyperglycemia and related growth factors. They can also contribute to diabetic complications via changes in inflammation, fibrosis, ER stress, oxidant stress, and mitochondrial dysfunction. Undoubtedly, as we learn more about the transcriptome of diabetic tissues, we will identify more lncRNAs that function in the development of diabetic complications. It is important to note that lncRNAs can be individual specific. Analysis of 10 transcriptomes from granulocytes from different individuals revealed that lncRNAs expression varies among individuals at a level that is higher than protein-coding genes (31). Understanding how expression of a specific lncRNA increases an individual's susceptibility to diabetic complications will be useful. In addition, SNPs identified to be associated with diabetes and its complications located within lncRNAs loci (51, 52) can be used to identify individuals with elevated risk of diabetes. Although valuable catalogs of lncRNAs expressed in normal cells and tissues have been recently published (24), transcriptomic analysis of diseased tissues to uncover dysregulated lncRNAs and further analysis of their molecular function remain to be performed.
Recently, a number of studies have found that RNAs previously classified as noncoding can code for polypeptides that are involved in muscle cell processes (3, 40). These unanticipated findings highlight the need for rigor in the study of lncRNAs. It is also particularly important to examine the molecular mechanisms of action of lncRNAs, their protein binding partners, and genomic targets. These efforts have been facilitated by emerging technologies such as chromatin isolation by RNA purification (ChIRP-seq) (14), RNA-immunoprecipitation coupled to mass spectrometry (25), which have identified protein partners important for lncRNA function, and RNA of isolated chromatin assay, which directly identifies RNA–chromatin interactions (30).
Given the impact of lncRNAs in disease states, development of therapeutic interventions targeting lncRNAs will be important. As indicated earlier, our laboratory has shown that chemically modified gapmer oligonucleotides with target-specific sequences can be used to inhibit the activity of lncRNAs in vivo in mouse kidney (28). This suggests that similar strategies can be applied to diabetic nephropathy. In addition, in the clinic, expression of lncRNAs in biofluids can be a valuable noninvasive biomarker for early detection of diabetic complications, a major unmet need in clinical management. Finally, integrative analysis of lncRNA and mRNA networks across tissues in normal and disease states will be useful to understand their function in the whole individual (Fig. 6).
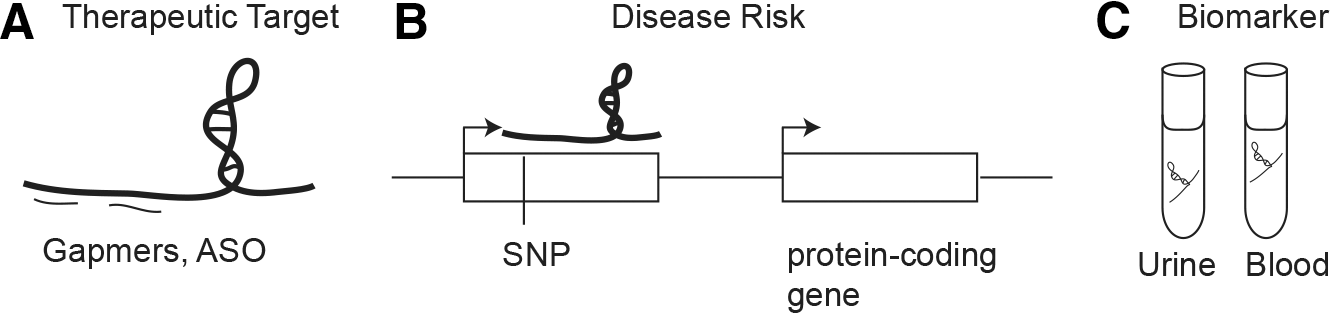
As evident by the recent discoveries of the role of lncRNA in metabolic tissues and under diabetic condition, the field will undoubtedly uncover more lncRNAs that are important in the development of diabetes as well as its associated complications.
Footnotes
Acknowledgment
The authors gratefully acknowledge funding from the National Institutes of Health, R01 DK 065073, R01 HL106089, and R01 DK081705 (to R.N.) and K01 DK104993 (to A.L.).