
Abstract
Aims:
Although the human pathogen, Mycobacterium tuberculosis (Mtb), is strictly aerobic and requires efficient supply of oxygen, it can survive long stretches of severe hypoxia. The mechanism responsible for this metabolic flexibility is unknown. We have investigated a novel mechanism by which hemoglobin O (HbO), operates and supports its host under oxygen stress.
Results:
We discovered that the HbO exists in a phospho-bound state in Mtb and remains associated with the cell membrane under hypoxia. Deoxy-HbO carries an autokinase activity that disrupts its dimeric assembly into monomer and facilitates its association with the cell membrane, supporting survival and adaptation of Mtb under low oxygen conditions. Consistent with these observations, deletion of the glbO gene in Mycobacterium bovis bacillus Calmette-Guerin, which is identical to the glbO gene of Mtb, attenuated its survival under hypoxia and complementation of the glbO gene of Mtb rescued this inhibition, but phosphorylation-deficient mutant did not. These results demonstrated that autokinase activity of the HbO modulates its physiological function and plays a vital role in supporting the survival of its host under hypoxia.
Innovation and Conclusion:
Our study demonstrates that the redox-dependent autokinase activity regulates oligomeric state and membrane association of HbO that generates a reservoir of oxygen in the proximity of respiratory membranes to sustain viability of Mtb under hypoxia. These results thus provide a novel insight into the physiological function of the HbO and demonstrate its pivotal role in supporting the survival and adaptation of Mtb under hypoxia.
Innovation
Although Mycobacterium tuberculosis (Mtb) is strictly aerobic, it can survive long stretches of oxygen limitation. The mechanism by which it adapts under low oxygen is not known. We discovered for the first time that hemoglobin O (HbO) carries a redox-dependent autokinase activity that changes its oligomeric state and facilitates its membrane association, thereby sustaining availability of oxygen to its host under hypoxia. This study thus provides a novel insight into the physiological function of hemoglobin, HbO, in Mtb and demonstrates its pivotal role in supporting the survival and adaption of its host under hypoxia.
Introduction
M
Mtb produces two single-domain heme proteins known as truncated hemoglobins, hemoglobin N (HbN) and hemoglobin O (HbO), at different phases of its growth cycle (8, 25). Their identical counterparts are also present in Mycobacterium bovis (11). The HbN of Mtb and M. bovis bacillus Calmette-Guerin (BCG) carries a potent nitric oxide (NO) scavenging activity (21, 24) and protects the host during intracellular infection (2, 27). Recently, it has been shown that the HbN gets posttranslationally modified by mannosylation in Mtb and modulates host/pathogen interactions during intracellular infection (2). The physiological function(s) of the HbO still remains unknown. Different functions, including involvement in cell respiration (15, 25), redox reactions (20), peroxidatic activity (3, 22), and environmental stress management (4, 22), have been proposed for the HbO of Mtb and Mycobacterium leprae. The complexity in understanding the cellular function(s) of truncated hemoglobins arises due to the fact that these are able to perform a variety of chemical reactions in vitro that may not reflect on their real physiological function(s).
The glbO gene, encoding HbO, has been identified in the majority of fast and slow growing mycobacteria and may be important for their cellular metabolism. In fact, the obligate intracellular pathogen, M. leprae, which has undergone reductive evolution to retain a minimum set of genes essential for its pathogenic lifestyle, lacks HbN but carries the HbO encoding gene (33), which suggests its essentiality for the mycobacteria. A basal level of HbO has been detected during the entire growth cycle of Mtb and M. bovis (20, 25), indicating its importance for the cellular metabolism of its host. In addition, a significant increase in the biosynthesis of HbO in Mtb during hypoxia (12, 26) also suggests its requirement under low oxygen, which the tubercle bacillus faces during its pathogenic life cycle. Thus, deciphering the molecular mechanism by which HbO functions in its host will be important to understand its physiological role in cellular metabolism and pathogenicity of Mtb and develop a novel target for designing more effective antimycobacterial drugs in tuberculosis treatment.
In this study, we have discovered a novel mechanism by which HbO may function in its native host. Here we have demonstrated that the HbO carries an autokinase activity and exists in a phospho-bound state in Mtb. It gets autophosphorylated in a deoxy state, which results in dissociation of its dimeric assembly into monomer that readily associates with the cell membranes and facilitates oxygen uptake. This study suggests a vital role of the HbO in supporting the adaptation and survival of its host under oxygen-limiting conditions.
Results
Mtb HbO carries an autokinase activity and gets phosphorylated in vitro
In silico analysis of Mtb HbO revealed that it carries a putative ATP-binding motif (GGAKT) within the connecting loop of A and B helices, resembling a deviant form of P-loop (xKT/S) motif (Fig. 1A), present in some ATP-binding proteins (14, 19). In addition, a putative phosphorylation site (TLDD) was also identified at the C-terminal region of HbO (Fig. 1A). In the X-ray structure of HbO dimer (18), these motifs are closely spaced in each subunit and are away from the interface (Fig. 1B, C), suggesting the possibility of posttranslational modification of the HbO via intrasubunit phosphotransfer. This assumption was substantiated experimentally by in vitro kinase assays, which demonstrated rapid incorporation of γ-32P from ATP on the HbO in a concentration-dependent manner (Fig. 1D and Supplementary Fig. S1) that was inactivated in the presence of anti-HbO polyclonal antibodies (Fig. 1E). Optimal autokinase activity of the HbO was observed in the presence of Mg++ that was changed marginally by other divalent ions, except Co++, which reduced this activity significantly (Fig. 1F), indicating that Co++ is not favored as a cofactor or the Co-ATP complex is unable to activate kinase activity of the HbO. To validate the phosphorylation site of HbO experimentally, we carried out autophosphorylation of HbO in vitro and checked intact mass as well as tryptic digest of the protein through mass spectrometry. Phospho-bound HbO exhibited the increase of nearly 950 Da in the protein mass (Supplementary Fig. S2), which indicated incorporation of single phosphate group with the protein. A single phospho-bound peptide was identified and the liquid chromatography with tandem mass spectrometry (LC-MS-MS) spectra confirmed the phosphate group on Thr103 (Fig. 2A, B, and Supplementary Fig. S3). The ATP binding and phosphorylation sites of HbO were further verified after site-directed mutagenesis of Lys13 within the ATP-binding loop and Thr103 of the phosphorylation site, which abrogated autophosphorylation ability of the HbO (Fig. 2D). The circular cichroism spectra suggested that the secondary structures of these mutants are similar to wild-type HbO (Supplementary Fig. S4). Phosphorylation of the HbO did not occur when Lys13 and Thr103 mutants were included together in a kinase reaction in 1:1 molar ratio, which ruled out the possibility of intersubunit phosphotransfer (Fig. 2D). Phospho-threonine (phospho-Thr) antibodies identified the wild-type HbO, but not the Thr103 mutant (Fig. 2E), suggesting that Mtb HbO is specifically phosphorylated at the Thr103 residue. These complementary approaches confirmed that the HbO carries an autokinase activity and gets phosphorylated by intrasubunit phosphotransfer at the Thr103 residue.

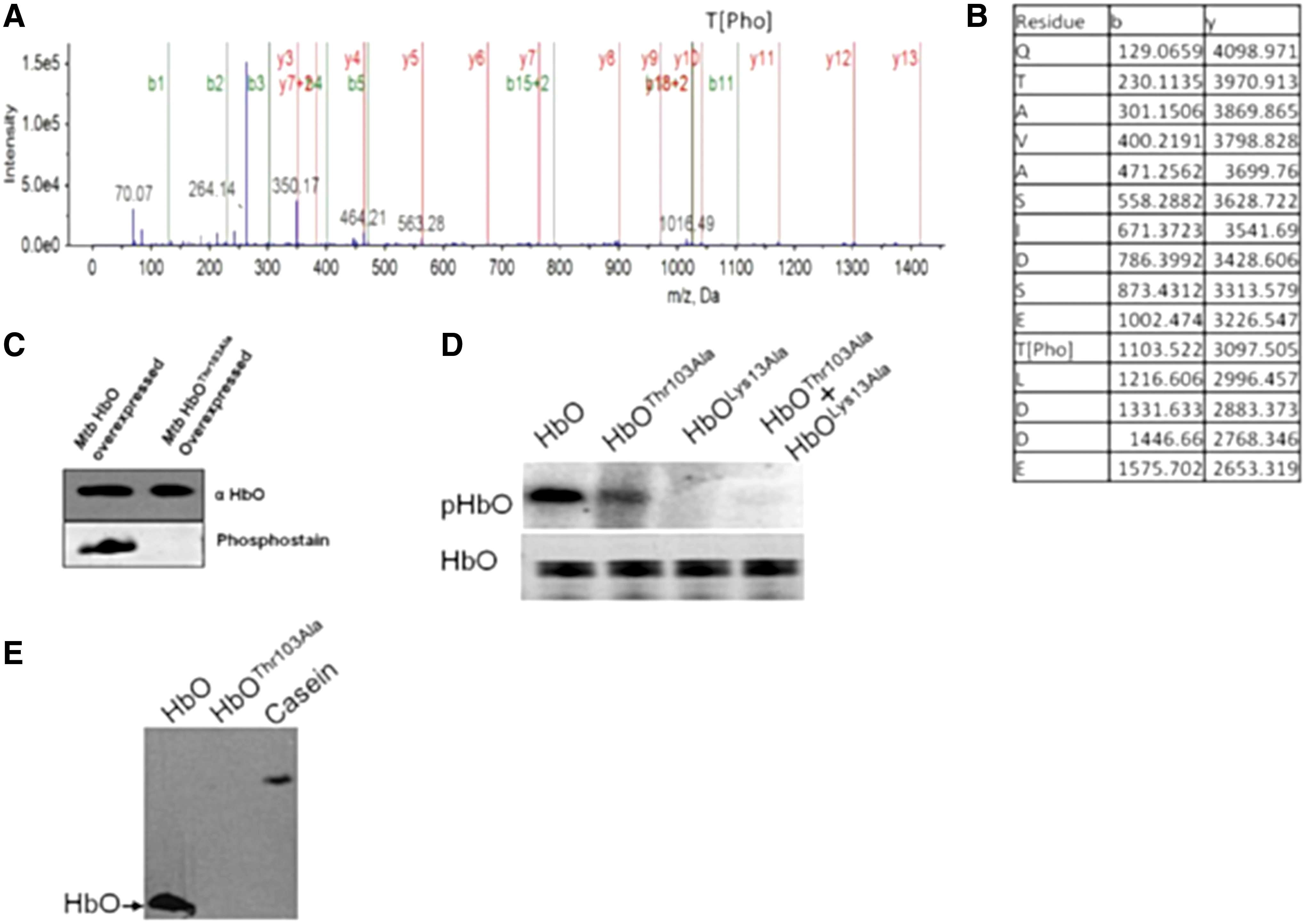
The HbO exists in a phospho-bound state in Mtb
To corroborate the in vitro results, we investigated the in vivo status of HbO in its native host. Therefore, we overexpressed the wild-type and the HbOThr103Ala mutant in Mtb and isolated the protein after coimmunoprecipitation using HbO monospecific polyclonal antibodies. When these protein samples were probed with phosphostaining, a positive signal appeared only with wild-type HbO but not with HbOThr103Ala mutant (Fig. 2C). Simultaneously, we separated total cellular proteins of the HbO overexpressing Mtb and its isogenic control cells. Proteomic profiles of these cells were analyzed after two-dimensional gel electrophoresis, and an HbO spot was identified on parallel gels after general protein stains and phospho-protein staining (Supplementary Fig. S2). The position of HbO spot on the gel was checked separately after Western blotting and authenticity of the HbO spot was confirmed through LC-MS-MS analysis of its tryptic digest and the peptides, which matched with the HbO sequence (Supplementary Fig. S5e). Taken together, these results confirmed that HbO exists in a phospho-bound state in its native host.
Autokinase activity of the HbO is regulated by its redox states
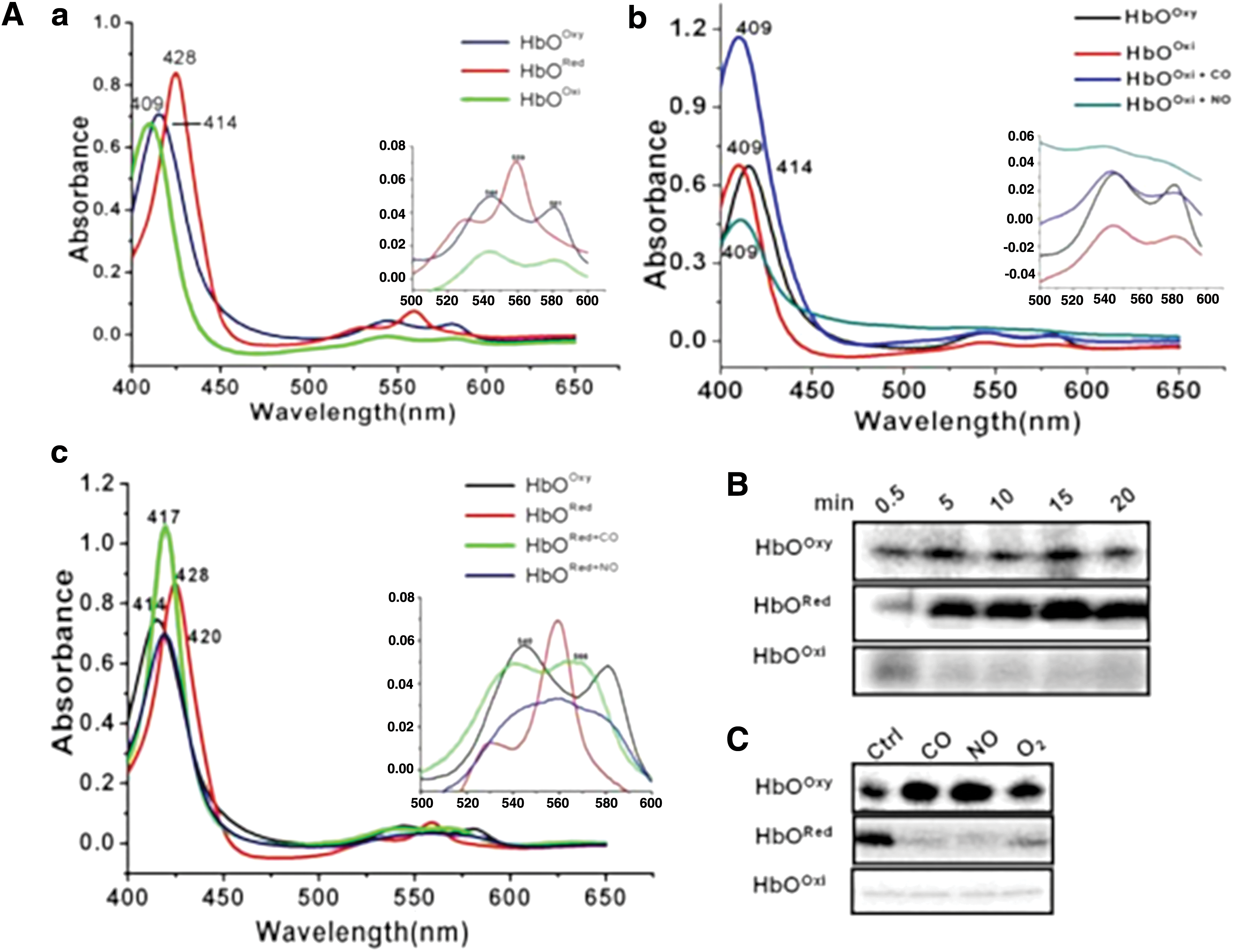
Since heme proteins can interact with various gaseous ligands and are key regulators of adaptive responses toward redox changes in the cell, we checked whether these are regulatory ligands for the autokinase activity of HbO. Spectral characteristics of HbO in ligand-bound and ligand-free states were checked (Fig. 3A and Supplementary Fig. S6), which matched closely with previous reports (20). Optical spectra of the oxygenated HbO exhibited a sharp Soret peak at 414 nm along with ὰ and β peaks at 544 and 581 nm, respectively, whereas ferric species of the protein appeared as a mixture of low- and high-spin heme, having Soret peak at 409 nm and ὰ and β peaks at 540 and 580 nm, respectively. The ligand-free ferrous species of the HbO displayed a penta-coordinated high-spin state with absorption maxima at 428 and 559 nm (Fig. 3A, panel a). In vitro kinase assay demonstrated that the autokinase activity of oxy-HbO remains low but increases significantly in a deoxy state (Fig. 3B). A time-dependent increase in the autophosphorylation of the HbO occurred in a deoxy state, which decreased significantly in the presence of O2 and remained completely blocked in the presence of gaseous ligands, NO and CO (Fig. 3C), as opposed to autophosphorylation of oxyHbO, which did not change much when exposed to these gaseous ligands. In contrast, oxidized HbO(Fe3+) did not show any reaction alone or in the presence of any gaseous ligand (Fig. 3C and Supplementary Fig. S7). Interactions of ligands (NO, CO) with different redox states of the HbO were checked spectrophotometrically (Fig. 3A, panels b, c). These results suggest the involvement of HbO in sensing of oxygen and/or redox states of the environment.
Autophosphorylation disrupts dimeric assembly of the HbO into monomer
Since the redox state of HbO modulates its autophosphorylation ability, we checked the subunit association properties of phospho-bound HbO in vitro and analyzed its oligomeric states in cells growing under aerobic and hypoxic conditions. Recombinant HbO, isolated from aerobically growing cells of Escherichia coli, appeared mainly in an oxygen-bound state as a mixture in dimer to monomer ratio of ∼3:1 (Fig. 4A, panel I). When this preparation of the HbO was subjected to in vitro kinase reaction, it was converted into monomer (Fig. 4A, panel III), suggesting that subunit contacts of the HbO are disrupted after ATP binding and phosphorylation. To understand the functional correlation between autophosphorylation and physiological states of HbO in vivo, we further checked the oligomeric state of HbO isolated from cells grown under hypoxia. Interestingly, HbO isolated from hypoxia grown cells appeared largely as a monomer, having monomer and dimer in the ratio that varied from 2:1 to 3:1 (Fig. 4A, panel II), unlike aerobic condition, where the major fraction of HbO appeared in a dimeric state. The oligomeric state of HbO collected from different gel filtration fractions was confirmed on native polyacrylamide gel electrophoresis (PAGE) to validate changes in its subunit association properties (insets of Fig. 4A and Supplementary Fig. S8).
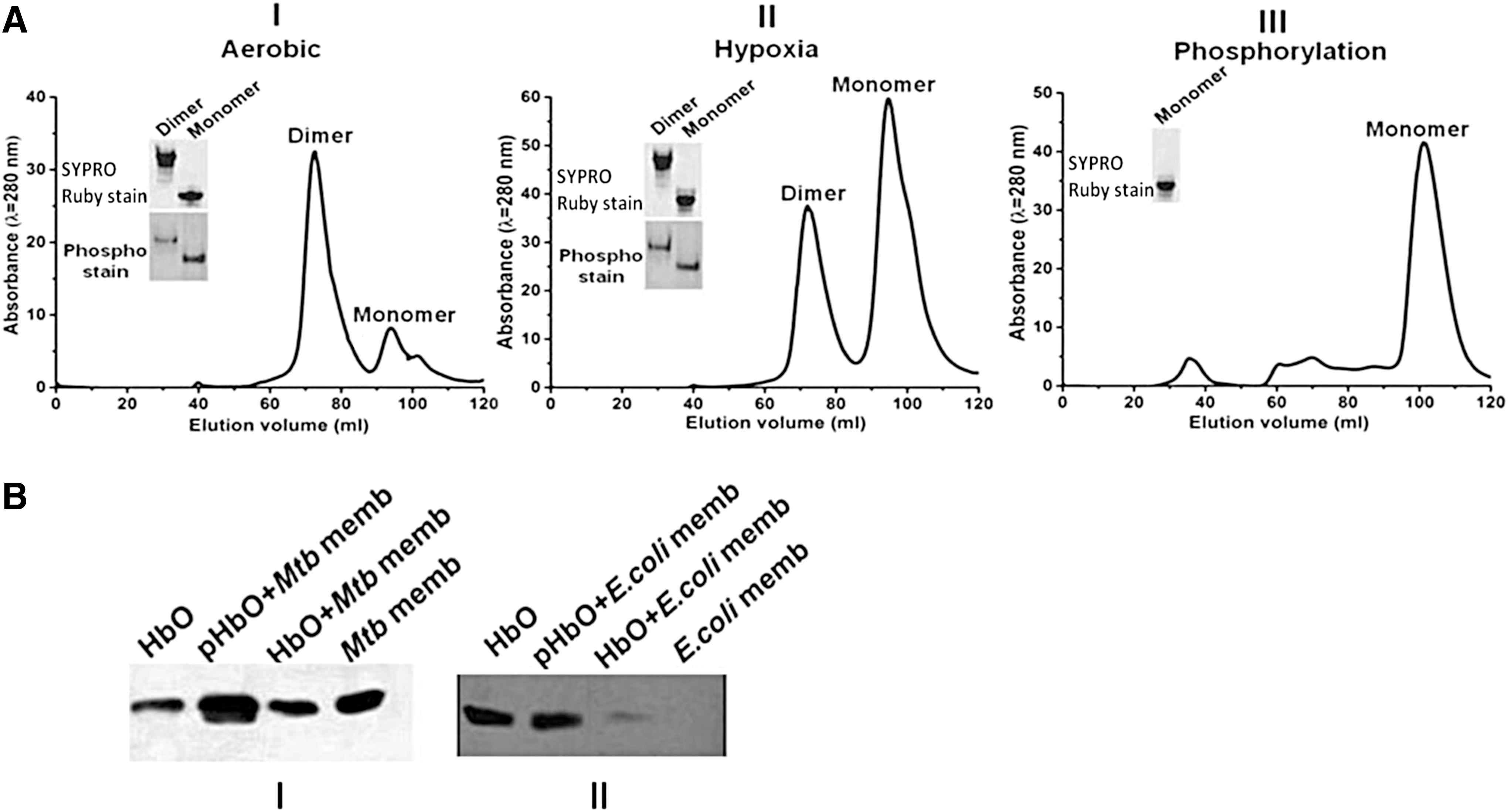
Since the oligomeric state of the protein plays a vital role in controlling the function of a protein, we investigated whether Mtb HbO also exists in different oligomeric states in its native host. When whole-cell lysates of Mtb cells, grown under aerobic condition and hypoxia, were analyzed on nonreducing 15% sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE), two distinct bands, corresponding to the HbO monomer (14 kD) and dimer (28 kD), were identified on Western blots using anti-HbO antibodies (Supplementary Fig. S9), as opposed to reducing SDS-PAGE, where HbO has been identified as single band in the cell lysate of Mtb (20, 25). Although it is difficult to accurately determine the ratio of monomer and dimer of the HbO in Mtb under aerobic and microaerobic conditions, these results suggest that HbO exists in two oligomeric states in Mtb.
Phospho-bound HbO monomer associates with the cell membrane
Primary studies on subcellular fractions of Mtb indicated that the HbO is localized in both cytoplasmic and membrane fractions of aerobically growing cells, and therefore, we analyzed membrane association properties of HbO in different oligomeric states to check whether cellular localization of the HbO, in any way, is related to its oligomeric state. When different oligomeric forms of the HbO, collected from gel filtration chromatography, were allowed to interact with the cell membrane of Mtb, the signal for the phospho-bound HbO monomer appeared significantly higher after Western blotting (Fig. 4B, panel I and Supplementary Fig. S10), whereas the signal remained nearly the same in the case of unphosphorylated HbO and HbO dimer, which corresponded to the signal observed in the control membrane without any HbO interactions, possibly due to the background of native HbO already present in Mtb. These results were further supported by the observation that respiratory membranes of Mtb retained a higher amount of phosphorylated HbO in comparison with native HbO and other globins during gel filtration analysis (Supplementary Table S1). Furthermore, we checked whether the association of HbO is specific to the cell membrane of Mtb and tested the interactions of HbO with the cell membrane of HbO lacking heterologous host, E. coli. The membrane fraction of E. coli did not show any signal after Western blotting with monospecific HbO antibodies. When the cell membrane of E. coli was incubated with phosphorylated HbO, a strong signal appeared compared with the one incubated with the HbO, purified from aerobically growing recombinant E. coli (Fig. 4B, panel II), where HbO predominantly exists in a dimeric state. These results indicated that the membrane association properties of the HbO may not be restricted to Mtb only.
Disruption of the glbO gene attenuates growth and cell viability of M. bovis BCG under hypoxia
Genomic sequences of the glbO gene and its upstream regulatory region are identical in Mtb and M. bovis (11), suggesting similar function(s) of the HbO in these mycobacterial strains. Therefore, for safety reasons we used M. bovis BCG to disrupt the glbO gene and used it as a model to investigate the physiological function of HbO. Disruption of the glbO gene was confirmed through polymerase chain reaction (PCR) analysis of the genome of wild-type and mutant strains (Supplementary Fig. S11A) and Western blot analysis of whole-cell lysates of wild-type and glbO gene-disrupted strains (Supplementary Fig. S11B), which confirmed lack of HbO biosynthesis in the mutant strain of M. bovis BCG. The selected mutant strain, thereafter, was designated as ΔglbO M. bovis BCG. Growth profiles of wild-type and the ΔglbO M. bovis BCG appeared more or less similar under aerobic conditions (Fig. 5A), but cells of ΔglbO M. bovis BCG did not show any distinct increase in the cell density under hypoxia (Fig. 5B), whereas the parental wild-type strain exhibited a slow and steady increase in optical density under similar conditions. Complementation of the glbO gene of Mtb in ΔglbO M. bovis BCG rescued its growth inhibition under hypoxia, whereas expression of phosphorylation-deficient mutant of the HbO (HbOThr103Ala) did not show any advantage on cell growth of ΔglbO M. bovis BCG under similar condition (Fig. 5B). These results demonstrated that autophosphorylation plays an important role in modulating the physiological function of HbO in M. bovis BCG as well as in Mtb. A significant increase in the cell density was observed under hypoxia when Mtb HbO was overexpressed in wild-type M. bovis BCG (Fig. 5B), which further substantiated that the presence of HbO confers growth advantage to its host under low oxygen.

We further investigated the role of HbO on cell viability under hypoxia after checking colony forming unit (CFU) counts of ΔglbO M. bovis BCG and HbO expressing ΔglbO M. bovis BCG and compared it with the isogenic wild-type counterpart. An equal number of cells (normalized by their optical density) were taken out and plated at different stages of their growth. Surprisingly, CFU counts of ΔglbO M. bovis BCG appeared very low compared with HbO expressing ΔglbO M. bovis BCG and its wild-type counterpart even at the starting point (Fig. 5D). The survival rate of ΔglbO M. bovis BCG, as determined by CFU counts, appeared 30 to 40% lower than the wild type and the HbO expressing ΔglbO M. bovis BCG (Fig. 5D). These results demonstrated that the presence of HbO increases the cell viability of its host and might be helpful in sustaining its survival.
HbO modulates the oxygen uptake ability of its host
To investigate how HbO enhances the ability of its host to grow and survive better under low oxygen, we set up experiments in a hypoxia model (6) to check the oxygen utilization ability of ΔglbO M. bovis BCG and compared it with the wild-type and HbO overexpressing counterpart. This was accomplished by culturing these strains in a sealed glass vial, thus limiting the total amount of oxygen available in the head space. Methylene blue was added in the culture to analyze depletion of oxygen during growth of these strains. Reduction in the color of this dye served as a visual indication of oxygen depletion, and OD600 measurements at regular intervals indicated the relative efficiency of oxygen consumption by these strains. No color change appeared in the vials having ΔglbO M. bovis BCG even after 20 days, indicating that the oxygen uptake ability of these cells might be attenuated. However, ΔglbO M. bovis BCG, complemented with Mtb HbO, exhibited a slow decrease in the color of the dye after 10 days, similar to the isogenic wild-type strain, indicating gradual uptake of oxygen by these cells. M. bovis BCG, overexpressing Mtb HbO, consumed oxygen more rapidly than all other strains (Fig. 5C). Decolorization of methylene blue occurred faster and was clearly visible after 40 days in HbO expressing Mtb compared with the isogenic wild type (Supplementary Fig. S12). Taken together, these results demonstrated the involvement of HbO in modulating the oxygen uptake ability of its host.
Discussion
The present study provides a novel insight on the physiological function of Mtb HbO and demonstrates for the first time that it carries a redox-dependent autokinase activity that allows its recruitment near the cell membrane and modulates cellular metabolism of Mtb under low oxygen. Being a strict aerobe and intracellular pathogen, Mtb cells would require an efficient mechanism for the trapping of oxygen and its delivery to the cell membrane for sustaining the respiratory activity during long exposure of hypoxia. The high oxygen binding affinity and slow oxygen releasing rate (20) of membrane-associated HbO may allow it to sequester and collect oxygen near the respiratory apparatus and support slow and steady respiration of Mtb for an extended period of oxygen limitation. This is supported by the observations that HbO exists in a phospho-bound state and being associated with the cell membrane in its native host promotes its growth and viability under oxygen limitation. Owing to the fact that tubercle bacillus encounters severe hypoxia during its intracellular regime and HbO undergoes posttranslational modification to alter its oligomeric state that enhances its proximity to the respiratory membranes and improves its oxygen uptake, the existence of HbO may be crucial during the pathogenic life cycle of Mtb within the macrophages and calcified granuloma, where it resides during long latent periods of infection and faces severe oxygen paucity (1, 31, 32).
Mtb HbO appeared unique among single-domain hemoglobins in having an ATP-binding motif and a phosphorylation site within the truncated globin fold, which gave initial indication for its posttranslational modification in vivo via autophosphorylation. Recombinant HbO from aerobically growing cells of E. coli has been purified mainly as a dimer, which was converted into a monomeric state after in vitro kinase reaction. Proximity of ATP-binding and phosphorylation sites in each subunit, away from the interface in the X-ray structure of the HbO (18), suggested the possibility of intrasubunit phosphotransfer during phosphorylation of HbO, which might affect protein backbone motion and/or alter protein dynamics to disrupt its intersubunit contacts. It seems credible in view of a very small association area at the interface and weak intersubunit contacts between two monomeric units of the HbO (Supplementary Table S2). This is supported by the observation that the highest complex formation significance score of potential dimer forming chains, A/G in the dodecamer structure of the HbO, is 0.156 and the surface area between these chains spans only 723.1 and 624.5 Å2 for G and A chains, respectively. Such weak intersubunit contacts have been found advantageous in protein systems that require dynamical movement of subunits to mediate important cellular functions (16). This may also be true for the HbO. The autokinase activity of Mtb HbO increases maximally in a deoxy state, suggesting its role in oxygen and/or redox sensing. It has been reported that transcription of the glbO gene gets upregulated in Mtb under hypoxia and during macrophage infection (12, 26), and thus, the involvement of HbO under these conditions may be envisaged. This is supported by the observations that disruption of the glbO gene attenuates growth of M. bovis BCG under hypoxia and complementation of Mtb glbO gene rectifies these inhibitory effects. In addition, overexpression of the glbO gene in Mtb enhances its oxygen uptake and cell viability under hypoxia, and advantages conferred by the HbO to its host are drastically reduced during expression of a kinase dead mutant of HbO. These results demonstrated that autophosphorylation of HbO plays a vital role in modulating the function of HbO. The physiological role of dimeric HbO remains unknown at present. Our initial studies indicated that the expression of glbO gene may not affect hypoxia-related DevR regulon of Mtb but may be under the control of sigma factor, SigH (unpublished results), which is involved in the management of multiple stresses, including hypoxia (17). Thus, involvement of HbO in modulating the physiology of Mtb under other environmental stresses cannot be ruled out.
Existence of HbO is ubiquitous in mycobacteria, including M. leprae (33) that has undergone reductive evolution to retain a minimum set of genes required for its pathogenic life cycle, suggesting that the presence of HbO is required for the cellular metabolism of its host. It is intriguing why mycobacteria, being a single-cell organism, need an oxygen binding protein. Although most mycobacteria are strict aerobe, their cell wall is highly complex, thick, and impermeable, which might restrict easy diffusion of oxygen. Thus, the presence of an oxygen binding protein adjacent to the respiratory membrane of pathogenic mycobacteria such as Mtb, M. bovis may be highly beneficial for supporting their aerobic metabolism during their intracellular regime where they are exposed to long stretches of oxygen paucity. Localization of the HbO increases on the cell membrane in hypoxia grown cells of Mtb, indicating that close interaction of the HbO with respiratory membranes might be required during oxygen limitation. This is in line with our previous observations that recombinant E. coli and Mycobacterium smegmatis, expressing Mtb HbO, exhibit higher respiratory activities and the addition of purified HbO to the cell membrane of E. coli enhances its oxygen uptake (23). These in vitro studies implicate the role of HbO as an oxygen collector and supplier to allow the cells to maintain respiratory activities, required for their survival during long stretches of hypoxia and latency. Although the involvement of Mtb HbO in oxygen uptake has been ruled out due to in vitro observation (18, 20) that this protein carries a high affinity for oxygen binding and dissociates oxygen very slowly, our study suggests that HbO still can act as an oxygen sequester and supplier in its native host. Mtb is able to survive in its host for years by existing in a latent state where it remains viable and metabolizes, but at an extremely slow pace (31, 32). Thus, proximity of oxygen-bound HbO with the cell membrane under hypoxia and its slow oxygen releasing properties may be able to support slow metabolism and safeguard viability of its host during extended periods of hypoxia and latency by maintaining a buffer of oxygen near the respiratory membranes. In fact, if oxygen dissociation by the HbO would have been faster, then the contribution of HbO-mediated oxygen flux becomes insignificant for Mtb during long periods of oxygen limitation.
It has been reported that Mtb HbO interacts with the membrane lipids and mediates advantage on cell growth of E. coli (15). The molecular mechanism by which HbO interacts with the respiratory apparatus of its native host is not known at present. The autokinase activity of HbO increases in a deoxy state and disrupts its dimeric assembly into monomer that displays high affinity toward the cell membrane. The crystal structure of the HbO (18) provided an insight to explain the mechanism by which the HbO monomer might interact with cell membrane. The HbO carries a positively charged EF loop region, carrying five arginine residues and a histidine residue, which remains partially masked in dimeric assembly of the HbO and gets fully exposed in a monomeric state. In addition, a hydrophobic patch, spanning from 55 to 70 residues, remains buried at the interface of the HbO dimer and may get surface exposed after dissociation of the HbO subunits. Exposure of these hydrophobic and positively charged regions in the HbO monomer may allow it to interact strongly with negatively charged cell membrane. This is supported by the fact that phospho-bound HbO associates strongly with the cell membrane of both Mtb and E. coli. Our study suggests that phosphorylation of HbO in Mtb might be important for its recruitment on the cell membrane. This is in line with our observation that the expression of a kinase dead mutant of the HbO is unable to provide growth and survival advantage to its host under hypoxia, unlike native HbO. It is likely that the contact between HbO and the cell membrane may also affect the oxygen affinity of protein that may facilitate unloading of oxygen close to the respiratory apparatus. As a matter of fact, lipid-mediated changes in the oxygen affinity of Vitreoscilla hemoglobin (VHb) have been observed previously (30).
Taken together, this study unravels a novel mechanism by which HbO may function in its native host. Functional properties of the HbO are modulated by its ability to get autophosphorylated in a redox-dependent manner. Our experimental studies support the dual role of HbO in mycobacteria: as an oxygen sensor that senses the paucity of oxygen and transduces the signal to change the oligomeric state of the protein to facilitate the association of protein with cell membrane. Accumulation of oxygenated HbO in the proximity of the respiratory apparatus may facilitate required delivery of oxygen to the respiring membranes and sustain the energy status of Mtb during long exposure of hypoxia and latency due to its unique oxygen binding properties. One of the bottlenecks in tuberculosis control is the latency of the tubercle bacillus when it respires slowly and maintains extremely slow metabolism, which makes it refractory toward therapeutic drugs as well as host-generated bactericidal mechanisms. Thus, HbO might be an attractive system to target Mtb, including latent Mtb, for the development of new antimycobacterial agents.
Materials and Methods
Bacterial strains, plasmids, and growth conditions
E. coli, JM109 and BL21(DE3), were routinely used for cloning and expression of recombinant genes. Cultures of E. coli were grown in Luria–Bertani or Terrific broth (containing 24 g of yeast extract, 12 g of Bacto-peptone, 12.3 g K2HPO4, 2.3 g KH2PO4). Mtb H37 Ra, M. bovis BCG, and M. smegmatis strains of mycobacteria were used for experimental work. Mycobacterial cultures were grown in Middlebrook 7H9 medium (Difco) having 10% (vol/vol) oleic acid-albumin-dextrose catalase enrichment (Difco), 0.5% glycerol, and 0.05% Tween-80 in liquid culture, and Middlebrook 7H10 agar (Difco) was used as solid medium. When required, kanamycin (30 μg/mL), hygromycin (50 μg/mL), and ampicillin (100 μg/m) were added for the selection. Cultures were grown at 37°C at 200 rpm unless otherwise stated. Bacterial growth was monitored by measuring OD600.
Cloning and expression of recombinant proteins
E. coli JM109 and E. coli BL21DE3 were used routinely for cloning and expression of recombinant genes. The glbO gene (Rv2470) was amplified from the genomic DNA of Mtb H37Rv (Mtb) using primer pairs, HbONdeIF (5′-GATCATATGCCGAAGTCTTTCTACGAC-3′) and HbOBamHIR (5′-GGATCCTCAAAACGGGGAGTTGACCAGCGA-3′), and amplified product was digested with restriction enzymes, NdeI and BamHI. The digested product was cloned in pET9b and pET28c for the expression in E. coli as a native or His-tagged protein, respectively, and named as pET9bHbO and pET28cHbO. Expression level of the HbO was checked on 15% SDS-PAGE and verified after Western blotting using polyclonal HbO-specific antibodies. Mutants of the glbO gene were generated using overlap PCR extension. Primer sets HbOATPKEX1TR (5′-GTCGGCGGCGCCGCAACCTTCGAC-3′), HboATPKEX2F (5′-TACGACGCGGTCGGCGGC-3′), and HbONdeI IF (5′-GATCATATGCCGAAGTCTTTCTACGAC-3′) were used for the mutation of Lys13 to Ala and the plasmid construct was named as pET28C HbOLys13Ala. Second primer sets HbOTXXDmutOVR (5′-GTGCTCGTCATCGAGGGCTTCTGA-3′) and HbOTXXDmutOVF (5′-TCAGAAGCCCTCGATGACGAGCAC-3′) were used for the conversion of Thr103 to Ala and the plasmid construct was named as pET28C HbOThr103Ala. These HbO mutants were designated as HbOLys13Ala and HbOThr103Ala. Authenticity of cloned genes was confirmed by DNA sequencing.
Construction of HbO overexpressing strain of mycobacteria
Construction of HbO expression plasmid for mycobacteria has been described previously (25). Briefly, the coding region of the glbO gene was amplified by PCR from the genomic DNA of Mtb, using gene-specific primer pairs, carrying a BamHI site at 5′ end and PstI site at 3′ end. The PCR product was digested with BamHI and PstI and cloned at complementary sites of mycobacterial expression vector, p19Kpro, under the constitutive promoter of the 19 kDa antigen of Mtb. Resulting expression plasmid was designated as p19KproHbO and transformed into Mtb and M. smegmatis. Expression of recombinant HbO in mycobacteria was visualized after running the cell lysate of HbO expressing mycobacteria on 15% SDS-PAGE and probing with HbO-specific polyclonal antibodies.
Purification of recombinant HbO and gel filtration analysis
Recombinant HbO, expressed in E. coli, was purified from the cell lysate as described previously (25). Subunit association properties of the HbO were analyzed after gel filtration chromatography on Superdex-75 column, equilibrated with 50 mM Tris.HCl and 100 mM NaCl (pH 8). For the ATP binding experiment, purified HbO was incubated with cold ATP in a kinase buffer (50 mM Tris.HCl, pH 7.4, 50 mM NaCl, 10 mM MgCl2) at 25°C for 6 h and then analyzed via gel filtration chromatography. Oligomeric states of the HbO, isolated after gel filtration, were checked on native PAGE.
Optical absorption spectra
Optical absorption spectra of the HbO were recorded using a Cary 100 spectrophotometer (Varian). Ferric form of the HbO was prepared at room temperature by oxidation of the oxy form using 10-fold excess potassium ferricyanide as described elsewhere (20). The protein was reduced in the presence of sodium dithionite, and oxy samples were prepared after exposing the ligand-free ferrous protein to air by vortexing vigorously. These protein samples of the HbO were exposed to gaseous ligands by bubbling CO, NO, or O2 for 1 min and were immediately used for phosphorylation assays.
In vitro kinase reaction and autophosphorylation assay
In vitro autokinase assay was performed by incubation of a specified amount of purified HbO in 20 μL of kinase buffer containing 2 μCi of [γ-32P] ATP) at 37°C for 40 min. The reaction was stopped after adding sodium dodecyl sulfate (SDS) containing gel loading buffer (50 mM Tris.HCl, pH 6.8, 1% beta- mercaptoethanol, 2% SDS, 0.1% bromophenol blue, and 10% glycerol). Phosphorylated protein was resolved by electrophoresis on a 15% SDS-PAGE and analyzed using a Phospho-Imager (FLA-9000; Fujifilm).
Determination of phospho-bound state of the HbO in vivo
Mycobacterial expression plasmid vector p19kpro, carrying wild-type or mutant HbO (HbOT103Ala), was transformed into Mtb via a standard electroporation procedure and recombinant cells expressing HbO were probed with HbO antibodies after Western blotting to confirm the expression of protein. Recombinant mycobacterial strains, expressing wild-type and mutant HbO (HbOT103Ala), were cultured into 7H9 liquid broth till the OD600 reached to 0.9 or 1. Twenty milliliters of each culture was harvested by centrifugation and lysed after sonication or by French Press. Cell lysate was centrifuged at 14,000 rpm for 20 min and the clear supernatant, thus obtained, was incubated with HbO-specific antibodies at 37°C for 1h and HbO was retrieved using the Protein G immunoprecipitation kit (Sigma) following the manufacturer's instruction. The HbO protein, thus obtained, was run on 15% SDS-PAGE, transferred on polyvinylidene difluoride membrane, and probed with Pro-Q Diamond phospho-stain (Invitrogen) and phospho-Thr antibodies (Invitrogen). These blots were further stripped in 2% SDS/100 mM DTT, 62 mM Tris (pH 7), for 30 min at 50°C and analyzed after probing with HbO-specific antibodies.
Mass spectrometric analysis of proteins
Protein samples of the HbO, before and after in vitro kinase reaction, were digested in gel with trypsin for 8 h and injected into the LC/MS system. Mass spectrometry measurements of the HbO were done in linear and reflector modes on a Voyager MALDI-TOF system (Applied Biosystems) equipped with an N2 laser source. Mass spectra of digestion mixtures or isolated peptides were acquired on linear and reflector modes using a matrix solution of cyano-4-hydroxycinnamic acid in 0.2% trifluoroacetic acid in acetonitrile-H2O (50%, V/V) and were externally calibrated using a mixture of peptide standards (Applied Biosystems). Mass spectra of trypsin-digested HbO were compared before and after phosphorylation. The product ion spectra were searched against a custom Mtb database using MASCOT.
Membrane binding assay
Native and phosphorylated forms of the HbO protein (10 μg) were incubated with the membrane fraction (80 μg) of E. coli at room temperature for 30 min to allow binding of protein to the membrane and thereafter centrifuged at 40,000 rpm to remove unbound HbO protein. The supernatant was then discarded and pellet was suspended in SDS gel loading buffer and loaded on 15% PAGE gel, followed by Western blotting after transferring the gel onto a nitrocellulose membrane and immunoprobing with anti-HbO antibodies.
Membrane association of the HbO was also checked via gel filtration analysis as described earlier (29) for VHb. Briefly, 50 mg of Sephadex G-100 was suspended in 1.5 mL of 0.1 M potassium phosphate buffer (pH 7.2) and allowed to swell for 4 h at room temperature. Thereafter, 0.5 mL of the buffer solution containing the membrane or the HbO alone or in combination was added. This suspension was incubated for 10 min at room temperature with continuous stirring and then allowed to settle down for 20 min and 100 μL aliquot of the supernatant was removed. The concentration of unbound HbO was checked in the supernatant by using heme absorption at 414 nm.
Disruption of the glbO gene encoding HbO
Since glbO gene and its upstream regulatory sequences of Mtb are identical to that of M. bovic BCG (11), we used M. bovis BCG to disrupt the glbO gene encoding HbO and used it as a model to evaluate the function of HbO. Disruption of the glbO gene in M. bovis BCG was carried out following the published procedure (23) in two crossover steps. Briefly, 1000 bp upstream and downstream regions of the glbO gene were amplified from the genomic DNA of M. bovis BCG and cloned on PmlI and KpnI sites of a plasmid vector, p2NIL (23). The hygromycin cassette from pGOAL19 (19), cloned into the single PacI site, was excised and cloned in between these two fragments at PacI site to generate suicide delivery vector p2NILHbO. This vector was pretreated with UV light and electroporated in M. bovis BCG, and transformants were selected on hygromycin/X-Gal plates. Deletion of the glbO gene in the selected transformants was confirmed via PCR and designated as ΔglbO.
Growth profile of mycobacteria under hypoxia
For checking the growth profile of mycobacteria under hypoxic conditions, a required amount of seed culture (1%) of wild-type, ΔglbO gene deleted and HbO expressing cells of M. bovis BCG, were inoculated into a number of (30–40) 100 mL culture flasks carrying 75 mL medium, and initial OD600 was normalized to 0.1 in each flask. Flasks were sealed and cultures were allowed to grow at 37°C with 75 rpm. Flasks, in triplicate, were taken out at different time intervals and OD600 was measured and simultaneously plated after appropriate dilution to determine CFU for checking the cell viability. In another set of experiments, cell cultures were inoculated into 20 mL medium in 25 mL screw-capped flat-bottomed culture tubes containing methylene blue (0.5 mg/mL) as an indicator of oxygen depletion, as described elsewhere (6). Tubes were sealed and subjected to slow rotation (75 rpm) at 37°C. Decolorization of methylene blue, as an indicator of oxygen consumption, was checked periodically by taking OD600.
Footnotes
Acknowledgments
The authors are thankful to Prof. Tanya Parish, Infectious Disease Research Institute, Seattle, Washington, for providing plasmid vectors, p2NIL, pGOAL18, and pGOAL19 and to Prof. Pradip K. Chakraborti for his support and helpful discussion. Support provided by NASI-Senior Scientist Fellowship to KLD by the National Academy of Sciences, India, and the research facility provided by CSIR-IMTECH and Panjab University are thankfully acknowledged. Fellowship/assistantship has been provided to M.D.H., D.S., H.D., N.T., and S.S. by ICMR, DST, and DBT, Govt. of India. These funders had no role in the study, data analysis, or preparation of this article.
Author Disclosure Statement
The authors declare that there are no commercial associations that might create a conflict of interest.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
Supplementary Table S1
Supplementary Table S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.