
Abstract
Aims:
Nicotinamide adenine dinucleotide phosphate oxidases (NOX-es) produce reactive oxygen species and modulate β-cell insulin secretion. Islets of type 2 diabetic subjects present elevated expression of NOX5. Here, we sought to characterize regulation of NOX5 expression in human islets in vitro and to uncover the relevance of NOX5 in islet function in vivo using a novel mouse model expressing NOX5 in doxycycline-inducible, β-cell-specific manner (RIP/rtTA/NOX5 mice).
Results:
In situ hybridization and immunohistochemistry employed on pancreatic sections demonstrated NOX5 messenger ribonucleic acid (mRNA) and protein expressions in human islets. In cultures of dispersed islets, NOX5 protein was observed in somatostatin-positive (δ) cells in basal (2.8 mM glucose) conditions. Small interfering ribonucleic acid (siRNA)-mediated knockdown of NOX5 in human islets cultured in basal glucose concentrations resulted in diminished glucose-induced insulin secretion (GIIS) in vitro. However, when islets were preincubated in high (16.7 mM) glucose media for 12 h, NOX5 appeared also in insulin-positive (β) cells. In vivo, mice with β-cell NOX5 expression developed aggravated impairment of GIIS compared with control mice when challenged with 14 weeks of high-fat diet. Similarly, in vitro palmitate preincubation resulted in more severe reduction of insulin release in islets of RIP/rtTA/NOX5 mice compared with their control littermates. Decreased insulin secretion was most distinct in response to theophylline stimulation, suggesting impaired cyclic adenosine monophosphate (cAMP)-mediated signaling due to increased phosphodiesterase activation.
Innovation and Conclusions:
Our data provide the first insight into the complex regulation and function of NOX5 in islets implying an important role for NOX5 in δ-cell-mediated intraislet crosstalk in physiological circumstances but also identifying it as an aggravating factor in β-cell failure in diabetic conditions.
Introduction
Reactive oxygen species (ROS) are key modulators of islet insulin production and release. As for the sources of these radicals, the members of the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX-es) family emerged as of particular interest due to their strict intracellular compartmentalization, regulated ROS production, and their capacity to provide a link between cellular metabolism/energy state (NADPH) and ROS production (45, 59). NOX enzymes are essential regulators of physiological insulin secretion but may exert harmful effects when chronically overactivated (2, 18, 38, 42 –45). The NOX family comprises five isoforms (NOX1–5) and two dual oxidases (DUOX1–2) (35). In rodent and human pancreatic islets, the expressions of NOX1, NOX2, and NOX4, and the accessory subunits NOXA1, p22 phox , p47 phox , and p67 phox have been confirmed (2, 23, 38, 53, 64). NOX5 is a particular member of the NOX family as it is expressed in diverse human tissues and cell lines but lacks a homolog in rats and mice (4, 5, 7, 11, 16, 25). The presence of NOX5 messenger ribonucleic acid (mRNA) in human islets has been reported in recent studies using reverse transcription polymerase chain reaction (RT-PCR), real-time polymerase chain reaction (PCR), and RNA sequencing; however, these studies did not conduct any follow-up investigations to establish the cell-type-specific expression or the pathophysiological significance of NOX5 (2, 38). A possible link between NOX5 and islet function was suggested by gene expression profiling that showed elevated NOX5 mRNA levels in islets of type 2 diabetes mellitus (T2DM) patients compared with islets of nondiabetic subjects (Geoprofiles GDS3882/220641_at).
Innovation
Islet cells contain different nicotinamide adenine dinucleotide phosphate oxidase (NOX) isoforms that play modulatory roles in insulin secretion. In this article, we provided evidence for islet-cell-specific expression of NOX5 and its upregulation in β-cells under high-glucose condition in vitro. We created a suitable mouse model to study the role of NOX5 in vivo and demonstrated a detrimental effect of NOX5 on insulin secretion under diabetogenic conditions. Our data imply NOX5 as a potential therapeutic target to improve β-cell function and insulin secretion in obesity-related diabetes.
The molecular structure and the characteristics of its enzymatic activity render NOX5 a particularly intriguing molecule in islet function. Indeed, NOX5 contains four Ca2+-binding elements and several phosphorylation sites that control NOX5 activity in a concerted manner (20, 46). Calcium binding is essential for NOX5 superoxide production, while phosphorylation of specific residues increases the Ca2+ sensitivity of the EF hands allowing NOX5 to respond to regulatory cues at physiological (10–100 nM) Ca2+ concentrations (4, 6, 20, 46, 48). Calcium signals, NADPH, and ROS form a complex network to modulate islet glucose sensing and insulin secretion (40). Increase in glucose concentrations leads to cytosolic Ca2+ accumulation that triggers a biphasic wave of β-cell insulin release. NADPH acts as one of the “amplifying factors” sustaining the second phase of insulin secretion (51). In line with these studies, insulin secretagogues increase the NADPH/NADP ratio in rodent islets, and inhibition of NADPH formation reduces insulin secretion (3, 27, 39). Last but not least, a transient elevation in cellular ROS was identified as an essential second messenger promoting insulin secretion (45, 50). NOX5 produces ROS through NADPH oxidation in a Ca2+-dependent manner, but its potential physiological role or pathological effect in islet insulin secretion has not yet been investigated.
The lack of substantial data concerning NOX5 in islet function urged us to (i) characterize the expression of NOX5 in situ in human pancreatic sections and in vitro in isolated islets, (ii) assess its role in insulin secretion in human islets by employing small interfering ribonucleic acid (siRNA)-mediated knockdown of NOX5, and (iii) assess the potential of NOX5 role in vivo in diabetes-related impairment of β-cell function using a novel mouse model. To achieve the latter goal, we generated mice with doxycycline-inducible, β-cell-specific NOX5 expression and evaluated their islet insulin-secretory capacity in chow-diet (CD)-fed condition and after 14 weeks of a high-fat diet (HFD) regime.
Results
Glucose-induced upregulation of NOX5 expression in human islet β-cells
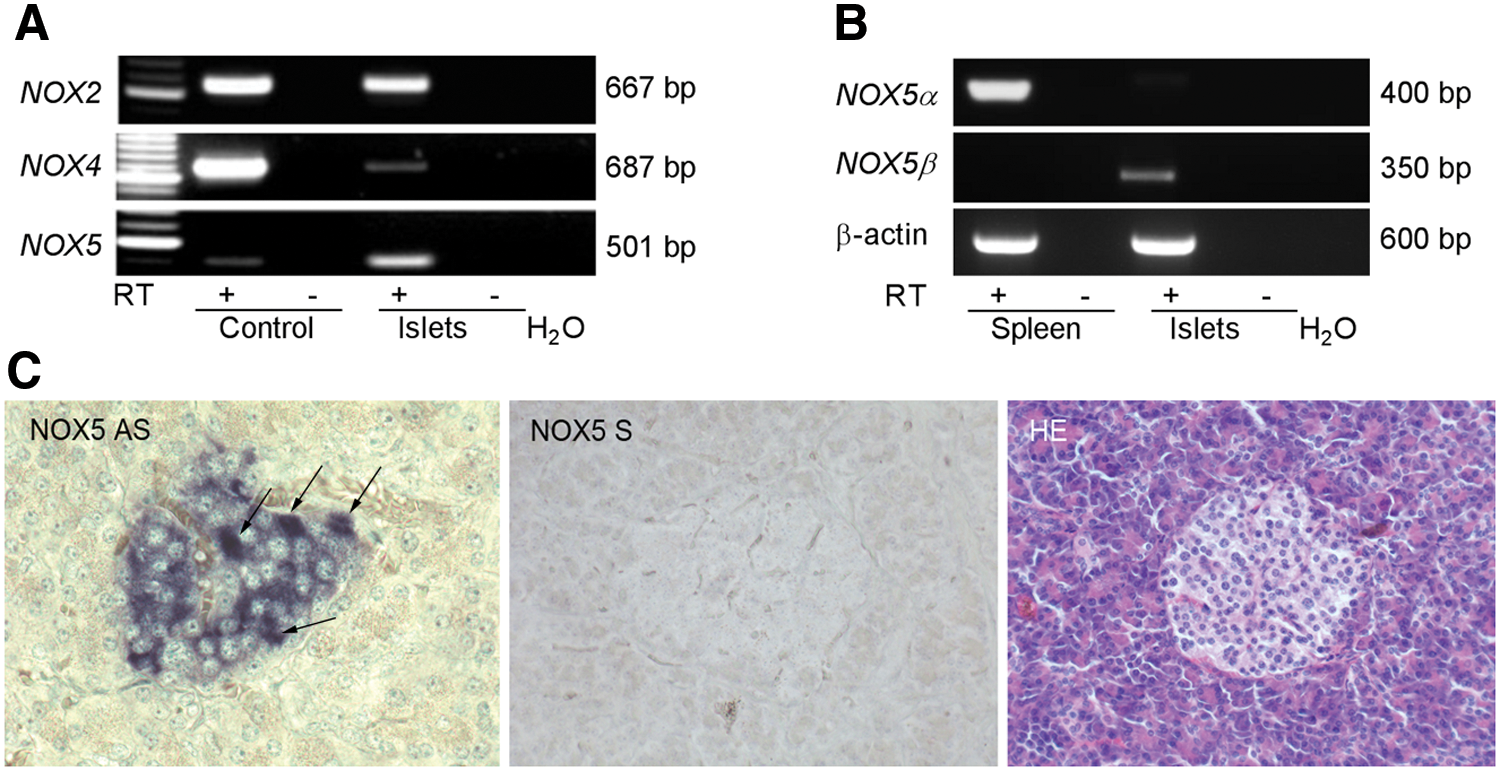
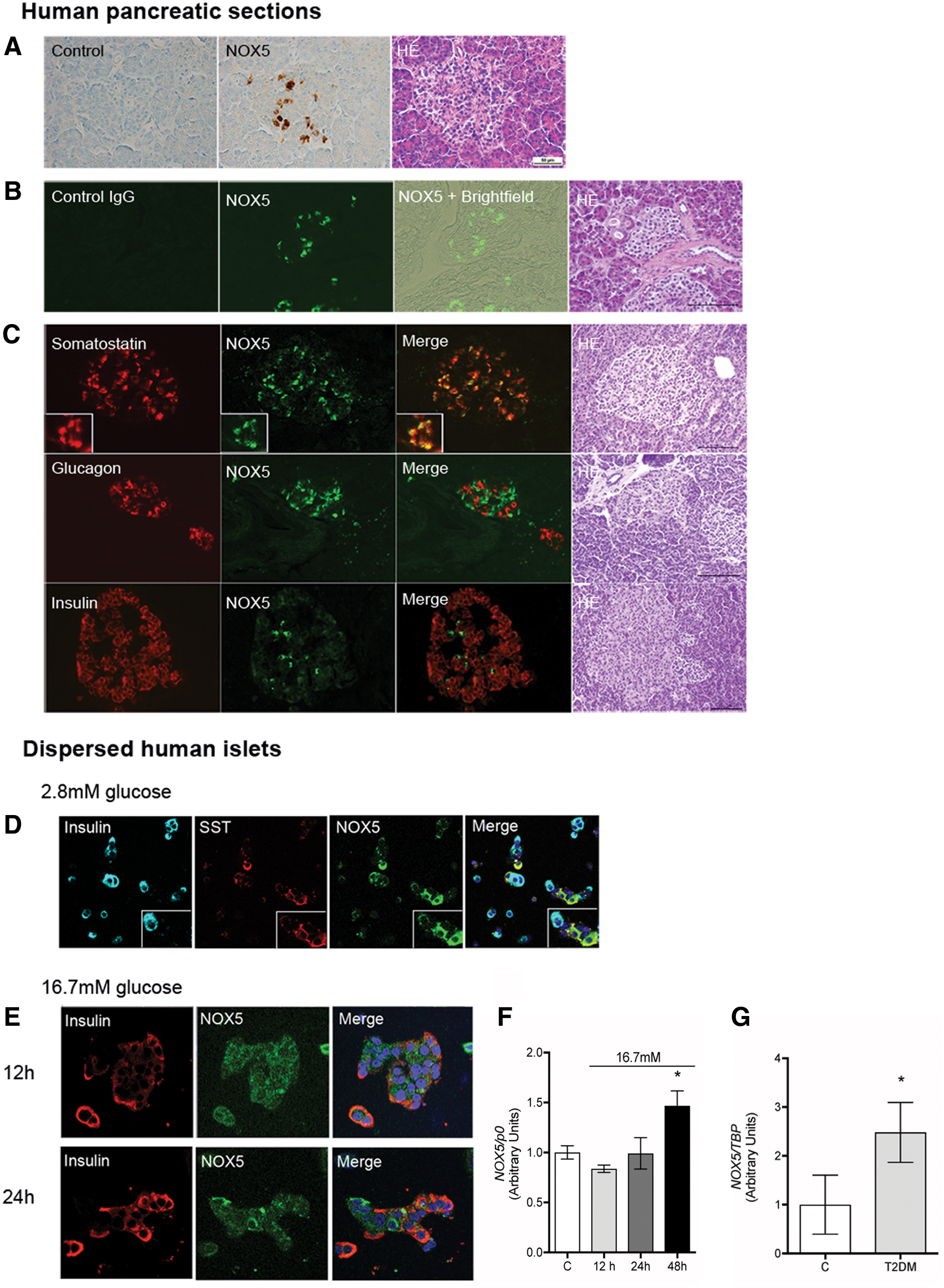
Conventional RT-PCR validated the presence of NOX2, NOX4, and NOX5 mRNAs in isolated whole human islets (Fig. 1A and Supplementary Fig. S1). NOX5 mRNA can undergo differential splicing giving rise to different NOX5 isoforms. The two major forms are termed NOX5α and β, and have been cloned from the spleen and testis, respectively (4, 20). Primers selectively recognizing either the α or β isoform demonstrated the presence of Nox5β mRNA in islets (Fig. 1B and Supplementary Fig. S1). The functionality of RT-PCRs for NOX5α was confirmed by the amplicon obtained in the reaction using human spleen complementary DNA (cDNA). The correct identity of the band obtained in islets with the NOX5β isoform-specific primers was validated by gel purification of the band and subsequential sequencing. In situ hybridization using pancreatic sections confirmed the presence of NOX5 mRNA in the majority of islet cells with occasional cells displaying more prominent signal intensity. Exocrine pancreas was devoid of NOX5 labeling. No signal was obtained when using the negative control NOX5 sense probe (Fig. 1C). NOX5 protein expression was then evaluated by immunohistochemistry and immunofluorescence using human paraffin-embedded pancreatic sections. For immunohistochemistry, a previously tested monoclonal NOX5-specific antibody was employed (1, 11) revealing prominent immunodetectable NOX5 protein in numerous islet cells (Fig. 2A). Identification of islet cells with the highest NOX5 protein expression was then achieved by immunofluorescence employing an unrelated NOX5 antibody (HPA019362; Atlas Antibodies). The specificity of the antibody was established by Western blot and immunofluorescence, comparing wild-type (WT) and NOX5-overexpressing HEK293 (human embryonic kidney) cells. This antibody detected a prominent band of ∼72 kDa in the NOX5-transfected but not in WT HEK293 cells. By confocal immunofluorescence, NOX5-transfected cells displayed dispersed cytoplasmic labeling while no signal was observed in WT HEK293 cells (Supplementary Fig. S2A). The specificity of the NOX5 antibody in immunofluorescence application using paraffin-embedded tissues was ascertained on human pancreas serial sections that were incubated either with a nonrelevant isospecific IgG (control) or with NOX5 antibody (NOX5). Images taken in brightfield light were superposed to NOX5-stained sections for islet identification (NOX5+Brightfield). In addition, serial sections were used for hematoxylin–eosin staining (Fig. 2B). Next, NOX5-expressing cells in human islets were identified by immunofluorescence costaining with the aforementioned NOX5 antibody in conjunction with antibodies recognizing the three major islet hormones: somatostatin (SST), glucagon, and insulin. High NOX5 expression was detected in cells that were also positive for SST. Glucagon and insulin-positive cells did not show NOX5 positivity detectable by this antibody (Fig. 2C). Next, we employed immunofluorescence on dispersed whole human islets to investigate the regulation of NOX5 expression. Dispersed islet cells incubated under basal (2.8 mM) glucose concentrations revealed prominent NOX5 labeling in SST-positive (δ) cells and in colocalization with SST-containing vesicles. At this basal glucose concentration, no NOX5 staining was observed in insulin-positive (β) cells (Fig. 2D). Next, immunofluorescence was applied to islets subjected to 16.7 mM glucose media for 12 and 24 h. Upon high-glucose insults, a lower intensity, punctuated NOX5 labeling was visible in insulin-positive cells, though not in colocalization with insulin-containing vesicles (Fig. 2E). Glucagon-positive (α) cells were devoid of NOX5 signal in both basal (2.8 mM) and high-glucose (HG, 16.7 mM) conditions (Supplementary Fig. S2B). NOX5 expression is mainly controlled at the protein level through proteasomal degradation (15). In line with a post-transcriptional regulation, we did not observe any changes in NOX5 mRNA levels assessed by real-time PCR in whole islets incubated with 16.7 mM glucose for 12 and 24 h. However, glucose-induced upregulation of NOX5 expression was observed in islets subjected to high-glucose media for 48 h (Fig. 2F). Transcription of other NOX isoforms detected in islets (Nox2 and Nox4) remained unchanged in the same conditions, while Duox2 mRNA levels showed a tendency toward a nonsignificant decrease after 48 h of high-glucose media incubation (Supplementary Fig. S2C). To substantiate the in vivo relevance of NOX5 upregulation, we compared NOX5 expression in samples of islets of a previously described cohort of control and T2DM subjects (24). Islets of T2DM subjects displayed significantly increased NOX5 mRNA expression compared with nondiabetic controls (Fig. 2G). Expressions of NOX1, 2, 3, 4 and DUOX2 were statistically not different between control and T2DM subjects (Table 1).
Nicotinamide Adenine Dinucleotide Phosphate Oxidase Isoform Messenger RNA Expression Levels in Islets of Healthy Subjects and Subjects with Type 2 Diabetes Mellitus
D/H, expression ratio between subjects with Type 2 diabetes mellitus and healthy subjects; DM, diabetes mellitus, DUOX, dual oxidase; NOX, nicotinamide adenine dinucleotide phosphate oxidase; SD, standard deviation.
siRNA-mediated knockdown of NOX5 in human islets abrogates glucose-induced insulin secretion
To gain further insight into the relevance of NOX5 in islet insulin secretion, we performed siRNA-mediated knockdown of NOX5 in dispersed human islet cell cultures (Fig. 3). Efficiency of the NOX5 siRNA was verified in cultures of primary human skeletal muscle myotubes, showing a suppression of NOX5 mRNA levels of ∼35% of scrambled RNA-transfected cells (Supplementary Fig. S3). For the human islet transfection experiments, islets were dispersed and cultured in 5.6 mM glucose media overnight and transfected by NOX5-specific or control (scrambled) siRNAs, and further kept in culture for 48 h in low-glucose (LG, 5.6 mM) or HG (16.7 mM) media. At the end of the incubation period, a glucose-induced insulin secretion (GIIS) assay was performed using 16.5 mM glucose as stimulatory agent. Islets transfected with scrambled siRNA displayed an average 1.8-fold increase in insulin secretion in response to glucose stimulation (S) compared with their basal (B) secretion when cultured in LG media (Fig. 3, scrambled, LG). Forty-eight hours preincubation in HG media led to an increase in basal secretory rate with a blunted secretory response to glucose (Fig. 3, scrambled, HG). Islets transfected with NOX5-specific siRNA (siNOX5) displayed a tendency toward lower basal insulin secretion and an abrogated glucose-induced insulin-secretory response compared with scrambled siRNA-transfected islets (Fig. 3, siNOX5, LG). Upon HG preincubation, siNOX5-transfected islets also responded by an increase in basal insulin secretion but failed to reach the levels observed in control-transfected islets. Similar to control islets, glucose stimulation did not further increase their insulin secretion (Fig. 3, siNOX5, HG). Insulin contents were similar between scrambled and siNOX5-transfected islets (data not shown).

Creation of mice with inducible β-cell-specific expression of NOX5
The in vitro human islet experiments suggested that NOX5 is indeed involved in the regulation of islet insulin secretion and the adaptation to HG conditions. However, these experiments could not properly distinguish between the effects of the lack of NOX5 in δ- or β-cells and could not provide information concerning the role of NOX5 in islet function in vivo. The Nox5 gene is present in the human genome, yet rats and mice, the most frequently employed species in islet function research, are natural NOX5 gene knockouts (4). Thus, to study NOX5 function in vivo, diverse mouse models with cell-specific knock-in of Nox5 were created. These models have successfully been used to characterize the role of NOX5 in diabetic kidney disease, hypertension, and stroke (12, 28, 61). To distinguish the role of NOX5 between islet β- or δ-cells in vivo, two novel NOX5-expressing mouse models with RIP (rat insulin promoter) or SST-promoter-driven knock-in of the Nox5 gene needed to be generated. In our first set of investigations that we present in this article, we set out to examine the direct role of NOX5 in β-cell insulin secretion by employing mice with doxycycline-inducible β-cell-specific expression of NOX5 (RIP/rtTA//NOX5β) mice. For this purpose, TetO/NOX5β transgenic FVB mice were crossed with RIPrtTA C57Bl6/CBA mice that express the tetracycline-activated transcription factor rtTA under the direction of the rat insulin 2 promoter (RIP) (41). In the presence of doxycycline (DOX), the rtTA transcriptional activator binds the TetO promoter triggering NOX5β expression in islet β-cells (Fig. 4A). NOX5 and rtTA transgene transmissions were verified by PCR (Fig. 4B and Supplementary Fig. S4A). Then, 5-week-old male mice with similar transgene intensity signals were provided drinking water with (DOX+) or without (DOX−) 1 mg/mL doxycycline for 6 weeks. Induction of NOX5 mRNA expression was verified by RT-PCR in isolated islets. DOX+ mice displayed a strong NOX5 amplicon, while no signal was detected in DOX− mice (Fig. 4C and Supplementary Fig. S4B). Besides islet β-cells, the RIP promoter is known to induce gene expression in selected hypothalamic neurons (32). In line with these data, NOX5 mRNA was also detected in the hypothalamus of DOX+ mice, whereas other organs (spleen and liver) remained devoid of NOX5 (Fig. 4D and Supplementary Fig. S4C). Immunohistochemistry confirmed NOX5 expression in the majority of islet cells in pancreatic sections of DOX+ mice with no signal observed in DOX− mice (Fig. 4E). Functionality of the NOX5 protein was validated in isolated islets by measuring superoxide production using the nitroblue tetrazolium (NBT) reduction test. Islets derived from DOX+ mice showed approximately twofold higher superoxide production than those derived from DOX− mice (Fig. 4F). The ROS-producing capacity of NOX5 is enhanced by increases in calcium concentrations. Thapsigargine is a noncompetitive inhibitor of the sarco/endoplasmic reticulum Ca2+ ATPase pump (SERCA) responsible for Ca2+ transport into the ER. Blocking ER Ca2+ transport leads to ER storage depletion resulting in secondary Ca2+ influx through the plasma membrane triggering a raise in cytosolic Ca2+ concentrations. ER Ca2+ store depletion leads to ER stress and the induction of Ca2+-dependent apoptotic pathways in islets (29, 49). To evaluate whether NOX5 is involved in the onset of Ca2+-mediated ER stress, we compared Thapsigargin-induced expressions of two ER stress marker proteins, 78-kDa glucose-regulated protein (GRP78) and C/EBP homologous protein (CHOP), between islets derived from DOX− and DOX+ mice (Fig. 4G). Expressions of these two proteins were comparable between mice with and without islet NOX5 expression.

Metabolic characterization of RIP/rtTA/NOX5β mice
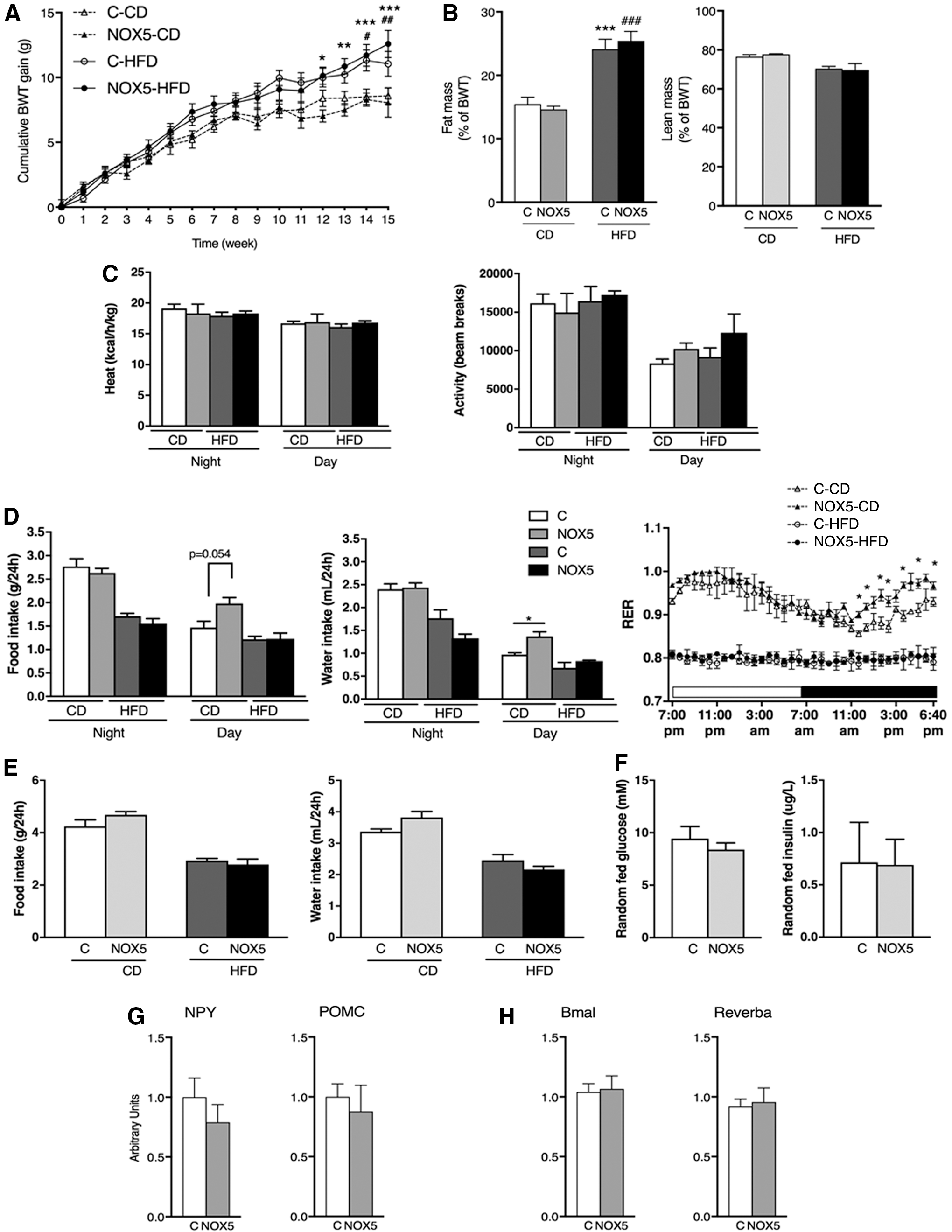
To investigate the potential role of NOX5 in relation to hypernutrition-induced impairment of insulin secretion, we studied four groups of male mice. Mice were first administered drinking water for 2 weeks with or without DOX addition, and were subsequently provided either a control CD or a HFD for 14 weeks while continuing with or without DOX administration. Two weeks of DOX application had no effect on body weight gain (3.70 ± 0.42 g and 3.30 ± 0.33 g in DOX− and DOX+ mice, respectively). Fourteen weeks of HFD feeding resulted in elevated body weight gain and fat deposition compared with the CD-fed groups, but there was no difference between DOX− and DOX+ mice within the same regime group (Fig. 5A, B). Similarly, no difference was observed in fed or starved glucose levels between DOX− and DOX+ mice regardless of whether they were kept under CD or HFD regime (Table 2). A detailed metabolic characterization was performed in calorimetric cages. Heat production and locomotor activity were similar between DOX− and DOX+ mice in both diet regimes (Fig. 5C). Interestingly, however, CD-fed DOX+ mice displayed increased diurnal food (p = 0.054) and water intake (p = 0.031), and an increase in the diurnal respiratory exchange ratio (RER) reflecting more prominent carbohydrate utilization (Fig. 5D). Twenty-four hours cumulative food and water intake showed no differences between CD-fed DOX− and DOX+ mice, pointing toward disturbed daily rhythm of feeding in NOX5-expressing mice (Fig. 5E). HFD feeding resulted in attenuated differences between light and dark period food and water intake, and in diminished RER values indicating prominent fat utilization in DOX− mice. HFD-fed DOX+ mice were similar to DOX− mice in respect to these parameters (Fig. 5D, E). To evaluate the effect of differences in circadian rhythm on metabolic homeostasis, we evaluated random fed glucose and insulin levels in CD-fed control and NOX5-expressing mice between 2:30 and 3:00 pm when their food intake and energy utilization showed the most noticeable differences. Glycemic values and insulin concentrations between control and NOX5-expressing mice showed no differences (Fig. 5F). At the same daylight period, we evaluated the expressions of hypothalamic orexigenic (neuropeptide Y, NPY) and anorexigenic (proopiomelanocortin, POMC) peptides that showed no difference between control (C) and NOX5-expressing (NOX5) mice (Fig. 5G). Similarly, hypothalamic mRNA levels of the circadian rhythm-regulating Bmal and Reverba genes were undistinguishable between these two groups of mice (Fig. 5H).
Glucose Levels in Chow-Diet- and High-Fat Diet-Fed Control and NOX5-Expressing Mice After 13 Weeks of High-Fat Diet Regime
CD, chow diet; HFD, high-fat diet.
Beta-cell-specific NOX5 expression aggravates diet-induced impairment of insulin secretion in vivo and in vitro
To characterize the effect of β-cell NOX5 expression on islet function, we compared GIIS between CD- and HFD-fed control and NOX5-expressing mice both in vivo and in vitro. CD-fed DOX− and DOX+ mice displayed similar serum insulin levels both in starved conditions (Basal, B) and after 5 min of 2 mg/kg intraperitoneal glucose injection (Stimulated, S) (Fig. 6A; left panel). Similarly, islets isolated from CD-fed DOX− and DOX+ mice displayed comparable insulin secretion in basal conditions (2.8 mM, Basal, B) and in response to 16.7 mM glucose challenge (Stimulated, S) in vitro. Maximum secretion capacity assessed as theophylline-induced (Theo) insulin secretion was also unaltered between control and NOX5-expressing mice (Fig. 5A; right panel). Islet insulin content was undistinguishable between CD-fed DOX− and DOX+ mice (data not shown). After 9 weeks of HFD feeding, DOX− but not DOX+ mice displayed elevated starved serum insulin levels (0.24 ± 0.01 and 0.39 ± 0.06 vs. 0.23 ± 0.02 and 0.25 ± 0.01 μg/L in CD and HFD-fed DOX− vs. CD and HFD-fed DOX+ mice, respectively) (Fig. 6B; left panel). Intraperitoneal glucose injection led to a modest increase in insulin levels in HFD-fed control mice but failed to induce any detectable increase in insulin secretion in NOX5-expressing mice. Islet number and size quantified on pancreatic sections revealed no differences between HFD-fed control and NOX5-expressing mice (Supplementary Fig. S5). Abrogated GIIS was also confirmed in vitro using islets isolated from DOX− and DOX+ mice sacrificed after 14 weeks of HFD (Fig. 6C; right panel). Indeed, islets of DOX− mice still retained an ∼30% increase in insulin release upon exposure to 16.7 mM glucose. By contrast, islets of DOX+ mice lacked any glucose-induced insulin-secretory response, and their theophylline-induced insulin release capacity was significantly diminished. Islet insulin content was unaltered between DOX− and DOX+ mice (data not shown). Enhanced lipotoxicity-associated β-cell secretory defect related to NOX5 expression was also implied by additional in vitro experiments. Seven-week-old male mice receiving drinking water with or without DOX supplementation for 6 weeks were sacrificed, the islets isolated and incubated in the presence of 0.5 mM palmitate for 72 h and subsequently subjected to GIIS test. Palmitate-exposed islets displayed abrogated insulin-secretory response to 16.7 mM glucose challenge, with no difference between islets of DOX− and DOX+ mice. However, theophylline-induced insulin secretion was significantly lower in NOX5-expressing compared with control islets (Fig. 6C). Theophylline induces cyclic adenosine monophosphate (cAMP) levels through inhibition of phosphodiesterase (PDE). To investigate the effect of NOX5 on cAMP-mediated insulin secretion from the opposite angle, we compared the effect of the GLP-1 analog exendine-4, an activator of adenylcyclase. Surprisingly, exendine-4 (Ex) raised insulin secretion in DOX− and DOX+ islets to the same extent. Total islet insulin content was similar between DOX− and DOX+ mice (data not shown).

Discussion
ROS are established bidirectional modulators of islet insulin secretion and survival exerting both beneficiary and detrimental effects depending on the amount and the source of ROS (50). This dual role of ROS in islet function has been highlighted by several studies [reviewed in Newsholme et al. (45)]. Indeed, a transient increase in β-cell ROS production after glucose administration has been attributed an essential function in insulin secretion (38, 44). In contrast, sustained ROS elevation has been linked to impaired insulin release (19, 36). NOX enzymes are major sources of cellular ROS. NOX-es are expressed in a tissue- and cell-specific manner, regulated by particular stimuli, and act as crucial mediators of various physiological cellular processes. On the contrary, chronic overactivation of NOX-derived ROS production has been linked to the onset of diverse pathologies (35, 45). Concerning islet function in specific, a physiological inhibitory role of NOX2 in GIIS was established by investigations conducted in NOX2-knockout mice employing both in vitro isolated islets and in vivo experiments (18, 38, 44). By contrast, deregulated activation of NOX2/NOX1 was associated with hampered islet function in diabetic or inflammatory conditions (42, 43). A similar dual modulatory role was also proposed for NOX4. Employing a chemical inhibitor of NOX4, Anvari et al. found that short-term activation of NOX4 was necessary for appropriate GIIS; however, prolonged overstimulation of NOX4 resulted in damaged β-cell function (2). The fact that in physiological conditions NOX enzymes can exert both antagonistic (NOX2) and agonistic (NOX4) effects on insulin secretion highlights the necessity of further exploration of their islet cell type (α, β, δ) distribution, subcellular localization, regulation, and signaling mechanisms. Most organs and cell types express several NOXes, but the function of each NOX isoform seems to be unique and not replaceable by other isoforms (6, 16). In line with a specific regulatory role for each NOX isoform, no report has suggested a compensatory upregulation of other NOX-es in mice rendered NOX deficient by classical gene recombination, siRNA-mediated gene silencing, or chemical inhibition (2, 18, 37, 38).
Our study examined the role of one of the least investigated NOX isoforms, NOX5 in islet function. Expression of NOX5 mRNA in human pancreas and isolated islets was previously observed by RT-PCR, real-time PCR, and RNA sequencing (2, 21, 38). Employing pancreatic sections, we validated the presence of Nox5 mRNA and protein in human islets in situ. Of note, that in situ hybridization detected Nox5 mRNA expression in the majority of islet cells implying the presence of Nox5 mRNA in β-cells. These data are in line with RNA sequencing analysis that reported Nox5 mRNA in human islet sorted islet β-cells (9). Compared with the more ubiquitous pattern of Nox5 mRNA expression immunodetectable NOX5 protein was observed in a more restricted number of cells scattered within the islet, identified by coimmunofluorescence staining as δ-cells. This divergence between mRNA and protein detection pattern might either be related to different sensitivity between the mRNA and protein detection methods, or alternatively, it might reflect a differential rate of hsp70/ubiquitin-mediated NOX5 protein degradation between δ- and β-cells. Expression of NOX5 is regulated at the protein level through hsp70-induced proteasomal degradation allowing upregulation of NOX5 protein levels without concomitant alterations in mRNA transcription in diverse cell types (13 –15). In line with this well-characterized regulatory mechanism of NOX5 expression, we showed divergent NOX5 mRNA and protein levels in human islet cell types. Indeed, our in situ hybridization experiments demonstrated the presence of NOX5 mRNA in the majority of islet cells, implying the presence of NOX5 mRNA in β-cells (insulin-positive cells) that make up >80% of islet cells. On the contrary, NOX5 protein expression was below the detection level of the two antibodies employed on both human pancreatic sections and dispersed human islet β-cells but was raised above detection level once β-cells were induced by HG concentrations. Of additional interest, the low ability of β-cells to increase HSP70 expression under HG/inflammatory challenges together with an increase of NOX activity has previously been proposed as a key factor in β-cell failure (34). Diminished proteasomal degradation of NOX5 could provide an explanation for the detection of NOX5 protein in β-cells in islets incubated in high-glucose media and could also account for the delay in the elevation of NOX5 mRNA levels (48 h) compared with protein levels (12 h) observed in these islets. A similar delayed elevation of Nox5 mRNA compared with the protein amount was also observed in angiotensin II-treated human podocytes (28). NOX enzymes and NOX5 in specific display cell-specific expression patterns within different organs with cell-specific functions and related pathologies (61). In this context, a cell-specific (β- and δ-cells) expression and regulation for NOX5 in different islet cell identities is in line with the current knowledge concerning NOX5. Our current study established this dual expression pattern and then proceeded to evaluate the lack of proper NOX5 function in human islets in vitro using an siRNA approach. These experiments indicated that in human islets cultured under LG conditions, NOX5-mediated signals are essential for proper GIIS. Decreased expression of islet NOX5 also attenuated upregulation of basal insulin release in response to chronic glucose exposure. Taken together, these results suggest dual roles for NOX5: a key mediator of insulin release in response to physiological cues and a mediator in the islet secretion compensatory mechanism when confronting pathologically elevated glucose levels. Unbridled insulin release is a hallmark of islets of T2DM patients. Thus, the lack of chronic HG-induced increase of basal insulin secretion together with the previously observed appearance of NOX5 protein in insulin-positive cells might suggest a negative role for NOX5 in β-cell insulin secretion in hyperglycemic conditions. The siRNA-mediated knockdown approach provides a suitable means for understanding the role of NOX5 in whole human islet insulin secretion, however, lacks the capacity to differentiate between the direct contribution of NOX5 in β-cells and the indirect regulatory effects originating from δ-cells. In addition, while these experiments highlighted the role of NOX5 in islet function in vitro, they did not provide appropriate information about the role of NOX5 in islet function in vivo. Therefore, in our next set of investigations presented in this article we focused our attention on the in vivo role of NOX5 in β-cells, the islet cell type directly accountable for insulin secretion.
To achieve this goal, we generated mice with β-cell specific expression of NOX5. A similar transgenic approach was successfully employed to uncover the role of NOX5 in vivo the development of diverse pathologies, notably diabetic nephropathy, high blood pressure, and stroke (12, 28, 33). Using our novel mouse model, we demonstrated a deteriorating effect for excess NOX5 on insulin release under hyperglycemic/diabetogenic conditions and linked this effect to enhanced degradation of cAMP, a critical amplifying component in insulin secretion. A previous study demonstrated NOX2 immunoreactivity in association with insulin-secretory vesicles together with an antagonistic effect for NOX2 on insulin release involving the cAMP-mediated pathway (38). In our investigations, we observed several critical differences between the islet effects of NOX5 and NOX2. First, as described before, NOX5 protein became detectable in β-cells only when islets were cultured in high-glucose conditions. Second, in line with the appearance of immunodetectable NOX5 in HG-incubated β-cells, the deteriorating effect of NOX5 on insulin secretion was observed only when islets were subjected to prior diabetogenic insults; for example, islets of mice with diet-induced diabetes were used, or when isolated islets were preincubated with palmitate in vitro. Third, NOX5 immunoreactivity was not observed in direct colocalization with insulin-containing vesicles suggesting a distinctive signaling pathway for NOX5 compared with NOX2. This observation is in line with the high specificity of cellular localization and action for each NOX isoform (35). And last but not least, the negative effect of NOX5 was most prominently observed during theophylline-induced insulin secretion, while no differences were noted upon the addition of exendine-4, a GLP-1 receptor agonist. Both theophylline and exendine-4 increase cAMP levels but through different pathways: theophylline is an inhibitor of PDEs (the enzymes degrading cAMP), while exendine-4 is an activator of adenylcyclase (the enzyme generating cAMP). cAMP is a critical amplifying signal of β-cell insulin secretion. NOX2-derived ROS was shown to act as an inhibitor of physiological GIIS by inhibiting adenylcyclase activation (38). In contrast, our results suggest that the detrimental effect of NOX5 on insulin release is relayed through activation of PDEs (52). In islets, different PDE isoforms are present, and their precise isoform-specific implication in GIIS is subject of intensive investigations (52). The identification of PDE isoform linked to NOX5 action by PDE isoform-specific inhibitors was not attempted in this study. Theophylline is also considered as an agonist of adenosine receptors that mediate their effects through modulation of intracellular cAMP levels (58). Inhibition of the A1 adenosine receptor action increases cellular cAMP levels and insulin secretion (66). The potential involvement of Gi-coupled adenosine receptor activation in the observed effect of theophylline in our experiments cannot be excluded and should be addressed in the future. It is noteworthy, however, that in our mouse model NOX5-linked modification of theophylline action occurs only under diabetogenic conditions that also exert an enhancing effect on β-cell NOX5 expression in human islets in vitro. The physiological relevance of glucose-induced NOX5 expression in β-cells in vitro was underlined by our data demonstrating elevated NOX5 levels in T2DM islets. The latter results are in line with previously reported expression profiling data comparing control and T2DM islets (Geoprofiles GDS3882/220641_at). Islet insulin secretion is modulated through crosstalk with hypothalamic neurons (54). The RIP promoter employed in our mouse model is also known to induce hypothalamic gene expression (32). The results of in vivo insulin secretion assays performed in the RIP-NOX5 mice were in line with the in vitro experiments performed on isolated islets. These results suggest (but does not completely exclude) that the effects of hypothalamic RIP expression did not directly affect islet function in our mouse model. Hypothalamic nuclei are major regulators of food intake, and RIP-expressing neurons participate in this process in mice. The precise function of these neurons in the regulation of nutrient intake is currently unknown with studies reporting both orexigenic and anorexigenic functions in different mouse models (55, 63). Mice with RIP-mediated NOX5 expression demonstrated a specific increase in light period food intake without any alterations in the total amount of daily caloric consumption, body weight gain, or body adiposity indicating a possible role of neuronal NOX5 for the regulation of diurnal rhythm of feeding behavior without any obvious alterations in whole-body metabolic homeostasis. Increased day-time chow pellet consumption is reflected in preferential carbohydrate utilization (higher RER values) in RIP-NOX5 mice. By contrast, control mice use more lipids as they are at rest (thus, eat less carbohydrate containing chow pellets) during the day-time period. In our investigations, we did not find any differences at present, the existence and possible role of hypothalamic insulin-producing neurons in humans are largely ignored (22). Similarly, no study conducted a systematic immunohistological scanning of human brain specimen to uncover NOX5-expressing neurons. Thus, at this point we did not aim to pursue any further studies concerning alterations in hypothalamus-related feeding behavior of NOX5-expressing mice but definitely consider these findings a worthy topic for follow-up experiments.
Glucose-induced upregulation of NOX5 in dispersed human islets was observed in β-cells; however, NOX5 was also prominently expressed in SST-producing δ-cells both in basal glucose conditions and during hyperglycemic insult in vitro. SST is crucial in intraislet communication exerting a negative effect on α- and β-cell secretion (26). The clinical relevance of SST-mediated signaling in the modulation of islet hormone release was recently highlighted by an elegant study, demonstrating an important role of SST in the suppressive action of insulin on glucagon release (62). Our results obtained from human islets transfected with siNOX5 suggest a positive regulatory role for NOX5 in human islet insulin secretion in vitro. This effect was observed also when islets were cultured in LG containing media when the most prominent NOX5 expression was observed in SST-positive cells. These results suggest that δ-cell NOX5 might be a modulator SST-mediated regulation of β-cell insulin secretion. Additional in vitro and in vivo investigations will be needed to determine the precise role of NOX5 in δ-cell-mediated α- and β-cell intraislet crosstalk. The hypothetical role of NOX5 in SST-mediated intraislet crosstalk is depicted in Figure 7 (upper panel).
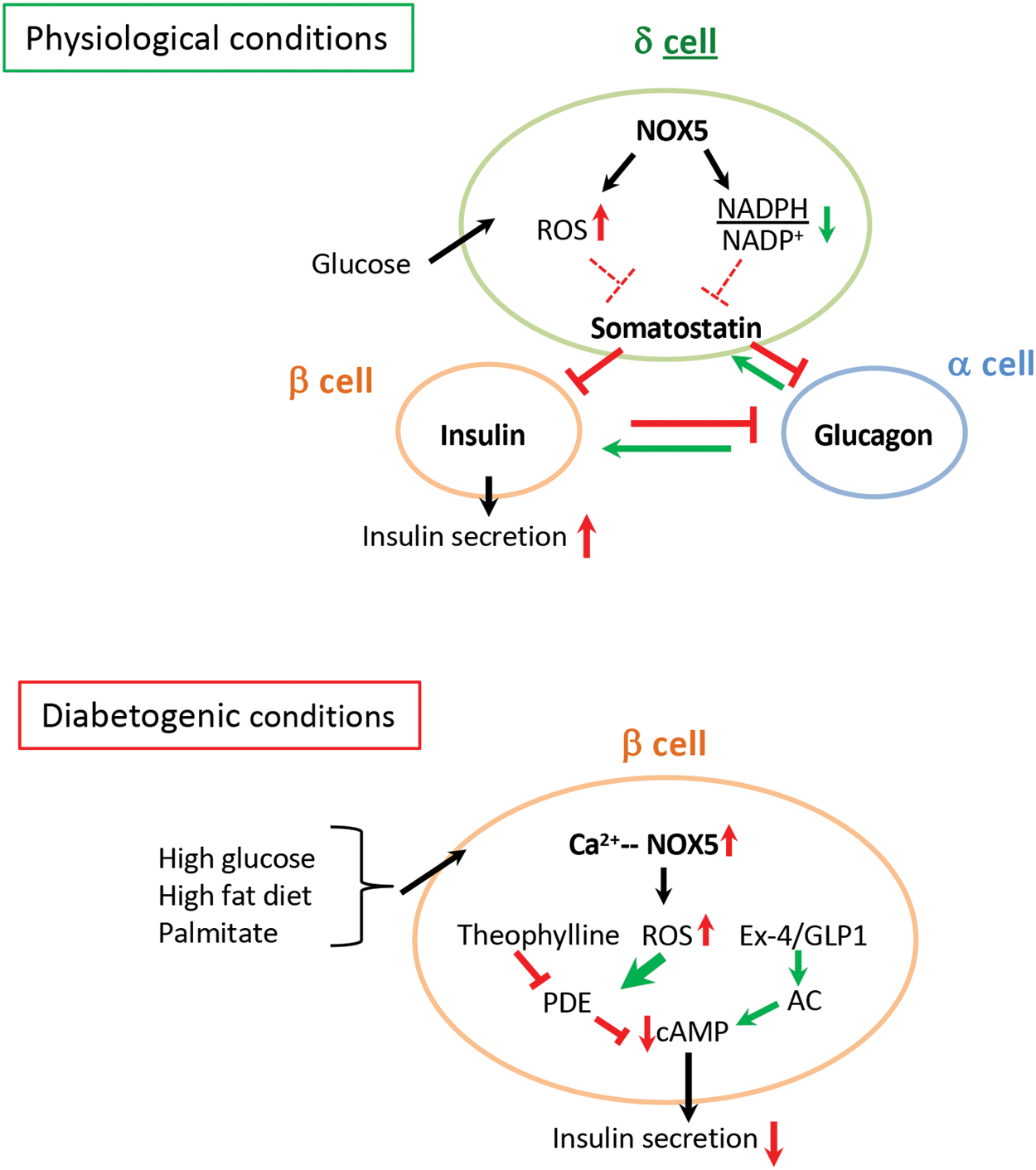
In this study, we aimed to elucidate the role of NOX5 in β-cells, in particular in diabetogenic conditions. The adverse effect of HG-induced ROS production is associated with increased β-cell calcium levels and the attenuation of acute glucose-induced Ca2+ response (8). The ROS-producing activity of NOX5 is calcium dependent responding with a dynamic increase in the same range of Ca2+ concentrations (100–150 nM) that has been observed in islets exposed to HG (4, 5, 8, 46, 47). Alterations in redox status are known modulators of insulin vesicle exocytosis (30). Our results showed diminished GIIS in diabetic circumstances, suggesting sustained NOX5-derived ROS production as a possible link between hyperglycemia, increased β-cell Ca2+ levels, and hampered functionality of the insulin releasing pathway. The impairment of this signaling pathway could be attributed to enhanced PDE-mediated cAMP degradation leading to diminished effectivity of cAMP, one of the crucial amplifying mediators of insulin secretion. The hypothesis concerning the role of NOX5-derived ROS production in diabetes-related β-cell dysfunction is depicted in the lower panel of Figure 7.
In conclusion, this study is the first one to demonstrate the cell type distribution and regulation of NOX5 in human islets in vitro, and explore its function in islet insulin secretion using a novel mouse model in vivo. Our data imply NOX5 as a contributor to overnutrition-induced β-cell damage and ultimate β-cell failure in T2DM. General ROS inhibitors that were employed to improve redox-related pathologies lack therapeutic efficacy, highlighting the need to identify specific ROS sources for more precise pharmacological targeting (56). In this respect, NOX enzymes are among the most promising molecules (17, 31). To our knowledge, our study is the first one suggesting NOX5 as a potential therapeutic target in the context of improving islet function in diet-induced diabetes.
Materials and Methods
Human islet culture
Human islets for research were provided thanks to the European Consortium for Islet Transplantation funded by Juvenile Diabetes Research Foundation (1-RSC-2014-100-I-X) or were obtained from Prodo Laboratories. Islets were cultured overnight at 37°C, in CMRL containing 5.6 mmol/L glucose and 10% fetal calf serum. Then, islets were transferred into fresh complete CMRL supplemented with 5.6 or 16.7 mM glucose, incubated for 12, 24, or 48 h and processed for RNA isolation.
Human islet NOX5 siRNA knockdown
Human islets were transfected by scrambled or a pool of four different siNOX5 (SMARTpool: ON-TARGETplus NOX5, Cat. No. 010195; Dharmacon) using the Darmafect 4 reagent (Cat. No. T-2004; Dharmacon) according to the manufacturer's protocol. Islets were incubated in CMRL medium containing 5.6 or 16.7 mM glucose for 48 h, and then processed for GIIS tests. Secreted insulin was measured by commercial enzyme-linked immunosorbent assay (ELISA) kit (Mercodia). Efficiency of siRNA-mediated knockdown was verified using differentiated myotubes of primary human skeletal muscle cultures obtained from three different healthy donors (Authorization H-A-2007-0016; Videnskabsetiske Komité A for Region Hovedstaden). Cells were cultured in HAM'S F10 media supplemented with 20% SVF, 1% Penicillin/Streptomycin, and 1% ABAM until confluence. Differentiation was induced in Dulbecco's modified Eagle's medium LG media supplemented with 2% SVF, 1% Penicillin/Streptomycin, and 1% ABAM during a 7-day culture period. siRNA transfection was performed as described earlier (10).
RNA preparation, RT-PCR, real-time PCR
Total RNA from human and mouse islets and other tissues was prepared with microRNeasy (Qiagen) and TRIZOL (Life Sciences), respectively. cDNA was synthesized by Superscript II Reverse Transcriptase (Invitrogen). Real-time PCR quantification of NOX5 in islets was achieved using TaqMan assay (hs-00225846-m1; Applied Biosystems) preceded by 15 cycles preamplification. Other genes were quantified using Sybergreen. All reactions were carried out in an ABI 7500 instrument (Applied Biosystems). Results were quantified by the ΔCt method using P0 or the TATA-binding protein (TBP) as standard nonvariable genes. Primers are listed in Table 3.
Sequences of Primers Used for Reverse Transcription Polymerase Chain Reaction, Real-Time Polymerase Chain Reaction, and Genotyping
PCR, polymerase chain reaction; Q-PCR, quantitative polymerase chain reaction; RIP, rat insulin promoter; rtTA, reverse tetracycline-controlled transactivator; TBP, TATA-binding protein.
In situ hybridization
In situ hybridization was performed on paraffin-embedded human pancreatic sections using digoxigenin-labeled complementary RNA (cRNA) as described previously (4).
Immunofluorescence
For immunofluorescence, a commercial NOX5 antibody (HPA-0119362; Atlas Antibodies) was employed. The antibody was validated by Western blot and immunofluorescence using HEK293 WT (negative control) and NOX5-transfected cells (positive control) provided by Prof. K.-H. Krause (University of Geneva) (Supplementary Fig. S2A) (4). For immunofluorescence, islets were rinsed in phosphate buffered saline and incubated in Accutase (Sigma). Single-cell suspensions were rinsed with complete CMRL, and aliquots of 100,000 cells were incubated for 24 h at 37°C and then cultured with or without the addition of 16.7 mM glucose in KREBS +0.1% bovine serum albumin (BSA) medium. Cells were subsequently fixed and stained with the appropriate antibodies (Table 4). Immunofluorescence was performed using confocal microscopy (Zeiss LSM510 Meta microscope) (60). Images were acquired and processed using LSM510 software (Carl Zeiss AG) and ImageJ (NIH). Immunofluorescence investigations were conducted on paraffin-embedded human pancreatic sections and on frozen sections obtained from mice with or without preceding doxycycline administration. Images were obtained by confocal microscopy (AxioPlan2; Zeiss) and processed by Axiovision software (Zeiss). Antibodies are listed in Table 4.
Antibodies Applied in Immunofluorescence Investigations
Immunohistochemistry
Immunohistochemistry was performed by applying a NOX5 monoclonal antibody (1, 11) to paraffin-embedded human pancreatic sections, or sections derived from mice with or without preceding doxycycline administration. Negative control was obtained by omitting the first antibody. NOX5 signal was revealed by 3,3′-diaminobenzidine (DAB).
Creation of rtTA/RIP/NOX5β mice and animal experiments
NOX5β cDNA was PCR amplified from the pDONRNox5β plasmid (GeneCopoeia) using high fidelity Phusion Polymerase (New England Biolabs) and modified primers with 5′ NotI and 3′ SalI recognition sequences. Purified PCR product and pBI backbone were digested by NotI/SalI and ligated together. A 3.7 kb TetO/NOX5β fragment was excised using EcoRV/XmnI digestion, the resulting band gel purified and provided to the University of Ottawa Core Transgenic Facility for pronuclear injection into fertilized FVB/N oocytes. Subsequent NOX5β founders on a pure FVB/N background were identified by PCR genotyping. Mice with β-cell-specific NOX5 expression were generated by intercrossing TetO/NOX5β mice with rtTA-RIP mice on a C57Bl6/CBA congenic background (41). NOX5 expression was induced by 1 mg/mL doxycycline (DOX) administration in the drinking water for 5 weeks. In a second set of experiments, mice were supplied or not with 1 mg/mL of DOX in their drinking water for 2 weeks. Then, mice were randomly assigned to receive either control chow (RM3; SDS) or high-fat-containing diet (HFD; 60% calories from fat; Ref. 2127; Kliba-Nafag) for 14 weeks while continuing with or without DOX administration. Ethical approval was granted by the Geneva Cantonal Animal Experimentation Committee.
Islet superoxide generation
Superoxide production was detected in islets isolated from mice with (DOX+) or without (DOX−) 6 weeks of DOX administration using the NBT assay; NBT is water soluble and yellow in color before reduction by superoxide that leads to the generation of the water-insoluble blue-colored formazan (57). Islets were incubated in 2.8 mmol/L glucose-containing KRBH/BSA buffer in the presence of 0.2% NBT for 90 min at 37°C. Then, islets were centrifuged at 13,000 rpm for 5 min, the cells were broken in 2 mol/L KOH, and the reduced NBT was dissolved in dimethyl sulfoxide (DMSO). Superoxide quantity was estimated by measuring absorbance at 620 nm. Data are expressed as optical density per 15 islets.
Islet gene expression measurements
Islets were isolated from male and female mice with (DOX+) or without (DOX−) treatment for a period of 6 weeks by pancreas collagenase digestion and Ficoll purification (65). Islets were incubated in 5.6 glucose media overnight. 50–60 islets were then stimulated or not with 25 μM Thapsigargin for 24 h. At the end of the incubation period, islets were collected by centrifugation and processed for RNA extraction.
Islet size determination
Pancreata were fixed in paraformaldehyde, embedded in paraffin, and serial sections were stained by hematoxylin/eosin. Morphometric measurements were conducted on duplicate sections derived from three different tissue levels. Images were obtained by Axiocam Imaging Sytem (Carl Zeiss AG). Islet diameter was calculated from computer-assisted image transformation (MetaMorph Image Analysis).
Metabolic phenotyping and body composition
Energy expenditure and RER were determined by indirect calorimetry, locomotor activity was recorded by infrared frame, food and water intake were measured by specific sensors in Labmaster metabolic cages (TSE). Body composition (fat and lean mass) was analyzed by magnetic resonance imaging (EchoMRI).
Static incubation GIIS
Islets were isolated from CD- or HFD-fed mice employing pancreas collagenase digestion and Ficoll purification (65). Islets were incubated overnight in RPMI 1640 medium with 10% fetal calf serum and with or without additional 2 μg/mL doxycycline. Then, aliquots of 30–150 islets were preincubated for 2 h at 37°C in Krebs-Ringer bicarbonate HEPES (KRHB) buffer supplemented with 0.1% BSA and 2.8 mM glucose. Basal secretion was measured for 1 h in fresh KRHB containing 2.8 mM glucose. Stimulated secretion was measured in KRHB containing 16.7 mM glucose with or without additional 5 mM theophylline or 0.1 mM exendin-4 for an additional hour. At each secretion stage, the incubation buffers were collected. At the end of the test, islets were lysed in acid-ethanol. Insulin secretion and content (acid-ethanol samples) were analyzed by ELISA (Mercodia). Insulin secretion was expressed as the percentage of total insulin content (the sum of insulin secreted during basal, glucose-/exendin-4, and theophylline-stimulated secretion added to acid-ethanol insulin content).
Statistical analysis
One- or two-way analysis of variance tests followed by Sidak's multiple comparison or Student's paired t-tests were used where applicable, p < 0.05 was considered significant.
Footnotes
Acknowledgments
We gratefully recognize the excellent technical help of M. Surini-Demiri (University Hospitals of Geneva) with immunohistochemical analyses and C. Vesin (University of Geneva) with islet isolation experiments. We express our gratitude to Prof. J-H. Doroshow (Center for Cancer Research, National Cancer Institute, Bethesda, MD) for providing us with the monoclonal NOX5 antibody. We are indebted to Prof. K. Steiger (Justus-Liebig University Giessen) for the in situ hybridization experiments. We thank C. Sinigaglia and D. Mattey-Doret (University of Geneva) for their technical help with islet insulin secretion experiments, E. Kovari (University Hospitals of Geneva) for imaging experiments, and Prof. P. Halban and Y. Gosmain (University of Geneva) for critical discussions..
Author Disclosure Statement
No competing financial interests exist
Funding Information
This study was supported by the Ernst and Lucie Schmidheiny Foundation (Geneva, Switzerland) and the Société Francophone du Diabète (Paris, France) to I.S. and by the Juvenile Diabetes Research Foundation (1-RSC-2014-100-I-X, European Consortium for Islet Transplantation [ECIT]).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.