
Abstract
Significance:
Hydrogen sulfide (H2S) at the right concentration is associated with numerous health benefits in experimental organisms, ranging from protection from ischemia/reperfusion injury to life span extension. Given the considerable translation potential, two major strategies have emerged: supplementation of exogenous H2S and modulation of endogenous H2S metabolism.
Recent Advances:
Recently, it was reported that hepatic H2S production capacity is increased in two of the best-characterized mammalian models of life span extension, dietary restriction, and hypopituitary dwarfism, leading to new insights into dietary and hormonal regulation of endogenous H2S production together with broader changes in sulfur amino acid (SAA) metabolism with implications for DNA methylation and redox status.
Critical Issues:
Here, we discuss the role of dietary SAAs and growth hormone (GH)/thyroid hormone (TH) signaling in regulation of endogenous H2S production largely via repression of H2S generating enzymes cystathionine γ-lyase (CGL) and cystathionine β-synthase (CBS) on the level of gene transcription, as well as reciprocal regulation of GH and TH signaling by H2S itself. We also discuss plasticity of CGL and CBS gene expression in response to environmental stimuli and the potential of the microbiome to impact overall H2S levels.
Future Directions:
The relative contribution of increased H2S to health span or lifespan benefits in models of extended longevity remains to be determined, as does the mechanism by which such benefits occur. Nonetheless, our ability to control H2S levels using exogenous H2S donors or by modifying the endogenous H2S production/consumption equilibrium has the potential to improve health and increase “shelf-life” across evolutionary boundaries, including our own. Antioxid. Redox Signal. 28, 1483–1502.
Introduction
H

H2S has historically been associated with toxicity at high levels. Environmental and/or occupational exposure to elevated concentrations of H2S can lead to irritation, respiratory distress, and even death (102). Extreme H2S conditions correlate with mass extinctions throughout geobiological history (92, 94, 123). Toxic levels can also be generated aberrantly in mammals by dysregulation of endogenous H2S metabolism, resulting in a range of pathologies from inflammation (103, 184) to β cell dysfunction and diabetes (167). Mechanistically, H2S toxicity in eukaryotes has been attributed to inhibition of mitochondrial respiration and induction of oxidative stress (31, 79, 86).
Too little H2S also has negative consequences. Mice with genetic defects in endogenous H2S generating enzymes cystathionine γ-lyase (CGL, also known as cystathionase, CTH, or CSE) or cystathionine β-synthase (CBS) are susceptible to hypertension (70, 190), neurodegenerative disorders (127), vascular complications associated with diabetes (167), and symptoms of osteoporosis (110). Because CGL and CBS have functions in addition to H2S production, it is important to note that addition of exogenous H2S corrects many of these problems in rodent knockout models, consistent with a specific role for H2S. In humans, reduced circulating H2S levels correlate with, and may be causally related to, cardiovascular disease (90).
While too much or too little H2S is detrimental, exposure to supraphysiological levels of H2S can be beneficial. For example, in rodents, H2S induces a hypoxia-resistant metabolic state resembling torpor (13), increases vasodilation of blood vessels and lowers blood pressure (197), protects against ischemia/reperfusion injury to multiple organs, including the heart (39) and liver (78), improves insulin sensitivity (188), delays cognitive decline in animal models of Alzheimer's disease (52), and alters many other physiological endpoints with medical relevance (181). In lower organisms including yeast and worms, exogenous H2S extends longevity (65, 114). These benefits of exposure to supraphysiological levels of H2S likely occur through distinct biological mechanisms, including posttranslational modification and functional alteration of cellular targets such as ion channels (110, 118) and growth factor and hormone receptors (171, 196), with many more likely yet to be deciphered.
Given the considerable translational potential of increased H2S in a number of biomedical areas (27, 136, 166), two major strategies have emerged: supplementation of exogenous H2S and modulation of endogenous H2S production. Each approach has unique challenges. Exogenous H2S supplementation is conceptually the most straightforward, with multiple different forms of H2S already available, from the short-acting gas itself (13) or aqueous forms of the H2S salt NaHS (2) to long-acting, slow-release H2S donors (100, 131). Nonetheless, toxicity at high levels and the corrosive and malodorous nature of the gas present challenges (6, 57). H2S is also highly reactive, so that even in orally available, slow-release formulations, targeted delivery to the proper cell type at the optimal dose, once this is fully elucidated, may be challenging to achieve.
The alternate approach to increasing H2S levels is to modulate endogenous production and/or consumption. On the production side, this could include modulation of expression, posttranslational modification, subcellular localization, allosteric regulation, and/or substrate-level control of H2S producing enzymes. Several examples of each of these are known. Acetylcholine and vascular endothelial growth factor both increase H2S production in endothelial cells by activating cognate receptors and increasing calcium/calmodulin-dependent CGL activation (126, 190). In vascular smooth muscle cells, CGL translocates upon endoplasmic reticulum (ER) stress to mitochondria (48), where concentrations of cysteine, a substrate for H2S production, are highest. In the liver, hypoxia increases mitochondrial CBS levels and H2S production by preventing oxygenation of its heme cofactor and thus inhibiting degradation by Lon protease (172). CBS activity is also stimulated by the methionine cycle intermediate S-adenosylmethionine via allosteric regulation (157). Inhibition of phosphodiesterase PDE5 with the drug tadalafil is best known for stimulating nitric oxide production, but also increases H2S production, in part, through cyclic guanosine monophosphate-dependent protein kinase G activation in the heart, which may work to increase CGL transcription via activation of the transcription factor SP1 (145). Tadalafil also activates H2S production in podocytes by increasing CGL protein levels (97). The antidiabetic drug metformin boosts H2S production in several mammalian tissues, although the mechanism remains unknown (185). In conclusion, there is clear evidence that endogenous H2S production is regulated on multiple levels.
On the consumption side, a major mechanism of H2S removal occurs in mitochondria via the sulfide quinone oxidoreductase (SQR), a complex II-like component of the mitochondrial electron transport chain that catalyzes the first step in H2S oxidation to thiosulfate (12, 64, 93). SQR also passes electrons donated by H2S to coenzyme Q, thus promoting proton pumping by the mitochondrial electron transport chain for cellular bioenergetics (48, 74, 93, 111, 117). Like H2S producing enzymes CBS and CGL that are expressed in different cells/tissues (83), SQR displays tissue-specific expression, with high activity in the gut and liver, and low activity in the brain (108). In addition to SQR, sulfur dioxygenase (ethylmalonic encephalopathy 1 protein [ETHE1]) is involved in sulfide oxidation to thiosulfate, which can be readily excreted (64). Because ETHE1 and SQR (as part of the mitochondrial electron transport chain) have direct and indirect oxygen requirements, respectively, H2S consumption and thus steady-state H2S levels can be affected by oxygen tension. Pathophysiological states such as hypoxia can trigger a rapid increase in H2S by modulating both production via CBS stabilization and removal via inhibition of oxygen-dependent detoxification by SQR/ETHE1. Finally, exposure of worms lacking SQR to H2S results in inhibition of protein translation via a mechanism involving phosphorylation and inactivation of the translational initiation factor eIF2α by the amino acid sensing kinase general control nondepressible 2 (GCN2) (68).
While many studies report benefits of exogenous H2S, including ongoing clinical trials of the H2S donor and polysulthionate SG1002 in the context of cardiomyopathy (89), precision manipulation of endogenous H2S levels is currently hampered by our limited knowledge of mechanisms controlling production and consumption, thus representing a major roadblock to progress on this complementary front.
Recently, the finding that hepatic H2S production capacity is increased in two of the best-characterized mammalian models of life span extension, dietary restriction (DR) (65) and hypopituitary dwarfism (66), has led to new insights into regulation of endogenous H2S levels. In this review, we summarize the role of sulfur amino acids (SAA) and endocrine signaling in control of endogenous H2S production, focusing mostly on the production side via regulation of expression of CGL. We discuss the importance of H2S in these longevity models, including the potential role of H2S as an intermediate in nutrient and endocrine/paracrine feedback control. In addition, we demonstrate the plasticity of CBS and CGL gene transcription in response to numerous stimuli, including diet, hormones, drugs, radiation, and disease using the NCBI GEO Profiles database. Finally, we discuss the potential impact of H2S on longevity extension.
Measuring Sulfide Levels
Before discussing the regulation of endogenous H2S production, it is important to consider the techniques used to measure H2S in biological samples and their limitations. The steady-state H2S level present in a cell, organ, or extracellular fluid is a function of enzymatic H2S production and consumption (Fig. 2). At neutral pH, H2S, with a pKa1 of ∼6.9, is in equilibrium with HS- at an ∼1:1 ratio. This is important because H2S, but not HS-, can pass readily through cell membranes into extracellular fluids and evaporates easily from aqueous solution, and thus can be lost (or trapped for later measurement if care is taken) in the gas headspace above a biological sample. It also allows the use of pH to maintain sulfide in solution (HS- at basic pH) or to release it into the gas phase (H2S at mildly acidic pH). The term sulfide is used heretofore when referring to both H2S and HS- species.
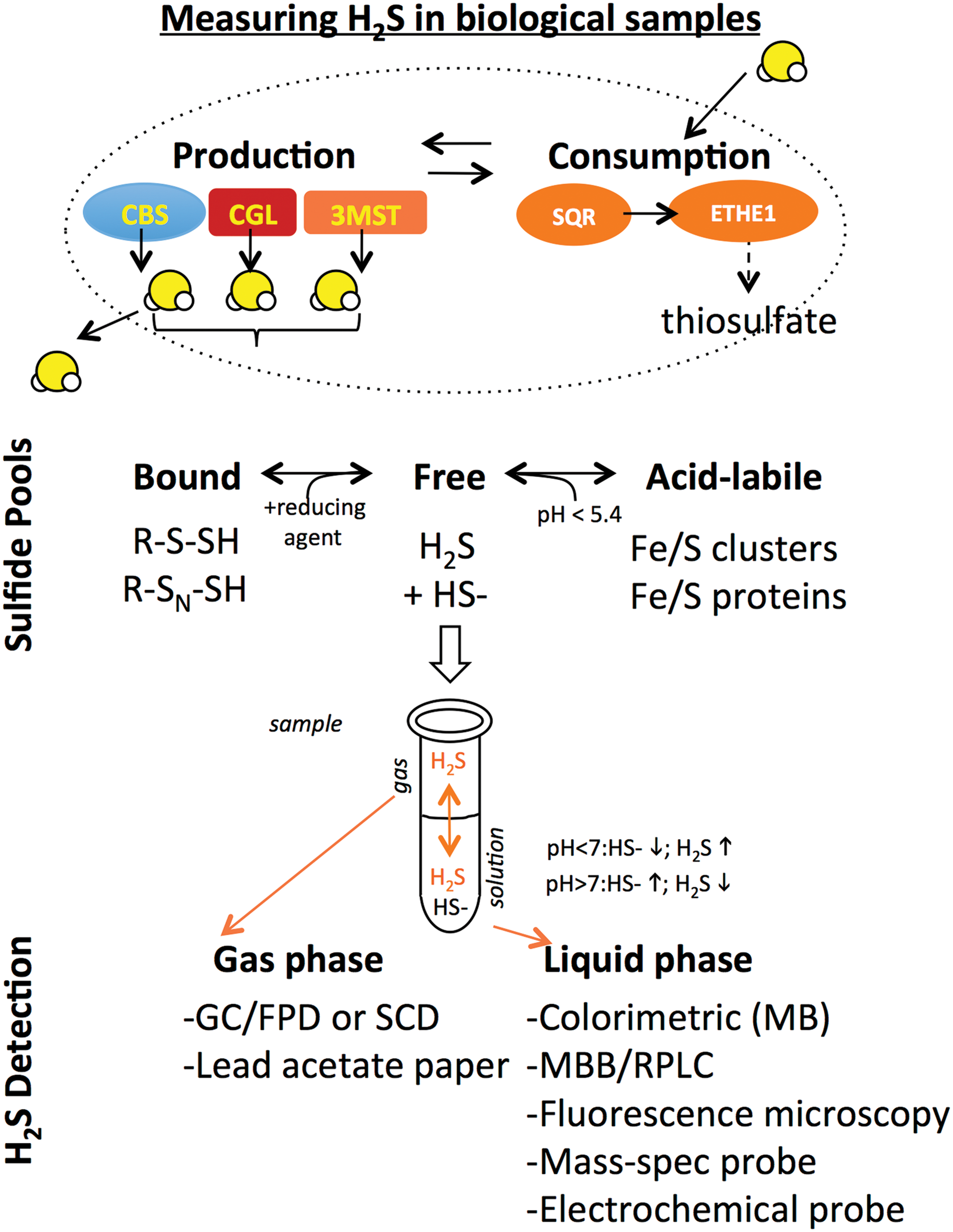
Sulfide present in biological samples exists in different pools that can be distinguished based on their biochemical properties. In addition to free sulfide, H2S can exist as sulfane sulfur, a pool that is functionally defined based on the liberation of free H2S in the presence of a reducing agent such as dithiothreitol or tris(2-carboxyethyl) phosphine. Sulfur present in iron/sulfur clusters can also give rise to free H2S under acidic conditions (pH <5.4), as can nonenzymatic reduction of molecular sulfur, however, the contribution of these sources to free H2S pools under physiological conditions remains unclear.
Techniques to measure absolute concentrations of sulfide present in biological samples have been reviewed extensively (61, 71, 119, 122, 170, 181). They can be grouped into two major categories based on whether the sulfide being measured is present in liquid or gas phases. H2S gas trapped/released into the headspace above its source can be measured by gas chromatography followed by flame photometric detection or sulfur chemiluminescence detection (177). Measuring sulfide in the liquid phase often requires prior derivatization to stabilize sulfide, for example, in the methylene blue method in which lead acetate reacts with sulfide to form lead sulfide, and then, in the presence of N,N-dimethyl-p-phenylenediamine under acidic conditions forms methylene blue, which can be detected by absorbance in a spectrophotometer with a lower detection limit of 1 μM (72, 161). Monobromobimane also reacts with and stabilizes sulfide in solution, which can then be detected by its fluorescence after reverse-phase liquid chromatography with a lower detection limit in the nM range (152). More recently, a number of fluorescent dyes have been reported that react specifically with H2S in vitro and in vivo, as well as a mass spectrometry-based probe targeted to mitochondria that can be used in vivo (4). Finally, electrochemical polarographic (122) or amperometric (190) probes can be used in vivo to directly measure changes in steady-state levels over time in response to perturbations such as hypoxia without sulfide derivatization.
Depending on the experimental design, sulfide measurements can report on either maximal production capacity or actual steady-state levels. Measurements of maximal production capacity in the presence of excess substrate and cofactor, typically done in cell-free lysates/extracts, utilize a variety of techniques to measure H2S in liquid or gas phase as described above. Because such measurements generate much more H2S than is present under physiological conditions, they can be performed in relatively high throughput using a variety of detection methods (e.g., lead acetate). They also lend themselves to the use of enzyme inhibitors (e.g., propargylglycine [PAG] for CGL; aminooxyacetic acid for CGL, CBS, or cysteine aminotransferase [CAT]; aspartate for CAT) and substrates that can help dissect the nature of the H2S-producing activity. A major disadvantage is that they do not report on the actual amount of H2S being generated in the sample in vivo. This is typically done in fresh or frozen samples from cells, tissues, or organs in which sulfide generation and consumption pathways are intact, and substrates for sulfide production present at physiological levels. After extraction and derivatization, sulfide is measured using a variety of techniques appropriate for either solution- or gas-based sulfide measurements. This also allows for measurement of different sulfide pools (bound; acid-labile) following appropriate extraction. A limited number of techniques that permit rapid detection of free sulfide in living biological samples (e.g., sulfide amperometric electrodes or polarography in circulating blood) further allow measurement of changes in steady-state free sulfide levels, for example, upon hypoxia. Nonetheless, quantitative assessment of intracellular H2S levels particularly in vivo remains challenging for a host of reasons, including the volatile and highly reactive nature of the molecule and interference from other thiol-containing molecules (121, 122).
DR and Nutrient-Based Regulation of Endogenous Transsulfuration Pathway and H2S Production
DR describes a variety of nutritional interventions in which reduced total calorie or nutrient intake is associated with health benefits, generally including life span extension in experimental organisms. DR also consistently protects against diseases for which age is the prime risk factor, including cancer, neurodegeneration, inflammatory/autoimmune disorders, and cardiovascular disease. DR has pleiotropic beneficial effects on multiple physiological systems, from glucose homeostasis to inflammation to oxidative stress resistance, some or all of which may contribute to its longevity and health effects (46, 160).
Experimental DR regimens vary widely. Classic calorie restriction (CR) regimens in rodents involve enforced restriction of total food intake by 20–60%. CR is also sometimes referred to as intermittent fasting (IF) because animals quickly eat their allotment of food and thus spend the remaining time between meals in a fasted state. Animals on CR/IF regimens are typically fed the same diet as the ad libitum fed controls and thus at a fixed ratio of calories from fat, carbohydrates, and protein, making it difficult to separate the effects of nutrient restriction from those of reduced energy intake. Nonetheless, adding back essential amino acids (EAA) to food restricted flies abrogates longevity benefits (53), likely through effects on nutrient signaling pathways independent of their calorie content.
Other DR regimens that restrict (or dilute) specific nutrients, including protein or EAAs, without periods of enforced food restriction can also extend life span and health span across evolutionary boundaries (99, 159). Restriction of the EAA methionine (methionine restriction [MR]) is one of the best-studied examples in which restriction of an individual EAA can alter physiology and extend longevity. MR consistently extends life span in multiple experimental organisms, including yeast, worms, flies, rodents, and human cells in culture (18, 23, 80, 91, 96, 124, 143). Classical MR regimens in rodents, in which cysteine is absent due to its methionine sparing effects (a better name would thus be “sulfur amino acid restriction”), result in a 42% increase in mean and 44% increase in maximum life span (141), as well as improvements in stress resistance and metabolic fitness (1, 26, 101, 163). Restriction of the EAA tryptophan can also extend life span in rodents (147), while restriction of branched chain amino acids can have metabolic benefits, including improved glucose homeostasis and body composition (43).
Multiple molecular mechanisms have been proposed to account for DR benefits. These nonmutually exclusive pathways include reduced anabolic signal transduction via insulin/insulin-like growth factor-1 (IGF-1) and mechanistic target of rapamycin (mTOR); activation of adaptive redox response pathways; improved metabolic function and nutrient utilization; increased autophagy; and an overall reduction in macromolecular damage from reactive oxygen and nitrogen species (45, 156). Nonetheless, the relative contribution of such molecular, metabolic, and physiological changes to longevity benefits remains poorly understood, as do the relationships between CR, MR, EAA restriction, and other DR regimens.
Recently, we reported that H2S production capacity is increased in multiple experimental models of DR and associated with multiple DR benefits, adding it to the list of potential molecular mechanisms underlying DR action (65, 67). In mice, we found that 1 week of 50% CR increases endogenous hepatic H2S production and improves resistance to the acute multifactorial stress of hepatic ischemia/reperfusion injury. In this model, H2S production and stress resistance correlate with increased CGL expression on the mRNA and protein levels, are prevented by pharmacological inhibition of CGL with the chemical inhibitor PAG or by genetic ablation of CGL, and augmented by CGL overexpression independent of diet. Interestingly, constitutive activation of mTOR in hepatocytes blocks DR-mediated hepatic stress resistance (60) and simultaneously suppresses CGL expression and H2S production capacity in the liver. In worms, flies, and yeast, H2S production capacity is augmented by organism-appropriate DR regimens, including genetic models of reduced food intake in worms (eat 2), MR in flies, and reduced glucose in yeast (65).
While this study was the first to explicitly link augmented endogenous H2S to pleiotropic DR benefits, it is consistent with a number of findings regarding age-related changes in H2S and expression of H2S-producing enzymes in the context of longevity and DR. In fruit flies, life span extension via DR correlates with increased expression and activity of CBS in whole fly homogenates and is blocked by knockdown of CBS or inhibition of CGL with PAG, while either total body or neuron-specific overexpression of CBS is sufficient to extend life span (81). In rats, there is an aging-related decline in CGL and CBS expression and H2S production in multiple tissues, and this is abolished by long-term 10–40% CR from 8 to 38 months of age (135). Similarly, 30% CR for 6 months in rats attenuates renal aging, boosts CGL and CBS expression, and increases H2S production in the kidneys (182). Even short-term DR in rats, in the form of SAA restriction for 1 week, results in increased hepatic CGL expression (155). In mice, SAA metabolic pathways are significantly affected on the transcriptional level by DR in multiple tissues (165). Twenty to forty percent CR results in a dose-dependent increase in hepatic H2S production in both male and female mice of different strain backgrounds, correlating with improved health, although intriguingly, not always with extended longevity (116). Caloric restriction between 0% and 40% for 3 months in male mice increases hepatic CGL expression in a dose-dependent manner (35), while SAA restriction for 5 weeks elevates hepatic CGL and CBS expression (130).
How does diet influence H2S generation, and by what mechanism is H2S increased on DR? One mechanism of increased H2S production is by transcriptional regulation of CBS and CGL. Regulation of these genes is best understood in the context of de novo cysteine biogenesis via the transsulfuration pathway (TSP), the presence of which explains why cysteine is considered a nonEAA. The first step is the condensation of homocysteine with serine to form cystathionine by CBS. Cystathionine is then hydrolyzed to cysteine (and α-ketobutyrate + ammonia) via CGL (Fig. 3). When cysteine levels are low, CGL is upregulated at the mRNA and/or protein level via increased protein expression of activating transcription factor 4 (ATF4) (98, 154, 155), which binds to sequences in the first intron of CGL to activate transcription under stress conditions, including amino acid deprivation (115). In liver-derived cells, cysteine deprivation induces ATF4 expression via the amino acid deprivation sensor GCN2 (98), a kinase that senses uncharged tRNAs and controls translation via phosphorylation of the translation initiation factor eIF2α (50). Cysteine can then be used in protein, glutathione, or taurine biosynthesis. Mouse embryonic fibroblasts lacking ATF4 show little or no CGL transcript or protein (36) and require the addition of cysteine or other antioxidants for growth (58). ATF4 expression is increased on CR or SAA restriction (105, 155), and likely plays a role in increased H2S production, in part, through transcriptional control of the H2S generating enzyme CGL in the liver and potentially other cells/organs (65, 130, 146), although the formal genetic requirement remains to be tested.
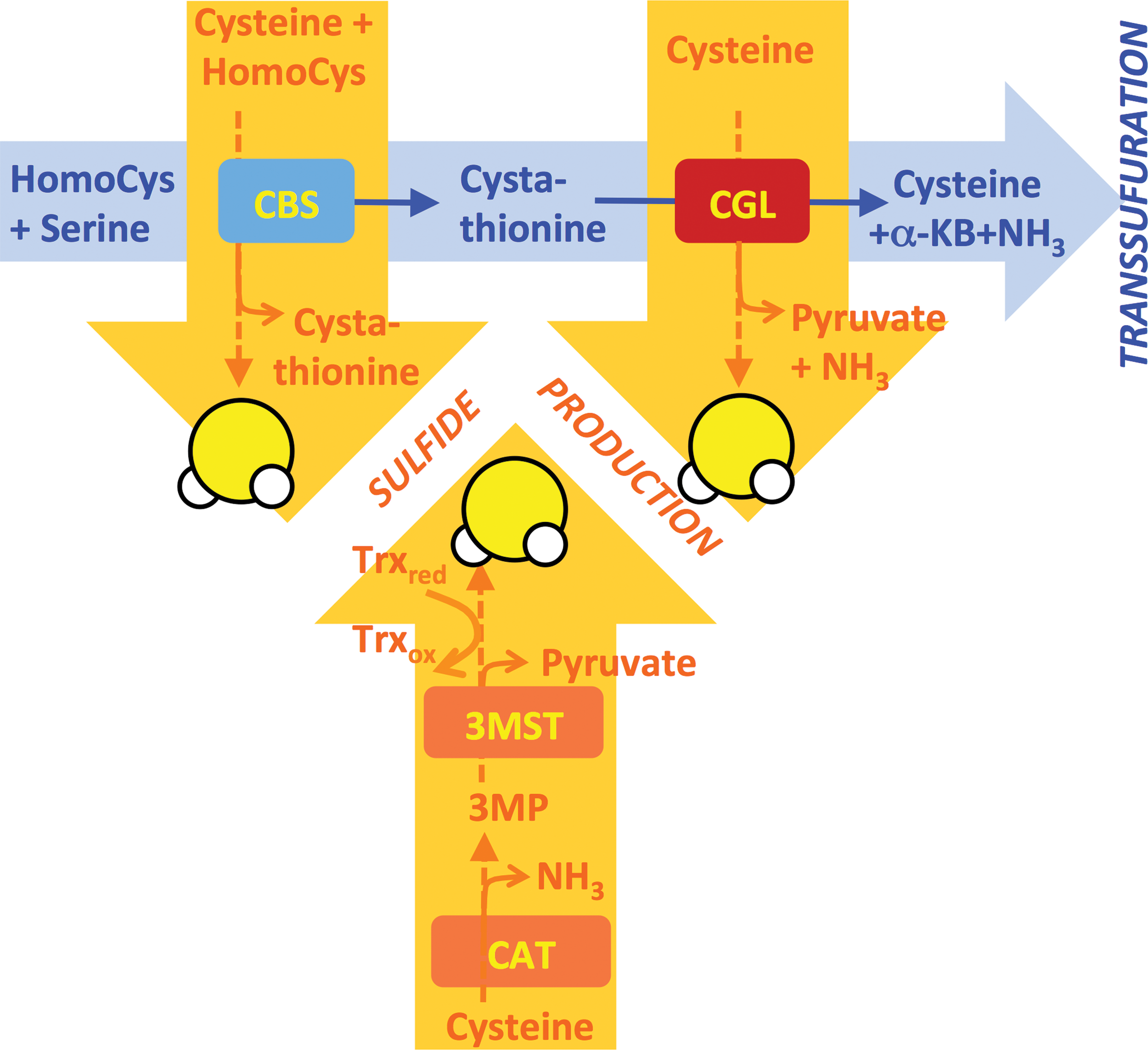
It is important to note that while CBS and CGL are both major H2S producers, H2S production is not a by-product of de novo cysteine biogenesis via the TSP. Rather, H2S is produced by a different series of reactions catalyzed by the same proteins with overlapping substrates and products (Fig. 3). CBS produces H2S from homocysteine and cysteine, and is the predominant H2S-producing enzyme in the kidney and brain under normal physiological substrate concentrations (28). In the liver, H2S production by CGL dominates (29), in part, due to CGL protein levels being 60-fold higher than CBS (83). While the catalytic efficiency for H2S production by CGL is higher with homocysteine than cysteine, under normal physiological conditions in the liver, cysteine is the predominant substrate for H2S production (29). Importantly, the catalytic efficiency for cysteine production from cystathionine by CGL is 30 times higher than H2S generation from cysteine (29). In other words, conditions favoring de novo cysteine biogenesis via TSP are different from those favoring H2S production from cysteine, consistent with TSP and H2S production being separable processes. Whether these two processes are mutually exclusive within a given cell, or can occur at the same time in different subcellular compartments, remains to be elucidated.
What is the relationship between TSP and H2S production, and does an increase in one necessarily affect the other? In other words, if DR increases TSP activity to replenish cysteine levels, increased cysteine utilization for H2S generation presents an apparent paradox. In fact, while the substrate preferences for H2S generation by CGL and CBS are described (157), the sources of free cysteine (or homocysteine) for H2S production are not known. Thus, one potential answer to this apparent paradox is that cysteine for protein translation or glutathione biosynthesis produced de novo by the TSP is in a distinct cellular pool or subcellular compartment than free cysteine fueling H2S production. Substrate for H2S generation may instead come from lysosomes via autophagy or via proteolysis, both of which are expected to increase on DR. If cysteine pools are distinct, it may be possible for cysteine generated de novo from TSP to drive glutathione and/or taurine biosynthesis in the same cell at the same time as H2S production is increased. Future studies are required to shed light on metabolic flux through TSP and H2S biosynthetic pathways as a function of diet, including DR.
Contrary to DR, dietary interventions such as high-fat diets (HFD), which can precipitate obesity, metabolic syndrome, and shortened life expectancy, have opposite effects on TSP gene expression and H2S production capacity. For example, mice fed a HFD for up to 16 weeks exhibit reduced CGL expression in the liver, lung, and aortic endothelium, with diminished H2S biosynthesis in liver, kidney, and lung tissues (129). Rats fed HFD for 18 weeks exhibit increased plasma homocysteine concentrations and decreased hepatic enzymatic activities of CBS and CGL (17). Rats fed a Western diet (21% fat) versus control chow (6% fat) for 12 weeks display decreased CGL expression and H2S in aortic tissue, and this correlates with decreased endothelial function and increased reactive oxygen species in this tissue (76).
Exercise appears to reverse some of the negative impacts of HFD feeding on TSP activity and H2S production. For example, in addition to improvements in body weight, hepatic steatosis, and glucose homeostasis, exercise restores CBS and CGL expression in the liver and H2S bioavailability in the plasma and liver in mice fed HFD for 24 weeks (180). Exogenous H2S reduces lipid accumulation, injury, and fibrosis in the kidneys of mice on HFD (186), suggesting that H2S itself may contribute to these benefits.
In addition to effects of macronutrient intake, dietary micronutrients can also affect TSP activity and H2S production. In Dahl rats, a high-salt diet induces hypertension and diminishes renal CBS expression and endogenous H2S content, while supplementation with H2S reverses effects on hypertension and aortic remodeling (70). Chronic intake of the artificial sweetener aspartame for 90 days in mice reduces hepatic TSP metabolites cysteine, S-adenosylmethionine, and S-adenosylhomocysteine, as well as CGL on the mRNA and protein levels (40).
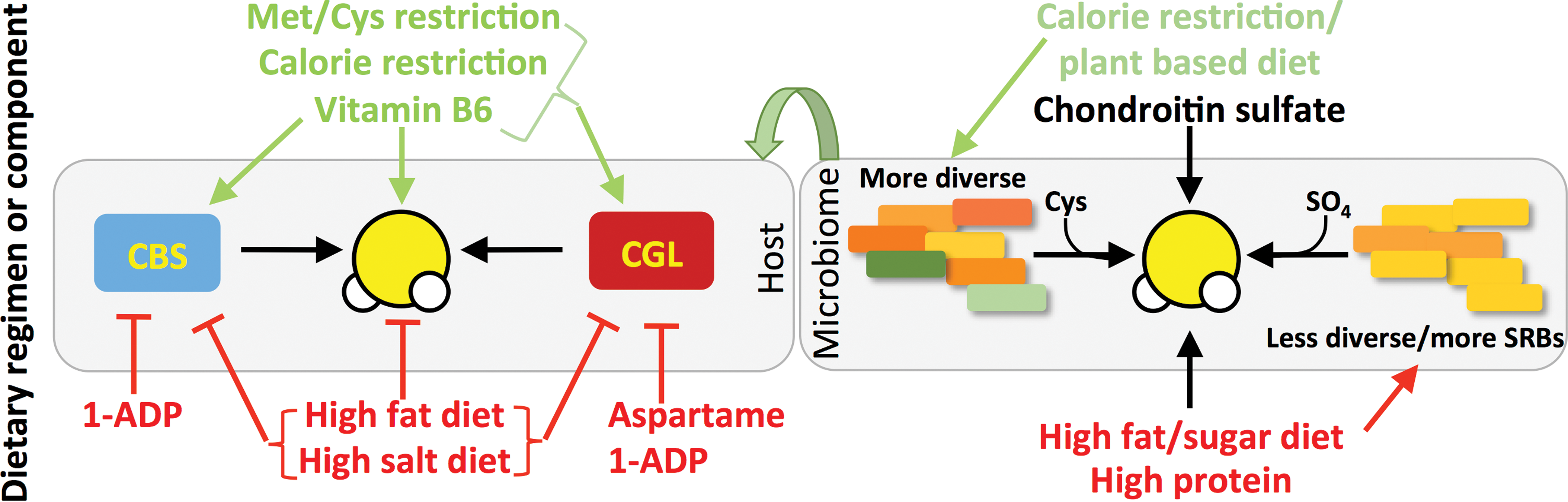
The bioactive form of vitamin B6, pyridoxal phosphate (PLP), is required as a co-factor for CGL and CBS enzymatic activities, with CGL displaying greater sensitivity to PLP deficiency than CBS. As a segment of the U.S. population is considered PLP insufficient (10–12%) or slightly insufficient (10–15%) (54), it brings to light the possibility that dietary PLP, or the lack thereof, can affect human health via control of endogenous H2S production. Interestingly, the metabolite 1-amino D-proline (1ADP), which is found in the common dietary foodstuff flaxseed, is a PLP antagonist, and rats fed a diet with 10 mg 1ADP/kg diet display reduced enzymatic activities of CBS and CGL (113). A summary of dietary effects on TSP and endogenous H2S production is presented in Figure 4.
Finally, H2S can also be generated on cysteine oxidation via 3-mercaptopyruvate sulfurtransferase (3-MST), which can produce H2S in the presence of a reducing agent from 3-mercaptopyruvate following deamination of cysteine by CAT (153). CAT (but not 3-MST) is a PLP-dependent enzyme like CBS and CGL, however, little is known about dietary regulation of H2S production by this cysteine oxidation pathway.
Although the focus up to this point has been primarily on the interaction of diet and endogenous H2S production in cells, tissues, and organs of eukaryotic cells, bacteria constituting our gut microbiome also produce H2S. Sulfate-reducing bacteria (SRB) such as those from the Desulfovibrio genus use inorganic sulfate as a terminal electron acceptor for respiration, thus generating H2S (25). A wide range of bacteria can also generate H2S via degradation of organic sulfur in the form of cysteine through cysteine desulfhydrase activity present in homologues of eukaryotic CBS, CGL, and CAT/3-MST enzymes. Interestingly, H2S generation by bacteria protects them from antibiotic-induced oxidative damage (150). The presence of high levels of SRB or other H2S producing bacteria such as Fusobacterium is associated with proinflammatory disorders, including inflammatory bowel disease (both ulcerative colitis and Crohn's disease), colorectal cancer, and irritable bowel syndrome (25, 158). Nonetheless, H2S can also protect gut barrier function through modulation of the host inflammatory response in a rodent model of colitis (179), and thus, it remains unclear whether elevated H2S associated with inflammation in the gut is causative of disease or merely an epiphenomenon.
Diet can influence both the composition of the microbiome and its propensity to produce H2S. Individuals with low bacterial richness and diversity have an increased potential for H2S formation from SRB, and are also more likely to be obese and insulin resistant, gain more weight over time, and have increased inflammatory phenotypes (95). Rodents with symptoms of metabolic syndrome induced by HFD also show higher levels of SRB in gut microbiota (194). Mice fed a diet high in fat and simple sugars, as well as the dietary supplement chondroitin sulfate, are associated with increased cecal levels of Desulfovibrio and H2S production, but without compromised gut barrier function (140). Diets high in animal protein are also associated with increased fecal H2S concentrations independent of fat and sugar intake (112). Interestingly, humans on a calorie-restricted (50% fewer calories than their control counterparts), mostly plant-based, diet have greater phylogenic diversity in their fecal microbial populations than controls (55), although the levels of H2S are yet to be established.
Host responses to changes in gut microbiome-derived H2S are also beginning to be characterized. Induction of gut bacterial H2S production via high-protein diet feeding results in an increase in SQR gene expression in host colonocytes (12). Eukaryotic cells, and in particular colonocytes, have the ability to utilize H2S as an electron donor to fuel ATP production via this mitochondrial electron transport chain protein (93). Thus, the increase in SQR in response to augmented bacterial H2S production is indicative of a host adaptive response to either detoxify H2S or utilize it as an inorganic fuel source for ATP production. In gnotobiotic mice lacking a microbiome, H2S levels are reduced in plasma, adipose, and lung tissues, and CGL activity is reduced in many organs (151) consistent with bidirectional effects of microbiome and host on sulfide metabolism (Fig. 4). Future studies should thus consider the effects of dietary macro- and micronutrient composition on both host and microbiome sulfide metabolism and TSP activity.
Long-Lived Hypopituitary Dwarfism and Growth Hormone/Thyroid Hormone Regulation of Endogenous TSP-Dependent H2S Production Capacity
Disruption of the hypothalamic–pituitary axis signaling via spontaneous or induced mutations results in dwarfism, but also extreme longevity, increased stress resistance and improved metabolic fitness in a number of rodent models (10, 149), including genetic deficiency in growth hormone releasing hormone (GHRH) (164) or its receptor (growth hormone releasing hormone receptor, aka little mouse) (42); the anterior pituitary developmental transcription factors Pit1 (Snell dwarf) (42) or Prop1 (Ames dwarf) (19) or the receptor for growth hormone (GHR) (32, 33). In addition, there is a significant inverse correlation in both small and large mammals between thyroid hormone (TH) levels and life expectancy (16). Hence, states that allow for decreased growth hormone (GH) and TH signaling offer prolongevity and antiaging benefits.
In humans, polymorphisms in the thyroid gland receptor for thyroid-stimulating hormone (TSH) resulting in increased TSH but decreased TH production and signaling are associated with extreme longevity in centenarian studies (7, 8, 75, 142). Furthermore, humans with defective GHR signaling (Laron dwarfism), while not apparently longer lived, are resistant to several aging-related diseases, including type 2 diabetes and cancer (56). Conversely, increased TH signaling through active triiodothyronine (T3) binding to the TH receptor (TRs) beta isoform results in DNA damage and premature senescence, two major drivers of aging and aging-related pathologies (193). Likewise, transgenic overexpression of GH in rodents results in large size, hyperinsulinemia, insulin resistance, and decreased life expectancy (11).
What is the mechanistic relationship between longevity extension by DR and hypopituitary dwarfism? Reduced GH/TH signaling leads to CR/MR-like alterations in SAA and energy metabolism without altering food intake (20), while DR alters hypothalamic activation of the pituitary gland (34, 41) and lowers GH (47) and TH signaling (44). This could be due, in part, to a reduction in DR in the adipose-derived hormone leptin, which is implicated in control of hypothalamic thyrotropin-releasing hormone (TRH) levels during fasting via melanocortin and neuropeptide Y signaling (176). Protein restriction decreases GH release from the pituitary and blunts pituitary responsiveness to GHRH (59). Fasting also directly targets the liver and alters the metabolism and bioavailability of TH (77, 187) as well as expression of GHR (162).
Functional interaction studies indicate that benefits of CR or MR on longevity and metabolic fitness are dampened or abrogated entirely in hypopituitary or growth hormone receptor knockout (GHRKO) dwarf mice (5, 14, 15, 22). Conversely, GH supplementation concurrent with DR reverses several of the metabolic effects of DR (30, 51). Together, these data suggest that the mechanisms of DR underlying increased life span and health span work primarily, or at least significantly, through modulation of the hypothalamic–pituitary axis and GH production.
Recently, we identified increased H2S production as another commonality between DR and hypopituitary dwarfism. Hepatic CGL expression and H2S production are elevated in long-lived genetic mouse models of reduced GH and/or TH signaling in vivo and in cultured cells on serum withdrawal, a model of reduced growth factor signaling in vitro (66). Negative regulation of hepatic H2S production by GH and TH is additive and occurs via distinct mechanisms. TH acts primarily via TRs in the liver to repress CBS and CGL gene expression, while GH acts via GHR signaling to control substrate-level H2S production, likely via negative regulation of autophagy. As discussed above, autophagy is a potential source of free cysteine for H2S generation independent of de novo cysteine biogenesis via the TSP. Hepatic H2S production is also increased in long-lived mouse models overexpressing FGF21 (195) or lacking IRS-1 (148). FGF21 regulates both GHR (73) and TH (37) signaling, and is induced on DR (101). IRS-1 is a docking protein that facilitates insulin and IGF-1 signaling, but is also implicated in GHR signaling (106).
Similar to our previous study linking H2S to DR, this was the first study to explicitly associate GH and TH signaling with regulation of endogenous H2S production in the liver. While any causality between health benefits observed in dwarf mice and increased H2S production remains to be established, regulation of H2S production by GH and TH is consistent with a larger body of work linking these hormones to a broader regulation of SAA metabolism, including TSP and glutathione metabolism, as described below.
The connection between GH and TH on SAA metabolism was first observed in hypophysectomized or thyroidectomized/radioiodine ablated rats, in which changes in activities of TSP and transmethylation pathway (TMP) enzymes were noted (62, 63). Similar effects on SAA metabolism are present in long-lived genetic models of GH and TH deficiency despite normal calorie and SAA intake. For example, in Ames dwarf mice, hepatic TSP enzyme activities, as well as those of the TMP involved in methionine-dependent methyl donation and methionine recycling, are significantly increased (173), leading to a decrease in steady-state levels of the universal methyl donor, SAM, and an increase in metabolic flux of methionine through the TSP (20, 174). Increased flux is accompanied by increased expression of TMP and TSP genes, including CBS and CGL in the liver and increased methionine flux through the TSP in the brain and kidney. SAA metabolism is also altered in similar directions in Snell dwarf mice (178). Similar to hypopituitary dwarf mice, GHRKO and growth hormone releasing hormone knockout mice show perturbations in metabolic and gene expression signatures of SAA and glutathione pathways, including increased expression of CBS and CGL (3, 21). Pharmacological inhibition of TH production results in similar changes in hepatic SAA metabolism, CGL expression, and H2S production (66).
Changes in hepatic SAA metabolism described above are coincident with increased expression of TMP and TSP genes, including CBS and CGL, which are likely driven, in part, by ATF4. ATF4 is increased on the protein (but not mRNA) level in multiple genetic and dietary models of longevity (88, 105). While the mechanism in dietary models is likely via GCN2 kinase activation as discussed above, the mechanism in hypopituitary models remains unknown. However, mouse models of subclinical hypothyroidism display increased ER stress (199), a known trigger of ATF4 activation (49). Our recent finding that hepatic ATF4 is also increased specifically in the hypothyroid state (66) suggests a direct or indirect role of TH signaling in ATF4-mediated regulation of CGL. Nonetheless, mechanistic details of ATF4 stabilization under hypothyroid states and direct evidence of a causal role of ATF4 in hypothyroid-induced H2S production remain to be tested in these models.
How could H2S specifically contribute to extended longevity? A variety of nonmutually exclusive possibilities are likely. H2S is best characterized by its ability to posttranslationally modify exposed cysteine or disulfide residues on the surface of target proteins, including membrane receptors, thus altering their structure, stability, and/or function. For example, H2S regulates multiple physiological endpoints via sulfhydration of ATP-dependent potassium channels (KATP channels) present on specific cell types as discussed previously (85, 198). On vascular smooth muscle cells, stimulation of KATP channels results in vasorelaxation (128), while in pancreatic beta cells, KATP stimulation by H2S decreases insulin secretion (192). Thus, increased H2S could contribute to alterations in lipid metabolism and insulin sensitivity observed in these long-lived mouse models via effects on KATP channel activity in these or other cell types.
H2S may also contribute to the reduction of hormones negatively associated with longevity, including IGF-1. Recently, we found that H2S is required for the reduction of circulating IGF-1 and thyroxine (T4) levels found in drug-induced hypothyroidism or upon fasting (66). H2S may also contribute to the decrease in serum T3, T4, blood glucose, and oxygen consumption in rats and chickens fed garlic and garlic-derived polysulfides (134, 168, 169, 175). This role of H2S in the negative regulation of TH and GH action, and thus positive regulation of its own production, is consistent with the ability of NaHS supplementation to increase renal CBS and CGL expression and activity and to improve H2S bioavailability in kidney, plasma, and urine of aged mice (69). Future studies will be required to determine the mechanism by which H2S contributes to the downregulation of these hormones, and whether it involves KATP channel polarization or some other mechanism.
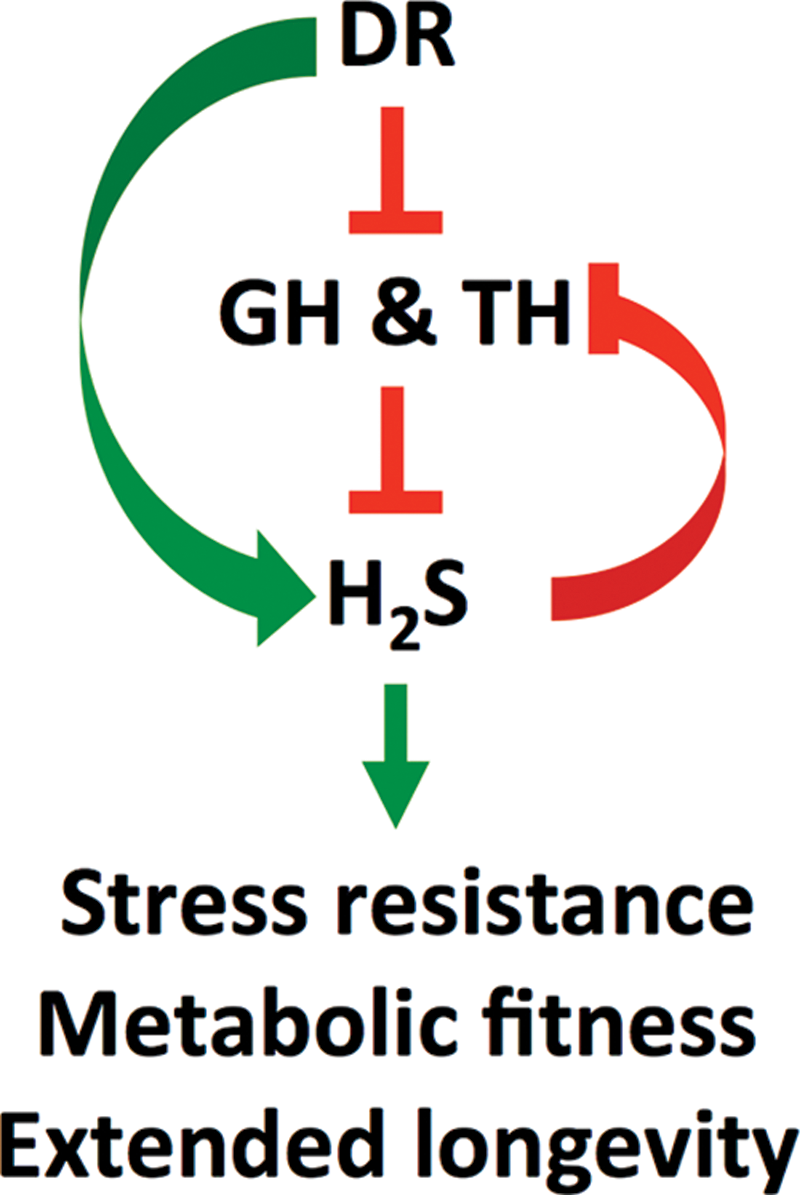
Finally, epigenetic changes could also serve as a link between increased transmethylation and H2S production in dwarf models, with implications for longevity. Treatment of hypopituitary mice with GH early in life abrogates longevity benefits, presumably through epigenetic changes that occur in the presence of GH during this narrow developmental window (125). Intriguingly, hepatic H2S production capacity, which remains high in 18-month-old dwarf mice, is also reduced late in life by early-life GH exposure (66). Whether this is a result of epigenetic changes in TSP genes themselves or some other mechanism, for example, changes in hypothalamic inflammation (144), remains to be seen. A summary of hormonal regulation of endogenous H2S production and the interaction with DR in animals is given in Figure 5.

In summary, genetic models of longevity due to perturbations in GH and TH signaling lead to profound alterations in SAA metabolism, including the expression of H2S generating enzymes CGL and CBS, particularly in the liver. Similar changes are observed on DR. An integrated and composite model of the regulation and cross talk between GH, TH, diet, and H2S is presented in Figure 6.
Plasticity of TSP Gene Expression in Response to Different Stimuli: An Unbiased Approach Using the NCBI GEO Profiles Database
We next used an unbiased approach to identify factors affecting TSP gene expression with the potential to modulate H2S production in human, mouse, and rat tissues and cells using the NCBI Gene Expression Omnibus (GEO Profiles) Database (9, 38). Table 1 summarizes the reported effects of nutrition, hormone activity, drugs, environmental and occupational pollutants, medical treatments, and health-related conditions on CGL and CBS gene expression. A wide range of stimuli affect TSP gene expression. In terms of dietary influences, red meat consumption in humans is associated with decreased CGL expression in the colon of irritable bowel disease patients, but not in control patients (GDS3471/217127_at/CTH). In mice, HFD (GDS4783/10503023/Cth), ketogenic diet (GDS2738/1426243_at/Cth), “fast food diet” (GDS3232/1426243_at/Cth), high-salt diet (GDS1545/D17370_at/Cth), and zinc deficiency (GDS4614/1367838_at/Cth) are all associated with a reduction of CGL expression in various tissues, while fasting increases CGL expression in the liver (GDS3893/9031/Cth). CGL expression in the liver is also subject to circadian control (GDS3084/1367838_at/Cth). Whether or not the circadian-related expression pattern is due to the periodicity of feeding and/or physiological changes during the course of 24 h is yet to be determined.
The NCBI GEO Profiles repository was used to extract data regarding the effects of dietary, hormonal, medical, environmental, and genetic stimuli on regulation of CGL (referred as CTH in the data set identifiers) and CBS gene expression. In addition, the biological relevance/endpoint correlating with the change in CGL or CBS gene expression is given in the “Change in H2S Producing Enzyme Column,” when provided in the GEO Profiles repository. For more information on each study, the GEO Profiles number in the far right column can be used to search for more details on the study, the original publications arising from the data set, and the study authors at the GEO Profiles website:
ATF4, activating transcription factor 4; CBS, cystathionine β-synthase; CGL, cystathionine-γ-lyase; iPSC, induced pluripotent stem cells; IPF, idiopathic pulmonary fibrosis; IR, ionizing radiation; VALI, ventilator-associated lung-induced injury.
Endocrine and hormonal influences on CGL and CBS expression are also evident in the GEO Profiles database. In human papillary thyroid carcinoma, CGL expression is decreased compared to adjacent normal tissue (GDS1665/206085_s_at/CTH), while CBS expression is decreased in human MCF7 breast cancer cells on silencing of the estrogen receptor expression (GDS4061/212816_s_at/CBS). Serum/growth factor withdrawal increases CGL expression in human smooth muscle cells, while GH addition to male rats decreases hepatic CGL expression (GDS862/8.2.2.19/Cth). Finally, leptin supplementation (GDS3653/ILMN_2733193/Cth) and retinoic acid treatment (GDS799/2827/Cth) decrease CGL expression in liver and smooth muscle cells, respectively. Future experiments are required to better understand how these changes in TSP gene expression impact H2S production in vivo.
H2S in Longevity Extension
CBS and CGL are multifunctional proteins with important roles in H2S generation, but are also directly responsible for de novo cysteine biogenesis, with indirect effects on protein translation, central one-carbon metabolism, and cellular redox state. Such pleiotropic effects will make it difficult and time-consuming to dissect the specific contribution of endogenous H2S to health and longevity phenotypes in the context of DR or long-lived mutants in rodent model systems. Nonetheless, data are rapidly accumulating in support of the ability of exogenous H2S to slow the aging process and extend longevity across evolutionary boundaries.
Longevity extension by H2S was first shown in the nematode worms Caenorhabditis elegans. Exposure to 50-ppm H2S in air during early development and continuing throughout life extends mean life span by 70% (114). Treatment early in adulthood with the H2S donor molecule and garlic constituent diallyl trisulfide also extends worm longevity in a skn-1-dependent manner (133) as does the slow-releasing H2S donor GYY4137 (137). Deletion of the H2S generating enzyme mpst-1 (3-MST) reduces worm life span, and this can be rescued with exogenous H2S in the form of GYY4137 (138). Increased H2S production is also required for extended longevity on removal of germ cells (183), a distinct model of longevity extension in worms. In the yeast Saccharomyces cerevisiae, the presence of exogenous H2S donors NaHS and GYY4137 during early logarithmic growth extends yeast chronological life span (65).
While the effects of exogenous H2S on rodent longevity are not yet known, there is an aging-dependent decline in endogenous H2S in tissues from wild-type rats (135) and mice (69) that is attenuated by exogenous H2S supplementation (69). Exogenous H2S in the form of injected NaHS or inhaled H2S also prevents neurodegeneration and neurovascular dysfunction in mouse models of Parkinson's induced by intracerebral homocysteine injection or administration of the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, respectively (84, 87). On a cellular level, CGL deficiency in mouse embryonic fibroblasts results in premature replicative senescence, a cellular model of aging, and this is rescued by H2S (NaHS) supplementation (191).
H2S can also extend life span and defer senescence in plants. During postharvest of the daylily flower, endogenous H2S production is diminished due to decreased activities of the plant H2S-producing enzymes L- and D- cysteine desulfhydrases, coinciding with senescence of the plant. Exogenous NaHS blocks the decrease in endogenous H2S production, slows senescence, and extends postharvest life of daylilies (109). H2S also alleviates postharvest senescence of broccoli while activating expression of antioxidant defense-related genes (104). Treatment of grapes with a 1 mM NaHS solution decreases rotting and threshing, maintains antioxidant systems, and prevents senescence-related breakdown of chlorophyll and accumulation of carotenoid (120). Kiwi fruit senescence and tissue softening are similarly delayed and antioxidant systems maintained with NaHS treatment in a dose-dependent manner (200).
What are the implications of exogenous H2S exposure on humans? While acute high-dose exposures are lethal, the health effects of chronic low-dose exposure remain inconclusive (107). For example, a series of epidemiological studies on people from Rotorua, New Zealand, with long-term exposure to elevated levels of ambient H2S (average of ∼20 ppb, with a range of 0–64 ppb), found that people in the upper quartiles of H2S exposure had slightly improved psychomotor reaction times and episodic memory (139), without any association with cognitive dysfunction, mood impairment, or peripheral neuropathy (132, 139).
Conclusions
At the right concentration, exogenous H2S has clear health benefits, including longevity extension in experimental model organisms. Increased endogenous H2S production is common to numerous experimental models of extended longevity, including genetic models with shared defects in GH and/or TH signaling (Ames, Snell, GHRKO, FGF21o/e, IRS-1KO), as well as various forms of DR, along with broader changes in SAA metabolism with implications for DNA methylation and redox status. The relative contribution of increased H2S to health span or life span benefits in these models remains to be determined, as does the mechanism by which it occurs. In conclusion, our ability to control H2S levels using exogenous H2S donors or by modifying the endogenous H2S equilibrium has the potential to increase “shelf life” across evolutionary boundaries, including our own.
Footnotes
Acknowledgments
This work was supported by the following grants: nos. AG050777 (C.H.); DK090629 and AG036712 (J.R.M.); DK098525 and DK056123 (A.N.H.).