
Abstract
Aim:
Neutrophil infiltration and increased oxidative stress are involved in the pathogenesis and severity of psoriasis. Although the therapy of psoriasis remains elusive, targeting treatment to reduce oxidative stress is considered a potential option. Our study demonstrates the anti-inflammatory effects of a natural furocoumarin, imperatorin, on activated human neutrophils and psoriasiform dermatitis in mice.
Results:
Imperatorin inhibited superoxide anion generation, neutrophil adhesion, and migration in N-formyl-
Innovation and Conclusion:
Our results demonstrate that imperatorin inhibits human neutrophil respiratory burst, adhesion, and migration through the elevation of cAMP/PKA to inhibit Akt, ERK, JNK, and Ca2+ mobilization. Imperatorin is a natural inhibitor of PDE4A/B/C and may serve as a lead for developing new therapeutics to treat neutrophilic psoriasis. Antioxid. Redox Signal. 35, 885–903.
Introduction
Psoriasis is a common inflammatory cutaneous disorder that affects ∼2%–3% of the global population. This chronic disease manifests as erythematous plaques and papules with silver scaling, and is associated with many comorbidities (13, 37, 43). Sterile inflammation or pathogen infection triggers adhesion and migration of neutrophils to the inflamed site. Marked neutrophil infiltration and keratinocyte proliferation in the epidermis are dominant in psoriatic skin. Upregulation of oxidative stress, chemotaxis, and the adhesion capacity of neutrophils have been found in patients with psoriasis (17, 33, 36). The generation of superoxide anion and associated reactive oxygen species (ROS) from neutrophils contributes to a vicious cycle of increasing neutrophil activation, and is related to the pathogenic process and disease severity of psoriasis (15, 39).
Innovation
Using a phosphodiesterase (PDE)4 inhibitor to reduce oxidative stress is a useful strategy for psoriasis treatment. This study provides the evidence of this target by suppressing neutrophilic oxidative stress against this inflammatory cutaneous disorder. Our results demonstrate that imperatorin inhibits key inflammation reactions of activated neutrophils involved in psoriasis pathogenesis, and ameliorates imiquimod- and interleukin-23-induced mouse psoriasiform dermatitis disease. Imperatorin inhibits human neutrophil respiratory burst, adhesion, and migration through the elevation of cAMP/protein kinase A to inhibit protein kinase B (Akt), extracellular regulated kinase, c-Jun N-terminal kinase, and Ca2+ mobilization. Imperatorin is a natural inhibitor of PDE4A/B/C and may serve as a potent therapeutic to treat neutrophilic psoriasis. This study also suggests that imperatorin can act as a lead compound for developing selective PDE4A/B/C inhibitor.
The etiology and pathogenesis of psoriasis remain unclear. Although there is currently no cure for psoriasis, steroids and vitamin D3 analogs remain the gold-standard first-line therapy. However, in some patients and in those with severe psoriasis, corticosteroids and vitamin D3 analogs may be poorly tolerated, have low efficacy, or produce severe side effects with long-term use (38). Thus, it is important to develop agents with better safety and efficacy by targeting disease-defining pathogenic pathways. One drug newly developed using this strategy is a phosphodiesterase (PDE)4 inhibitor. Of all the PDE families, the PDE4 subfamily is the most common subtype in neutrophils and is involved in the pathogenesis of many neutrophilic inflammatory diseases (31, 57). PDE4 mainly hydrolyzes intracellular cAMP. The PDE4 inhibitor exerts anti-inflammatory effects by decreasing oxidative stress and CD11b integrin expression via increasing cAMP levels and protein kinase A (PKA) activity in neutrophils (24, 52, 54). Elevation of the cAMP level raises PKA activity, which in turn inhibits the activity of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and reduces the respiratory burst (59). Currently, the number of PDE4 inhibitors available for clinical application is limited. One promising agent is apremilast, which is currently approved for the treatment of psoriasis (22).
Imperatorin [9-(3-methylbut-2-enyloxy)-7H-furo[3,2-g]chromen-7-one] was isolated from Angelica hirsutiflora by our team (27). It was reported to possess a variety of pharmacological effects, including antihypertensive (6), antitumorigenic (9, 10, 28), and anti-inflammatory activities (18, 20, 35, 46), in published studies. Recent animal studies showed that imperatorin reduces neutrophil infiltration in lipopolysaccharide (LPS)-induced acute lung injury (46) and carrageenan-induced paw edema (20). However, the action mechanisms and therapeutic effects of imperatorin in neutrophils remain elusive.
We found imperatorin is a selective PDE4 inhibitor that shows suppressive effects on the production of superoxide anion and reactive oxidants, CD11b expression, neutrophilic adhesion, and migration in N-formyl-
Results
Imperatorin suppresses fMLF-stimulated superoxide anion release and associated ROS formation in human neutrophils
Respiratory burst and protease degranulation are two major components of neutrophilic inflammatory responses. To investigate the anti-inflammatory effects of imperatorin (Fig. 1A) on human neutrophils, we first tested whether imperatorin could reduce superoxide anion release and associated ROS formation in fMLF-treated neutrophils. The data showed that imperatorin (0.03–1 μM) inhibited superoxide anion production with a half-maximal inhibitory concentration (IC50) value of 0.17 ± 0.02 μM (Fig. 1B). Imperatorin also suppressed fMLF-induced total ROS formation in neutrophils (Fig. 1C).
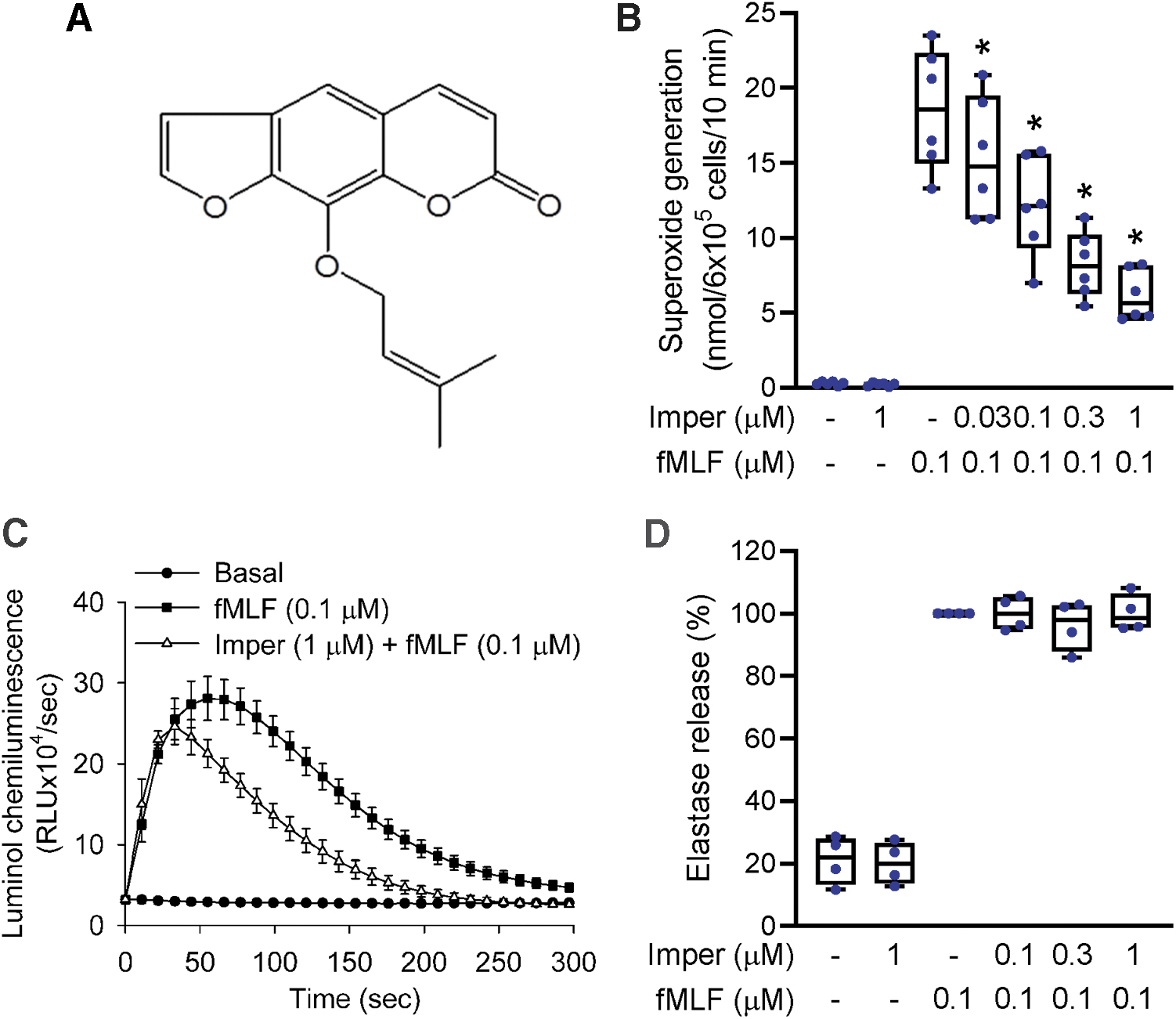
Protease degranulation of neutrophils was analyzed by release of elastase, the major granule-derived protease in neutrophils. Figure 1D shows that imperatorin failed to inhibit fMLF-stimulated elastase degranulation from neutrophils. Imperatorin and isoimperatorin are isomeric. We have also studied the effect of isoimperatorin in human neutrophils (Supplementary Fig. S1). Our results showed that imperatorin, compared with isoimperatorin, had much stronger inhibitory effects on superoxide anion release in fMLF-activated human neutrophils.
Imperatorin significantly inhibits fMLF-stimulated neutrophil adhesion and chemotaxis
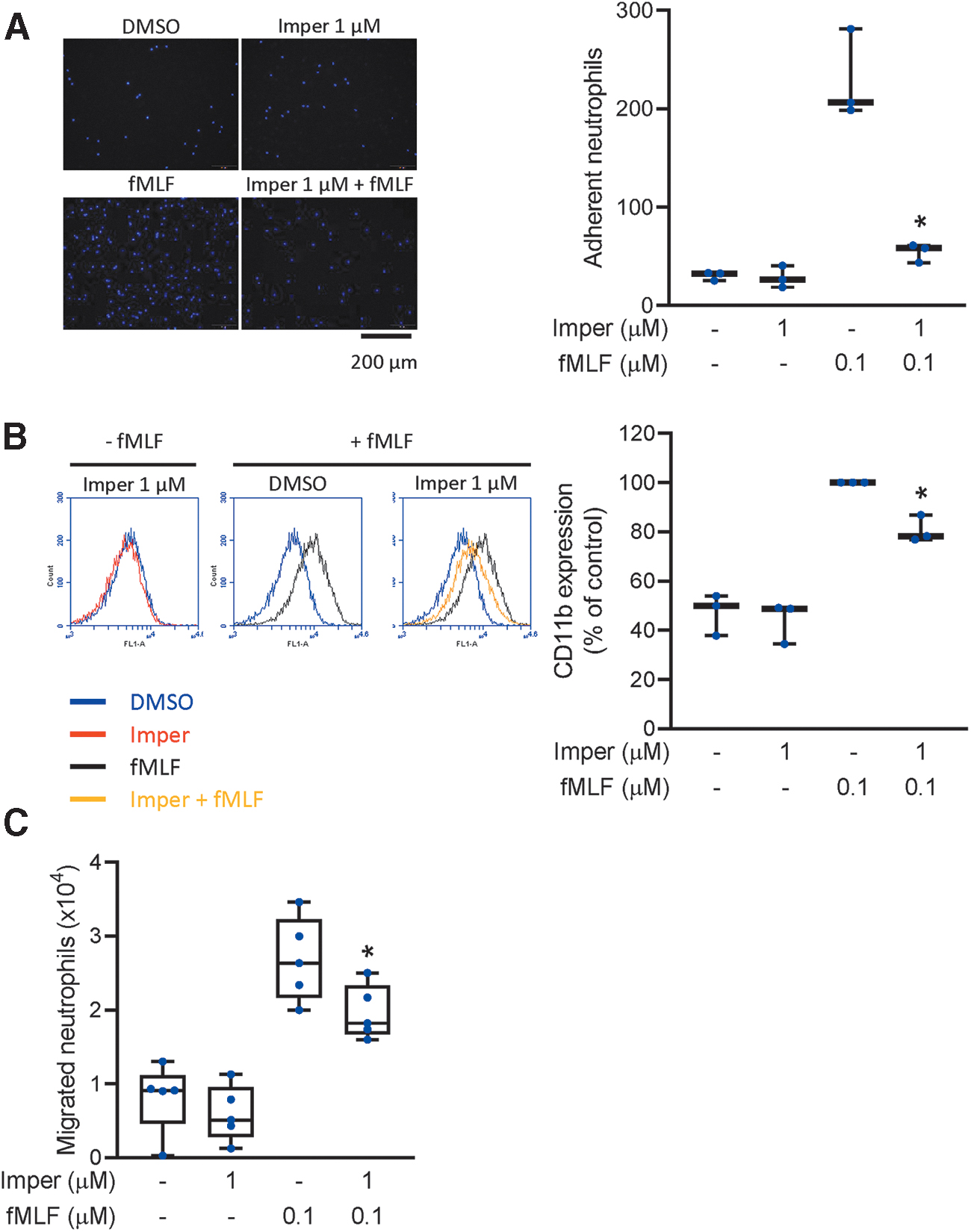
Neutrophil chemotaxis is a directional movement toward sites of inflammation. Adhesion molecules on circulating neutrophils are expressed during inflammation to facilitate neutrophil adhesion and migration. We evaluated the chemotactic responses of activated neutrophils using three different methods: the adherence of neutrophils to endothelial cells, expression of the adhesion molecule CD11b, and neutrophil migration. Imperatorin diminished the number of neutrophils adhered to the surface of endothelial cells (Fig. 2A). Imperatorin also significantly reduced fMLF-induced CD11b expression on neutrophils (Fig. 2B) and impaired the migration of fMLF-attracted neutrophils (Fig. 2C). All these data confirmed that imperatorin inhibited adhesion and chemotaxis in fMLF-stimulated neutrophils.
The inhibitory effects of imperatorin are not mediated through radical-scavenging effects, subcellular NADPH oxidase activity, cytotoxicity, or protein kinase C activation
The xanthine/xanthine oxidase system and a 1,1-diphenyl-2-picrylhydrazyl (DPPH)-scavenging assay were used to investigate whether the inhibitory effects of imperatorin on oxidative stress were directly mediated through its antioxidant activity. Our results showed that imperatorin did not alter the reduction of WST-1 and DPPH in cell-free systems (Supplementary Fig. S2A, B) where superoxide dismutase (SOD) and α-tocopherol were used as positive controls. Therefore, the suppressive effect of imperatorin on superoxide anion generation was not mediated directly by radical scavenging in human neutrophils.
NADPH oxidase is a multicomponent enzyme complex that is involved in the respiratory burst in neutrophils (48). To assay whether NADPH oxidase was involved in suppression of superoxide anion generation by imperatorin, neutrophil fractions were isolated to test their reconstituted NADPH oxidase activities. As shown in Supplementary Figure S2C, imperatorin had no effect on the generation of superoxide anion mediated through activated subcellular NADPH oxidase. Diphenyleneiodonium (10 μM), a NADPH oxidase inhibitor, was used as a positive control. This result supported that the inhibitory effects of imperatorin on respiratory burst are not mediated by directly blocking NADPH oxidase activity.
Neutrophil viability was examined by detection of lactate dehydrogenase (LDH) release. Imperatorin (0.3–30 μM) showed no toxicity to neutrophils when they were coincubated for 15 min (Supplementary Fig. S2D). Our results confirmed that none of the suppressive effects of imperatorin were due to direct cytotoxicity. In addition, imperatorin failed to reduce the generation of superoxide anion stimulated by phorbol myristate acetate (PMA) (Supplementary Fig. S2E). Ro318220 (1 μM), an antagonist of protein kinase C (PKC), was used as a positive control. These data suggested that PKC pathway did not mediate the inhibitory effects of imperatorin.
Imperatorin blocks cAMP-specific PDE activity to increase cAMP concentration and PKA activity in neutrophils
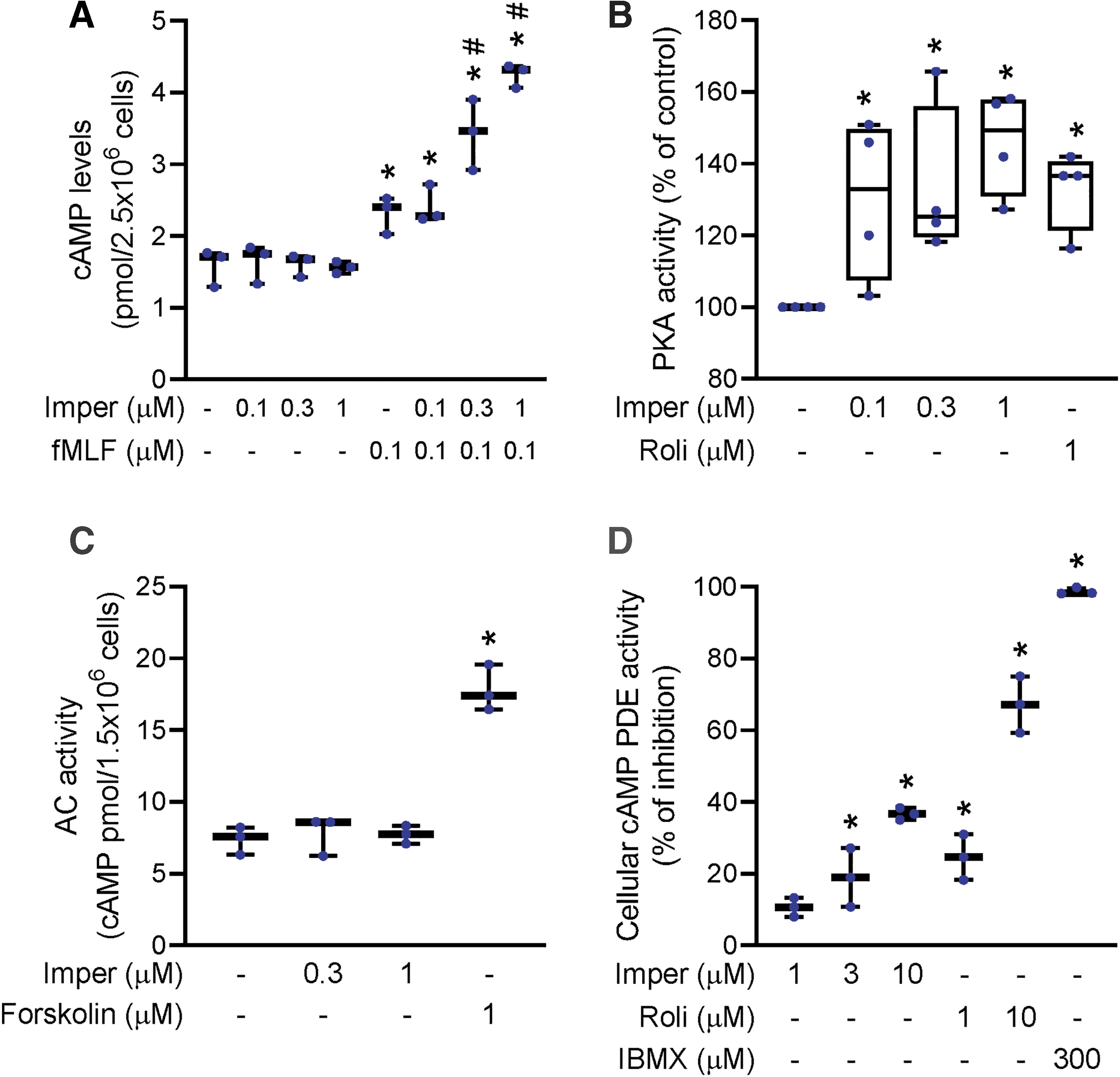
Increased levels of the second messenger cAMP mediate the cAMP/PKA pathway and inhibit inflammatory processes in human neutrophils (3, 52). We analyzed the effects of imperatorin on cAMP concentration and PKA activity in neutrophils, and demonstrated that imperatorin (0.1, 0.3, and 1 μM) induced a synergistic increase in cAMP concentration in fMLF-stimulated neutrophils (Fig. 3A). However, imperatorin had no effect on intracellular cAMP concentrations when it was coincubated alone with neutrophils. Imperatorin also increased PKA activity in fMLF-stimulated neutrophils (Fig. 3B). Intracellular cAMP is synthesized by adenylyl cyclases (ACs) and degraded by PDEs. Our results indicated that imperatorin had no effect on the activity of ACs in neutrophils (Fig. 3C). Significantly, imperatorin reduced cAMP-specific PDE activity (Fig. 3D). Rolipram (a PDE4 inhibitor), forskolin (an AC activator), and 3-isobutyl-1-methylxanthine (IBMX; a nonselective inhibitor of PDEs) were used as positive controls in the respective assays. Our data indicated that imperatorin-induced elevation of cAMP level and PKA activity in stimulated neutrophils was mediated via suppression of intracellular PDE activity.
Imperatorin selectively inhibits PDE4 activity
The subtypes of cAMP-specific PDEs in human neutrophils are PDE3, PDE4, and PDE7 (14). In this study, imperatorin selectively inhibited the activity of PDE4 subtypes, but not of PDE3 or PDE7 (Fig. 4). PDE4 subtypes include PDE4A, PDE4B, PDE4C, and PDE4D. In further PDE4 subtype analyses, imperatorin (1, 3, and 10 μM) significantly reduced the activities of PDE4A, PDE4B, and PDE4C (Fig. 4A–C), but had a weaker inhibitory effect on PDE4D (Fig. 4D). The IC50 values of imperatorin on PDE4A, PDE4B, and PDE4C are 6.28 ± 0.69, 5.80 ± 1.11, and 5.64 ± 0.16 μM, respectively. Rolipram (PDE4 inhibitor), cilostamide (PDE3 inhibitor), and BRL50481 (PDE7 inhibitor) were used as positive controls.

The inhibitory effects of imperatorin on neutrophil activation are mediated through PKA signaling
To confirm that the imperatorin-induced reduction in neutrophil functions including ROS formation, adhesion, and migration was mediated through PKA signaling, the PKA inhibitors, H89 and KT5720, were used. The suppression of superoxide anion formation by imperatorin in fMLF-stimulated neutrophils was reversed when cells were pretreated with H89 or KT5720 (Fig. 5A, B). These results suggest that imperatorin inhibition of superoxide anion production in neutrophils was mediated through the cAMP/PKA pathway. In addition, imperatorin-mediated inhibition of total ROS production in fMLF-stimulated neutrophils was blocked by H89 (Fig. 5C). Our data also showed that the inhibitory effects of imperatorin on neutrophilic adhesion and migration were markedly reduced by the pretreatment of PKA inhibitor (Fig. 5D, E). Rolipram, a PDE4 inhibitor, was used as positive control. Rolipram inhibited neutrophil superoxide anion release, total ROS production, adhesion, and migration. These inhibitory effects of rolipram were also reversed when neutrophils were pretreated with PKA inhibitor (Fig. 5A–E).
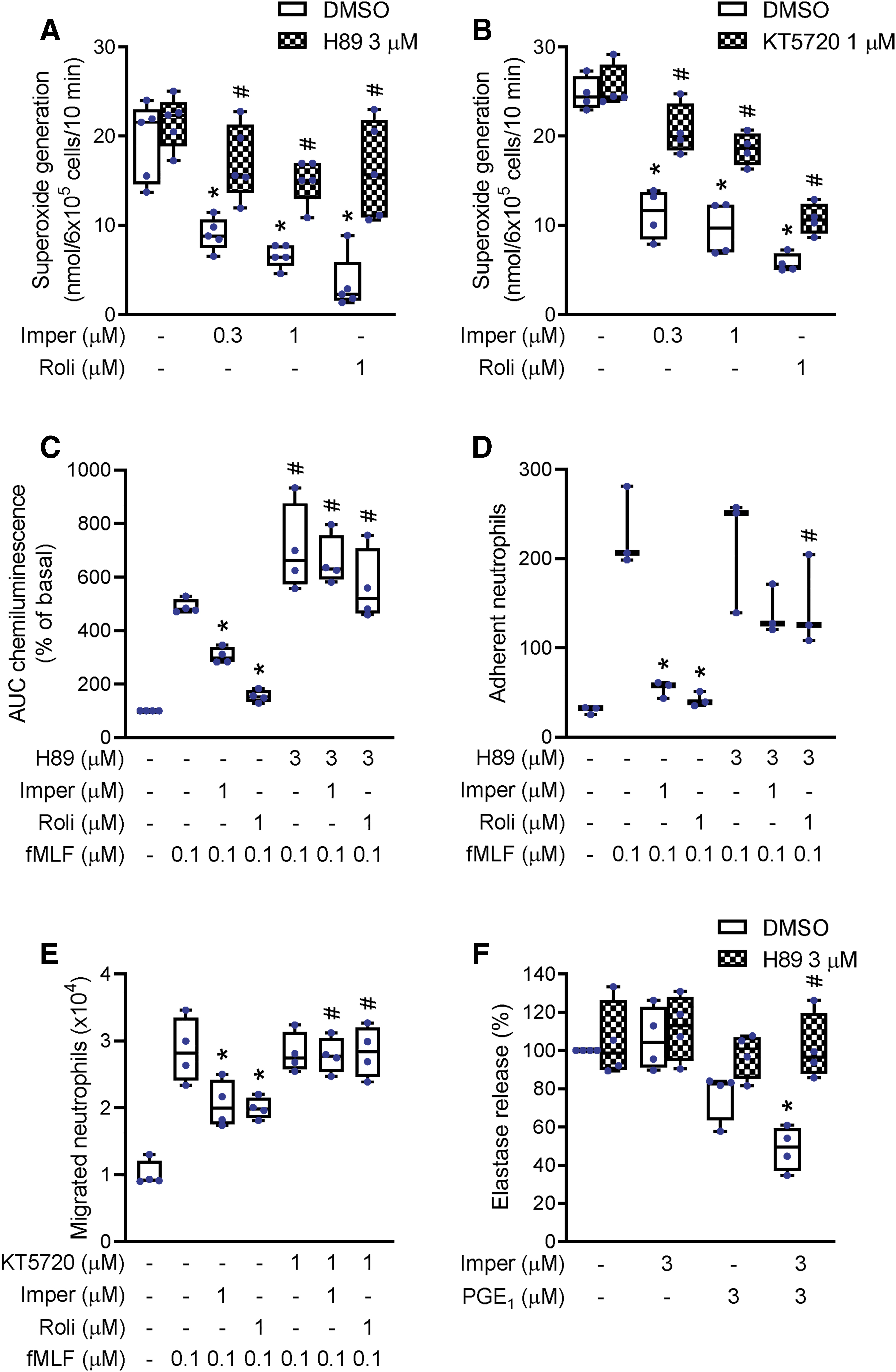
Prostaglandin (PG)E1 (3 μM) slightly reduced elastase release, but PGE1 combined with imperatorin showed synergistic effect, indicating that imperatorin enhanced PGE1-mediated effect on elastase release. This synergistic suppression of elastase release was also reversed by H89 (Fig. 5F). These results confirmed that the inhibitory effects of imperatorin on neutrophil functions are mediated through PKA signaling.
Imperatorin suppresses phosphorylation of protein kinase B, extracellular regulated kinase, and c-Jun N-terminal kinase, and reduces the duration of Ca2+ increase via PKA signaling
Our data showed a rapid phosphorylation of protein kinase B (Akt), extracellular regulated kinase (ERK), and c-Jun N-terminal kinase (JNK) in fMLF-stimulated neutrophils. Imperatorin significantly suppressed fMLF-induced phosphorylation of Akt473, Akt308, ERK, and JNK, and these inhibitory effects were abolished by H89 (Fig. 6). The intracellular Ca2+ level ([Ca2+]i) transiently and rapidly increases after fMLF activation. This response results in neutrophil activation via a G protein-coupled receptor. The modulation of [Ca2+]i can affect neutrophil activation (5). In fMLF-stimulated neutrophils, imperatorin reduced the t 1/2 (defined as the time taken to decline to half of its peak value) of the increase in [Ca2+]i, but was unable to reduce the peak [Ca2+]i. The inhibitory effect on t 1/2 was reversed by H89 (Fig. 7). These data suggest that the inhibitory effects of imperatorin on neutrophils are mediated through regulation of PKA activity to suppress Akt, ERK, JNK, and Ca2+ signals.

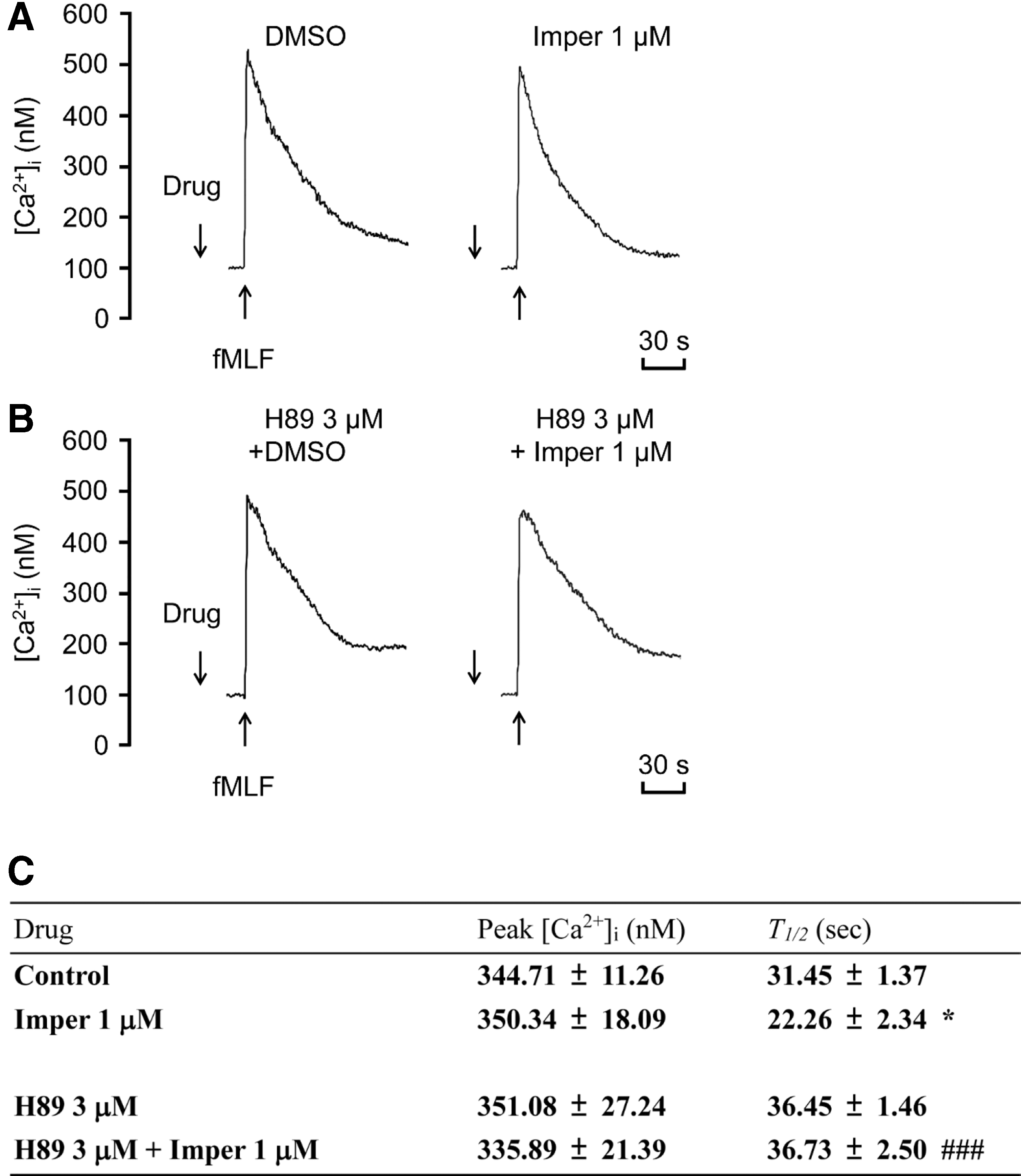
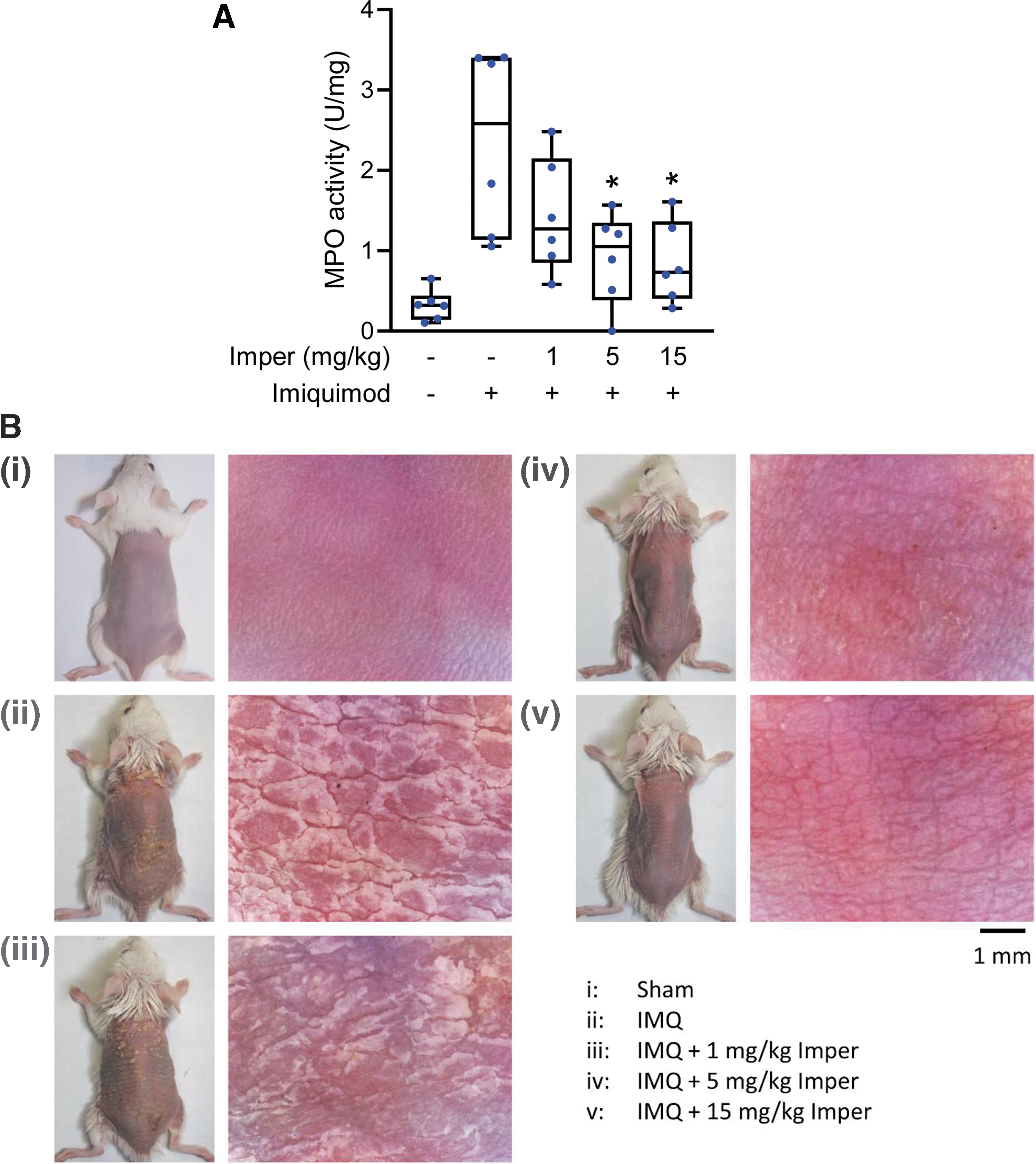
Imperatorin alleviates imiquimod-induced psoriasiform dermatitis in BALB/c and C57BL/6 mice
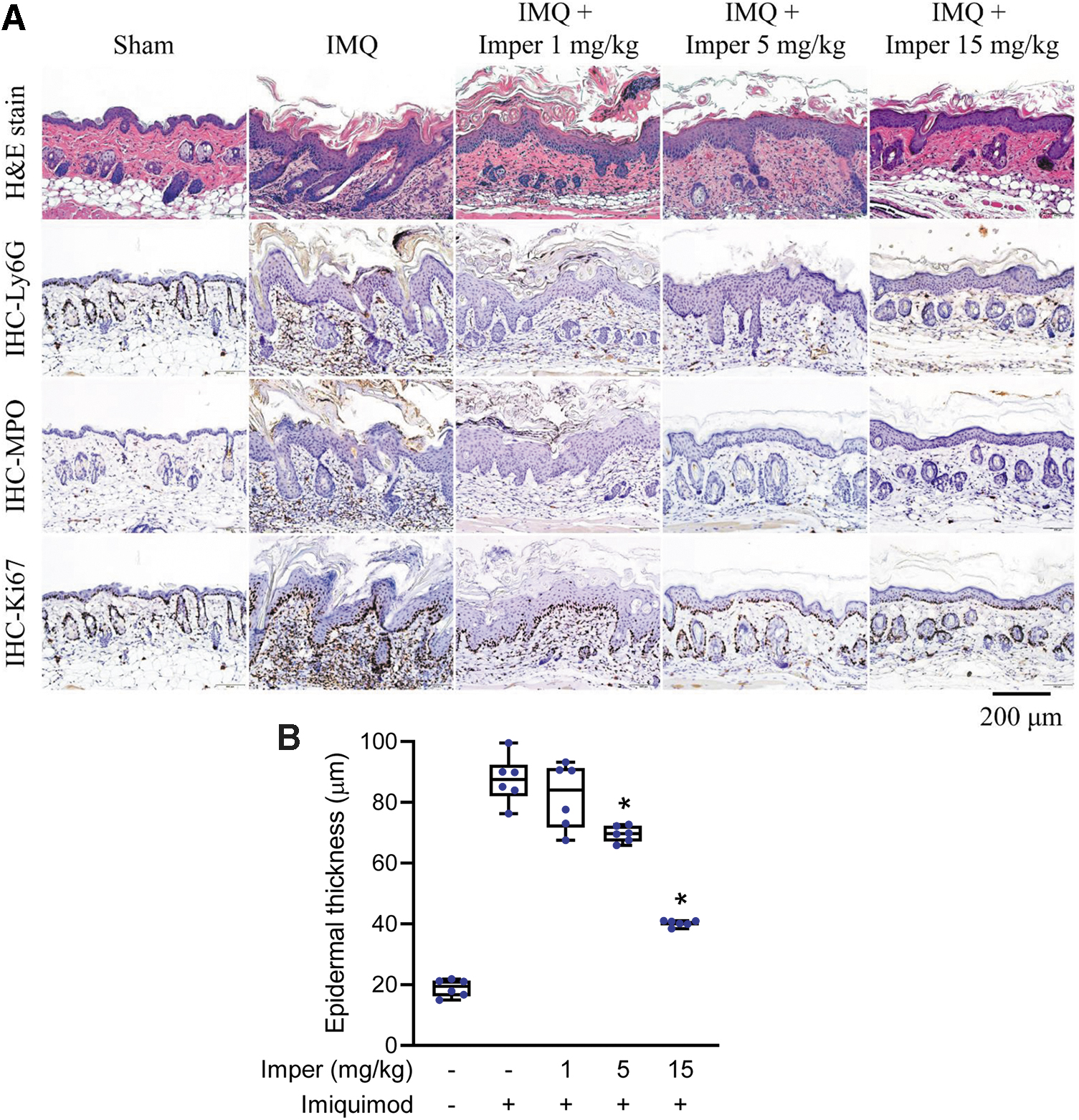
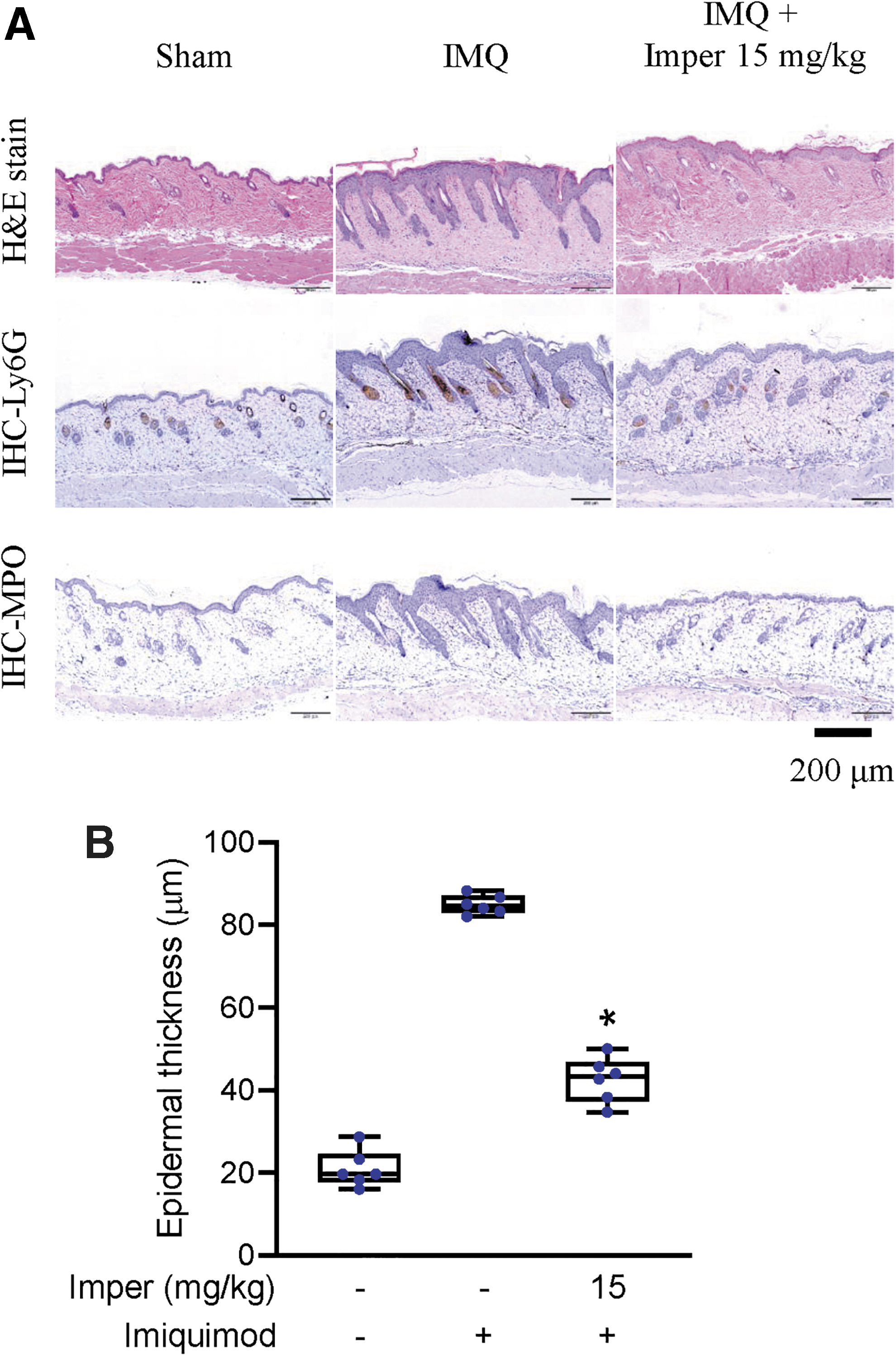
We evaluated the therapeutic effects of imperatorin in imiquimod-induced psoriasiform dermatitis in BALB/c and C57BL/6 mice. The therapeutic effects of imperatorin on psoriasiform dermatitis in mice were investigated in sham-operated mice treated with vehicle (50% dimethyl sulfoxide [DMSO] in Tween 80); imiquimod-induced psoriasiform dermatitis mice treated with vehicle; and imiquimod-induced psoriasiform dermatitis mice treated with imperatorin at doses of 1, 5, or 15 mg/kg. Imiquimod topically applied to the dorsal skin of BALB/c mice resulted in obvious scaling and redness. It also increased neutrophil infiltration into the skin, which was detected as increases in skin myeloperoxidase (MPO) activity, compared with that observed in sham-operated BALB/c mice (Fig. 8A). Imperatorin relieved the scaliness and redness in the inflamed skin area (Fig. 8B). Figures 9A and 10A show the histological analyses of skin from each group stained with hematoxylin–eosin (H&E) and after immunohistochemical (IHC) staining. Microscopically, imiquimod-treated skin showed features of significant desquamation and epidermal thickening compared with the sham-operated group. The increase of epidermal thickness and keratinocytes in the basal layer resulted in acanthosis and hyperproliferation in the imiquimod-treated groups in BALB/c and C57BL/6 mice (Figs. 9 and 10). The neutrophil infiltration and cell proliferation in the imiquimod-treated groups were greater than those in the sham-operated group based on the presence of Ly6G+, MPO+, and Ki67+ cells in the dermis layer. All these pathological features in BALB/c and C57BL/6 mice were significantly reduced by imperatorin treatment (Figs. 9 and 10 and Supplementary Figs. S3 and S4).
Imperatorin alleviates IL-23-induced psoriasiform dermatitis in C57BL/6 mice
The therapeutic effect of imperatorin on psoriasiform dermatitis was further investigated in IL-23-induced C57BL/6 mice. IL-23-treated skin showed features of desquamation, epidermal hyperplasia and acanthosis, and reduction in granulosa layer compared with the sham-operated group. The neutrophil infiltration in the IL-23-treated groups was greater than that in the sham-operated group. All these pathological features were significantly reduced by imperatorin treatment (Fig. 11 and Supplementary Fig. S5).

Imperatorin ameliorates LPS-induced acute lung injury
The therapeutic effects of imperatorin on acute respiratory distress syndrome (ARDS) were tested in LPS-induced acute lung injury model in C57BL/6 mice. LPS increased neutrophil infiltration into the lung, which was detected as increases in MPO activity (Fig. 12A). Histopathologic studies of H&E-stained lung section revealed that LPS induced severe lung injury as ARDS-like lung inflammation, including alveolar damage, edema, and hemorrhage as well as neutrophil infiltration (Ly6G+ and MPO+ cells) (Fig. 12B). Treatment of imperatorin (5 mg/kg) significantly attenuated MPO activity and neutrophil infiltration as well as improved the pathological changes (Fig. 12 and Supplementary Fig. S6).

Discussion
Psoriasis is an incurable inflammatory cutaneous disease that can disable patients and limit their quality of life. Innate immunologic reactions recognized as an initial trigger are central to the development of psoriatic skin lesions. Inflamed skin is infiltrated by activated neutrophils, macrophages, T cells, and dendritic cells (4). Neutrophils are effector cells of innate immunity that are activated in psoriasis to secrete proinflammatory mediators, ROS, proteases, and neutrophil extracellular traps (15, 44). Increased ROS generation and insufficient antioxidant activities are involved in the pathogenesis of psoriasis (17, 33, 36). Although the mechanisms of psoriasis pathogenesis remain unclear, targeting treatment to reduce oxidative stress is considered a potential option (25). In psoriatic lesions, there is massive infiltration of neutrophils that migrate through the endothelium into the epidermis. The peripheral blood neutrophil count and neutrophil-to-lymphocyte ratio are correlated with psoriasis severity, and seem to be strong predictors of the disease (41). The chemotactic and migratory responses of neutrophils are greatly enhanced by chemotactic factors, which therefore augment the inflammatory process (44). Increased oxidative stress, degranulation of proteases, and induction of proinflammatory mediators all contribute to the vicious cycle of inflammation (8, 15). Recently, the use of PDE4 inhibitors to target disease-defining pathogenic pathways and restrain neutrophil activation and its induced oxidative stress has shown promise as a therapy for psoriasis patients (38). Here, we demonstrate the antipsoriasis effects of imperatorin, a natural furanocoumarin PDE4 inhibitor, and clarify its signal transduction mechanisms in human neutrophils.
As neutrophils are activated, they mediate the respiratory burst and protease degranulation to regulate inflammatory reactions. During the respiratory burst reaction, neutrophils generate a considerable amount of ROS via NADPH oxidase activation. Dysregulated or overabundant oxidative stress in inflamed tissues is related to the pathogenesis of inflammatory diseases. This study demonstrated that imperatorin suppressed superoxide anion formation and total ROS production in fMLF-stimulated human neutrophils (Fig. 1B, C). Upon infection, neutrophils are recruited from the vasculature into inflamed tissue via adhesion and migration processes. Circulating peripheral blood neutrophils adhere to endothelium near the inflamed area and then migrate into the area. These reactions require the production of chemoattractants to attract and activate neutrophils. Imperatorin clearly inhibited the fMLF-stimulated adhesion of human neutrophils to endothelial cells and reduced the expression of adhesion molecule CD11b on activated neutrophils (Fig. 2A, B). The chemotaxis assay demonstrated that imperatorin suppressed chemotactic migration of fMLF-stimulated neutrophils (Fig. 2C). All the above data suggest that imperatorin blocks the increase in oxidative stress, adhesion, and chemotaxis in activated neutrophils. Therefore, imperatorin shows potential as an adjuvant therapy for psoriasis.
Imperatorin was unable to scavenge superoxide anion and DPPH radicals in cell-free systems (Supplementary Fig. S2A, B), indicating that imperatorin mediates its anti-inflammatory effects through regulating intracellular signaling pathways. NADPH oxidase in neutrophils catalyzes electron transfer from NADPH to molecular oxygen, thereby producing superoxide anion and increasing oxidative stress (34, 58). Imperatorin did not inhibit neutrophil NADPH oxidase activity (Supplementary Fig. S2C), indicating that imperatorin inhibits neutrophil activation by regulating upstream signaling pathways of NADPH oxidase.
The intracellular cAMP plays a negative-regulatory role in neutrophil activation (50). Our study showed that imperatorin increased the intracellular cAMP concentration and PKA activity in activated human neutrophils (Fig. 3A, B). These results suggest that the anti-inflammatory effects of imperatorin in neutrophils are through cAMP/PKA-dependent pathway. Consistent with this hypothesis, we demonstrate that the inhibitory effects of imperatorin on superoxide anion formation, total ROS production, adhesion, and migration were reversed by inhibiting PKA (Fig. 5). The concentration of cAMP is modulated by a balance of synthesis by ACs and degradation by PDEs. Our data showed that imperatorin attenuated the activities of cAMP-specific PDEs, but not that of ACs (Fig. 3D, C). Imperatorin has been identified as a potential PDE4 inhibitor by Ivey et al. in a fission yeast-based screening assay (23). Previous studies have reported that PDE3, PDE4, and PDE7 are the essential cAMP-specific PDE subtypes in neutrophils (14), and that PDE4 is the major subtype present. Our screening demonstrated that imperatorin selectively inhibited activities of PDE4 subtypes, but not that of PDE3, or PDE7 (Fig. 4). Furthermore, we showed that imperatorin decreased the activity of PDE4A, PDE4B, and PDE4C, but not PDE4D. A major concern with PDE4 inhibitors is the occurrence of adverse effects, such as nausea, emesis, and gastrointestinal reactions. Animal studies suggest that PDE4D activity in the area postrema of medulla possibly involves in emesis control (26, 32). The inhibition of PDE4D activity by PDE4 inhibitors in central nerve system can cause severe nausea or vomiting. Our study indicated that imperatorin showed more sensitive inhibitory effect on PDE4A, PDE4B, and PDE4C than PDE4D. Therefore, our data suggest that imperatorin may perform effective therapeutic activity with less adverse effects. We provide experimental evidence to suggest that imperatorin, a natural furocoumarin, can act as a lead compound to develop selective PDE4A/B/C inhibitor for treating neutrophil-dominant inflammatory diseases.
Imperatorin is a selective PDE4A/B/C inhibitor. PGEs increase the intracellular cAMP level via Gs-coupled receptors (11). Elevation of cellular cAMP levels by Gs activator, but not by PDE4 inhibitor, shows inhibitory effect on elastase release in activated neutrophils (54). Imperatorin did not inhibit elastase degranulation in activated neutrophils (Fig. 1D). Interestingly, imperatorin showed a synergistic effect on elastase release when combined with an AC activator, PGE1, and this synergistic effect was also blocked by a PKA inhibitor (Fig. 5F).
The phosphorylation of Akt and MAPKs as well as increase in [Ca2+]i involves in neutrophil activation (52 –54). Immunoblotting assays demonstrated that imperatorin inhibited fMLF-induced phosphorylation of Akt, ERK, and JNK, and that these effects were abolished by a PKA inhibitor (Fig. 6). The [Ca2+]i rapidly increased after treatment with fMLF in neutrophils. Imperatorin did not reduce the peak [Ca2+]i, but did reduce the t 1/2 in fMLF-stimulated neutrophils (Fig. 7). This effect on t 1/2 was also reversed by PKA inhibition. All these data led to the suggestion that the anti-inflammatory effects of imperatorin in neutrophils are mediated through inhibiting Akt, ERK, JNK, and Ca2+ by PDE4 inhibition.
Imperatorin has been reported to possess a variety of pharmacological effects, including anticancer, anti-inflammation, antivirus, antibacteria, antiangiogenesis, vascular relaxation, antiosteoporosis, and anticystic fibrosis (7). Imperatorin suppresses arachidonate metabolism by inhibiting cyclooxygenase and 5-lipoxygenase in stimulated mouse peritoneal macrophages (1). Imperatorin inhibits T-cell proliferation by blocking binding of nuclear factor of activated T cells to DNA (30). Imperatorin shows anti-inflammatory effects through inhibition of NF-kappaB pathway (61). Many furanocoumarins have been well established as photochemotherapy for inflammatory skin diseases (42). The efficacy of phototreatment with 8-methoxypsoralen bath has been confirmed in psoriatic patients (55). Furanocoumarins have multitargets, and their chemical structures can be further developed as drug candidates for several therapeutic fields (7). We cannot rule out the off-target effect of imperatorin in human neutrophils. Our study showed that imperatorin inhibits PDE4 activity, and increases intracellular cAMP level and PKA activity in human neutrophils. These suppressive effects of imperatorin on cell responses and signaling were reversed by PKA inhibitor, suggesting that PDE4 inhibition is involved in the anti-inflammatory effects of imperatorin.
The study of new therapeutic drugs for treating psoriasis is an important issue. The histological characteristics of human psoriasis include skin erythema, scaling, desquamation, increased epidermal thickness, and keratinocyte hyperproliferation, as well as the infiltration of innate immune cells (56). It is well known that the number of infiltrated neutrophils and the upregulation of oxidative stress are correlated with disease severity in psoriasis (34). However, the pathogenic factors of psoriasis are very complex, and no animal model of psoriasis can fully recapitulate the complex features of lesions in psoriasis patients (16). van der Fits et al. reported that imiquimod-induced dermatitis in BALB/c mice closely resembles human psoriasis lesions (56). Imiquimod-induced mouse psoriasiform dermatitis is a convenient and rapid animal model for studying psoriasis. We focused on evaluating the therapeutic effects of imperatorin on psoriatic lesional skin: skin desquamation, epidermal thickening, keratinocyte hyperproliferation, and neutrophil infiltration. Aldara cream (5% imiquimod) was used to induce psoriasiform dermatitis. Neutrophils are an important histological hallmark and play a critical pathogenic role in psoriasis (40). Neutrophil depletion has been reported to reduce the psoriasis phenotype in the imiquimod-induced psoriatic lesions in BALB/c mice (19). Sumida et al. reported that depletion of neutrophils ameliorated the disease severity of imiquimod-induced mouse psoriasis (45). Therefore, drugs targeting neutrophils are potential therapeutic strategy for treating psoriasis. In imiquimod-induced psoriasiform dermatitis mice, imperatorin reduced the MPO activity (Fig. 8A), relieved the scaliness and redness in the inflamed skin area (Fig. 8B). Histological and IHC examinations demonstrated that imperatorin significantly alleviated the desquamation, the increase in epidermal thickness, keratinocyte hyperproliferation, and neutrophil infiltration (Fig. 9 and Supplementary Fig. S3). Our results show that imperatorin is a potential adjuvant for psoriasis therapy.
Previous study demonstrated that imiquimod can induce different types of dermatitis in different mouse strains (47). In BALB/C mice, imiquimod seems to more replicate an infection phenotype but not a psoriasis phenotype. Nevertheless, imiquimod more closely replicates a psoriatic dermatitis in C57BL/6 mice. Therefore, we tested the therapeutic effect of imperatorin in imiquimod-induced psoriasiform dermatitis in C57BL/6 mice. Our results showed that imperatorin inhibited imiquimod-induced increases in desquamation, epidermal thickness, keratinocyte hyperproliferation, and neutrophil infiltration in C57BL/6 mice (Fig. 10 and Supplementary Fig. S4). IL-23-induced psoriasiform dermatitis is a well-recognized psoriasis animal model (12). IL-23 intradermal injection induced neutrophil infiltration, epidermal hyperplasia and acanthosis, and reduction in granulosa layer in C57BL/6 mice. Our study revealed that imperatorin efficiently reduced IL-23-induced psoriasiform dermatitis (Fig. 11 and Supplementary Fig. S5). We further checked the therapeutic effects of imperatorin on ARDS, a neutrophil-dominant disease (60). As shown in Figure 12, LPS induced lung edematous changes and neutrophil infiltration, as shown by staining with H&E, Ly6G, and MPO, as well as MPO activity. Imperatorin showed decreases in lung neutrophil infiltration and edematous changes (Fig. 12 and Supplementary Fig. S6). Our results confirm that imperatorin has anti-inflammatory effects in human neutrophils and displays therapeutic potential to treat neutrophil-associated inflammatory diseases.
We identify that imperatorin is a selective inhibitor of PDE4A/B/C subtypes. To our knowledge, this is the first study to demonstrate the signaling pathways mediated by imperatorin in human neutrophils and to prove the therapeutic effects of imperatorin on psoriasis. The anti-inflammatory effects of imperatorin are mediated by reducing PDE4 activity to enhance the cAMP/PKA-dependent suppression of Akt, ERK, and JNK activation and Ca2+ mobilization (Fig. 13). Psoriasis remains an incurable and relapsing disease that is associated with many comorbidities. This study shows that imperatorin, a natural compound, shows promise as an alternative treatment for psoriasis.

Materials and Methods
Reagents
Imperatorin (Fig. 1A) was isolated from A. hirsutiflora by our team with a purity >97% (27). Aldara cream (5% imiquimod) was purchased from 3M Health Care (Leicestershire, United Kingdom). Dextran and Ficoll-Paque were purchased from GE Healthcare (BioTech, Stockholm, Sweden). Hank's balanced salt solution (HBSS) was purchased from Gibco (NY). PGE1, rolipram, and N-(2-((p-bromocinnamyl)amino)ethyl)-5-isoquinolinesulfonamide (H89), and KT5720 were obtained from Calbiochem (CA). Fluo-3 acetomethoxy (Fluo-3/AM) ester was purchased from Invitrogen (OR). The PDE subtype enzymes were purchased from BPS Bioscience (CA). The tetrazolium salt WST-1 was obtained from Dojindo Laboratories (Kumamoto, Japan). The information of antibodies is summarized in Supplementary Table S1. The other reagents were purchased from Sigma (MO).
Neutrophil preparation
This study was approved by the ethics committee of Chang Gung Memorial Hospital, and the study design followed the principles of the Declaration of Helsinki 1975. Blood samples were drawn from healthy donors aged 20–30 years after obtaining written informed consent. Neutrophils were purified by standardized methods, that is, dextran sedimentation, gradient centrifugation in Ficoll-Hypaque solution, and the hypotonic lysis of erythrocytes (52). The purity of isolated neutrophils was determined by CD66b staining and analyzed by flow cytometry. The purity of isolated neutrophils was ≥92.91%. Purified neutrophils that contained >98% viable cells were resuspended in Ca2+-free HBSS solution at 4°C before use.
Measurement of extracellular superoxide anion release
Oxidative stress is related to the pathogenesis of many inflammatory diseases. The assay of superoxide anion formation was based on the SOD-inhibitable reduction of ferricytochrome c (2). After administration of ferricytochrome c (0.5 mg/mL), neutrophils (6 × 105 or 3 × 105 cells/mL) were treated with vehicle (0.1% DMSO) or imperatorin at 37°C for 5 min. Neutrophils were then activated with 0.1 μM fMLF or 5 nM PMA for another 5 or 10 min. Neutrophils were treated with cytochalasin B (CB, 1 μg/mL) for 3 min before fMLF stimulation. Superoxide anion release was analyzed continuously using a spectrophotometer (U-3010; Hitachi, Tokyo, Japan) to measure the changes in absorbance at 550 nm.
Determination of ROS production
ROS levels were determined by a luminol–peroxidase system. Neutrophils (3 × 105) were pretreated with luminol (37.5 μM) and horseradish peroxidase (6 U/mL) at 37°C for 5 min, and then treated with vehicle or imperatorin for 5 min. Next, neutrophils were stimulated by fMLF (0.1 μM)/CB (1 μg/mL) for another 5 min. Chemiluminescence signals were monitored using a chemiluminometer (Fluostar Optima; BMG Labtech, Offenburg, Germany).
Determination of elastase degranulation
Neutrophil degranulation was evaluated by elastase release. After administration of elastase substrate, methoxysuccinyl-Ala-Ala-Pro-Val-p-nitroanilide (100 μM), neutrophils (6 × 105/mL) were treated with vehicle or imperatorin at 37°C for 5 min. Elastase release was induced by the addition of fMLF (0.1 μM)/CB (0.5 μg/mL), and changes in absorbance at 405 nm were continuously monitored. The data are normalized as a percentage of the elastase release in the control group.
Neutrophil adhesion study
Hoechst 33342 (1 ng/mL)-stained human neutrophils were incubated with vehicle or imperatorin (0.3 and 1 μM) for 5 min before the addition of fMLF (0.1 μM)/CB (1 μg/mL) for another 15 min. bEnd.3 endothelial cells were treated with LPS (2 μg/mL) for 4 h. After washing, LPS-pretreated endothelial cells were coincubated with the labeled neutrophils (1 × 105 cells/mL) for 15 min. Neutrophils adhering to endothelial cells were enumerated using a motorized inverted microscope (IX81; Olympus, Japan). Six random areas (0.572 mm2) were selected for cell number counting with a 10 × objective.
Detection of CD11b expression
The adhesion molecule CD11b is a member of the integrin family. Human neutrophils (2.5 × 106 cells/mL) were incubated with vehicle or imperatorin at 37°C for 5 min, and then treated with fMLF (0.1 μM)/CB (0.5 μg/mL) for 5 min. After activation, cells were cooled and centrifuged at 4°C. Cells were resuspended in HBSS (80 μL), and then labeled with fluorescein isothiocyanate-conjugated anti-CD11b antibody (1.5 μg) for 90 min on ice. The immunofluorescent CD11b was detected using flow cytometry.
Neutrophil migration study
A microchemotaxis chamber (Millipore, Darmstadt, Germany) with 3-μm pore size was used to analyze neutrophil migration. Human neutrophils (2.5 × 106 cells/mL) were preincubated with vehicle or imperatorin for 5 min at 37°C in the top chamber, and fMLF (0.1 μM) as chemoattractant was added to the bottom chamber. The number of migrated cells was calculated using a Coulter counter (Beckman Coulter, Inc., CA).
Assay of superoxide anion scavenging
The ability of imperatorin to scavenge superoxide anion was assayed using a cell-free xanthine/xanthine oxidase system (49). Xanthine oxidase (0.02 U/mL) was incubated with DMSO or imperatorin in assay buffer (50 mM Tris, 0.3 mM WST-1) for 3 min at 37°C, and then xanthine (0.1 mM) was added for 10 min. The absorbance change induced by superoxide anion-induced WST-1 reduction was analyzed at 450 nm.
DPPH-scavenging study
An ethanol solution of DPPH (100 μM) was treated with imperatorin, α-tocopherol, or DMSO for 15 min at 25°C. The absorbance changes were analyzed at 517 nM.
Determination of NADPH oxidase activity
After incubation with phenylmethylsulfonyl fluoride (PMSF, 1 mM) at 4°C for 30 min, neutrophils were lysed by sonication in relaxation buffer (3 mM NaCl, 100 mM KCl, 3.5 mM MgCl2, 10 mM piperazine-N,N′-bis(2-ethanesulfonic acid), 1 mM ATP(Na)2, and 1 mM Ethyleneglycol-bis(β-aminoethyl)-N,N,N′,N′-tetraacetic Acid (EGTA)). Samples were centrifuged for 5 min at 300 g to remove unbroken cells. The cytosol and membrane fractions were separated from the supernatant of samples by centrifugation at 100,000 g for 30 min. The two fractions were mixed with ferricytochrome c (0.5 mg/mL) and GTPγS (2 μM) in Pi2M buffer (138 mM NaCl, 2.7 mM KCl, 2 mM MgCl2, and 10 mM NaH2PO4/Na2HPO4 · 12H2O) at 30°C. After sodium dodecyl sulfate (SDS, 100 μM) was added to the mixture for 2 min, imperatorin was added for 1 min before treatment with NADPH (0.16 mM) for another 10 min. NADPH oxidase activity was represented by the change in absorbance at 550 nm.
Cytotoxicity assay
LDH release was assayed using a commercially available kit (Promega, WI). LDH release represented the cytotoxicity of the tested reagent in a cell-free medium at 37°C. Data were compared with the total LDH level that was obtained by lysing the neutrophils with Triton X-100 (0.1%).
Assay of AC
Human neutrophils (5 × 107 cells/mL) were lysed by sonication in lysis buffer (25 mM Tris-HCl, 100 μM PMSF, 5 mM MgCl2, 0.25 M sucrose, 2 mM ethylenediaminetetraaceticacid, 10 μM pepstatin, and 10 μM leupeptin) at 4°C. After removal of the nonlysed neutrophils by centrifugation, the supernatant was further centrifuged at 100,000 g for 40 min at 4°C. The pellet fraction was used to measure AC activity. cAMP levels were detected using a commercial enzyme immunoassay kit as described below.
Determination of cAMP concentrations
cAMP levels were detected using a commercial enzyme immunoassay kit (Amersham Biosciences, Buckinghamshire, United Kingdom). Human neutrophils were incubated with imperatorin or positive controls for 5 min before activation with or without fMLF for another 1 min. Dodecyltrimethylammonium bromide (0.5%) was added to stop the reaction, and the mixtures were then centrifuged at 3000 g for 5 min at 4°C. The cAMP concentrations of the supernatants were assayed (52).
Assay of PKA activity
A nonradioactive protein kinase assay was used to detect intracellular PKA activity (Biosource, Camarillo, CA). Imperatorin or rolipram was administered to human neutrophils (2 × 107/mL) at 37°C for 5 min before fMLF was added for another 1 min. The reaction was terminated by cooling to 4°C. The supernatants were removed after samples were centrifuged at 4°C, and cell pellets were immersed in Omnia cell extraction buffer for cell lysis. After being sonicated and centrifuged at 500 g for 10 min at 4°C, supernatants were analyzed for PKA activity.
Analysis of PDE activities
Human neutrophils were sonicated and lysed in lysis buffer, and then the cytosolic fraction was prepared by centrifugation at 1400 g for 15 min.
To measure the activities of cellular PDEs or standard PDEs (PDE3B, PDE4A1A, PDE4B2, PDE4C1, PDE4D2, or PDE7A), imperatorin or positive controls were mixed with the cytosolic fraction or PDE enzymes for 10 min. cAMP (60 nM) was added to the active reaction for 45 min, and then a fluorescence donor (anti-cAMP antibody) and fluorescence acceptor (Dye2-labeled cAMP) (Cysbio International, Bagnol-sur-Ceze, France) were coincubated for 60 min. The residual cAMP concentration was detected by fluorescence change using a microplate reader (Infinite® 200 Pro; Tecan).
Immunoblotting analysis
Human neutrophils were treated with DMSO or imperatorin for 5 min before stimulation with or without fMLF (0.1 μM) for 30 s at 37°C. Cells were incubated with H89 (3 μM) for 5 min before the addition of imperatorin. After being centrifuged at 4°C, the cell pellets were lysed in 150 μL lysis buffer (pH 7.4, 20 mM Tris-HCl, 150 mM NaCl, 1 mM NaF, 2 mM Na3VO4, 1 mM PMSF, 1% dilution of Sigma protease inhibitor cocktails, 1 mM EGTA, and 1% Triton X-100). Samples were prepared by centrifugation at 14,000 g for 20 min at 4°C. The proteins in the samples were separated by 12% SDS-polyacrylamide gel electrophoresis. The blots were transferred to nitrocellulose membranes (Whatman; Perkin-Elmer Life Science) and blocked with 5% skim milk in Tris-buffered saline containing 0.1% Tween 20. The immunoblots were then incubated, and identified with targeted antibodies and horseradish peroxidase-conjugated secondary antibodies (Cell Signaling). The following rabbit polyclonal antibodies were used: phospho-Akt473 (1:1000 dilution), phospho-Akt308 (1:1000 dilution), phospho-ERK (1:3000 dilution), phospho-JNK (1:1000 dilution), Akt (1:1000 dilution), ERK (1:3000 dilution), and JNK (1:1000 dilution) (Supplementary Table S1). A UVP Biospectrum imaging system (UVP LLC, CA) was used to determine the intensities of bands by detecting peroxidase activity with an enhanced chemiluminescence kit (Perkin-Elmer Life Science, MA). Uncropped Western blotting images are shown in Supplementary Figure S7.
Determination of calcium concentration ([Ca2+]i)
The increase of the intracellular calcium concentration ([Ca2+]i) was stimulated by adding fMLF to Fluo-3 AM (2 μM)-labeled human neutrophils. The [Ca2+]i was determined by monitoring the fluorescence change (excitation/emission wavelengths, 488/520 nm) using a spectrofluorometer (Hitachi F 4500; Tokyo, Japan) (21).
Imiquimod-induced psoriasiform dermatitis models in BALB/c and C57BL/6 mice
Study protocols were conducted in accordance with the approval of the Institutional Animal Care and Use Committee of Chang Gung Memorial Hospital. Seven-week-old female BALB/c and male C57BL/6 mice (CD IGS; BioLasco, Ilan, Taiwan) were rested in a temperature-controlled environment for 1 week before the animal study.
The procedures for inducing imiquimod-induced psoriasiform dermatitis were performed as previously reported (29). The back of mice was shaved using depilatory machine and cream on day 1. From the next day, imiquimod (62.5 mg/day) was topically applied to the bared dorsal skin for the next five consecutive days. Imperatorin (1, 5, or 15 mg/kg) or vehicle (40 μL; 50% DMSO and 50% Tween 80) was administered daily via the tail vein from days 3 to 6. On day 7, mice were humanely euthanized, and the back skin was removed for further studies. There are six mice in each group (n = 6 independent animals).
IL-23-induced psoriasiform dermatitis model in C57BL/6 mice
Seven-week-old male C57BL/6 mice were used for the animal study. The procedure for inducing IL-23-induced psoriasiform dermatitis was performed as previously reported (12). The back of mice was shaved using depilatory machine and cream on day 1. Skin inflammation was developed in mice by intradermal injection of recombinant mouse IL-23 (500 ng; BioLegend, CA) in every 2-day interval to the bared dorsal skin for consecutive 12 days. Imperatorin (15 mg/kg) or vehicle (40 μL; 50% DMSO and 50% Tween 80) was administered via intraperitoneal injection in the same day of IL-23 injection. On day 14, mice were humanely euthanized, and the back skin was removed for further studies. There are six mice in each group (n = 6 independent animals).
LPS-induced ARDS mouse model
This animal study used 7-week-old male C57BL/6 mice. All animals were randomly assigned to four groups: sham-operated mice treated with vehicle or imperatorin (5 mg/kg), ARDS animals treated with vehicle or imperatorin (5 mg/kg) intravenously. ARDS was induced by instilling LPS (2 mg/kg, Escherichia coli serotype 0111:B4) into mouse trachea after treatment of imperatorin for 1 h. Mice were sacrificed after LPS induction for 6 h. There are six mice in each group (n = 6 independent animals) (51).
Measurement of MPO activity in mouse skin
The MPO activity of skin or lung samples was measured as previously described (52, 54). Skin or lung samples of ∼100 mg were soaked in pH 6.0 buffer (0.5% hexadecyltrimethylammonium bromide in a 50 mM phosphate buffer), and then homogenized by sonication. After centrifugation at 2000 g at 4°C, the supernatants of skin or lung homogenates were used for analysis. The supernatants were mixed with hydrogen peroxide and o-dianisidine dihydrochloride in phosphate buffer for 5 min. The MPO activity of samples was determined by analyzing absorbance changes at 460 nm. MPO activity was expressed as units per gram of skin or lung tissue, and the values were referenced to a standard curve from commercial MPO (Sigma-Aldrich).
Histological examination
The skin or lung samples were immersed in 10% formalin for 1 day, embedded in 100% paraffin wax, and then sectioned at 5 μm thickness. The sliced sections were stained with H&E, and the tissue morphology was viewed under a microscope (Eclipse TS100; Nikon, Tokyo, Japan). For IHC stains, the protocol was as previously reported (52). Ly6G (Gr-1) and MPO are expressed in neutrophils, and thus were used to identify neutrophils. Ki67 antigen is a cell-cycle-related nuclear protein that is clearly expressed by proliferating cells. It is used to evaluate cellular proliferation. Ly6G, MPO, and Ki67 were immunohistochemically detected on paraffin-embedded sections using rat monoclonal antimouse-Ly6G (1:200 dilution), rabbit polyclonal antimouse-MPO (1:50 dilution), and rabbit monoclonal antimouse-Ki67 (1:200 dilution) primary antibodies (Supplementary Table S1). Primary antibodies were incubated with tissue sections for 1 h, followed by a corresponding polyclonal horseradish peroxidase-conjugated antirat immunoglobulin G antibody. The immune complex was visualized using the chromogen diaminobenzidine. IHC sections were counterstained with hematoxylin and mounted. The morphology was analyzed under a microscope (Eclipse TS100; Nikon). The pathological features of H&E and IHC sections were quantitated using ImageJ.
Statistical analysis
Data are reported as means ± standard error of the mean (SEM). N values are independent experiments. Electronic laboratory notebook was not used. All data are presented as box-and-whiskers plots (median, min–max) or line plots (mean, SEM). Statistical analysis was performed using one-way or two-way (for Figs. 5–7) analysis of variance followed by Tukey's multiple comparison test. A p value <0.05 was regarded as indicating significance.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported by the grants from the Ministry of Science Technology (MOST 106-2320-B-255-003-MY3, MOST 104-2320-B-255-004-MY3, MOST 106-2320-B-182A-002-MY2, and MOST 107-2320-B-182A-003), Ministry of Education (EMRPD1G0231 and EMRPD1H0381), Chang Gung Memorial Hospital (CMRPF1F0011–3, CMRPF1F0061–3, CMRPF1G0241–3, CMRPG3G1201, and BMRP450), and Chang Gung University of Science and Technology (ZRRPF3H0101 and ZRRPF3H0111), Taiwan. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.