
Abstract
Significance:
Oxidative stress contributes to numerous pathophysiological conditions such as development of cancer, neurodegenerative, and cardiovascular diseases. A variety of measurements of oxidative stress markers in biological systems have been developed; however, many of these methods are not specific, can produce artifacts, and do not directly detect the free radicals and reactive oxygen species (ROS) that cause oxidative stress. Electron paramagnetic resonance (EPR) is a unique tool that allows direct measurements of free radical species. Cyclic hydroxylamines are useful and convenient molecular probes that readily react with ROS to produce stable nitroxide radicals, which can be quantitatively measured by EPR. In this work, we critically review recent applications of various cyclic hydroxylamine spin probes in biology to study oxidative stress, their advantages, and the shortcomings.
Recent Advances:
In the past decade, a number of new cyclic hydroxylamine spin probes have been developed and their successful application for ROS measurement using EPR has been published. These new state-of-the-art methods provide improved selectivity and sensitivity for in vitro and in vivo studies.
Critical Issues:
Although cyclic hydroxylamine spin probes EPR application has been previously described, there has been lack of translation of these new methods into biomedical research, limiting their widespread use. This work summarizes “best practice” in applications of cyclic hydroxylamine spin probes to assist with EPR studies of oxidative stress.
Future Directions:
Additional studies to advance hydroxylamine spin probes from the “basic science” to biomedical applications are needed and could lead to better understanding of pathological conditions associated with oxidative stress. Antioxid. Redox Signal. 28, 1433–1443.
Introduction
R
Current Methods for ROS Detection
Various methods for ROS measurement have been recently reviewed in references 20, 21, 32, 72, 82. It is important to note that there is no method for ROS detection that is currently accepted as a “gold standard,” since every sensor or reagent has advantages and disadvantages including cyclic hydroxylamine spin probes.
Spin trapping is the most unambiguous method for detection of free radicals (36, 66). Spin traps react with the short-lived free radical via covalent bond formation (radical addition reaction) to produce persistent nitroxide called spin adduct, which would have a “signature” electron paramagnetic resonance (EPR) spectrum specific for each trapped radical. Regretfully, spin trapping of many free radicals proceeds with much lower rates as compared with their scavenging with cellular antioxidants. For instance, superoxide dismutase (SOD) and ascorbate may prevent O2 •−detection in cells and tissues (96). Moreover, intracellular reductants such as ascorbate and flavin proteins reduce radical adducts to EPR-silent diamagnetic products, severely limiting the use of spin traps in biological systems (99).
Fluorescent probes produce stable products upon reactions with ROS and provide high sensitivity detection. Unfortunately, applications of fluorescent probes in ROS studies are hindered by serious issues: (i) formation of multiple nonspecific oxidation products, (ii) light sensitivity, and (iii) redox cycling of the probes (41). For example, dihydroethidium and mitoSOX form two fluorescent products, ethidium produced by nonspecific redox reactions, and 2-hydroxyethidium, a specific adduct of O2 •− (98). The fluorescent spectra of ethidium and 2-hydroxyethidium overlap making fluorescence detection nonspecific, therefore, it is necessary to use high performance liquid chromatography analysis of samples containing O2 •− specific products and nonspecific oxidation products such as ethidium to correctly analyze the fluorescence signal (97). Dichlorodihydrofluorescein diacetate (DCFH-DA) is a commonly used oxidative stress marker. Despite the popularity of this assay, it cannot be reliably used to measure intracellular H2O2 and other ROS due to indirect oxidation and redox cycling of 2,7-dichlorodihydrofluorescein (DCF) (41). For these reasons, the editorial board of Free Radicals in Biology and Medicine journal stated that DCFH should not be used.
What Are Hydroxylamine Spin Probes?
The terms “cyclic hydroxylamines” and “hydroxylamine spin probes” are actually used for primary products of stable nitroxides reduction. The hydroxylamine spin probes previously used for ROS measurements are listed in Figure 1.
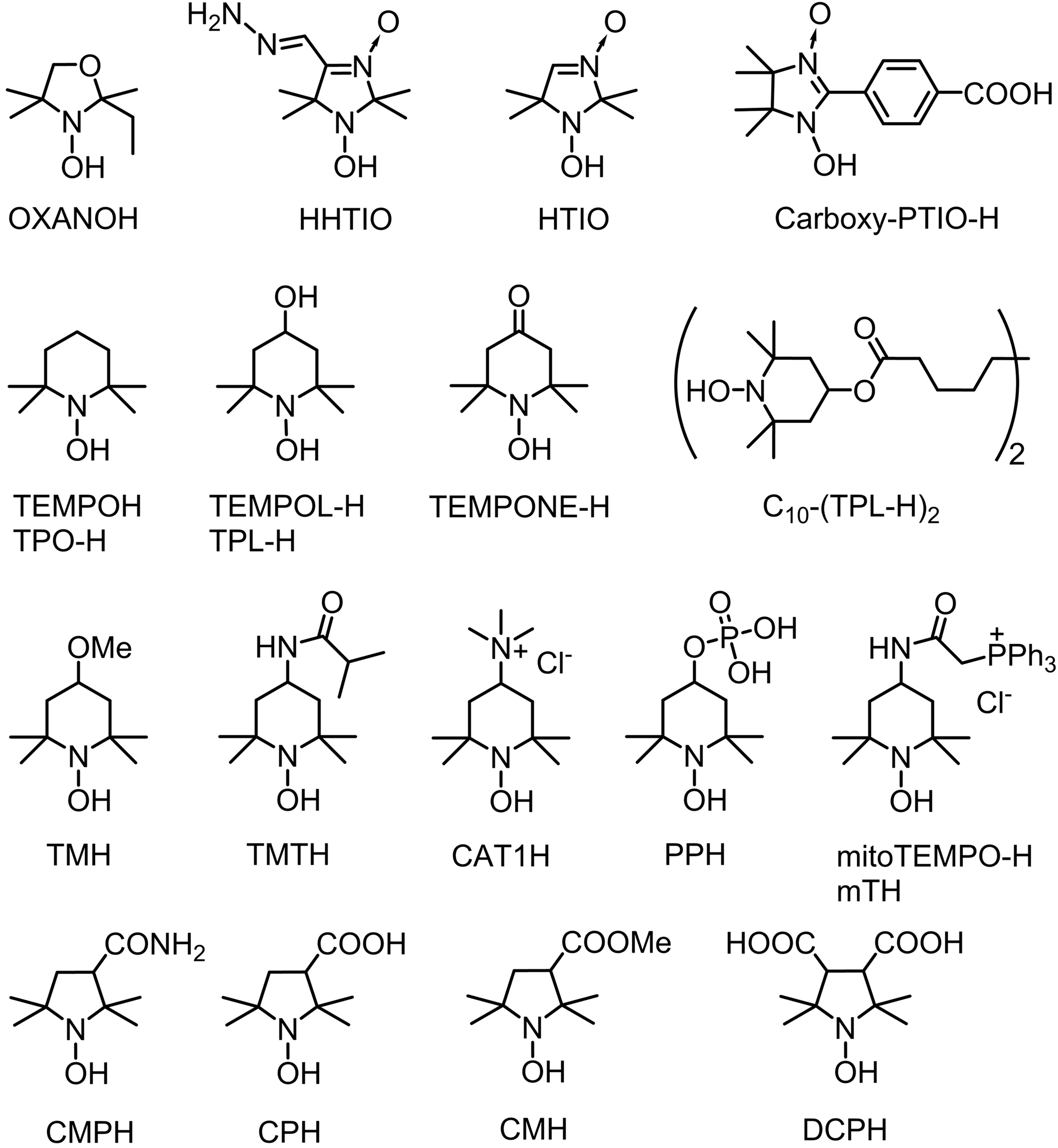
Most of the probes are commercially available or can be prepared from simple commercially available nitroxides (4, 15, 33, 51, 70, 75, 79, 87, 91, 93). Nitroxides are certainly the broadest family of stable free radicals. Overwhelming number of various structures have been synthesized with variable physical and chemical properties. Well-developed chemistry of nitroxides offers great possibilities for design of hydroxylamine spin probes. Hydrolytically stable hydroxylamines of pyrrolidine and piperidine series can form hydrochlorides that are fairly stable crystalline compounds and may be stored for a long time in dry atmosphere. These samples are usually free from background EPR signal of corresponding nitroxide. Both hydroxylamines and corresponding nitroxides demonstrate little or no cytotoxicity; therefore, application of cyclic hydroxylamine spin probes normally produce negligible disturbance to the cellular metabolic processes.
It is important to distinguish spin traps from hydroxylamine spin probes. Hydroxylamines are not spin traps: they cannot bind free radicals. They can undergo oxidation to stable nitroxides, and the nitroxide accumulation can be followed by EPR. Transformation of hydroxylamine to nitroxide requires one-electron oxidation, thus, the reagents, which can accept one electron to give thermodynamically stable products, provide higher oxidation rates. Hydroxylamines rapidly react with oxygen-centered free radicals, including superoxide (Fig. 2). Fast oxidation can occur with some molecular oxidants, such as peroxynitrite, if stable radical species are released in the reaction.
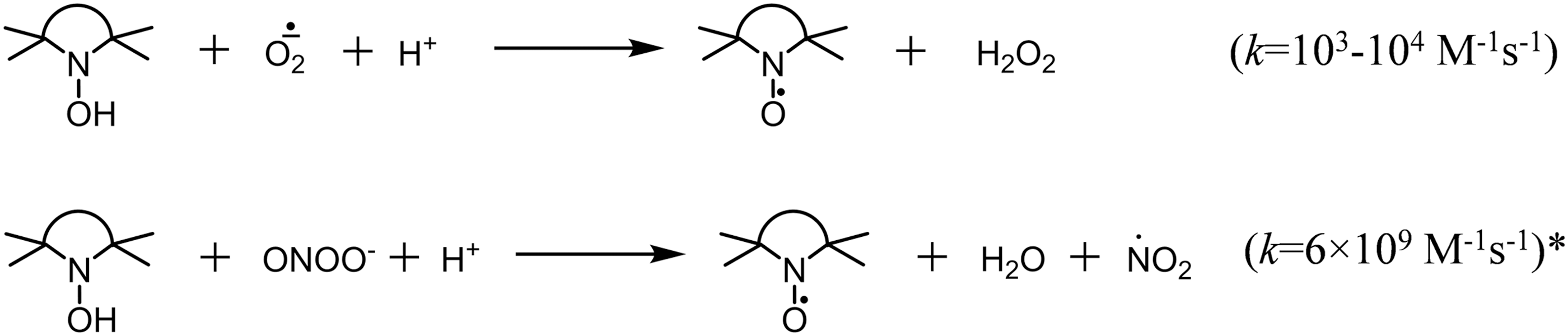
In contrast, reaction with oxygen, H2O2, and organic peroxides is very slow in the absence of mediator capable of one-electron transfer (22, 84). Transition metal ions can increase spontaneous oxidation of hydroxylamines in oxygen-containing solutions (65, 75, 88). To decrease the spontaneous oxidation, stock solutions and buffers should be treated with Chelex and Fe/Cu chelators such as diethylenetriaminepentaacetic acid (DTPA) or ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA) should be added. These reagents should also be added to tissue cellular and subcellular samples where natural iron- and copper-chelating structures are partly removed or diluted.
Reaction of hydroxylamine spin probe with different oxidants leads to accumulation of essentially the same nitroxide. Specific scavengers of certain ROS have to be used to identify particular ROS or particular enzymatic system responsible for nitroxide accumulation since hydroxylamine spin probes neither show specific reactivity to certain ROS type nor allow for EPR spectral identification of different ROS. Specificity of superoxide detection is often confirmed by inhibition of EPR signal with SOD or cell-permeable polyethylene glycol conjugated SOD while peroxynitrite detection can be confirmed by inhibition of EPR signal with urate supplementation (22). Alternatively, specific stimulation or inhibition of ROS-generating enzymes (e.g., Fas ligand for stimulation of NADPH-dependent superoxide production, Nox2 inhibitor gp91ds, or addition/removal of the enzyme substrate can be used to clarify the role of this enzymatic system (65).
Factors Influencing Efficacy of ROS Detection Using Hydroxylamine Spin Probes
Reactivity
Hydroxylamines are bases, for most of them equilibrium is observed between free base and protonated form in neutral solutions. The electron-withdrawing substituents expectedly decrease the pK a of protonated forms, shifting the equilibrium toward the free base. It is likely that hydroxylammonium cations have diminished reactivity with ROS (37). The apparent rate constants of reaction with superoxide and pK a values of some hydroxylamines are listed in Table 1. The data show that many of hydroxylamine spin probes have pK a close to 7 and, therefore, are at least partly converted into nonreactive protonated form at physiological pH. This factor hides other structural effects, such as influence of ring size and electronic effects of substituents upon the rate of oxidation. However, five-membered-ring pyrrolidine hydroxylamines generally are stronger reductants than those of piperidine series and, therefore, they are more disposed to artifacts. For example, 1-hydroxy-3-methoxycarbonyl-2,2,5,5-tetramethylpyrrolidine (CMH) was found to react directly with components of photosynthetic electron transport chain (PETC) to form corresponding nitroxide, whereas 1-hydroxy-4-isobutyramido-2,2,6,6-tetramethylpiperidine (TMTH) did not react (51).
pK a values, rate constants of reaction with superoxide, partition coefficients in octanol–water mixtures (Kp).
CAT1H, 1-hydroxy-2,2,6,6-tetramethylpiperidin-4-yl-trimethylammonium; CMH, 1-hydroxy-3-methoxycarbonyl-2,2,5,5-tetramethylpyrrolidine; CPH, 3-carboxy-1-hydroxy-2,2,5,5-tetramethylpyrrolidine; DCPH, 3,4-dicarboxy-1-hydroxy-2,2,5,5-tetramethylpyrrolidine; mTH, mitoTEMPO-H, 1-hydroxy-4-[2-triphenylphosphonio)-acetamido]-2,2,6,6-tetramethylpiperidine; OXANOH, 2-ethyl-1-hydroxy-2,5,5-trimethyl-3-oxazolidine; PPH, 1-hydroxy-4-phosphono-oxy-2,2,6,6-tetramethylpiperidine; TMH, 1-hydroxy-4-methoxy-2,2,6,6-tetramethylpiperidine; TMTH, 1-hydroxy-4-isobutyramido-2,2,6,6-tetramethylpiperidine.
Nitroxide stability
Efficacy of ROS measurement using hydroxylamine spin probes depends not only on the rate of hydroxylamine accumulation but also on stability of resulting nitroxide. Half-life of some nitroxides in biological samples may exceed hours. The nitroxides are known to undergo bioreduction with low-molecular biogenic antioxidants (ascorbate, glutathione, etc.) and with enzymatic systems, the primary products being hydroxylamines. Factors influencing nitroxide decay rates in biological samples and in vivo have been thoroughly investigated (49). Intracellular reductions proceed faster, and for cell-permeable nitroxides, it makes major contribution to overall nitroxide decay. In relation to hydroxylamine spin probes, this means that intracellular ROS measurement is more difficult. It is hard to compare extracellular and intracellular ROS production even if we use specifically targeted probes. Nitroxides differ much in their ability to reduce. The main structural features determining the reduction rate include ring size (44), charge and electronic effects of the substituents (63), and steric hindrance (39, 47). Sterically shielded nitroxides have never been used for hydroxylamine spin probe preparation yet. Five-membered pyrrolidine nitroxides demonstrate the highest stability to reduction compared with other structural types of nitroxides. For example, 4-isobutyramido-TEMPO (TMT nitroxide) is reduced by components of PETC inside the thylakoid membrane, whereas 3-methoxycarbonyl-proxyl (CM nitroxide) is not (51).
Permeability and targeting
Majority of ROS have limited lifetime and limited ability to diffuse. Therefore, to make the hydroxylamine oxidation efficient, it is important to ensure that spin probe can reach the source of ROS, or even accumulate in tissue or cellular compartment where ROS are generated. Ability of organic substances to diffuse through cellular membranes is mainly determined by partition coefficients (Kp). Kp octanol/water of some hydroxylamines is listed in Table 1. Remarkably, CMH usually gives the most intensive EPR spectra of nitroxide in cellular cultures and in tissues, because it is cell permeable, shows the highest reactivity among pyrrolidine derivatives, and gives highly stable nitroxide of pyrrolidine series.
It is always reasonable to test more than one hydroxylamine for measurements, choosing the probe with optimal redox properties, minimal artifacts, and maximal cellular responses. Specifically targeted hydroxylamines are of special interest as they can provide information about site of ROS origin (22).
Superoxide Measurement
Superoxide radical anion is a product of one-electron reduction of oxygen molecule, which is constantly produced in biochemical redox chains and may be generated by various enzymatic systems. It should be noted that some popular superoxide-producing enzymatic systems, such as xanthine–xanthine oxidase system, produce large amount of H2O2, which can cause secondary processes accompanied with production of free radicals. Therefore, supplementation with catalase is highly desirable. Enzymatic systems usually produce ROS with constant rate; therefore, at high hydroxylamine concentration we observe a linear growth of nitroxide EPR signal, allowing for quantitative measurement of superoxide production rate. Biological function of superoxide is broad. On one hand, it is an important mediator in signaling system regulating cell growth, differentiation, migration, and proliferation (94). On the other hand, it is implicated in cell death (8). Superoxide is often considered an initial source of all biogenic ROS, including highly reactive oxygen-centered radicals (alkoxy, alkylperoxy) and highly reactive molecules (H2O2, peroxynitrite). Unlike majority of oxygen-centered radicals demonstrating close-to-diffusion rates of reaction with various organic compounds, superoxide is a moderately reactive radical. Owing to high biological importance of superoxide, many methods have been suggested for its measurement, including specially designed spin traps and fluorescent dyes.
Superoxide measurement is the main field of application of hydroxylamine spin probes. After invention by Rosen et al. (75), it was successfully used by numerous researchers. Reaction of hydroxylamine spin probes with superoxide is relatively fast (Table 1), two orders of magnitude faster than spin trapping, and the nitroxides formed are more stable than spin adducts. As a result, strong EPR signals are obtained in cells with small amount of cyclic hydroxylamine spin probes (25–500 μM). High reactivity of cyclic hydroxylamine spin probes toward superoxide and high resistance of nitroxides to reduction allows for detection of intracellular superoxide. Sensitivity of this method is high enough for measurements of basal levels of superoxide production by cells.
The oxidation of hydroxylamine spin probes with superoxide can be completely suppressed by SOD. Application of SOD allows for validation of superoxide detection and separation from nonspecific oxidation, making the data on superoxide measurement unambiguous. Indeed, the rate of nitroxide accumulation usually has inverse dependence on SOD supplementation, but it is not affected by scavengers of other ROS, such as urate, catalase, or dimethylsulfoxide (54). Cu, Zn-SOD, and Mn-SOD are cell impermeable; therefore, for validation of intracellular superoxide detection, one can use cell-permeable polyethylene glycol-conjugated SOD (50 U/mL) or use cells overexpressing SOD1 or SOD2 (22).
Superoxide was shown to oxidize nitroxides to oxoammonium cations in the cell-free samples and the rate of this reaction is much higher than that of reaction of hydroxylamine and superoxide (95). Oxoammonium cations are highly reactive strong oxidants, they can oxidize superoxide to form oxygen and nitroxides. In biological media, they usually undergo fast two-electron reduction with various reductants (nicotinamide adenine dinucleotide, reduced form [NADH], thiols, alcohols, amines, etc.) to hydroxylamines. Thus, a redox cycle of interconversions occurs between hydroxylamine, nitroxide, and oxoammonium cation mediated by superoxide and cellular reductants, which accounts for catalytic “SOD-mimetic” activity of nitroxides (hydroxylamines) (49, 53, 95) (Fig. 3). The rate of ROS formation can be underestimated if the ratio nitroxide/hydroxylamine exceeds 1% (19). To perform quantitative measurements, one should have low degree of conversion (dilute cells or increase spin probe concentration) to allow linear time-course accumulation of nitroxide. Direct reduction of nitroxides with cellular reductants can also contribute to the equilibrium position. If the hydroxylamine concentration is not high enough, accumulation of the nitroxide can deviate from expected kinetics (68). In cell-free enzymatic systems and in cultured cells, there are usually no significant deviations from linear kinetics of nitroxide accumulation at conversions <1% (22).

The influence of hydroxylamine structure on the kinetics of nitroxide accumulation strongly depends on the system where the measurement is performed. For instance, piperidine-based hydroxylamine probe CAT1H reacts with superoxide with higher rate than pyrrolidine-based probes, and this leads to faster accumulation of CAT1 in cell-free systems; however, in cell cultures and in tissues, piperidine nitroxides are more rapidly reduced to EPR-silent hydroxylamines compared with pyrrolidine nitroxides, and, therefore, pyrrolidine hydroxylamines provide stronger EPR signals of corresponding nitroxides.
Measurement of ROS in Cell-Free Systems
The method was successfully used for measurement of activities of oxidative enzymes, producing superoxide: xanthine/xanthine oxidase (17), 1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase (84), NADPH-cytochrome P-450 reductase (25, 75), and flavinadenindinucleotide (FAD)-containing monooxygenase (75), for testing antioxidant capacity of preservation solutions (77), etc. It was found that in the absence of a competing scavenger, such as SOD, 3-carboxy-1-hydroxy-2,2,5,5-tetramethylpyrrolidine (CPH) reacts with 90% of superoxide from the membrane fraction (18). Thus, O2 •− detection by CPH should be used in cell-free systems or cellular fractions wherein detection of superoxide can be confirmed by inhibition of the EPR signal with SOD.
Production of superoxide in the presence of nitric oxide leads to formation of peroxynitrite. Hydroxylamines have been successfully used for quantification ofperoxynitrite in various systems, and urate is the efficient reagent for validation of these measurements (17).
It was already mentioned that H2O2 does not react with hydroxylamines with significant rate. However, in presence of H2O2, horse radish peroxidase can oxidize acetamidophenol to corresponding phenoxyl radical, and the latter readily oxidizes hydroxylamines to corresponding nitroxides (3). EPR measurement of the nitroxide formed provides quantitative measurement of H2O2 with detection limit of 3 pmol/test (4). The validation of the method can be performed using catalase. Owing to high rate of reaction of hydroxylamines with acetamidophenoxyl radical, the results of H2O2 measurement are not affected by the presence of NADPH (25–500 μM) or xanthine (25–500 μM) (18). The method can be used for measurement of H2O2 production by cells or mitochondria (18, 26). It should be noted that supplementation of superoxide-generating systems with SOD leads to H2O2 formation and quantification of H2O2 is an alternative to direct superoxide measurements (25, 34). Measurement of H2O2 along with superoxide production in subcellular fractions may give more information about enzymatic activity and function (18).
Since H2O2 is formed in various enzymatic reactions, measurement of H2O2 is a useful approach for measurement of enzymatic activity or substrate concentration. For instance, Nox4 NADPH oxidase activity was measured in membrane fractions of smooth muscle cells (18, 46). H2O2 formation in the enzymatic oxidation of glucose can be used for quantification of glucose. Highly sensitive immunoassays have been developed for measurement of thyroid-stimulating hormone (detection limit 0.0025 μU/mL) (2) and for clinical detection of hepatitis B virus (2, 61).
Measurements of ROS in Cellular and Subcellular Systems
Basal production of superoxide by intact nonstimulated cells and subcellular compartments (e.g., mitochondria) is usually very low; therefore, reliable measurements in cellular systems require good X-band EPR spectrometer with high-sensitivity resonator. Specific stimulation can strongly increase production of ROS, which can be measured by hydroxylamine spin probes. The nitroxide accumulation rate under these conditions depends on cellular location of ROS source and on permeability of the hydroxylamine used.
Cell permeability has been estimated for a set of hydroxylamines, showing the following order: CMH > PPH >> mTH > TMH ≈ TMTH >> CAT1H (22). This row does not show complete correlation with Kp values because of specific accumulation of some hydroxylamines. For example, 1-hydroxy-4-phosphono-oxy-2,2,6,6-tetramethylpiperidine (PPH) can be accumulated in cells via active transport, similar to that of phosphates and was shown to be mitochondria dependent (22); however, PPH does not permeate isolated mitochondria (69). Intracellular and intramitochondrial accumulation of mitoTEMPO-H, 1-hydroxy-4-[2-triphenylphosphonio)-acetamido]-2,2,6,6-tetramethylpiperidine (mTH) is driven by electrical transmembrane potential. Low lipophilicity of CPH most likely results from ionization of the carboxylic group (pK a is ca. 5) at physiological pH values. However, ionization of the carboxylic group only attenuates the cell permeability but does not abolish it completely (22). Difference in cell permeability of various hydroxylamines opens unique possibilities for identification of sources of cellular ROS. For example, site-specific superoxide formation was studied in human lymphoblasts with NADPH oxidase activator phorbol-12-myristate-13-acetate (PMA) or with stimulator of mitochondrial superoxide production antimycin A in the presence of cell-impermeable CAT1H probe and mitochondria-targeted mTH. PMA stimulation leads to nitroxide accumulation, which was five times higher in the presence of CAT1H compared with the presence of mTH. In contrast, stimulation of mitochondrial superoxide production by antimycin A strongly increased oxidation of mTH, whereas nitroxide accumulation with CAT1H probe was not affected by antimycin A (23).
CMH is the most efficient spin probe for measurement of overall ROS production. It allows for ROS detection in various cellular compartments and in different types of cells, usually giving the highest nitroxide accumulation rate (35, 40, 55, 67, 85, 89, 90). However, it also shows high background oxidation rate. Comparison of efficacy of various hydroxylamines in cellular systems and the use of inhibitors to identify ROS can be found in Dikalov et al. (22).
Measurement of ROS in Tissues and In Vivo
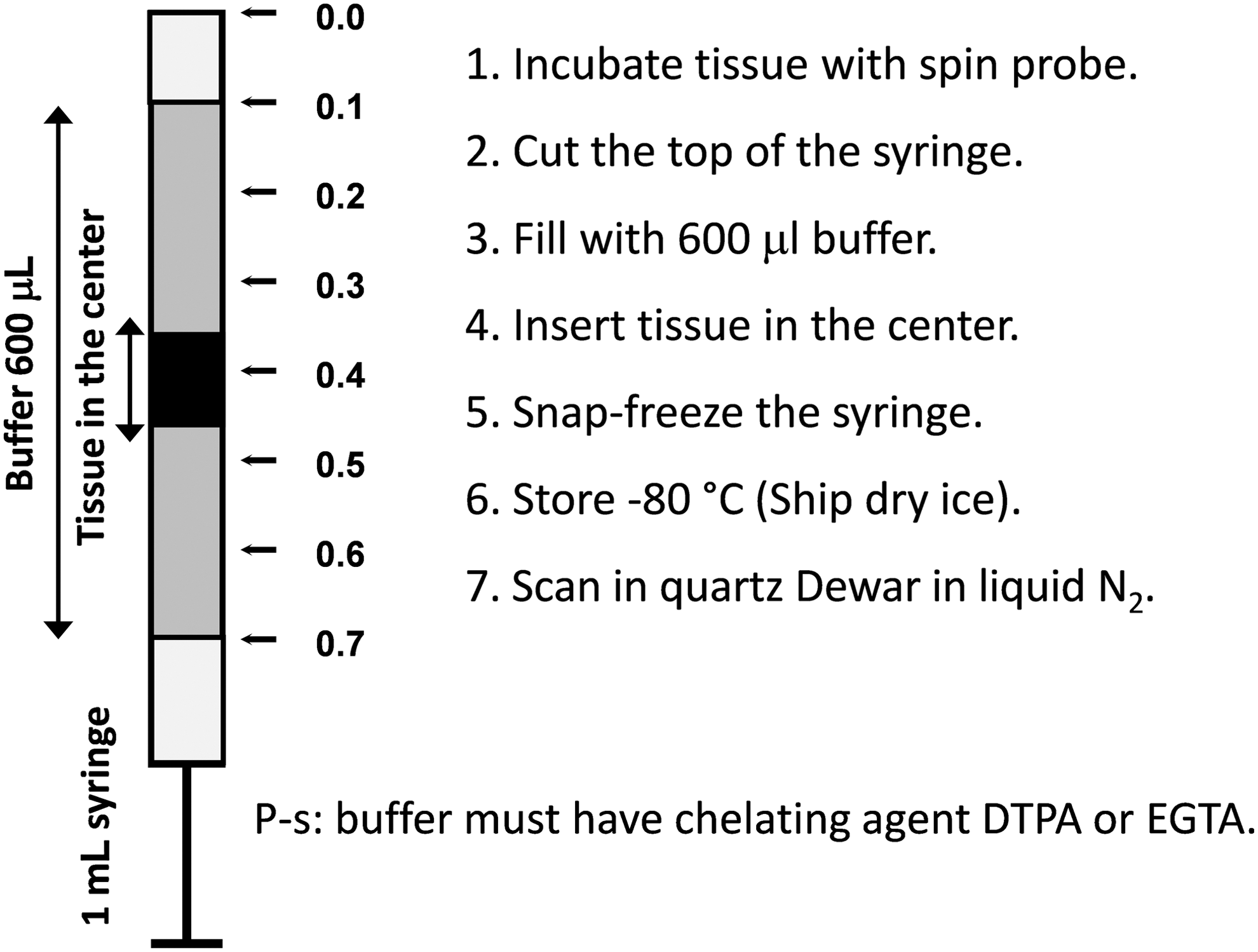
Hydroxylamines have found broad application in investigation of various oxidative stress-related phenomena in living tissues. Superoxide production can be measured by intraperitoneal (i.p.) infusion of cyclic hydroxylamines followed by ex vivo analysis of the blood or tissue samples using X-band (9 GHz) EPR spectroscopy (13, 29, 50). The typical protocols for ROS measurements are given in Dikalov et al. (14) and Fabbri et al. (28). Similar procedures were used in numerous publications (30, 31, 55, 56, 58, 64, 71, 73, 74, 78). Typically, the samples are recorded immediately after incubation with hydroxylamine spin probe (usually CMH or CPH). If the immediate measurements after incubation are not possible, the samples can be frozen. Freezing stops the enzymatic reactions leading to ROS formation; however, after thawing the background oxidation rate may increase (28). To avoid this effect, a special procedure has been developed with registration of immobilized nitroxide spectrum in the frozen sample (1, 7, 6, 12, 14, 57, 81, 86). Specifically, animals can be injected with hydroxylamine probe (20 mg/kg) for 60 min, sacrificed, and isolated tissue can be placed in the center of 1 mL plastic syringe (Fig. 4), snap frozen and stored at −80°C. Then frozen block can be removed by gentle push from the warmed up syringe and analyzed in the quartz Dewar with liquid nitrogen. As a reference one can freeze standard solutions of stable nitroxide product and use the EPR intensity of immobilized nitroxide to calculate the amount of detected ROS in vivo.
It should be noted that these measurements cannot be attributed to certain types of ROS without validation using specific inhibitors or scavengers. It has been shown that supplementation of wild-type mice myocardium samples with polyethylene glycol-conjugated superoxide dismutase (PEG-SOD) decreased amount of nitroxide after incubation with CMH (42). Transgenic animals such as overexpressing SOD2 mice or SOD2-depleted mice (SOD2−/+) can also be used for validation. In cell culture experiments, one can confirm specific site of O2 •− detection by testing the effect of supplementation with cell-impermeable Mn-SOD, cell-permeable PEG-SOD, or overexpression of SOD2 (22).
To provide a cell and tissue-specific measurements of superoxide, we recommend treating samples with hydroxylamine probe ex vivo. In these experiments, blood, isolated cells, or dissected tissue is incubated with the probe (50–500 μM) for 30 min. Tissue samples can be placed at the center of the syringe (Fig. 4). Blood or cell suspension can be placed into the 1 mL precut syringe and snap frozen in liquid nitrogen as shown in Figure 5. After incubation with the probe, cultured cells attached to the bottom of the dish can be quickly resuspended in 600 μL of fresh buffer and placed into the syringe.

Combination of cell-impermeable spin probe (e.g., CAT1H) and a spin probe capable of intracellular/intramitochondrial accumulation may provide information about mechanisms of cellular response development. Recently we have found that simultaneous use of CAT1H-15N, D12, and mTH can reveal peculiarities of various agents known to stimulate phagocytic NADPH oxidase and mitochondrial O2 •− production (20).
ROS Imaging in Animals
Numerous efforts have been made to visualize ROS production in tissues of living animals; however, ROS imaging in vivo still remains a challenge. There are following problems:
Technical aspect
Standard commercial EPR spectrometers operate at 9.5 GHz (0.34 T). This frequency is efficiently absorbed by liquid water in biological systems; therefore, the EPR spectra can only be recorded in ampoules with rather low cross section. Measurements in living animals have to be performed at much lower frequencies: L-band, around 1 GHz, 30–40 mT for objects smaller than 20 mm (mice or isolated organs), or radio frequencies below 300 GHz, 10 mT for larger objects (70 mm, rat or rabbit) (5, 45, 52, 59). Recording of a spectrum is usually not enough for in vivo studies, because nitroxide content in tissues is wide ranging. EPR imaging (EPRI) is a time-consuming procedure, because it requires a set of spectra recorded at different directions of the magnetic field gradient. Thus, there is always a tradeoff between temporal and spatial resolution. Moreover, EPRI does not give any references on anatomy; therefore, to use EPR images one should overlay them with magnetic resonance imaging (5). Mixed technologies, such as overhauser-enhanced magnetic resonance imaging (PEDRI), are currently considered more promising for radicals imaging (5, 62). Most of the advances in this field were made using self-made equipment.
Chemical aspect
Dynamics of nitroxide concentration in tissues upon injection of hydroxylamine reflects pharmacokinetics of both hydroxylamine and nitroxide, their metabolic (redox) transformations, and clearance by excretion. It has been shown that i.p. injection of TEMPOL or TEMPOL-H in mice leads to essentially the same concentration of TEMPOL in blood in <20 min (80). From this point of view, injection of nitroxide or hydroxylamine should lead to the same result. Most of the researchers prefer to follow kinetics nitroxide decay, because this allows to observe signal decay at higher nitroxide concentration and, therefore, requires lower device sensitivity. Another question is whether these measurements actually reflect ROS production in tissues. Peculiarities of pharmacokinetics of hydroxylamine and of corresponding nitroxide can play a major role. For example, higher level of 3-carboxy-2,2,5,5-tetramethylpyrroline-1-oxyl in brain tumors obviously results from blood-brain barrier damage (60). Nitroxide reduction rate was found oxygen dependent, or even determined by redox state of oxygen-consuming enzymatic complexes (10, 49, 83). However, in vivo the nitroxide–hydroxylamine equilibrium is different for different types of cells and tissues and depends on the pathophysiological condition of the living animal (11).
Acyl-protected hydroxylamines (acetoxyamines) represent an alternative to hydroxylamine or nitroxide spin probes (Fig. 6). These compounds themselves do not react with ROS, but they can be converted into reactive hydroxylamine spin probes by intracellular esterases.
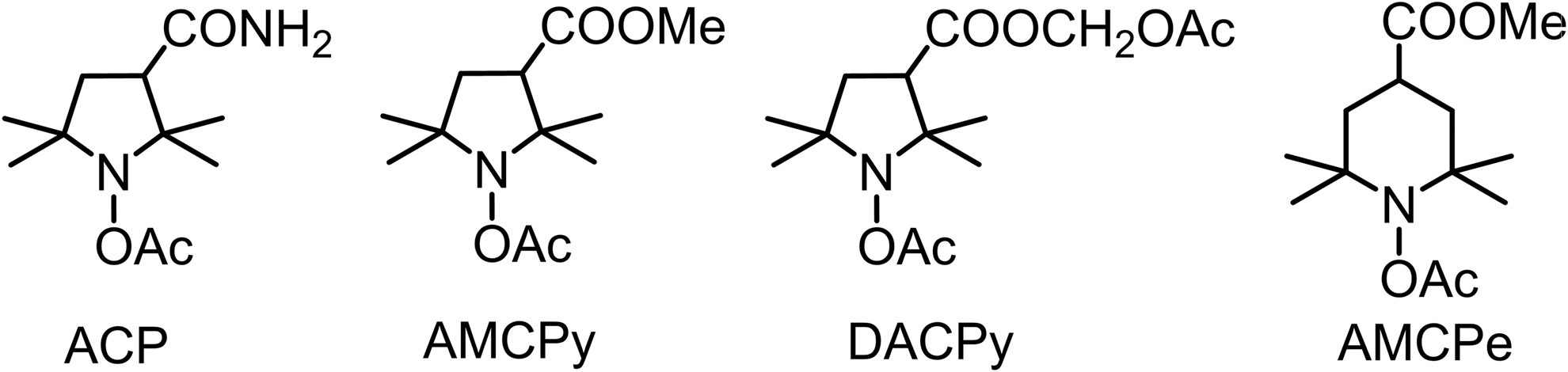
Investigation of the 1-acetoxy-3-carbamoyl-2,2,5,5-tetramethylpyrrolidine (ACP) in cellular system indicated that SOD can suppress >50% of nitroxide accumulation in PMA-treated neutrophils. This means that after deprotection with esterases, hydroxylamine can diffuse out of the cell and this hydroxylamine (and corresponding nitroxide) can redistribute in the system (38). It has been shown that in mice after i.p. or intravenous. injection, ACP quickly reaches equilibrium with the other tissues. Deacylation proceeds very fast within liver and, probably, kidney; the rates of hydrolysis in other tissues were much lower, and resulting hydroxylamine was redistributed in organs. The measurement of hydroxylamine concentration in blood and majority of organs showed almost homogeneous distribution with minor changes in a period 10–30 min after injection, whereas accumulation of the compound in brain and muscle proceeded with slower rate. In 30 min almost all ACP was converted into hydroxylamine, and the nitroxide concentration never exceeded 10–20% of that of hydroxylamine (76).
1-acetoxy-3-methoxycarbonyl-2,2,5,5-tetramethylpyrrolidine and 1-acetoxy-3-acetoxymethoxycarbonyl-2,2,5,5-tetramethylpyrrolidine in mice showed somewhat different pattern of distribution of corresponding hydroxylamine and nitroxide, with the highest concentration of both compounds 10 and 60 min after i.p. injection observed in kidney. AMCP hydroxylamine reached higher concentration in all tissues than DACP, whereas 1-acetoxy-4-methoxycarbonyl-2,2,6,6-tetramethylpiperidine hydroxylamine did not show significant accumulation (92).
ACP was used for rat brain EPRI after kainic acid (KA)-induced epileptic seizures. Difference in accumulation of the nitroxide in different areas of brain in control and KA-treated rats was used to reveal brain structures subjected to stronger oxidative stress (92).
In general, the acyl-protected hydroxylamines do not show apparent advantage over free hydroxylamines because the molecules are not retained by cells, and the products of their hydrolysis (deacylation) and oxidation are redistributed in tissues in agreement with their pharmacokinetics. In this respect, design of acyl-protected hydroxylamine probes with specific targeting groups might be a fruitful approach.
Conclusions
Cyclic hydroxylamine probes are the effective scavengers of superoxide radical. Cyclic hydroxylamines provide quantitative measurements of superoxide radical with high sensitivity both in vitro and ex vivo in cells and tissue samples. SOD-inhibited nitroxide formation can confirm the amount of detected superoxide. Cyclic hydroxylamines can also be used to detect peroxynitrite. Inhibition by peroxynitrite scavengers such as urate shows the amount of trapped peroxynitrite. Production of H2O2 can be measured by co-oxidation of CPH in peroxidase–acetamidophenol reaction. Cyclic hydroxylamines have been successfully used to assay the production of superoxide radical and H2O2 by NADPH oxidases, mitochondria, intact neutrophils, vascular cells, and tissue. To minimize nonspecific oxidation, buffers must be treated with Chelex resin and supplemented with iron and copper chelators (Desferal, DTPA, EGTA). New cell-permeable and acyl-protected cyclic hydroxylamines have a great potential in the study of intracellular ROS production. These new EPR techniques demonstrated great potential to study ROS in biological samples.
Footnotes
Acknowledgments
This work was supported by funding from National Institutes of Health (R01HL124116) and Russian Foundation for Basic Research (17-03-01132-a).