
Abstract
Significance:
Stomata sense the intercellular carbon dioxide (CO2) concentration (Ci) and water availability under changing environmental conditions and adjust their apertures to maintain optimal cellular conditions for photosynthesis. Stomatal movements are regulated by a complex network of signaling cascades where reactive oxygen species (ROS) play a key role as signaling molecules.
Recent Advances:
Recent research has uncovered several new signaling components involved in CO2- and abscisic acid-triggered guard cell signaling pathways. In addition, we are beginning to understand the complex interactions between different signaling pathways.
Critical Issues:
Plants close their stomata in reaction to stress conditions, such as drought, and the subsequent decrease in Ci leads to ROS production through photorespiration and over-reduction of the chloroplast electron transport chain. This reduces plant growth and thus drought may cause severe yield losses for agriculture especially in arid areas.
Future Directions:
The focus of future research should be drawn toward understanding the interplay between various signaling pathways and how ROS, redox, and hormonal balance changes in space and time. Translating this knowledge from model species to crop plants will help in the development of new drought-resistant crop species with high yields.
Introduction
Stomata are tiny pores formed by a pair of guard cells on the surfaces of plant leaves and stems. Their primary role is to maintain an adequate supply of carbon dioxide (CO2) for photosynthesis while limiting water loss through transpiration. To adapt to ever-changing environmental conditions, plants are constantly adjusting their stomatal apertures to control leaf CO2 and water content.
Guard cells sense the concentration of CO2 in the substomatal cavity (intercellular CO2 concentration or Ci) and are able to respond rapidly to changes in Ci (31, 98). When conditions are optimal for photosynthesis in C3 plants, CO2 is consumed by carboxylation reactions in the chloroplasts of mesophyll cells. This leads to a decrease in Ci below the ambient CO2 concentration (∼400 ppm) and triggers stomatal opening to maintain CO2 supply for the Calvin–Benson cycle (CBC) in mesophyll chloroplasts. In contrast, an increase in Ci leads to stomatal closure; this helps to conserve water but can also lead to increased leaf temperature and reduced uptake of nutrients by the transpiration stream. Such regulation may occur within minutes and is achieved by controlled transport of osmoregulatory ions, mainly potassium (K+), chloride, and malate through different types of ion channels in the guard cell membranes (45, 65).
Under conditions that limit photosynthesis, such as darkness, Ci increases due to reduced CO2 fixation through the CBC and stomatal aperture decreases (Fig. 1). Conversion of CO2 into bicarbonate (HCO3 −) mediates the stomatal response to changes in Ci (28, 52, 53). CO2 is spontaneously dissolved in water with formation of HCO3 −. Inside plant cells, the rate of this reaction is accelerated by carbonic anhydrases (CAs) (26). A Raf-like protein kinase, HT1 (high leaf temperature 1) is a highly CO2-specific stomatal regulator (44, 45, 52) involved in controlling the activity of the slow anion channel 1 (SLAC1) (49).

Stomatal opening occurs via activation of the guard cell plasma membrane hydrogen ATPase (H+ATPase), which causes hyperpolarization of the membrane and subsequent uptake of K+ through polarization-dependent inward rectifying K+ channels. Stomatal closure occurs through inactivation of the H+ATPase and activation of the guard cell anion channels, and this leads to depolarization of the membrane and activation of outward rectifying K+ channels (65).
During drought, water uptake by roots is limited and to avoid water loss by transpiration plants close their stomata. This is mostly regulated by the stress hormone abscisic acid (ABA). The first steps in ABA signaling leading to stomatal closure are well characterized. The key genetic components in the ABA signaling pathway include the following: ABA receptors pyrabactin resistance (PYR) 1/PYR1-like (PYL)/regulatory components of ABA receptors (RCARs) (76, 112), a group of type 2C protein phosphatases (PP2Cs), such as ABA-insensitives (ABI) 1 and 2 (140, 143), the protein kinase open stomata 1 (OST1/SnRK2.6), and calcium-dependent protein kinases (CDPKs, in Arabidopsis CPKs).
In the absence of ABA, PP2Cs are active and function as constitutive inhibitors of OST1 and CDPKs. Binding of ABA to its receptors inactivates the PP2Cs, and OST1 is activated either by autophosphorylation (11) or phosphorylation by some other protein kinase. Once activated, OST1 is involved in the activation of the guard cell anion channels SLAC1 and quick anion channel 1 (QUAC1) (57, 73) and inactivation of the inward rectifying Shaker family K+ channel (KAT1) (122). Activation of anion channels leads to an efflux of anions and small metabolites, such as malate, and in combination with the deactivation of the plasma membrane H+ATPase 1 (AHA1) (88) causes plasma membrane depolarization and the consequent activation of voltage-dependent K+ efflux channels (2). The resulting efflux of anions and K+ leads to the loss of guard cell turgor and the closure of stomatal pores.
In addition to adjustments of stomatal aperture, plants also react to long-term changes in environmental conditions by adapting the stomatal density in newly developed leaves. Mechanisms underlying the regulation of stomatal density in response to environmental changes have been recently reviewed (18, 28) and will not be discussed here. In this study, we address how stomata sense the changes in CO2 concentration and water availability in C3 plants, how drought-induced stomatal closure leads to increased production of reactive oxygen species (ROS), and how ROS signals regulate stomatal movement.
The major forms of ROS, singlet oxygen, superoxide anion, hydrogen peroxide (H2O2), and hydroxyl radical are formed in different subcellular compartments of plants. This occurs mostly through photorespiration-related reactions in peroxisomes, in mitochondrial electron transport chains, by over-reduction of the chloroplast electron transport chain, and by specific ROS-producing enzymes (15, 92, 106). This review covers the current knowledge of how signaling cascades relating to ROS, redox, and changing environments are involved in the adjustment in stomatal aperture.
Coordination of Mesophyll Photosynthetic Processes with Stomatal Aperture
Stomatal responsiveness to light and CO2 is suppressed when the epidermis is detached from the leaf, whereas re-establishment of the contact between mesophyll and detached epidermis restores stomatal responsiveness. This suggests the existence of diffusible chemical or vapor-phase signals released from the mesophyll (71, 99, 121). The nature of the mesophyll-driven signals has, however, remained elusive but a broad range of substances, including sucrose and malate, have been considered (44, 71, 121). There is evidence that CO2 concentration inside the leaf rather than outside the leaf influences stomatal aperture (98; Fig. 2). This notion is supported by several studies implying that red light-induced stomatal opening is mediated by the reduction of Ci, which is in turn caused by the increased photosynthetic activity of mesophyll cells (118, 119).
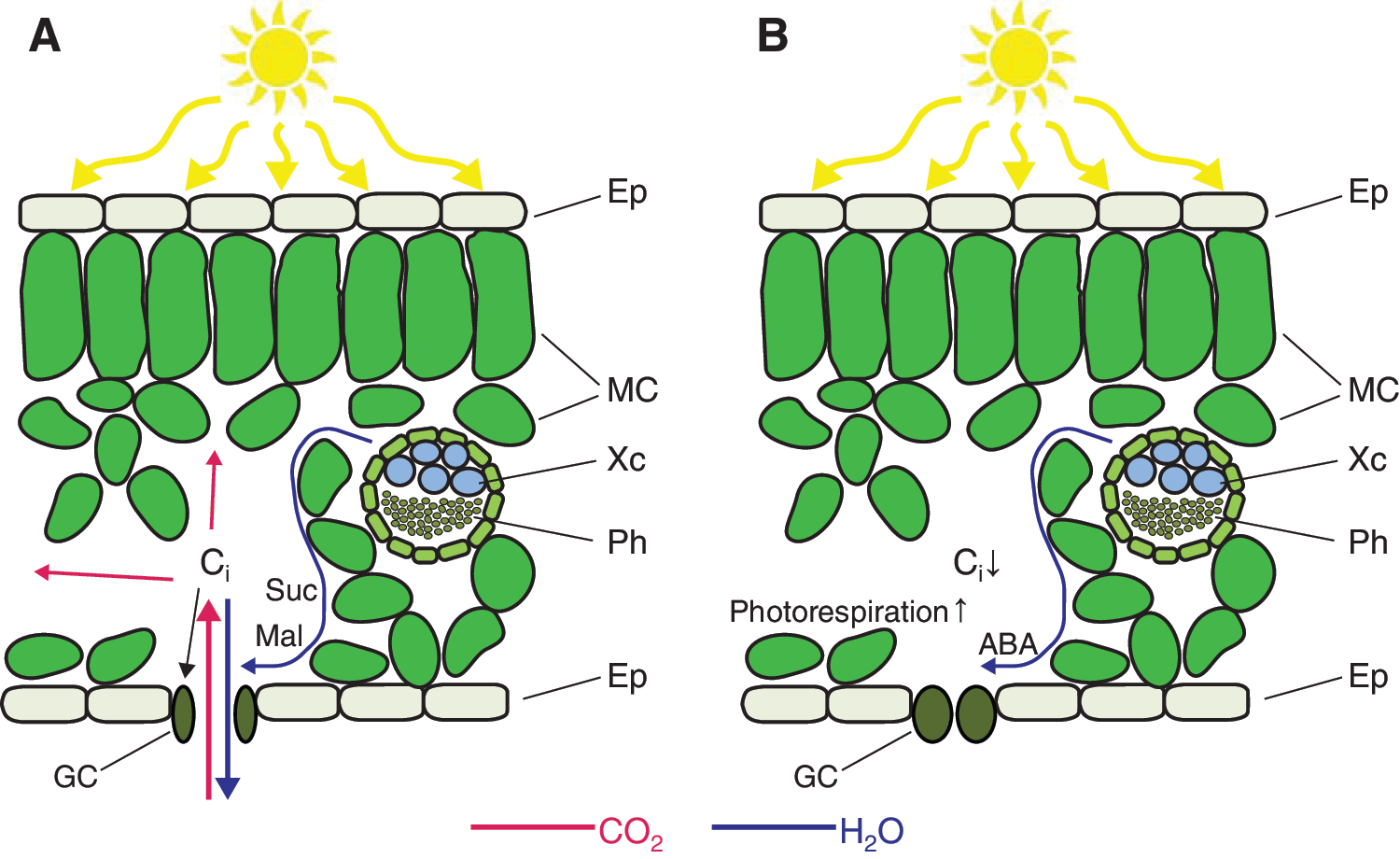
Guard cells are also known to have signaling components specific to CO2 responses. Plants carrying mutations in the highly CO2-specific protein kinase HT1 (42, 43, 49) showed severely suppressed stomatal opening in response to red light-induced decrease in Ci (80). However, Ci may not be the only signal through which guard cells get information on mesophyll processes. Functional red light-induced stomatal opening under artificially sustained Ci suggested that mesophyll photosynthesis could coordinate stomatal regulation by Ci-independent mechanisms (68, 89). The existence of a Ci-independent signal was further supported by unaffected stomatal conductance in plants, which had high Ci due to suppressed Rubisco activity (145).
Blue light, a factor affecting photosynthesis and stomatal opening, is directly perceived by guard cells (126, 127, 159). In addition, over-reduction of the plastoquinone pool in the mesophyll cell chloroplasts was recently suggested to induce ROS-mediated stomatal closure (147). Taken together, it seems that guard cells are able to recognize both changes in environmental conditions, such as light quality, and inner Ci-dependent and Ci-independent signals from mesophyll. Although the influence of mesophyll cells on stomatal aperture has been demonstrated by several studies, many details of this interplay remain unknown and should be elucidated in the future.
The Role of Guard Cell Chloroplasts in Stomatal Signaling
In most species, guard cell chloroplasts are smaller, present in a lower number, and have a less developed thylakoid structure with reduced granal stacking than mesophyll cell chloroplasts (3, 154). Despite these differences, guard cell chloroplasts still have functional photosystems I and II as well as CBC activity (10, 69, 70, 83, 117) and they can significantly contribute to guard cell metabolism (67). Similarly, regulation of photosynthesis in guard cells and in mesophyll by environmental factors can provide an indirect sensing mechanism coordinating stomatal behavior with mesophyll demands for CO2 (121, 133). There exist, however, some plant species such as the orchid genus Paphiopedilum (21, 104) that do not have chloroplasts in their guard cells but still display stomatal responses to high CO2 concentration and changes in light conditions (104), independent of photosynthesis in guard cells.
Guard cell chloroplasts are thought to be involved in osmoregulation of stomatal movements through photosynthetic carbon fixation, which produces osmotically active sugars. However, estimations of photosynthesis-derived osmotica in guard cells vary from 2% to 40% of the total pool of osmotically active substances depending on plant species and experimental approaches (67). Starch degradation in guard cell chloroplasts, which can be initiated by blue light and low CO2 (50, 111), can also contribute to the formation of guard cell turgor by releasing monosaccharides and/or provide phosphoenolpyruvate for CO2 fixation by cytosolic phosphoenolpyruvate carboxylase, leading to the formation of malate (50, 67, 121). Plants with impaired starch synthesis, both in mesophyll and in guard cells, demonstrated reduced stomatal responsiveness to elevated CO2, indicating that conversion of osmotically active carbohydrates has a role in the reduction of osmotic pressure during stomatal closure (8).
Although guard cell photosynthesis is important for the energization of stomatal opening (8, 133), it does not seem to be directly involved in the regulation of stomatal aperture as stomata of plants without chlorophyll in guard cells still remained responsive to CO2 and ABA (9, 104). However, numerous studies have highlighted the importance of guard cell chloroplast in stomatal regulation through chloroplast-dependent ROS accumulation (128, 149), calcium ion (Ca2+) release (107, 153), and retrograde signaling (113) (see the ROS Production and Sensing in Guard Cell Signaling During Drought section below). In conclusion, the function of guard cell chloroplasts may not be compulsory for CO2- and ABA-triggered stomatal regulation but appears to be important for amplifying and fine tuning processes through light-derived control of other signals.
Mechanism of Stomatal Opening Induced by Lower Than Ambient Concentrations of CO2
Decrease of CO2 concentration in the leaf intercellular air spaces is a powerful stimulus for the regulation of stomatal aperture since it can induce stomatal opening even under conditions that normally promote stomatal closure, such as darkness when photosynthesis is not possible and low air humidity, which poses the risk of wilting (85). Accordingly, mechanisms of low CO2-induced stomatal opening are likely to have an early evolutionary origin as stomata of ancient vascular plants, lycophytes and ferns, displayed rapid stomatal opening in CO2-free air but only a weak response to high CO2 concentrations (13, 65). The stomata of the lycophyte Selaginella responded to both elevated and reduced CO2 concentrations as well as to ABA (120), and stomatal closure in response to elevated CO2 and ABA was present in some fern species and had been possibly lost in others (48).
Although there are major gaps in our understanding of how low CO2 triggers stomatal opening, it is obvious that it must involve signaling systems that control the activity of H+ATPases (118). This can be achieved either by enhanced translocation of H+ATPases from internal membranes into the plasma membrane or by the regulation of the H+ATPase activity (Fig. 3A). The importance of H+ATPase translocation was demonstrated by impaired stomatal opening in response to low CO2 in the Arabidopsis mutant patrol1, which has a mutation in the endosome-localized proton ATPase translocation control 1, a protein involved in the translocation of the major guard cell H+ATPase, AHA1, into the plasma membrane (41). The mechanisms controlling AHA1 activation, however, are not known.

Other transporters also contribute to the production of osmotic pressure in guard cells during low CO2-induced stomatal opening. For example, plants with defects in the nitrate transporter 1.1. demonstrated decreased stomatal opening in CO2-free air, accompanied by reduced nitrate accumulation in guard cells (40). Malate transporter ATP-binding cassette B14 (ABCB14) can also promote stomatal opening by uptake of malate from the apoplast (72). The involvement of other transporters and regulatory pathways in guard cells activated by reduced CO2 concentration still awaits identification.
Mechanism of High CO2 Concentration-Induced Stomatal Closure
Stomatal closure is triggered by an increased concentration of CO2 and hence elevated Ci induces anion efflux through anion channels in the guard cell plasma membrane, followed by K+ efflux, and subsequent water outflow and a reduction of guard cell volume (Fig. 3B). Plants with defective S-type anion channel SLAC1 or with defects in the mechanisms that control SLAC1 activation display severely impaired stomatal closure in response to an increase in CO2 concentration (49, 86, 103, 141, 157).
The role of apoplastic malate for high CO2-induced stomatal closure was demonstrated already in 1993 (44). Malate can be transported from mesophyll to guard cells and could act as a mesophyll-driven signal linking mesophyll metabolism with stomatal regulation (6, 109). The R-type anion channel QUAC1 can be activated by apoplastic malate. Accordingly, plants lacking QUAC1 in their guard cells demonstrated partially impaired stomatal response to high CO2 (57, 90). Guard cells can also control the level of apoplastic malate by its uptake via ABCB14 activity (72). ABCB14 acts as a negative regulator in high CO2-induced stomatal closure as demonstrated by accelerated and delayed stomatal responses to high CO2 concentration in the abcb14 mutants and ABCB14 overexpressors, respectively (72).
It has been suggested that Ca2+ play a role as a second messenger in high CO2-induced stomatal closure. This has been concluded based on experiments where Ca2+ accumulation in guard cells subjected to higher than ambient CO2 concentration was observed, and from the impaired high CO2-induced stomatal closure in the presence of Ca2+ chelators, such as BAPTA or EDTA (55, 124, 152).
Genetically encoded Ca2+ sensors revealed that guard cells displayed oscillations of cytosolic Ca2+ concentration [Ca2+]cyt and these patterns were often associated with changes in stomatal aperture (4). However, unexpectedly, guard cells exposed to reduced CO2 concentration demonstrated more [Ca2+]cyt transients than those under elevated CO2 concentration (161). As guard cells produced “spontaneous” cytoplasmic Ca2+ transients and Ca2+ is required for high CO2-induced stomatal closure, it was suggested that elevated CO2 concentration enhances sensitivity of stomatal closing mechanisms to [Ca2+]cyt. In agreement with this hypothesis, CO2-derived HCO3 − enhanced Ca2+ sensitivity of the S-type anion channel activation in guard cells (157).
HCO3 − as a Signaling Molecule in CO2-Controlled Stomatal Movements
A reduction or an increase of Ci should be sensed and translated into activation of corresponding signaling pathways in guard cells. The CO2 permeability of biological membranes in relation to direct diffusion of CO2 through the membranes versus the role of CO2-permeable aquaporin channels has been addressed in several articles and there are indications that specific aquaporins can have a significant role in CO2 uptake (see 38 for a review).
Recently, a plant aquaporin, plasma membrane intrinsic protein 2;1 (PIP2;1), was shown to function as a channel for CO2 diffusion in Xenopus laevis oocytes (145). However, knocking out PIP2;1 was not sufficient to impair stomatal CO2 responses (145). This could be explained by functional redundancy among guard cell aquaporins; there are 35 aquaporin homologues in Arabidopsis. Transport of CO2/HCO3 − to chloroplasts also depends at least partly on aquaporins, including the PIP1;2 that is located in chloroplast envelope (138). In addition to the proposed role for PIP2;1 in CO2 transport, it was also involved in ABA-triggered stomatal closure (37).
Conversion of CO2 into HCO3 − is an important step that mediates stomatal responses to changes in ambient CO2 and Ci (28, 52, 53). Although CO2 is spontaneously dissolved in water with formation of HCO3 −, in cells the rate of this reaction is accelerated by CAs (26). Among CAs expressed in Arabidopsis, the function of βCA1 and βCA4, localized in chloroplasts and in plasma membrane, respectively, was important for the rapid stomatal response to changes in CO2 levels (52; Fig. 3). While single βCA mutants did not display clearly altered CO2 sensitivity, the double knockout of both βCA1 and βCA4 significantly delayed stomatal responses to CO2 (28, 52, 53). Interestingly, PIP2;1 physically interacted with βCA4 and this connection has been suggested to enable the generation of CO2 concentration gradient and thus to enhance transport of CO2 into guard cells (145).
The importance of HCO3 − is further supported by experiments showing that the concentration of cytosolic HCO3 −, rather than CO2, activated S-type anion channels in guard cell protoplasts (157). The role of HCO3 − as a small signaling molecule in guard cells was also confirmed by reconstitution of CO2 signaling pathway in X. laevis oocytes coexpressing PIP2;1, βCA4, SLAC1, and CPK6/23 or OST1. In these experiments, the presence of these proteins was enough to confer HCO3 −-induced activation of SLAC1 anion currents in oocytes (145).
Despite the established connection between cytosolic HCO3 − and anion channels in guard cells, our knowledge about CO2 signaling in guard cells has still major gaps. As an example, it has not been resolved which proteins can bind and/or sense the changes in HCO3 − concentration in guard cells to transmit the signal that eventually leads to changes in ion channel activities.
Mitogen-Activated Protein Kinases MPK4 and MPK12 and HT1: A New Pathway Controlling SLAC1 Activation in Response to Changes in CO2
Mutant screens with different approaches have led to the identification of important components in guard cell CO2 signaling (41, 43, 103, 141). The Raf-like protein kinase HT1 was identified by using thermal imaging of mutagenized plants subjected to low CO2. HT1 is expressed in guard cells and is a highly CO2-specific regulator, since plant lines carrying mutations in HT1 displayed stomata completely insensitive to changes in CO2 concentration, but remained responsive to other stimuli such as light, ABA, and air humidity (42, 43; Fig. 4). HT1 plays a role in controlling the activation of SLAC1 anion channel as a response to changes in CO2 concentration (Fig. 3). Experiments carried out in the heterologous system, X. laevis oocytes, demonstrated that SLAC1 activation by OST1 and by receptor-like protein kinase guard cell hydrogen peroxide-resistant 1 (GHR1) was suppressed by HT1 (49, 137).
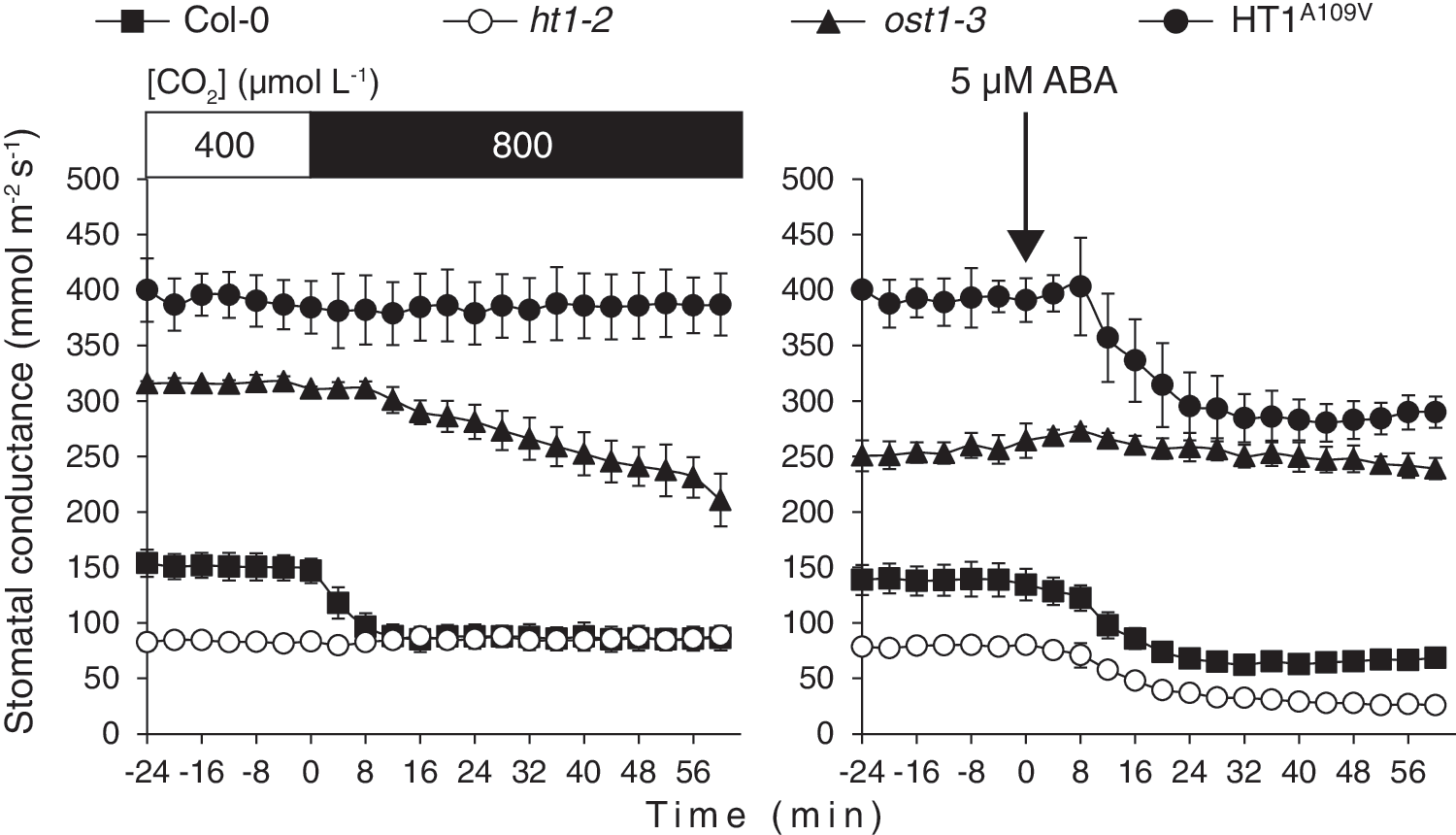
However, the mechanism how HT1 affects SLAC1 activation remains controversial. Some experiments have suggested that HT1 could phosphorylate OST1 and by that suppress SLAC1 phosphorylation by OST1 (137), however, these experiments were not confirmed in another study (49). Despite using various versions of the HT1 protein, no inhibition on OST1-induced phosphorylation of SLAC1 was observed in the presence of HT1. Instead, HT1 showed phosphorylation activity toward GHR1 and the N-terminus of SLAC1 in vitro; the functional outcome of these reactions, however, remained unclear (49). Thus, the mechanism by which HT1 controls anion channel activation during stomatal closure in response to elevated CO2 requires further research.
One should remember that results obtained in in vitro experiments and heterologous systems, such as X. laevis oocytes, do not necessarily reflect the regulatory interactions in planta due to missing components, and the models predicted in these artificial experimental systems need to be confirmed in plants before constructing regulatory models.
Studies with mitogen-activated protein kinase (MPK or MAPK) inhibitors and work focusing on the natural variation of water-use efficiency and ozone sensitivity among Arabidopsis natural accessions revealed that MPK12 is an important component of stomatal regulation (24, 58, 59). Further work showed that MPK4 and MPK12 are essential for CO2-dependent stomatal regulation (Fig. 3). Both of these MPKs inhibited HT1 activity in vitro, whereas experiments in X. laevis oocytes indicated that MPK12 was able to restore SLAC1 activation by GHR1 in the presence of HT1 (49, 58). It is noteworthy that CO2-induced stomatal responses in plants lacking MPK12 and MPK4 in their guard cells were fully abolished, similar to that observed for strong HT1 mutants. Thus, these MPKs seem to play a central role in controlling HT1 in CO2-induced stomatal regulation, however, the mechanism that relays changes in HCO3 − concentration to MPKs in guard cells remains to be addressed.
A mate-type transporter, resistant to high CO2 (RHC1), was also suggested to act as an upstream regulator of HT1. Its abundancy is high in guard cell plasma membranes and its activity was essential for stomatal response to high CO2 concentration (137). Phenotype of the rhc1 ht1-2 double mutant and oocyte experiments implied that RHC1 could act as a HCO3 − sensing element upstream of HT1, although its exact mechanism remained unknown (137). In contrast to these results, another study (149) showed that RHC1 alone was able to cause HCO3 −-insensitive ion currents in X. laevis oocytes, making the role of RHC1 in CO2/HCO3 − sensing unresolved.
ROS Production and Sensing in Guard Cell Signaling During Drought
Under certain conditions, stomata must close despite the mesophyll CO2 demands and low Ci. This type of stomatal closure can be induced, for example, by limited water availability, salt/osmotic stress, air pollution, or by pathogen attack, which is often referred to as stomatal immunity. Stomatal closure is one of the earliest responses of plants to water deficit. This rapid response is orchestrated by a complex network of signaling pathways where the main player, ABA, operates together with second messengers, Ca2+ and ROS (23, 100), and overrides the stomatal regulation by CO2. The participation of ABA in stomatal responses to drought is well known (35) and ROS and Ca2+ are important mediators in ABA signaling.
Stomatal closure is accompanied by increased ROS formation in the guard cell apoplast and chloroplasts in response to various treatments (128, 129; Fig. 5). Apoplastic ROS are generated mainly by two different types of enzymes: plasma membrane nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (respiratory burst oxidase homologues or RBOHs) and cell wall peroxidases. In Arabidopsis guard cells, there are two main isoforms of NADPH oxidases, AtRBOHF and AtRBOHD, which among other signals can also be regulated by ABA-depended processes (66). ABA-triggered stomatal response was significantly reduced in the atrbohF mutant and the phenotype was enhanced in the atrbohD atrbohF double mutant whereas the atrbohD single mutant did not differ from the wild type (66).
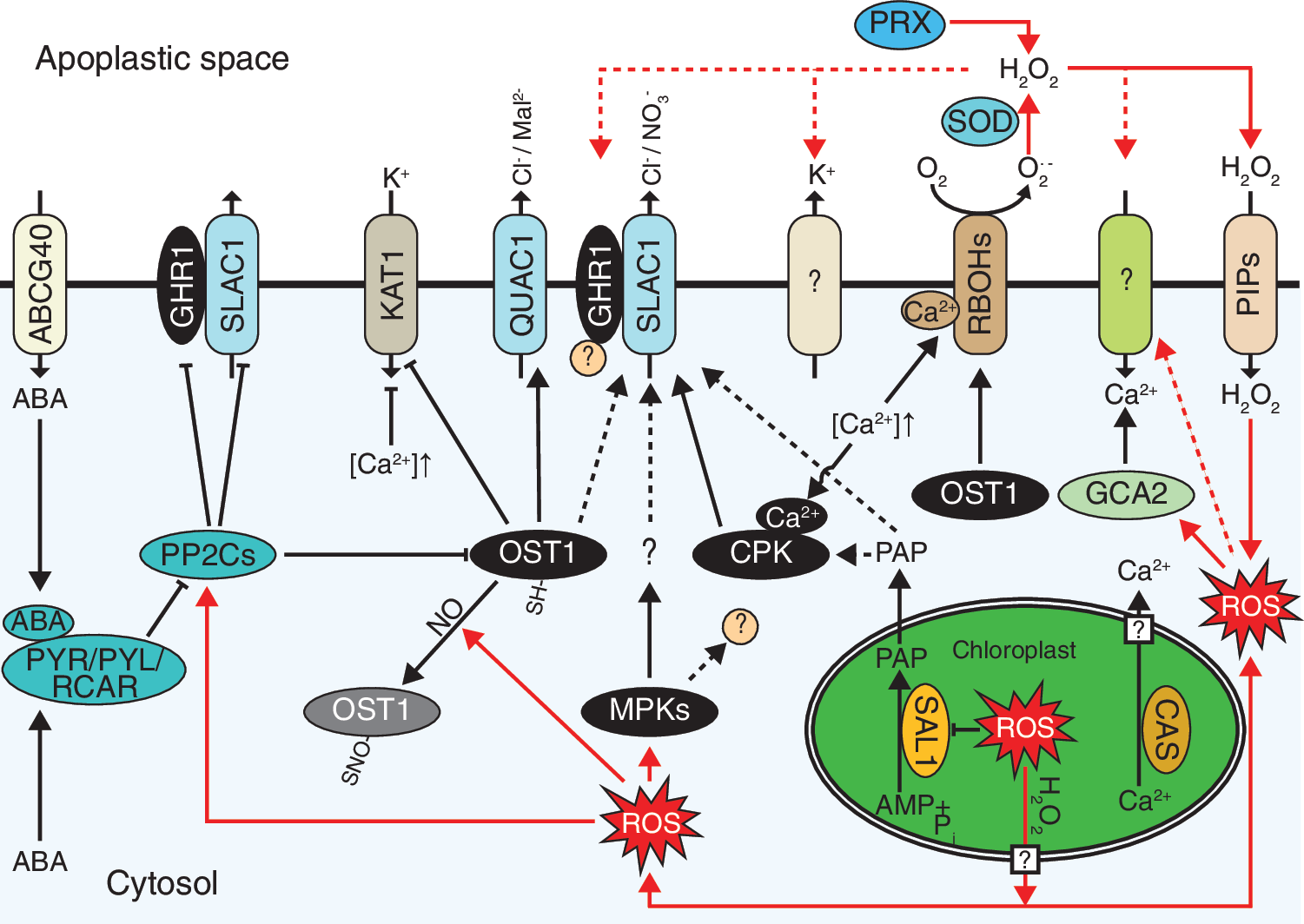
Due to its obvious role in pathogen-triggered ROS burst, RBOHD is more commonly recognized for its function in plant immune defense (77). However, recently, NADPH oxidases were shown to be involved also in the guard cell CO2 responses, and the CO2-induced ROS burst required ABA (17).
In addition to NADPH oxidases, also the cell wall-bound salicylhydroxamic acid (SHAM)-sensitive peroxidases take part in apoplastic ROS production around guard cells (61, 97). These peroxidases are involved in the pathogen-triggered ROS burst (61), but they may also be involved in the response to abiotic stress (93, 110). Apoplastic ROS are also produced by other oxidases, such as di- and polyamine oxidases (114). Copper amine oxidase and polyamine oxidases contribute to H2O2 production involved in the stomatal closure induced by ABA and ethylene in Vicia faba and Arabidopsis thaliana, respectively (5, 51). However, evidence for the involvement of peroxidases and amine oxidases in apoplastic ROS production has come from inhibitor studies and further research is needed to understand specific function, molecular identities, and significance of these proteins in ROS-induced stomatal regulation.
Apoplastic ROS production initiates the activation of plasma membrane Ca2+ channels leading to an increase in cytosolic Ca2+ levels. The molecular identity of these inducible plasma membrane Ca2+ channels is still not clear. In the cytosol, Ca2+ stimulates the activation of NADPH oxidases either directly by binding to their cytoplasmic EF-hands (63) or indirectly by affecting their phosphorylation by CPKs (27), or by complexes formed by calcineurin B-like proteins (CBLs) and CBL-interacting protein kinases (CIPKs) (131). A particular complex formed by CBL1/CBL9-CIPK26 phosphorylated and activated RBOHF (27).
The increase in cytoplasmic Ca2+ is sensed also in the chloroplasts where a thylakoid membrane-associated Ca2+-binding protein, calcium sensing receptor (CAS), is activated through yet unidentified mechanism. The activation of CAS was responsible for the release of Ca2+ from thylakoids and a chloroplastic ROS burst (107, 108, 142, 153), both of which contribute to the cytoplasmic Ca2+ oscillations, apoplastic Ca2+-induced stomatal closure, as well as retrograde signaling during plant immune defense (108). Moreover, the drought sensitivity of the Arabidopsis cas mutant is caused by the improper closure of stomata (148), which further highlights the importance of chloroplastic Ca2+ signaling in stomatal regulation.
The role and ability of OST1 in direct activation of SLAC1 have been recently discussed (128). First, phosphorylation of SLAC1 by OST1 has only been detected in vitro, and second, multiple mutants of CPKs that showed stomatal phenotype still have an active OST1, which nevertheless cannot activate SLAC1-mediated ion currents in vivo in the absence of specific CPKs. Furthermore, plants with impaired OST1 were shown to have wild-type-like stomatal closure in response to Ca2+ (102), possibly via activation of CPKs. This poses a question whether in vivo OST1 would actually be involved in the activation of guard cell anion channels indirectly through controlling the activation of CPKs, possibly by phosphorylation of RBOHF. The resulting ROS burst would activate Ca2+ channels, followed by CPK-dependent activation of SLAC1 (123, 128). In this model, OST1 would function upstream of ROS production and be negatively regulated by the PP2C ABI1, as has been shown (60, 75, 101). Furthermore, GHR1, and its negative regulator ABI2, another PP2C, would be involved in the downstream activation of plasma membrane Ca2+ channels and subsequent stomatal closure (54, 101; Fig. 5).
Although there is clear evidence for the involvement of ROS in the regulation of stomatal aperture, it is still not known how the ROS signals are sensed in the guard cell apoplast. Identification of the ROS and redox sensors has been one of the major challenges in plant ROS research during recent years. In guard cells, only a few ROS sensing mechanisms are known to be involved in the stomatal regulation. These are the redox regulation of the GHR1 apoplastic domain (54) and the redox regulation of OST1 (146) and CPK1 (139).
GHR1 is a plasma membrane-associated atypical leucine-rich repeat receptor-like protein kinase that has been proposed to be involved in apoplastic ROS perception. The apoplastic C-terminal domain of GHR1 has two conserved cysteines (C-57 and C-66) that are necessary for the correct function of the protein (54). As discussed earlier, GHR1 has been implicated as a central regulator of guard cell CO2 and early ABA responses but the molecular mechanism for its function is still unclear. GHR1 has been shown to interact with SLAC1 (54), but it is not likely to activate SLAC1 by phosphorylation as its cytoplasmic kinase domain lacks the conserved amino acids that are required for kinase activity (Sierla et al., unpublished data). Therefore, it is likely that there are other unidentified proteins involved in the GHR1-mediated SLAC1 activation.
The protein phosphatases ABI1 and ABI2 have also been shown to be inactivated in the presence of H2O2 (81, 82), but the mechanism for this redox regulation is still unknown. Another example for redox regulation of ABI1 and ABI2 in guard cell is a glutathione peroxidase-like 3 (GPXL3) enzyme in H2O2 scavenging and cytosolic redox regulation in response to ABA and drought stress (91). GPXL3 was suggested to interact with ABI1 and ABI2. In addition, similarly to H2O2 (81, 82), oxidized GPXL3 decreased the phosphatase activity of ABI2 by affecting its redox status in vitro. However, both proteomic data and subcellular localization of GPXLs as GFP-fusions (7) suggest that GPXL3 is in fact a type II transmembrane protein anchored to the endoplasmic reticulum and/or Golgi so that the catalytic side remains in the lumen and would not be able to interact with the PP2Cs in vivo. In the light of these results, the molecular basis for the drought-sensitive and drought-resistant phenotypes of the gpxl3 null mutant and GPXL3 overexpressor lines, respectively, (91) and the possible mechanisms of the redox regulation of the guard cell PP2Cs remain unknown.
The Role of Sulfate in Drought Sensing and the Emergence of a New Pathway
A number of studies with different plant species have shown that low soil water potential decreases stomatal conductance even before any measurable change in leaf water potential can be observed (34, 36). These results suggest that roots can sense low soil water potential and transmit a signal to guard cells initiating stomatal closure. The earlier hypothesis that root-borne ABA acted as a drought signal to leaves has now been questioned since stomatal closure appears to be dependent on foliar ABA production (19, 47, 87). In addition, the ABA that accumulates in roots during long-term drought conditions appears to be derived from the shoots (79). Other signals, such as chemical, electrical, and hydraulic, have been suggested to play a role in root to shoot signaling (56) and they all are likely to contribute to the long distance signaling through various signaling networks.
The role of sulfate in root to shoot signaling and stomatal regulation has been recently highlighted by several studies. The need for sulfate during drought is known to increase as many sulfur-containing compounds, such as glutathione, are involved in plant abiotic stress responses (1). Once taken up from the soil and transported to chloroplast, sulfate is converted into cysteine or 3′-phosphoadenosine-5′-phosphosulfate (PAPS), which is then used for the synthesis of sulfur-containing compounds and production of sulfated compounds (33). Cysteine plays an important role in a plant defense against abiotic stress as it is a precursor for glutathione biosynthesis (105) and it is required for the sulfuration of molybdenum cofactor, which at its sulfated form is required for the final step of ABA biosynthesis. Intriguingly, significant coregulation of ABA biosynthesis and sulfur metabolism takes place under stress conditions to ensure adequate cysteine supply needed for the final step in ABA biosynthesis (14). Sulfate concentration in xylem sap was increased in response to drought and this enhanced the effect of ABA on stomatal regulation during early stage of water stress in maize (29). Similarly, xylem-derived sulfate promoted stomatal closure by direct activation of the R-type anion channel QUAC1 and enhanced ABA biosynthesis (78).
Several sulfated compounds accumulate in plant leaves under drought. These are sulfated in the cytoplasm by a family of enzymes called sulfotransferases (SOTs) that catalyze the transfer of sulfuryl group from PAPS to several different compounds, such as glucosinolates, flavonoids, brassinosteroids, and salicylic acid (SA) (46). However, the role of sulfation of these compounds in drought resistance is not well understood. Instead, the by-product of SOT-catalyzed sulfation, 3′-phosphoadenosine-5′-phosphate (PAP), has been implicated in drought and high light signaling (30). Once produced in the cytosol, PAP is transported to chloroplasts where it is detoxified by dephosphorylation to adenosine monophosphate by the adenosine bisphosphate phosphatase 1, SAL1 (115). High light and drought inactivated SAL1 by redox-regulated dimerization causing the accumulation of its substrate, PAP (16, 30). It was suggested that PAP moves into the nucleus (32) where it is thought to inhibit the post-transcriptional gene silencing of stress responsive genes by 5′-3′ exoribonucleases (30). However, it is not clear whether chloroplastic or cytoplasmic PAP is responsible for gene regulation since the PAPS/PAP antiporter transports PAPS out and PAP into the chloroplast according to a concentration gradient (33, 34), which implies that inactivation of SAL1 results in an increase of cytoplasmic PAP due to a decrease of the concentration gradient-driven transport of PAP to chloroplast.
The involvement of PAP in ABA-dependent stomatal closure was also shown recently (113). The sensitivity of the guard cells of abi1-1 and ost1-3 for ABA was restored in mutant plants by genetically, or exogenously, increasing PAP levels. In addition, PAP upregulated the expression of many ABA and Ca2+ responsive genes, including several CPKs. It was suggested (113) that because of the transcriptional regulation, PAP-mediated chloroplast signaling could bypass the canonical ABA signaling pathway and activate SLAC1. However, PAP-induced stomatal closure required sufficient concentrations of Ca2+ and apoplastic ROS production by NADPH oxidases, but did not affect the activity of SLAC1 or the highly selective inward-rectifying K+ channels KAT1 or KAT2 in X. laevis oocytes. This suggests that PAP is dependent on ABA-mediated processes and works rather as a second messenger in ABA signaling. Intriguingly, exogenous application of PAP on Arabidopsis and barley leaf peels was able to trigger stomatal closure within a few minutes and the kinetics of this reaction was almost identical to that of exogenous ABA application (113). It is highly unlikely that stomatal closure through transcriptional regulation would occur as fast as by ABA-triggered post-transcriptional regulation. Therefore, PAP may also regulate SLAC1 activity through direct post-transcriptional regulation of other kinases such as CPKs or MAPKs (Fig. 5).
The Role of Other Plant Hormones in Guard Cell Drought Response
In addition to ABA, also other plant hormones and low-molecular-weight compounds have a role in the induction of stomatal responses to drought and in the mediation of ROS-related or ROS-dependent signal transduction leading to stomatal closure. Jasmonic acid (JA) and it its methyl ester (methyl jasmonate or MeJA) induce ROS production and stomatal closure through the activation of RBOHD and/or RBOHF (134). MeJA-induced stomatal closure, ROS production, and cytosolic alkalization were unaffected in the pyr1 pyl1 pyl2 pyl4 quadruple mutant, but was impaired in the SnRK protein kinase OST1 loss-of-function mutants, ost1 and srk2e, and in the ABA-deficient aba2-2 mutant (160). This suggests that the MeJA activation of RBOHD and/or RBOHF requires ABA priming (as also implied by previous studies; 51, 100) and OST1 function, but does not activate OST1 through the canonical ABA signaling pathway in guard cells. JA and MeJA have been suggested to regulate stomatal closure through transcriptional regulation of MeJA responsive genes and through ROS- and nitric oxide (NO)-triggered, Ca2+-dependent activation of CPK6 and its downstream target SLAC1 (23).
SA accumulates in plant leaves during drought stress and pathogen invasion and induces stomatal closure in response to apoplastic superoxide production (84, 94). SA-induced apoplastic ROS accumulation around guard cells was inhibited by the application of the peroxidase inhibitor SHAM but not by the NADPH oxidase inhibitor diphenyleneiodonium (DPI) (61, 97). This suggests that the SA-induced apoplastic ROS production is mediated through the cell wall-bound peroxidases. In addition, the SA accumulating mutants siz1 (93), cpr5 (12), and acd6 (116) have constitutively decreased stomatal aperture and show drought tolerance. The application of peroxidase inhibitors SHAM and azide compromised the narrow stomatal phenotypes of the mutants, while the application of the NADPH oxidase DPI had no effect (93, 110). These results also imply that peroxidase-facilitated ROS production is involved in the SA-mediated, drought-induced stomatal closure. However, it must be noted that SHAM is not a specific inhibitor of peroxidases but has been more commonly used as an inhibitor of the mitochondrial alternative oxidase (AOX), which is activated under conditions involving increased mitochondrial ROS production (96). Furthermore, low (1–5 mM) concentrations of SHAM act actually as peroxidase activators, when only higher concentrations (20 mM) inhibit peroxidases (130); in some published studies, the use of low SHAM concentrations has been interpreted as an inhibitory effect. Also, the inhibitory effect of SHAM on AOX should not be overlooked as it has been suggested that AOX helps to maintain the NO homeostasis in guard cell mitochondria by preventing the over-reduction of the electron transport chain, particularly during stomatal closure when NO concentration increases in the cytosol (20). Therefore, the mechanism of SA-induced peroxidase activation remains to be verified by further studies. Low concentrations of SHAM (less than 5 mM was also used in these studies).
In contrast to ABA, JAs, and SA, all of which positively regulate stomatal closure, ethylene can promote both stomatal opening and closure, although the reaction seems to be highly species dependent (23, 100). In general, there is great inconsistence in the results from different studies on the effect of ethylene on stomatal regulation. One possible explanation to the differences could be that these studies have mainly been performed with leaf disks, epidermal peals, or detached leaves and experiments on these samples do not always reflect the real response to studied stimuli. In addition, the effect of ethylene on stomatal aperture seems to be dependent on hormonal homeostasis and the detachment of leaves will disrupt the cellular balance.
Two independent studies have shown that in the absence of ABA, ethylene promoted stomatal closure, whereas in the presence of ABA it inhibited stomatal closure in Arabidopsis (25, 135). In addition, auxin and cytokinin, the major plant hormones involved in various aspects of plant growth and development, inhibited the ABA-induced stomatal closure by enhancing ethylene biosynthesis (136). Ethylene-induced stomatal closure was also dependent on the RBOHF-mediated ROS production (25), whereas opening or inhibition of ROS-induced stomatal closure could be promoted by the ethylene-induced accumulation of flavonols (150). Flavonols are plant metabolites with antioxidant properties and they accumulate in guard cells reducing ROS levels and consequently suppress stomatal closure (150). Taken together, ethylene seems to affect guard cell signaling mainly by controlling ROS homeostasis in the guard cells and its function is controlled by other hormones.
MPKs Play Multiple Roles in the Regulation of Stomatal Movement
In addition to CO2 signaling, MPKs are also suggested to have a role in guard cell ABA and pathogen signaling (22, 74). Whereas MPK9 and MPK12 were involved in the stomatal responses to ABA, MPK3 and MPK6 mediated pathogen signaling in guard cells (95). As discussed earlier, ROS are produced by RBOHs in response to both ABA and pathogen signaling, but while RBOHF is mainly responsible for the ROS production in response to ABA, RBOHD is involved in stomatal closure in response to recognition of potentially pathogenic microorganisms (60, 77). It would be tempting to speculate that the two NADPH oxidases, RBOHD and RBOHF, are regulating two separate MPK pathways, but recent research has indicated that reality is more complicated.
The activation of RBOHD was not required for the activation of MPK3 and MPK6 in response to bacterial pathogens (156). Moreover, it has been suggested that the rapid ROS burst and the activation of MPK3/MPK6 are two independent early signaling events during stomatal immune response in Arabidopsis.
More recently, these two signaling events were shown to belong to separate but interdependent signaling cascades that control stomatal movements (Fig. 6), and the loss-of-function of both MPK3 and MPK6 impaired pathogen-triggered stomatal closure (132). The activation of MPK3 and MPK6 was independent of the ABA-, SLAC1-, and RBOHD-mediated ROS burst. Instead of regulating anion channels, the two kinases controlled the metabolism of osmotically active organic acids such as malate and citrate. Under pathogen attack, the level of osmotically active metabolites in the cytosol decreased and the guard cell turgor was lost promoting stomatal closure. However, at the same time, the ABA-induced ROS production by RBOHD activated ABA signaling, leading to SLAC1 activation and stomatal closure (132). To what extent these interdependent signaling cascades interact and whether they share common mediators remain to be elucidated.
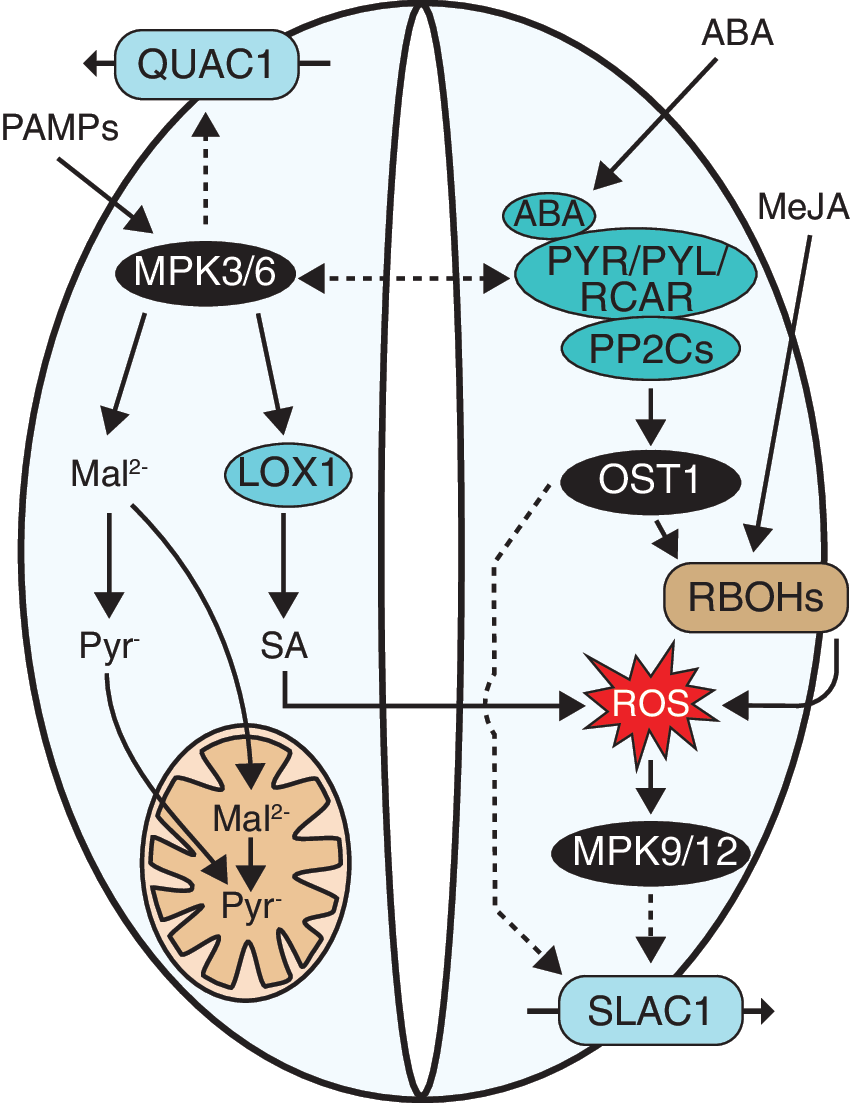
To further complicate the story, MPK3 and MPK6 have been suggested to regulate stomatal closure also through an ABA-independent oxylipin pathway (95). MPK3 and MPK6 activated guard cell-specific lipoxygenase 1 (LOX1), and SA was needed for the downstream signaling events leading to stomatal closure.
Both MPK9 and MPK12 are also involved in SA-mediated stomatal signaling in guard cells as SA activated S-type anion channels and elicited stomatal closure in wild-type Arabidopsis but not in the mpk9 mpk12 double mutant (62). It was suggested that the two kinases could be involved in the same signaling cascade through LOX1. However, the studies on MPK9 and MPK12 on ABA- and SA-mediated stomatal regulation have been performed mainly with TILLING mutants of mpk9-1 and mpk12-1 (containing, in addition to the mutations in MPK9 and MPK12, an undetermined number of point mutations elsewhere in the genome) and epidermal peels or guard cell protoplasts (59, 62). Point mutations can affect the protein function in different ways when compared to loss-of-function mutants. Similarly, experiments performed with epidermal peels or protoplasts are missing the mesophyll contact, as discussed earlier in the text. Therefore, the involvement of MPK9 and MPK12 in stomatal regulation by ABA and SA would require experiments with especially loss-of-function alleles and with intact plants to evaluate their role in stomatal processes.
The above studies on MAPK3/MAPK6 signaling cascades were focusing on pathogen-triggered stomatal closure. However, MPK3 and MPK6 are activated by both biotic and abiotic stresses, as well as by ABA (22). Decreased expression of MPK3 by guard cell-specific gene silencing resulted in impaired ABA-mediated inhibition of stomatal opening and H2O2-induced stomatal closure, but did not affect the ABA-induced stomatal closure (39). In addition, the mpk6 mutant guard cells were impaired in ABA-induced H2O2 accumulation (155). Taken together, it seems likely that MPK3/MPK6-regulated organic acid metabolism would also have a role in stomatal responses to abiotic stresses such as drought. However, this needs to be verified by testing the stomatal responses of the mpk3 mpk6 double mutant to abiotic stresses.
Negative Regulation of ABA Signaling
Stomata are generally considered to respond to abiotic and biotic stresses by decreasing their aperture. However, it is important to note that during the day, C3 and C4 plants rarely close their stomata completely. Instead, they have developed negative regulatory mechanisms to ensure minimal CO2 supply for photosynthesis by keeping stomata open during stress as well. As discussed earlier, ethylene negatively regulates ABA signaling in guard cells. In addition to hormonal regulation, cytoplasmic nitrosylation reactions are involved in the negative regulation of ABA signaling.
The ABA-dependent rapid accumulation of NO negatively regulated the OST1 function by S-nitrosylation of Cys137 near the catalytic site of the kinase (146). The S-nitrosylation of OST1 was observed as a late event in the ABA signaling, and thus, it has been suggested that this mechanism helps to reset ABA signaling. Considering the role of OST1 in the activation of RBOHF, it has been further suggested (128) that inhibition of OST1 by NO might also restrict ROS formation.
Cytoplasmic ROS participate also in the negative feedback regulation of CPK21 (139). Oxidation of CPK21 by H2O2 resulted in the formation of intramolecular disulfide bond that reduced the kinase activity. Conversely, CPK21 was activated by a thioredoxin H-type 1 (Trx-h). Thioredoxins are small proteins that catalyze the thiol to disulfide exchange reaction in their target proteins. Incubation of the oxidized CPK21 together with the Trx-h rescued the kinase activity, suggesting that CPK21 could be subjected to redox regulation under changing conditions (139). Furthermore, during stress, the inactivation of CPK21 by H2O2 could act as a negative feedback regulation of ABA-induced stomatal closure. It would be interesting to see if other CPKs are regulated in a similar manner.
Connection Between CO2 and ABA Signaling in Guard Cells
Since both ABA- and high CO2-induced stomatal closures involve activation of SLAC1 in guard cells, one could expect that ABA and CO2 signaling converges. Indeed, several mutations causing stomatal ABA insensitivity, such as abi1-1 and abi2-1 (86, 151) as well as ost1 and ghr1 (49, 86, 157; Fig. 4), display impaired stomatal responses to high CO2 concentrations. Stomata of the growth-controlled abscisic acid 2 (GCA2) mutant, gca2, which is related to CPKs, were insensitive to both ABA and high CO2 concentration. Furthermore, gca2 displayed altered patterns of cytoplasmic Ca2+ transients in response to these stimuli and was suggested as a convergence point between CO2 and ABA signaling (4, 161).
ABA receptors, PYR/PYL/RCARs, are also involved in CO2 signaling, as inactivation of several of these proteins impaired stomatal closure in elevated CO2 concentrations (17, 86). Due to a large number of PYR/PYL/RCAR proteins and a functional redundancy between them, further research is required to identify which combination of PYR/PYL/RCARs functions in the regulation of CO2-induced stomatal closure (17, 86, 157). Furthermore, recently developed fluorescent probes that enable real-time in vivo monitoring of ABA in plant cells (144) should be used in addressing the interplay between CO2 and accumulation of ABA in guard cells during changes in CO2 concentration (28).
Although several key components of ABA signaling are also connected with stomatal responses to high CO2 concentration, ABA-independent components also exist. ABA-induced stomatal closure was completely functional in the mutants of HT1 and MPK12, whereas these plants were deficient in CO2-controlled stomatal movements (49, 58). Moreover, experiments aimed to dissect which parts of the SLAC1 anion channel are important for ABA- and which for high CO2-induced stomatal closure showed that transgenic plants expressing SLAC1 anion channel without both C- and N-terminal regions were still able to respond to changes in CO2 concentration, but remained ABA insensitive. Thus, ABA-induced activation of SLAC1 seems to involve C- and N-terminal regions of the SLAC1, whereas CO2-induced stomatal closure seems to rely only on the transmembrane region (158).
The overlap between CO2 and ABA signaling suggests that ROS production in guard cells can increase in response to high CO2 concentration, similar to ABA-induced stomatal closure (Fig. 3B). Using a fluorescent probe H2DCF-DA, ROS accumulation was indeed observed in guard cells treated with HCO3 − or high CO2 concentration (17, 64, 125). Moreover, ROS scavengers impaired stomatal closure induced by CO2 (17, 64). A connection between CO2 and ABA signaling was further proved by the absence of ROS accumulation in stomata under elevated CO2 concentration in the ABA-deficient double mutant nced3 nced5, as well as in the triple pyr1 pyl1 pyl4 and the quadruple pyr1 pyl1 pyl2 pyl4 mutants (17). Similar to ABA, elevated CO2 induced ROS formation by NADPH oxidases (17, 125; Fig. 3). Thus, the rbohD rbohF double mutant demonstrated insensitivity of guard cells to HCO3 −/high CO2 concentration. These mutants also failed to produce ROS in guard cells in response to elevated CO2 (17, 64). Impaired accumulation of ROS in guard cells and decreased stomatal closure in response to high CO2 concentration were also observed in the tomato mutant rboh1 (125).
The current knowledge about high CO2-induced stomatal closure suggests at least three partially overlapping pathways. (i) Signaling through HT1/MPKs, which is ABA independent and is triggered by increased HCO3 − in guard cells (49, 58). (ii) Direct perception of HCO3 − by SLAC1 in the presence of protein kinases that activate SLAC1 (145). (iii) An ABA-dependent component that partially mediates high CO2-controlled stomatal closure (17, 86, 157). ABA signaling that activates OST1 and CPKs by suppression of PP2Cs could enhance SLAC1 sensitivity to HCO3 −, as well as directly trigger SLAC1 anion currents, although this hypothesis should be verified in the future. It is possible that plants under water stress should react to increased Ci faster and stronger than plants with satisfactory water supply to save water in leaves when CO2 supply for mesophyll cells is sufficient. This could explain the importance of ABA signaling for CO2-controlled stomatal movements, which would allow plants to adapt to changing environmental conditions.
Future Perspectives
Recent research has highlighted the complex interplay between apoplastic, cytoplasmic, and chloroplastic redox/ROS signaling, as well as hormonal regulation in the control of stomatal aperture. However, major gaps remain in the understanding of complex interactions within the guard cell signaling networks in response to changes in CO2 and water availability. Considerable efforts are needed for understanding how guard cells regulate, and are regulated by mesophyll photosynthesis. The outstanding key questions are related to how guard cells perceive and transmit signals from the surrounding environment and mesophyll cells. Furthermore, identification of proteins that can sense changes in HCO3 − and ROS in guard cells is also needed.
In the future, major breakthroughs will most likely come from the development of tools that enable real-time imaging of the cellular localization of ROS in guard cells in response to various stimuli. The focus of future research should be directed to understand the complex interactions between various guard cell signaling pathways, and how the guard cell hormones, ROS, and Ca2+ homeostasis modulate these interactions. In addition, the translation of such knowledge from model plants to important crop species, especially to those grown in arid areas, will be increasingly important in the near future.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist.