
Abstract
Significance:
Redox signaling is a common mechanism in the cellular response toward a variety of stimuli. For analyzing redox-dependent specific alterations in a cell, genetically encoded biosensors were highly instrumental in the past. To advance the knowledge about the importance of this signaling mechanism in vivo, models that are as close as possible to physiology are needed.
Recent Advances:
The development of transgenic (tg) redox biosensor animal models has enhanced the knowledge of redox signaling under patho(physio)logical conditions. So far, commonly used small animal models, that is, Caenorhabditis elegans, Drosophila melanogaster, and Danio rerio, and genetically modified mice were employed for redox biosensor transgenesis. However, especially the available mouse models are still limited.
Critical Issues:
The analysis of redox biosensor responses in vivo at the tissue level, especially for internal organs, is hampered by the detection limit of the available redox biosensors and microscopy techniques. Recent technical developments such as redox histology and the analysis of cell-type-specific biosensor responses need to be further refined and followed up in a systematic manner.
Future Directions:
The usage of tg animal models in the field of redox signaling has helped to answer open questions. Application of the already established models and consequent development of more defined tg models will enable this research area to define the role of redox signaling in (patho)physiology in further depth. Antioxid. Redox Signal. 29, 603–612.
Introduction
R
Genetically encoded biosensors have been applied for in vitro and in vivo studies. Cell lines are simple and powerful tools to study general cellular principles in the laboratory routine. Serial passaging of cell lines can cause genotypic and phenotypic variations over time. Therefore, researchers try to analyze primary cells whenever possible (21). As a matter of fact, data generated from either stable cell lines or primary cell culture might, however, lack important information closer to the in vivo situation. Intercellular communication between different cell types, biophysical properties, and local changes in the tissue environment such as pH, pO2 etc. are just some arguments for the need to study cells within the tissue context to reflect the physiological importance of the data analyzed. To this end, introducing genetically encoded biosensors as a transgene into a living organism is a new technical avenue in the field of redox biology.
Genetically Encoded Redox Biosensors
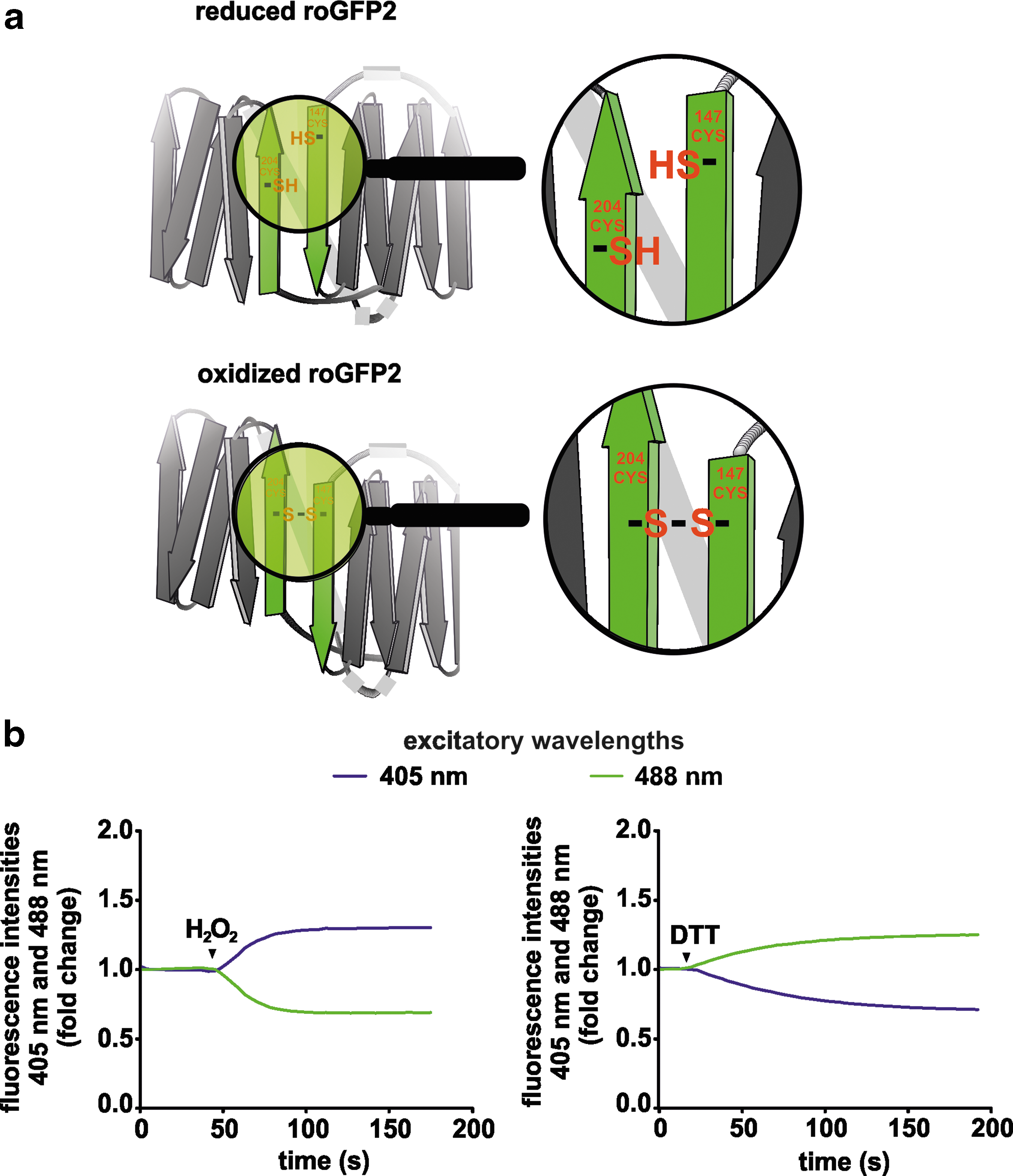
Early optical redox sensors relied mainly on small molecules changing their fluorescence behavior by reacting with the target of interest (27). The further development of imaging-based sensor techniques benefited from the cloning and recombinant production of fluorescent proteins. The green fluorescent protein (GFP) is composed of 238 amino acid residues (26.9 kDa) that exhibit bright green fluorescence when exposed to light in the blue to ultraviolet (UV) range (38). GFP was first isolated from the jellyfish Aequorea victoria. The fluorescence characteristic of GFP is without need of any co-factor, which makes it a suitable reporter protein. The groundbreaking importance for the biosciences made by the achievements regarding the characterization of GFP was honored by awarding the Nobel prize in chemistry in 2008 “for the discovery and development of the green fluorescent protein, GFP” to Osamu Shimomura, Martin Chalfie, and Roger Y. Tsien (11). Many mutations have been engineered in the GFP sequence, including some that affect color, pH stability, etc. The establishment of the reduction-oxidation sensitive (ro) GFP was highly instrumental for the redox biology field (19). roGFP (reduction-oxidation sensitive GFP) is a GFP variant in which surface-exposed cysteines are added to the molecule (Fig. 1a). roGFP, similar to wild-type GFP, has two excitation peaks (at ∼400 and ∼480 nm) and one emission peak at ∼510 nm. Oxidation of the cysteine residues in roGFP results in an increase in excitation at ∼400 nm (Fig. 1b). Reduction of those cysteines favors excitation at ∼480 nm. Two major roGFP variants, that is, roGFP1 and roGFP2, are commonly used. roGFP1 is derived from the wild-type chromophore with the amino acid S65 in the chromophore. It displays a dominant excitation peak in the UV range. The roGFP2 variant derived from enhanced GFP (EGFP) contains an S65T mutation and has a main excitation peak at 490 nm (19). The roGFP alone has a slow response to redox changes. To improve the specificity of the signal and the reaction rate, glutaredoxin-1 (Grx1) was fused to roGFP, which results in a rapid equilibrium between the glutathione (GSH)/glutathione disulfide (GSSG) couple and, thus, reflects the glutathione redox potential (E GSH) (16). Other genetically encoded fluorescence protein-based biosensors were designed to sense specifically hydrogen peroxide (H2O2), which is one of the main ROS generated by cells. The fusion of the yeast peroxidase (Orp1) to roGFP2 and the hydrogen peroxide sensor (HyPer) are examples for H2O2-sensing probes. HyPer is a fusion protein of the circularly permutated yellow fluorescent protein (cpYFP) and the H2O2 sensitive bacterial protein OxyR (4, 5, 12). Its fluorescence is highly pH sensitive, and measurements with HyPer should, therefore, always include simultaneous pH recordings. A more detailed overview about the available genetically encoded redox biosensors, including benefits and limitations, is given in a separate manuscript in this Forum.
Genetically Modified In Vivo Redox Biosensor Models
Genetically modified plant or animal models represent the most efficacious method of functional gene analysis. For overexpression of a gene in plants, nematodes, fish, and mice, transgenesis is commonly applied. These model organisms were also used to generate transgenic (tg) redox biosensor models in the past (Fig. 2). In this review, we will summarize the results of the so far established redox biosensor models with a major emphasis on the mouse models described.

Redox Imaging in Plants, Insects, Nematodes, and Fish
Plant (Arabidopsis thaliana)
Plants can frequently encounter different forms of environmental stress. Maintenance of the redox balance is vital for successfully responding to a change in the environment (30). tg expression of roGFP1 and roGFP2 in the model plant A. thaliana proved to be a powerful tool to monitor redox changes in vivo in different subcellular localizations in real time (22, 29). Most interestingly, the average redox potential determined with roGFP either targeted to the mitochondria or expressed in the cytosol was found to be distinct in the roots, demonstrating that plants maintain a subcellular-specific redox homeostasis (22, 40). Moreover, the elongation zone of the Arabidopsis root was found to be more oxidized than either the root cap or meristem (22). Compartment and subcellular-specific redox characteristics were further extended to organelle-specific oxidation by analyzing roGFP targeted to chloroplasts, mitochondria, peroxisomes, and endoplasmic reticulum (7). The established methodology in Arabidopsis was subsequently used for understanding compartment and organelle-specific responses to different stress forms. It allowed analyzing the response toward nitrogen, drought, water, extended darkness, and salt stress (7, 25, 36, 37, 40). Employing redox biosensor Arabidopsis models, thus, helped to uncover compartment and subcellular redox potentials.
Insect (Drosophila melanogaster)
tg Drosophila flies were established by employing the GSH (Grx1-roGFP2 or the inverted domain order roGFP-Grx1) and H2O2 (Orp1-roGFP2) biosensors with either cytosolic or mitochondrial localization under the control of the tubulin promoter. Living larvae can be imaged in vivo based on their transparency during development. For imaging adult flies, preparations of the multicellular non-transparent tissues need to be performed. Albrecht et al. could elegantly demonstrate that during tissue preparations a non-specific oxidation of GFP-based biosensors can occur (1). This can be prevented by adding N-ethylmaleimide (NEM) during sample preparation. NEM is a thiol-blocking agent that shields thiol groups from oxidation. Having solved the problem of non-specific oxidation, the authors went on to image the different roGFP tg fly models. They demonstrate both tissue-specific and cellular compartment-specific changes in the GSH redox state and H2O2 levels as well as the involvement of redox alterations for aging processes (1, 2). Comparing the results obtained with the GSH-specific Grx1-roGFP and the H2O2-specific Orp1-roGFP biosensors revealed that changes in the GSH redox state and changes of H2O2 levels are not necessarily coupled (1). This is important evidence, which demonstrates that the GSH pool can be regulated independently from altered H2O2 production in living organisms.
Nematode (Caenorhabditis elegans)
HyPer and Grx1-roGFP2 tg C. elegans under the control of the large ribosomal subunit L17 promoter were one of the first redox biosensor nematode models that allowed to detect distinct redox properties in vivo (3). The successful establishment of these models resulted in uncovering regions with distinct H2O2 production and GSSG/2GSH ratios as well as the characterization of redox mechanisms during development and aging (26). Subsequently, a mitochondria-targeted roGFP1 C. elegans strain as well as roGFP1-expressing strains with a restricted expression in pharyngeal muscle (myo-2 promoter), intestine (sur-5 promoter), or mechanosensory neurons (mec-4 promoter) were developed and permitted the analysis of subcellular compartments or compartments of organs (23, 35). These studies demonstrated a spatial organization of the E GSH at the tissue and sub-tissue level.
Zebrafish (Danio rerio)
The transparency of tg zebrafish makes this model highly useful for bioimaging. In zebrafish, mainly different versions of HyPer (6, 18, 31) and Grx1-roGFP2 (41) were used for transgenesis and helped to elucidate mechanisms of leukocyte recruitment to a site of sterile inflammation as well as heart regeneration. These results were reviewed recently elsewhere (10).
Redox Biosensor tg Mouse Models
Generation and application of tg redox biosensor-expressing mice have been developed as a powerful technical tool to analyze redox signaling in the past years. They allow semi-quantitative analysis in complex tissue preparations (Table 1). Analyzing biosensor signals in intact mice is challenged by the necessity to apply suitable imaging techniques, including confocal, two-photon, or wide field microscopy imaging techniques. Based on the size of a mouse and the wavelengths of the biosensors involved, it is, however, still difficult to image internal organs in situ. Isolation of cells, tissue slices, or isolated and ex vivo perfused organs are alternatives that are as close as possible to the in vivo situation. To this end, it is understandable that brain slices, isolated neurons or cardiomyocytes, Langendorff-perfused hearts, and the skin are the most frequently analyzed cells/tissues applying genetically modified redox biosensor mice up to now.
Table gives an overview of the up-to-date developed redox biosensor mouse models, their application in terms of redox imaging, and the observed phenotype.
CMV, cytomegalovirus; EF1α, elongation factor 1α; E GSH, glutathione redox potential; Grx1, glutaredoxin-1; roGFP, reduction-oxidation sensitive green fluorescent protein; UVA, ultraviolet A.
A recently developed redox histology protocol, in which the in situ redox state is fixed by applying NEM on cryosections and is quantified by calibration with diamide and dithiothreitol (DTT), is a further important milestone in the usage of mice as a “redox” model organism (14). Redox histology in combination with confocal microscopy allows imaging of the biosensor in histological sections. Adjacent cryosections of a snap-frozen tissue are either left untreated or treated with NEM, DTT, or diamide. DTT- and diamide-treated sections allow visualization of the biosensor in its fully reduced and oxidized form, respectively. The DTT and diamide response should be carefully titrated to ensure the full oxidation and reduction response. The NEM-treated cryosection makes an insight into the actual redox status of the biosensor within the tissue possible. A comparison of the ratios of the fluorescence intensities obtained from the NEM-treated cryosection with the data obtained from the DTT- and diamide-treated samples allows semi-quantitative insights into redox signaling at the tissue levels.
The protocol was developed as an endpoint experiment. Analyses at different time points after adding DTT or diamide within the same sample are, therefore, not possible. The redox histology protocol was successfully applied to healthy, non-diseased, and malignant tissue samples. The biosensors tested with this protocol include roGFP2-Orp1 as well as Grx1-roGFP2. The Grx1-roGFP2 was driven by the neuronal Thy1 promoter, whereas roGFP2-Orp1 was driven by the artificial CAG promoter in the first description of the newly established protocol (14). The CAG promoter is a strong synthetic promoter containing the cytomegalovirus (CMV) early enhancer element, the first exon and intron of the chicken α-actin gene, and the splice acceptor of the rabbit β-globin gene, which are frequently used to drive high levels of gene expression in mammalian expression vectors.
Cells with a typical histological signature are easily detectable in tissue cryosections. An example of redox-histology staining using cardiomyocyte-specific expression of Grx1-roGFP2 located to the mitochondrial matrix is shown in Figure 3a and b. The mitochondrial signals in the tg cardiomyocytes are visible. The fluorescence is restricted to one cell type due to the cell-type-specific promoter. Within the organ level, however, several cell types exist. This explains the necessity to further develop a possibility to distinguish between different cell types, for example by immunohistochemistry techniques in combination with the redox histology protocol. This would enable researchers to unequivocally detect the response of a specific cell type when using a ubiquitously active non-cell-type restricted promoter.

In the medical sciences, the refined development of tg biosensor mouse models and respective imaging techniques is of utmost importance since the mouse is still an important model organism for many relevant diseases. Genetically modified mice and experimental interventions mimicking relevant diseases have been developed. The combination of tg redox biosensor mouse models crossed to a wide range of available knock out mice or applied to disease models is, thus, the next logical step. This could help to advance the knowledge about the involvement of specific redox signaling pathways in physiology and pathology within the next years. The already established tg biosensor mouse models using either a restricted cell-type-specific or a ubiquitous promoter prove that they are a sound basis for studying the importance of redox signaling in health and disease.
Neuron-Specific roGFP Mice
Various neuron-specific roGFP-based mouse models have been established. They differ in the biosensor protein used, the promoter-driven-specific cellular expression, and the subcellular compartment analyzed.
Tyrosine hydroxylase promoter-driven roGFP2 tg mice
Calcium entry can affect the mitochondrial oxidation status in autonomous pacemaking cells. This is of special importance for the disease progression of Parkinson's disease (PD), which is a neurodegenerative disorder characterized by a selective loss of dopaminergic neurons in the substantia nigra. A decreased viability in the dopamine-producing cells is associated with increased mitochondrial ROS and energy deficiency (34). Guzman et al. generated a tyrosine hydroxylase promoter-driven tg mouse model as well as control mice, in which the tg expression of roGFP2 is driven by the CMV promoter (17). The roGFP2 biosensor is targeted to the mitochondrial matrix in both models. The tyrosine hydroxylase promoter results in a restricted expression of the roGFP2 in dopamine-producing cells, whereas the CMV promoter allows ubiquitous expression. Oxidation of the biosensor in neurons was analyzed by two-photon laser scanning microscopy employing midbrain slices. Relative oxidation of the sensor was analyzed after fully reducing or oxidizing the cells with DTT and aldrithiol, respectively. This strategy allowed comparing the relative sensor oxidation in dopaminergic neurons in the substantia nigra compacta and in the ventral tegmental area as well as in cholinergic neurons in the dorsal motor nucleus of the N. vagus. These cells are affected during the onset and progression of PD. The results of the roGFP2 analyses revealed that autonomous pacemaking cells in the PD related areas share a functional relationship between membrane depolarization, spike activity, calcium entry, and low calcium buffering capacity (15).
Calcium must be extruded from the pacemaking cells under consumption of ATP, which is produced during oxidative phosphorylation in the mitochondria. Using the mouse models described earlier, the authors were able to convincingly show that calcium entry in the cells, indeed, affected the oxidation status of the mitochondrial matrix. This effect was further enhanced in a loss-of-function mutation of DJ-1, a gene associated with early onset form of PD, which points to a functional involvement in the pathogenesis of this disease (17).
In contrast to Grx1-roGFP2, non-coupled roGFP2 is in equilibrium with various other redox couples and, thus, is not suitable, for example to measure specific ROS or the E GSH. It is important to note that the redox status of roGFP, thus, is not necessarily equivalent to the increased production of ROS. Nonetheless, the CMV-roGFP2-expressing mouse model was used in a later study to analyze the importance of mitochondria-derived ROS for antigen-specific T cell activation (42).
Thy-1 promoter-driven roGFP tg mice
Two independent groups generated thymocyte differentiation antigen/CD90 (Thy-1.2) promoter-driven roGFP-based tg mouse models (9, 44). Breckwoldt et al. used Grx1-roGFP2 targeted to the mitochondrial matrix, whereas Wagener et al. employed the uncoupled roGFP1 either located to the cytoplasm or targeted to the mitochondrial matrix. Since Grx1-roGFP2 and roGFP1 pick up the specific GSH redox couple versus the more general cellular redox balance, the analyses are not directly comparable. Breckwoldt et al. used their mouse model for analyzing the mitochondrial redox potential in different neurological diseases, including spinal cord injury and amyotrophic lateral sclerosis. On injury, they observed an influx of extracellular Ca2+, which initiated a permanent mitochondrial GSH oxidation that spread from the injury site along the affected axons. This was followed by mitochondrial shortening and fragmentation. Spontaneous redox signals in neurons at the single mitochondrion level where transients of GSH oxidation go along with shortening and re-elongation of the organelle were also observed in healthy mitochondria, which further increased on stimulating electrical activity (8).
Wagener et al. focused in their study on mapping redox conditions at the subcellular level by concentrating on comparing the roGFP1 signals expressed in either the cytosol or the mitochondrial matrix (44). They calibrated the sensor response in both compartments by treating hippocampal slices with 10 mM DTT or 5 mM H2O2. Based on this calibration, they determined the relative oxidation of the roGFP and quantified the redox potential. This revealed a significantly more oxidized E roGFP1 in the mitochondrial matrix compared with the cytosol in various brain regions. A striking difference was additionally detected by comparing the soma and the dendrites with a more oxidized roGFP1 in the soma (Table 1).
Erythrocyte-Specific roGFP2
Although oxygenated hemoglobin [Hb(FeII)O2] is considered a relatively stable molecule, it can physiologically auto-oxidize to methemoglobin [Hb(FeIII)]. Auto-oxidation is almost entirely responsible for ROS generation inside red blood cells. Erythrocytes are exposed to a wide variety of different pO2 levels in the venous and arterial blood, which renders the erythrocytes prone to redox modifications. Xu et al. developed tg mice, which express roGFP2 driven by a β-globin mini-promoter (47). Around 50% of peripheral red blood cells expressed roGFP2 in this mouse line. In unperturbed tg red blood cells, a E roGFP2 of −318.5 ± 8.07 mV was calculated based on the degree of roGFP2 oxidation after treatment with 1 μM H2O2 and 10 mM DTT. The pro-hemolytic compound phenylhydrazine resulted in a dose-dependent increase of roGFP oxidation. A similar effect was observed as part of red blood cell aging, implying that oxidative alterations contribute to red blood cell senescence and damage.
Polypeptide Chain Elongation Factor 1α Promoter-Driven roGFP
The polypeptide chain elongation factor 1α (EF1α) promoter in combination with the roGFP1 biosensor was used in tg mice for monitoring oxidative stress in vivo (46). RoGFP expression driven by the EF1α promoter was found in various organs. In the skin, the expression was mainly restricted to keratinocytes, demonstrating that the roGFP1 oxidation status can be tested in the skin of this tg mouse model as fluorescence of these cells is optically accessible in freely moving mice. The established mouse model is, thus, one of the rare examples for monitoring roGFP in vivo non-invasively. To this end, the authors applied three light-emitting diodes turned on sequentially and synchronized to a charge-coupled device camera. The contribution of autofluorescence, however, was significant when using C57BL/6J mice in this setup and demonstrates one current drawback of applying genetically encoded biosensors in vivo. The authors were able to reduce the autofluorescence problem, in part, by using albino hairless mice, decreasing the autofluorescence generated by the fur and melanin.
Cardiomyocyte-Specific roGFP-Based tg Mice
αMHC promoter-driven cyto and mito Grx1-roGFP2 mice
Various cardiovascular diseases have been strongly associated with an altered redox signaling (39). Especially, the ROS-generating NADPH oxidase enzyme complexes 2 and 4 are regarded to be of importance for detrimental or protective effects during the course of heart failure development in consequence of ischemia, mechanical load, etc. (48). The intracellular compartmentalization and organization of cardiomyocytes is dominated by the sarcomere units. Contractile activity depends on mitochondrial ATP production. Cellular metabolism, cellular redox signaling, and excitation contraction coupling are, thus, inevitably linked.
The recently established Grx1-roGFP2 tg mice, in which the transgene is driven by the cardiomyocyte-specific αMHC promoter, allowed gaining insights into the changes of the cellular E GSH in resting cardiomyocytes and after stimulation (43). Since Grx1-roGFP2 was either located in the cytoplasm or targeted to the mitochondria, the E GSH was compared in these two subcellular compartments. Most interestingly, the mitochondrial matrix was found to be more reduced (E GSH −278.9 mV) compared with the cytoplasm (E GSH −254.8 mV) (Table 1). This difference observed in isolated cardiomyocytes was further confirmed by analyzing Langendorff-perfused whole hearts. Intriguingly, the difference between the E GSH in the cytoplasm and the mitochondrial matrix in cardiomyocytes is in sharp contrast to the redox potential as determined in the Thy-1 promoter-driven roGFP1 tg mice described above. The discrepancy might be due to the different sensors, that is, Grx1-roGFP2 versus roGFP1, applied or is due to the different cell types, that is, neurons versus cardiomyocytes, analyzed. Stimulation of the cardiomyocytes isolated from the Grx1-roGFP2 tg mice with isoprenaline or angiotensin II demonstrated that the E GSH in the cytoplasm and the mitochondrial matrix responds with a specific pattern in each compartment.
The formation of cellular ROS and alterations in redox signaling are linked to aging and age-related diseases (20). Because of conflicting data, the link between ROS and aging, however, is not exactly clear. In some aging studies, ROS are found to decrease lifespan and in other studies, they are found to act as a pro-survival factor (45). Regarding the E GSH, mostly a pro-oxidant shift accompanied by an elevation of protein-GSH mixed disulfides have been previously reported in D. melanogaster (32) and different tissues of mice (33). However, these studies lack information about the E GSH in different subcellular compartments. Tg redox biosensor animal models make it possible to answer open questions like these by following the biosensor response in aging mice. With increasing age, mice develop an impaired heart function. In case of the αMHC Grx1-roGFP2 mice expressing either the sensor in the cytoplasm or the mitochondrial matrix, a significantly impaired fractional area shortening and ejection fraction is visible at an age of more than 80 weeks (Fig. 4a). The E GSH in isolated cardiomyocytes from young versus old mice was analyzed by treating the cells with H2O2 or DTT. Based on the maximum oxidation and the maximum reduction response of the Grx1-roGFP2 biosensor, the E GSH was calculated as described earlier (43). The E GSH of cardiomyocytes isolated from the aged mice compared with young animals revealed significant differences. The E GSH in the cytoplasm of the aged mice was significantly reduced compared with the young animals. Interestingly, this was accompanied by no change in the mitochondrial matrix, demonstrating that both compartments respond independently from each other. The strict difference of the E GSH in the cytoplasm and the mitochondrial matrix as seen in the young animals was lost with aging. This data set demonstrates how tg redox biosensor animal models can be used to define the role of redox signaling in (patho)physiology in further depth.

As outlined earlier, multiple tg biosensor organisms have been developed and have been employed to explore spatiotemporal changes of redox signaling in multicellular organisms. More cell-type-specific characteristics in redox signaling are worthy of being analyzed at the tissue level in the future. These analyses will rely on the further development of more cell-specific tg biosensor mice and other model organisms. However, there are several technical issues, which still need to be solved. Redox biosensor responses in tg animal/plant models cannot be easily quantified. So far, only redox potentials of GSH (Grx1-roGFP) and roGFP1/2 have been quantified in mouse models. This is partly due to the fact that calibration of the fluorescence signal in complex tissue preparations is difficult. Careful titration of the concentration of H2O2 or diamide and DTT needs to be performed to determine the concentrations required to achieve full oxidation or reduction of the biosensor, respectively. Such experiments cannot be conducted for all tissues in vivo. Further, in all experiments, a strict control for autofluorescence needs to be done. Moreover, not all tissues can be easily accessed by the applicable imaging techniques in vivo based on insufficient penetration depth. If sample or tissue preparation is needed to image otherwise inaccessible parts, a non-specific artificial oxidation during sample preparation needs to be prevented. Regarding pH-sensitive redox biosensors such as HyPer, possibilities to monitor pH changes or pH control biosensors simultaneously need to be considered.