
Abstract
Dr. Hideo Kimura is recognized as a redox pioneer because he has published an article in the field of antioxidant and redox biology that has been cited >1000 times, and 29 articles that have been cited >100 times. Since the first description of hydrogen sulfide (H2S) as a toxic gas 300 years ago, most studies have been devoted to its toxicity. In 1996, Dr. Kimura demonstrated a physiological role of H2S as a mediator of cognitive function and cystathionine β-synthase as an H2S-producing enzyme. In the following year, he showed H2S as a vascular smooth muscle relaxant in synergy with nitric oxide and its production by cystathionine γ-lyase in vasculature. Subsequently he reported the cytoprotective effect of H2S on neurons against oxidative stress. Since then, studies on H2S have unveiled numerous physiological roles such as the regulation of inflammation, cell growth, oxygen sensing, and senescence. He also discovered polysulfides (H2Sn), which have a higher number of sulfur atoms than H2S and are one of the active forms of H2S, as potent signaling molecules produced by 3-mercaptopyruvate sulfurtransferase. H2Sn regulate ion channels and transcription factors to upregulate antioxidant genes, tumor suppressors, and protein kinases to, in turn, regulate blood pressure. These findings led to the re-evaluation of other persulfurated molecules such as cysteine persulfide and glutathione persulfide. Dr. Kimura is a pioneer of studies on H2S and H2Sn as signaling molecules.
Professor Hideo Kimura

Background Development and Training
Dr. Kimura graduated from the University of Tokyo, Faculty of Pharmaceutical Sciences, in 1980 and received his PhD from the University of Tokyo in 1985. He studied neurotransmitters in the cerebellum using electrophysiological techniques at the National Defense Medical College (33, 64), and the gene structure of cytochrome P-450 at the Cancer Institute (35, 36). He completed his postdoctoral studies at the Salk Institute for Biological Studies where he identified a novel growth factor, Schwannoma-derived growth factor, as well as activin (28, 32, 34, 71). He continued working at the Salk Institute as a senior staff scientist, where he identified presenilin-binding protein (PBP), a novel guanine nucleotide exchange factor that activates Rac (27). PBP was later renamed as modifier of cell adhesion (9, 60), and finally as dedicator of cytokinases 3 (Dock3) (61).
Summary of Dr. Kimura's Top Contributions
Dr. Kimura showed the first time that hydrogen sulfide (H2S) is a novel signaling molecule in multiple body systems. He discovered a physiological role of H2S as a mediator of memory formation (1) (Fig. 1), and subsequently identified another role as a vascular smooth muscle relaxant in synergy with nitric oxide (NO) that was the first demonstration of a crosstalk between H2S and NO (21) (Fig. 2). Prominent neuroscientist Solomon Snyder commented in Science News, “They have very impressive evidence that H2S is a potential neurotransmitter. It's an exciting paper that should stimulate a lot of people's interest” (84).
Dr. Kimura discovered the cytoprotective effect of H2S on neurons from oxidative stress (39) (Fig. 3), and the effect on other tissues and organs such as heart and kidney was followed (14, 85). These findings led to the identification of numerous physiological roles of this molecule, including anti-inflammation, angiogenesis, oxygen sensing, and ATP formation (11, 51, 65, 67, 68, 82, 93, 94).

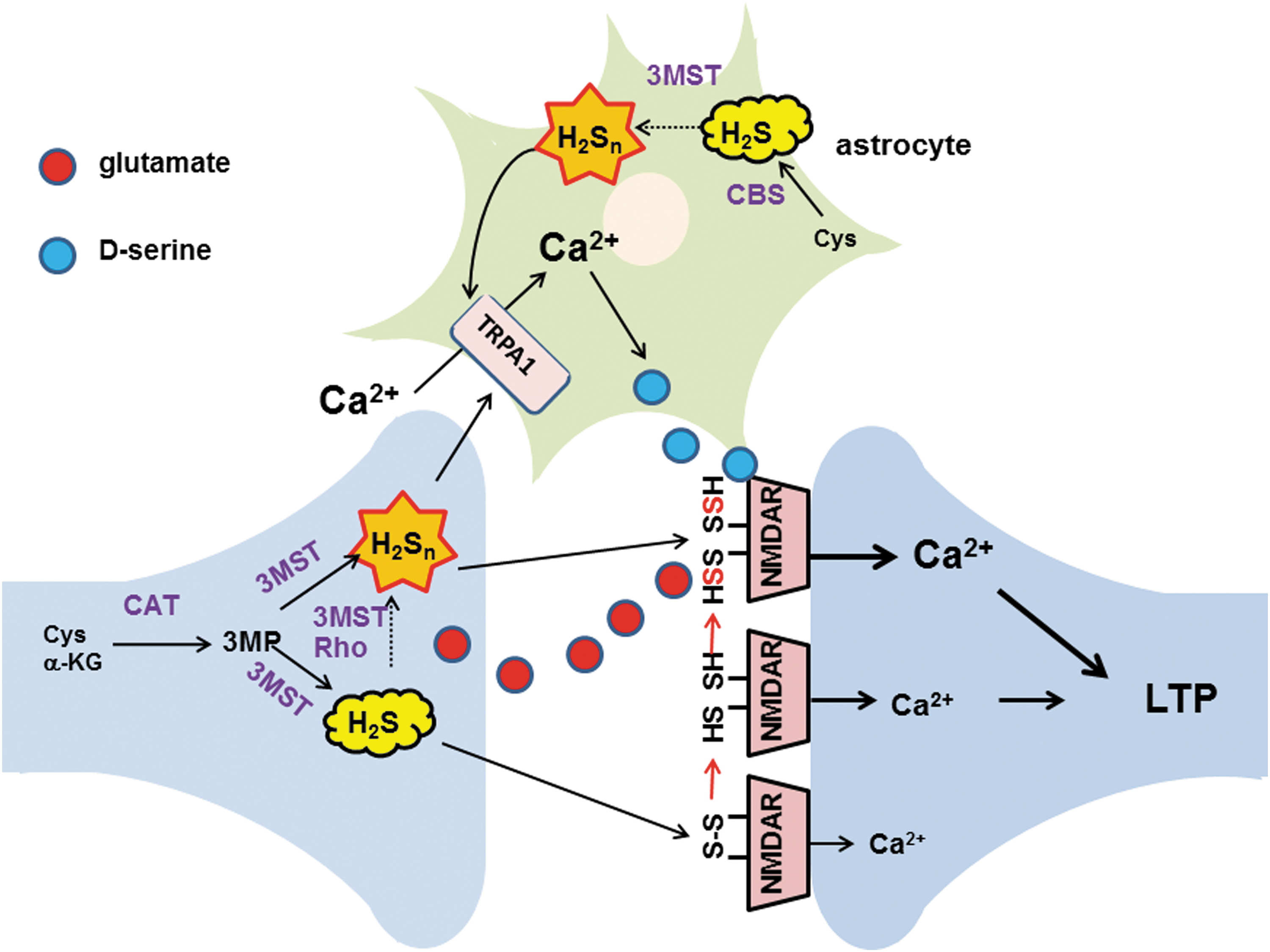
During further investigation of the physiological roles of H2S, Dr. Kimura discovered novel signaling molecules, H2Sn, produced by 3-mercaptopyruvate sulfurtransferase (3MST) (41, 42, 58, 59, 66) (Fig. 4). Since then, studies on H2Sn have unveiled various physiological roles such as the regulation of ion channels, transcription factors, protein kinases, and tumor suppressors, as well as the production of other per- and polysulfurated molecules (18, 41, 44, 80). Dr. Kimura also identified H2Sn produced by the chemical interaction of H2S with NO that may provide one of the mechanisms of the synergy between H2S and NO (52) (Fig. 5).

Description of Key Finding 1
In the library at the Salk Institute, Dr. Kimura found a metabolic map showing mammalian enzymes that can produce H2S. These enzymes were intensively studied from the 1950s to 1970s; cystathionine β-synthase (CBS), cystathionine γ-lyase (CSE), and 3MST together with cysteine amino transferase were found to have the capacity in vitro (5, 8, 48, 79), and their full-length cDNAs were cloned in the 1990s (56, 62, 81). However, rather than being recognized as a physiologically active molecule in these early studies, H2S was thought of simply as a byproduct of metabolic pathways or a marker for the evaluation of enzyme activity.
Three groups discovered endogenous sulfide in mammalian brains when measuring sulfide levels in intoxicated animals (17, 70, 88). Survivors of H2S poisoning experienced memory loss, and acute intoxication with H2S caused changes in the levels of neurotransmitters in the brains of animal models. Inspired by these findings, Dr. Kimura began studying H2S as a signaling molecule in 1993 when carbon monoxide had just been identified as a gaseous signaling molecule (47, 78, 86, 95), following on NO (4, 16, 23).
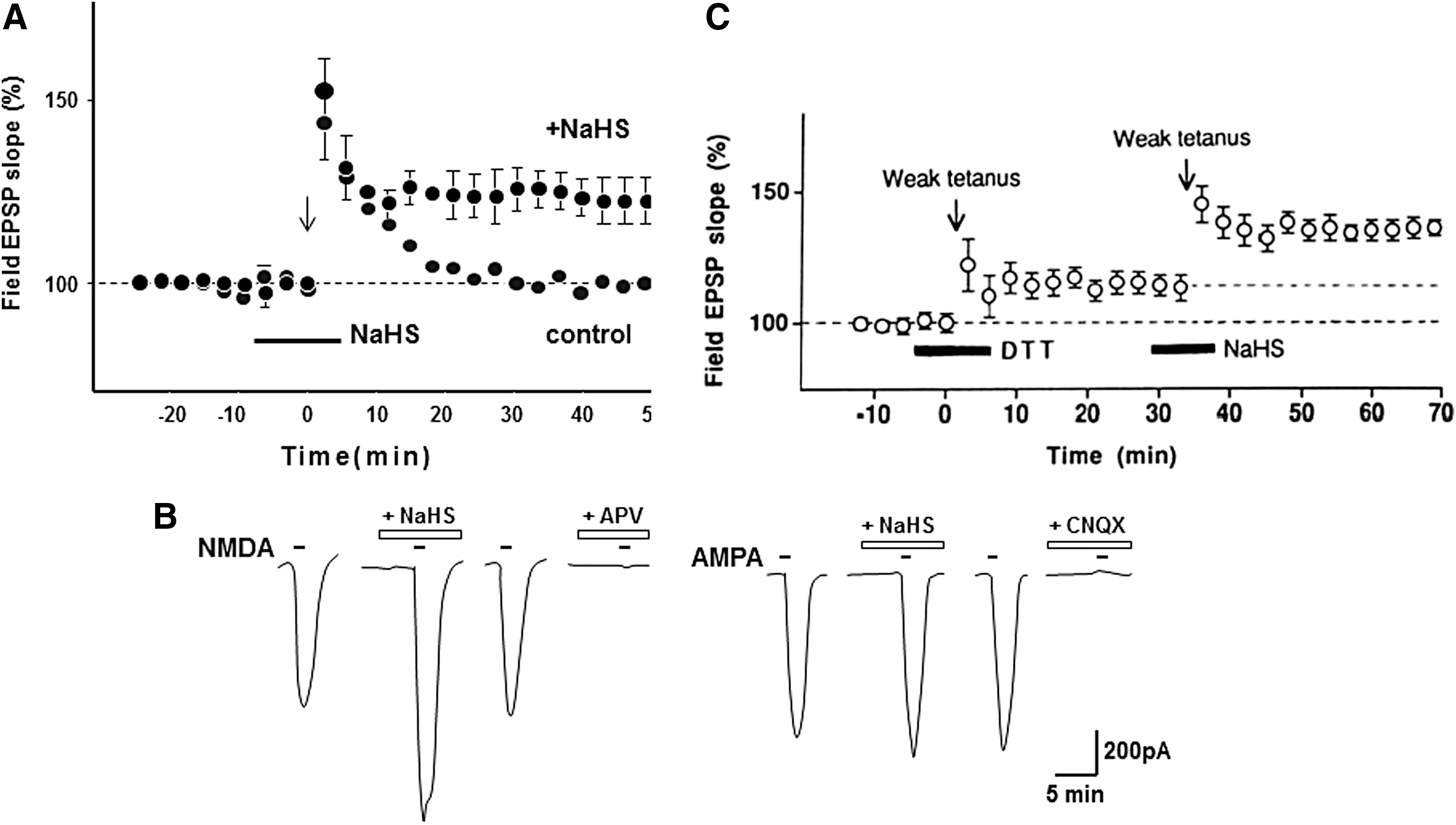
In 1996, Dr. Kimura together with his student Dr. Kazuho Abe demonstrated that H2S, which can be produced by CBS in the brain, facilitates the induction of hippocampal long-term potentiation (LTP), a synaptic model of memory formation (1) (Figs. 1 and 6). H2S enhances the activity of one type of glutamate receptor, N-methyl
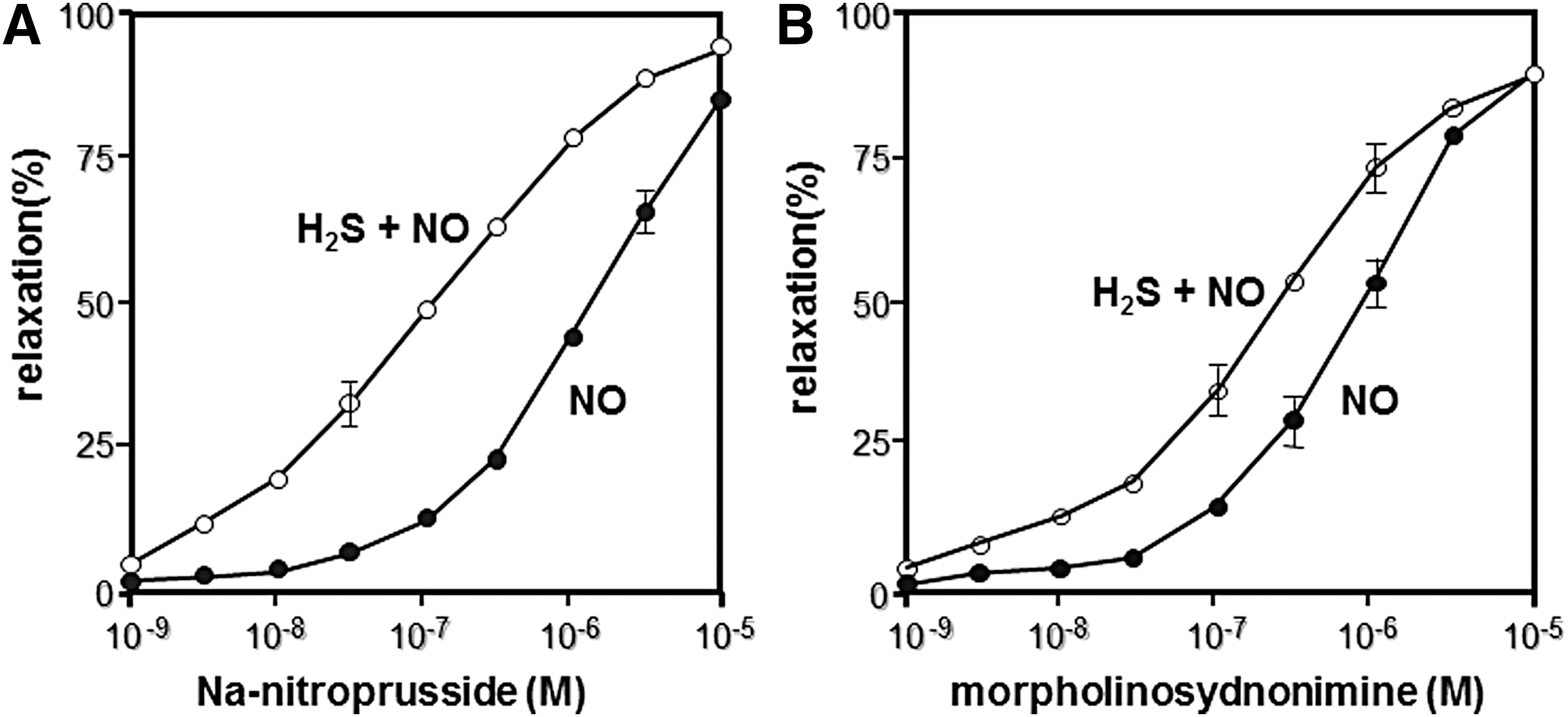
In 1997, Dr. Kimura demonstrated that H2S can be produced by CSE to relax vasculature in synergy with NO (21) (Fig. 2). This discovery also opened a field of crosstalk between H2S and NO (see also Key Finding 3). Subsequently, Wang and colleagues identified KATP channels as one of the targets of H2S to relax vascular smooth muscle (94), and Cirino and colleagues showed cyclic GMP-dependent protein kinase as another target to mediate H2S-induced vasorelaxation (6).
Description of Key Finding 2
Since H2S is a well-known toxic gas, its cytoprotective effects have been overlooked. Dr. Kimura found that H2S protects neurons from oxidative stress through enhancing the activity of the cystine/glutamate antiporter and cysteine transporter, as well as γ-glutamyl cysteine synthetase or glutamate cysteine ligase, a rate limiting enzyme in the production of glutathione (GSH), which is a major cellular antioxidant (38, 39) (Figs. 3 and 7). Dr. Kimura also showed that H2S enhances the activity of KATP and CFTR Cl- channels to suppress the excessive excitation of neurons by stabilizing membrane potential (37) (Fig. 7). This finding led to the identification of the cytoprotective effect of H2S on various tissues and organs, including the heart, kidney, retina, pancreas, and intestines, and the regulation of endoplasmic reticulum stress (14, 26, 45, 49, 72, 85, 93). Several H2S-based therapeutic compounds have been developed, with some undergoing clinical trials (87). H2S even plays an essential role in the development of bacterial resistance to antibiotics (73). H2S is a universal cytoprotectant effective in bacteria and mammals.
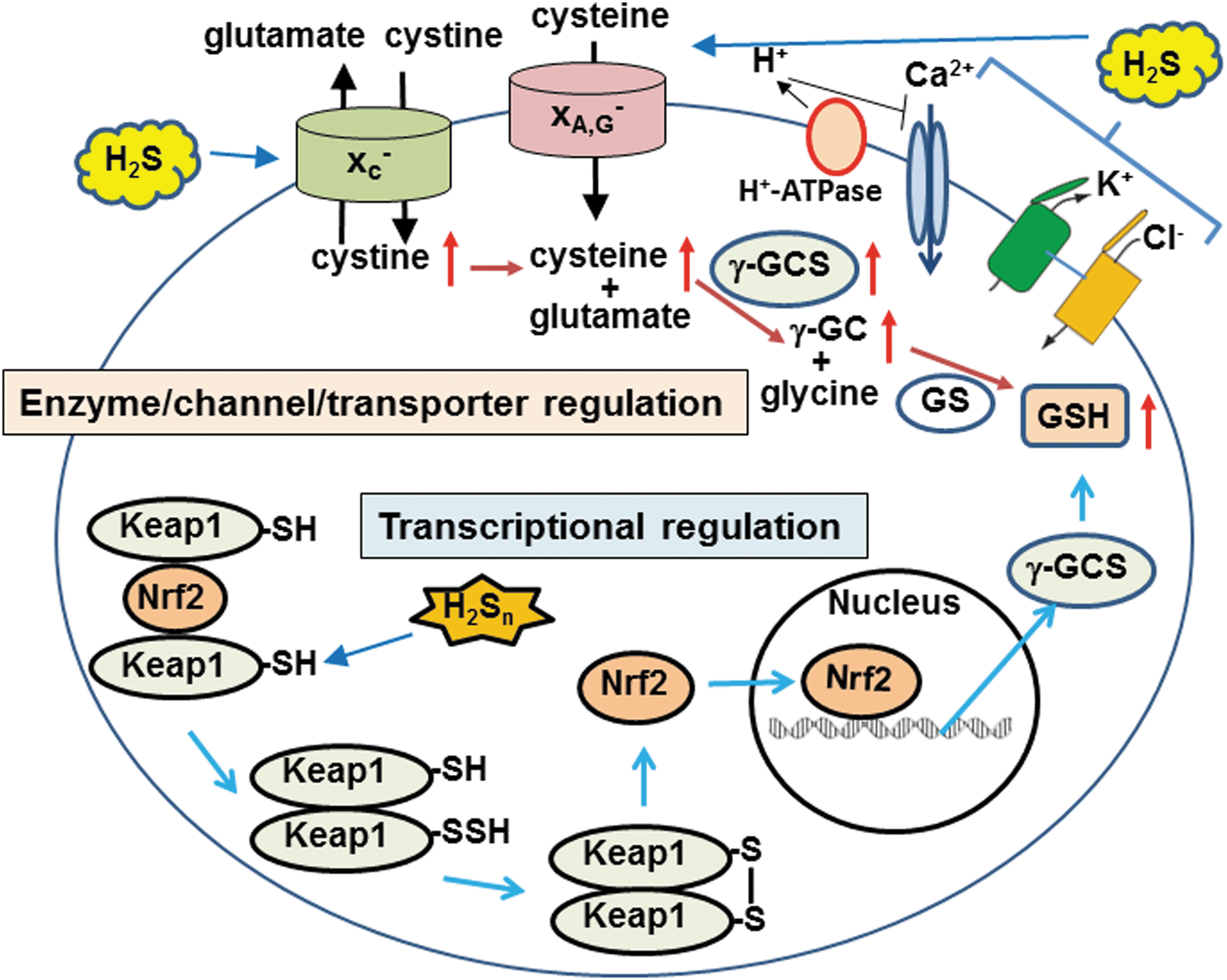
Dr. Kimura's group also identified the transcription factor specific protein 1 (SP1)-binding site at the 5′-noncoding region of the CSE gene (25) that contributes to the cytoprotection via NF-κB signaling. Snyder and colleagues identified a mechanism for the antiapoptotic actions induced by tumor necrosis factor alpha (TNFα), where TNFα stimulates the binding of SP1 to the CSE promoter that increases the levels of CSE, resulting in the production of H2S. H2S S-sulfurates (sulfhydrates) cysteine residues of the p65 subunit of NF-κB to facilitate the interaction with another subunit RPS3, leading to upregulation of antiapoptotic genes (72).
H2S-producing pathway from
Description of Key Finding 3
Astrocytes, a type of glia, were thought to merely support and provide nutrients to neurons, but were recently recognized as actively regulating neuronal activity by releasing gliotransmitters such as
H2S3 induced Ca2+ influx with an EC50 value (91 nM), approximately 1/1000th of that of H2S (116 μM) (Fig. 4). The effect of H2S3 was suppressed by inhibitors as well as siRNAs specific to transient receptor potential ankyrin 1 (TRPA1) channels, suggesting that H2S3 activates TRPA1 channels (41). In collaboration with Dr. Ohta, Dr. Kimura identified the target of H2S3 as two cysteine residues located at the amino terminus of TRPA1 channels (20). This finding suggests the additional mechanism for the facilitation of LTP induction (see Key Finding 1, and Fig. 6). H2Sn activates TRPA1 channels to induce Ca2+ influx in astrocytes (41, 59, 66), which, in turn, release gliotransmitters such as
Other roles of H2Sn were subsequently found (30). In collaboration with Dr. Ogasawara, Dr. Kimura found that H2Sn facilitate the translocation of nuclear factor-like 2 (Nrf2) to the nucleus by modifying its binding partner kelch-like ECH-associated protein 1 (Keap1) to upregulate the transcription of antioxidant genes (44) (Fig. 7). This system was initially reported to be activated by H2S (7). H2Sn regulate the activity of the tumor suppressor phosphatase and tensin homolog (18) and reduce blood pressure by dilating vascular smooth muscle through the activation of protein kinase G1α (80). These studies facilitated the identification of the production pathways for H2Sn.
Hylin and Wood reported that persulfurated cysteine residues of proteins were produced from 3MP, a substrate of 3MST (22). Bound sulfane sulfur, which is defined as the sulfur species that releases H2S under reducing conditions, includes H2Sn, free cysteine persulfide (Cys-SSH), GSH persulfide (GSSH), and persulfurated cysteine residues of proteins (24, 54, 63, 89). Dr. Kimura's group found that cells expressing 3MST contain higher levels of bound sulfane sulfur than cells expressing defective mutants of 3MST as well as cells without 3MST (76). Oral administration of
During studies on H2Sn production by 3MST, Dr. Kimura's group noticed that levels of cysteine and GSH were decreased when 3MST produces H2Sn. Dr. Kimura concluded that H2Sn readily reacted with Cys-SH and GSH to produce Cys-SSH and GSSH (40). Alternatively, 3MST may transfer sulfur from 3MP to cysteine and GSH to produce these persulfurated species (Fig. 8).
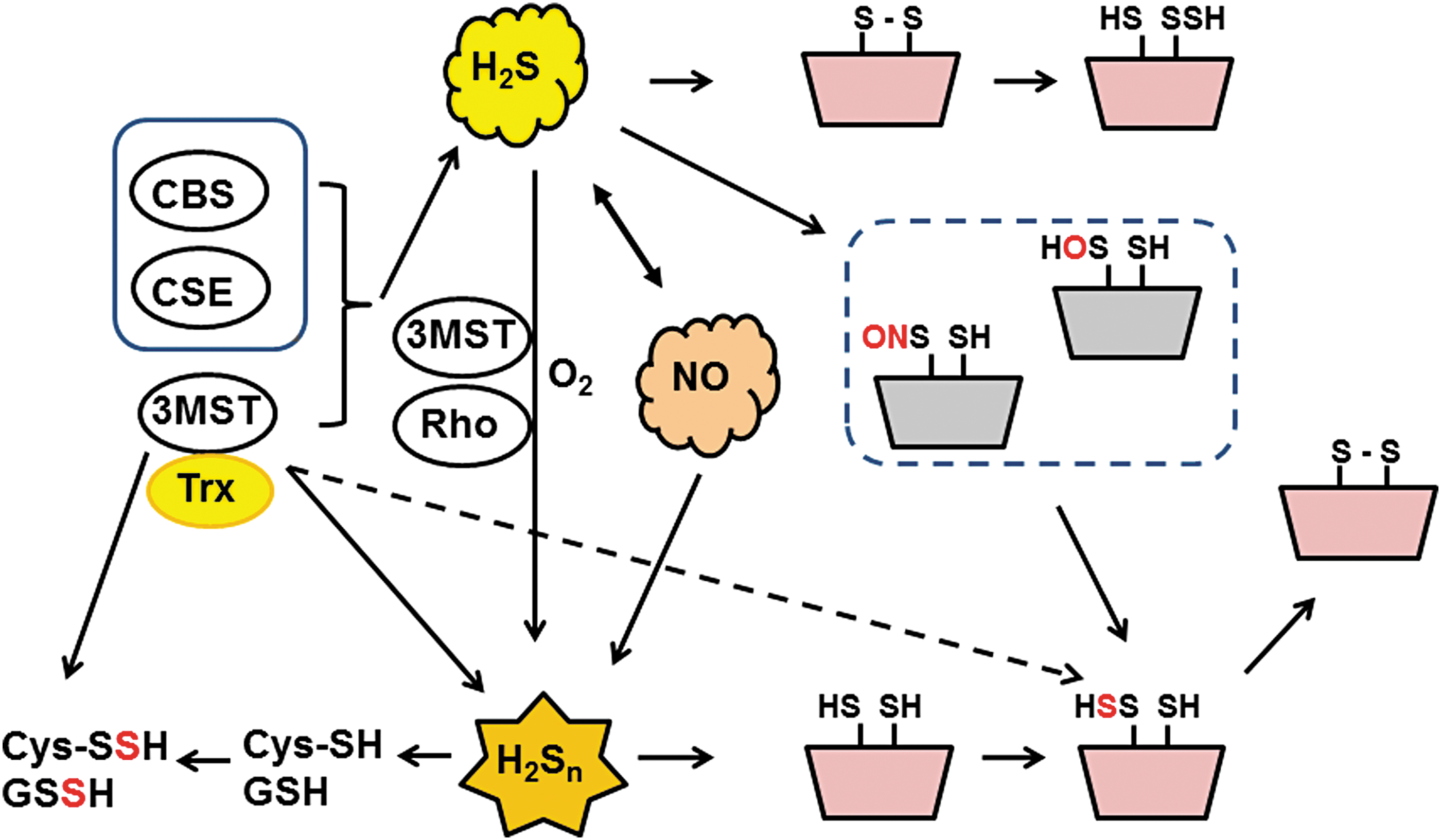
3MST requires a reducing substance to produce H2S, but an endogenous reducing substance was not known. From the structure of leishmania 3MST, which contains a thioredoxin domain in the molecule, the interaction of thioredoxin with 3MST had been predicted (90). Dr. Kimura's group determined thioredoxin as well as dihydrolipoic acid as potential endogenous reducing molecules necessary for 3MST to produce H2S (50, 57, 92).
3MST thiolates tRNA to maintain the accuracy of the genetic code and stabilize the tRNA structure (46, 91). There are two isoforms of 3MST that localize to the cytosol and mitochondria in humans (15). Cytosolic isoforms thiolate tRNA, whereas the mitochondrial isoform has a dual localization in both mitochondria and the cytosol, and not only thiolates tRNA in the cytosol but also supplies sulfur for iron–sulfur cluster formation in mitochondria (15). It is interesting to note that cysteinyl-tRNA synthetase was recently reported to have the activity to produce Cys-SSH (3).
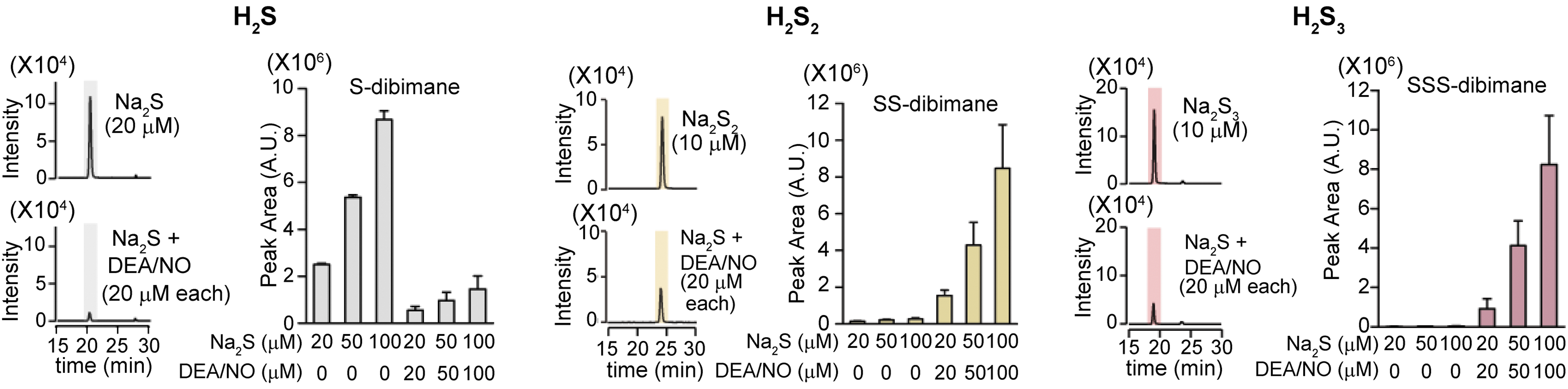
Eberhardt et al. and Cortese-Krott et al. reported that the chemical interaction of H2S with NO produces HNO and SSNO−, respectively (10, 13). Both groups showed H2Sn formation, but they did not consider their physiological relevance. Whereas Eberhardt et al. concluded that HNO is the chemical species responsible for activating the TRPA1-CGRP neuroendocrine signaling cascade and postulated that this pathway is essential for control of the general vascular tone, Dr. Kimura's group determined that H2S2 and H2S3 are the molecules that activate the TRPA1 channels of dorsal root ganglion neurons (52) (Fig. 5). Mustafa and Habara reported that H2Sn must be produced by the interaction of H2S and NO in mast cells (53). Molecules, which are produced from H2S and NO and activate TRPA1 channels, and H2Sn are degraded by cyanide (cyanolysis) and by reduction (52). In contrast, HNO is resistant to cyanolysis and SSNO− to reduction (10, 13). Considering these observations, Kimura's group suggested that the production of H2Sn from H2S and NO may be one of the mechanisms for the synergistic effects of both molecules on various tissues, including vascular smooth muscle relaxation (12, 21).
Other Achievements
In collaboration with Dr. Nagano, Dr. Urano, and Dr. Hanaoka, Dr. Kimura's group contributed to the development of H2S- and polysulfide-fluorescence probes (52, 69, 83) as well as inhibitors specific to 3MST (19).
Current Position
Dr. Kimura is a specially appointed researcher, National Institute of Neuroscience, National Center of Neurology and Psychiatry. He was recently appointed as Professor and Faculty of Pharmaceutical Science at the Tokyo University of Science, Yamaguchi (renamed Sanyo-Onoda City University in 2018), Japan.
He received the Promoting Award from the Japanese Pharmacological Society (1988), Human Frontier Science Program (1990), First Award from National Institute of Health (1994), Alzheimer Scholar Award from Alzheimer Association (1994), JB Award from Japanese Biological Society (2010), Research Front Award from Thomson Reuters (2016), Highly Cited Researcher from Clarivate Analytics (2017), and President Award from the National Institute of Neuroscience, National Center of Neurology and Psychiatry (2018). He has served as a nominator of Japan Prize since 2008.
Footnotes
Acknowledgments
Dr. Hideo Kimura would like to thank Professor David Schubert for encouraging and supporting him to promote the study of hydrogen sulfide. He also thanks late Professor Yutaka Sakai, Professors Hideomi Fukuda, Hiroshi Saito, Toshiharu Nagatsu, Sadaaki Iwanaga, Keizo Inoue, and Eijiro Ozawa for their suggestions and for supporting him. Dr. Kimura thanks all colleagues during their stay in the laboratories for their contribution to the study: Drs. Kazuho Abe, Norihiro Shibuya, Isao Ishii, Yasushi Enokido, Yasuo Nagai, Ken Umemura, Yoshinori Mikami, Kazuhiko Namekata, Mamiko Tsugane, Mari Ishigami, Shin Koike, Noriyuki Akahoshi, Yuriko Kobayashi, Makiko Tanaka, Yukiko Toyofuku, Ryo Miyamoto, and Yuka Kimura. He also thanks Professors Norio Matsuki, Makoto Suematsu, Yuki Ogasawara, Sachio Takashima, Noriyuki Nagahara, David Lefer, Kazuyuki Ishii, Tetsuo Nagano, Yasuteru Urano, Kenjiro Hanaoka, Toshihide Kimura, Yukiko Kaneko, Rumiko Hosoki, Akiko Ichinohe, Takeshi Kanaumi, and late Professor Ichiro Niki for their collaborations.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.