
Abstract
Significance:
Mycobacterium tuberculosis (Mtb) encounters reductive stress during its infection cycle. Notably, host-generated protective responses, such as acidic pH inside phagosomes and lysosomes, exposure to glutathione in alveolar hypophase (i.e., a thin liquid lining consisting of surfactant and proteins in the alveolus), and hypoxic environments inside granulomas are associated with the accumulation of reduced cofactors, such as nicotinamide adenine dinucleotide (reduced form), nicotinamide adenine dinucleotide phosphate, flavin adenine dinucleotide (reduced form), and nonprotein thiols (e.g., mycothiol), leading to reductive stress in Mtb cells. Dissipation of this reductive stress is important for survival of the bacterium. If reductive stress is not dissipated, it leads to generation of reactive oxygen species, which may be fatal for the cells.
Recent Advances:
This review focuses on mechanisms utilized by mycobacteria to sense and respond to reductive stress. Importantly, exposure of Mtb cells to reductive stress leads to growth inhibition, altered metabolism, modulation of virulence, and drug tolerance. Mtb is equipped with thiol buffering systems of mycothiol and ergothioneine to protect itself from various redox stresses. These systems are complemented by thioredoxin and thioredoxin reductase (TR) systems for maintaining cellular redox homeostasis. A diverse array of sensors is used by Mycobacterium for monitoring its intracellular redox status. Upon sensing reductive stress, Mtb uses a flexible and robust metabolic system for its dissipation. Branched electron transport chain allows Mycobacterium to function with different terminal electron acceptors and modulate proton motive force to fulfill energy requirements under diverse scenarios. Interestingly, Mtb utilizes variations in the tricarboxylic cycle and a number of dehydrogenases to dissipate reductive stress. Upon prolonged exposure to reductive stress, Mtb utilizes biosynthesis of storage and virulence lipids as a dissipative mechanism.
Critical Issues:
The mechanisms utilized by Mycobacterium for sensing and tackling reductive stress are not well characterized.
Future Directions:
The precise role of thiol buffering and TR systems in neutralizing reductive stress is not well defined. Genetic systems that respond to metabolic reductive stress and thiol reductive stress need to be mapped. Genetic screens could aid in identification of such systems. Given that management of reductive stress is critical for both actively replicating and persister mycobacteria, an improved understanding of the mechanisms used by mycobacteria for dissipation of reductive stress may lead to identification of vulnerable choke points that could be targeted for killing Mtb in vivo.
Introduction
Redox reactions constitute an integral part of life. Living cells harvest reductive energy stored in different chemical compounds during catabolism and utilize it for synthesis of biomolecules. A delicate balance exists in living systems to ensure a conducive environment for carrying out essential redox reactions. Cells need to maintain a reducing environment to avoid oxidation of cysteine in proteins. Furthermore, oxidative stress can lead to protein misfolding and damage the genetic material subsequently leading to cell death (11). This has warranted extensive examination of oxidative stress (11). However, the effect of reductive stress on cell physiology remains comparatively unexplored. Reductive stress is defined as an increase in the reduced equivalents (of a redox couple) in cells (60, 101). This includes an increase in the cellular glutathione (GSH) (reduced)/glutathione disulfide (oxidized glutathione) ratio, nicotinamide adenine dinucleotide (reduced)/nicotinamide adenine dinucleotide (oxidized) ratio (NADH:NAD+), flavin adenine dinucleotide (reduced)/flavin adenine dinucleotide (oxidized) ratio (FADH2:FAD), and other redox couples (101). The concept of reductive stress was introduced through an elegant study, which demonstrated that accumulation of GSH and nicotinamide adenine dinucleotide phosphate (NADPH) is associated with intracellular protein aggregation, and is related to cardiomyopathy in mice (38, 101). Several studies have analyzed the effect of reductive stress on cell physiology and have established that increased reducing equivalents are associated with impaired cell growth (136), intracellular protein aggregation (101), unfolded protein response (34), inhibition of protein synthesis (103), altered cellular metabolism (61), increased reactive oxygen species (ROS) generation (53), and even cell death (156). Therefore, it is essential for cells to maintain redox homeostasis to avoid oxidative or reductive damage.
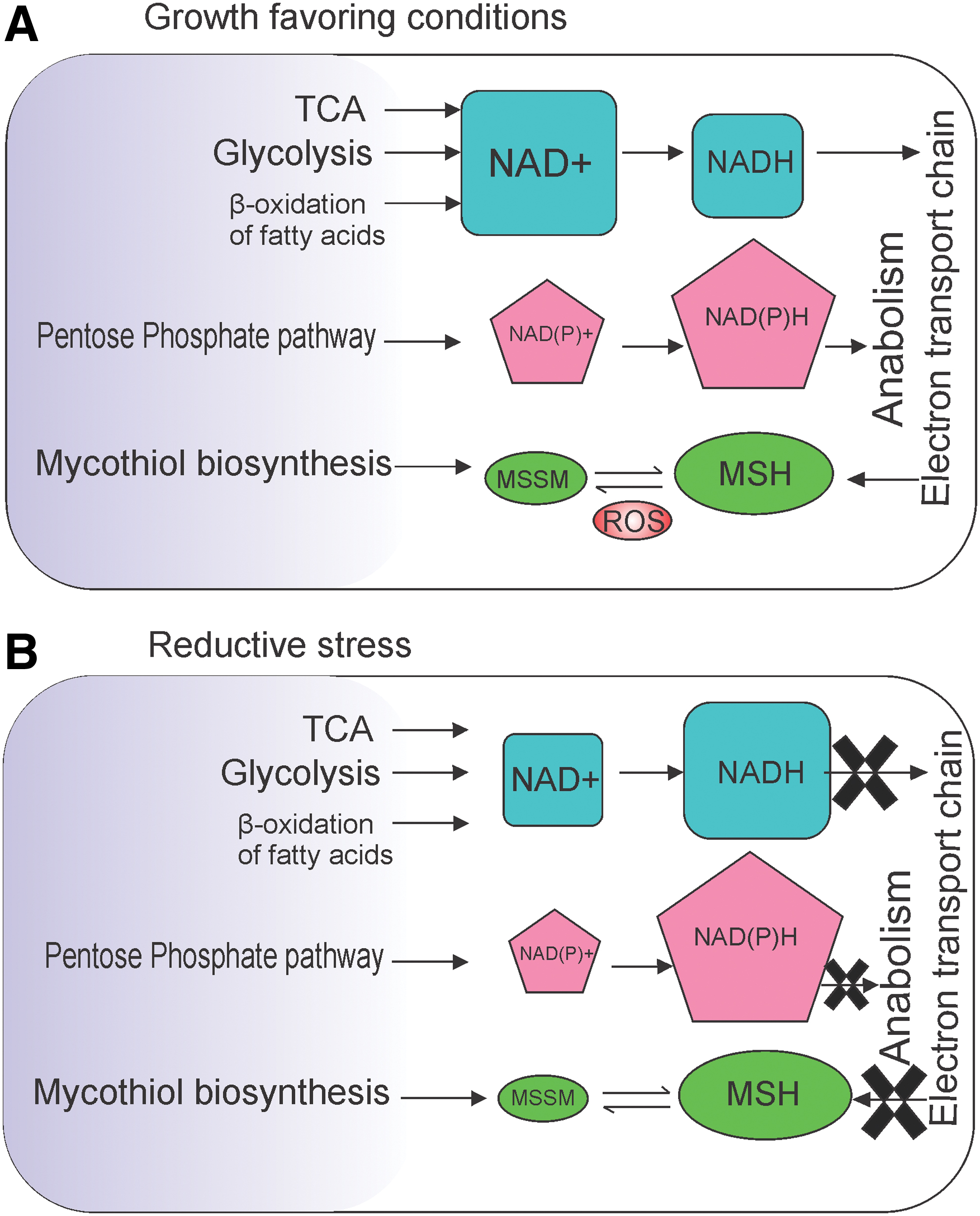
Mycobacterium tuberculosis (Mtb) is one of the most prevalent pathogens, causing 1.3 million mortalities annually, manifested in the form of tuberculosis (TB) disease (152). Approximately one-third of the world's population is estimated to be latently infected with Mtb and is at 10% lifetime risk of developing active disease (23). The synergy of human immunodeficiency virus-TB coinfection has further aggravated this as a major public health problem (97). During its infection cycle, Mtb is exposed to a number of redox stresses, such as hypoxia, increased levels of GSH inside granuloma, host-generated nitric oxide (NO), and carbon monoxide (CO), which have the potential of inhibiting bacterial respiration (71). However, Mtb survives many of these conditions. Experiments by Wayne and Hayes (148) have demonstrated that sudden exposure of Mtb to anaerobic conditions leads to cell death. However, gradual depletion of oxygen (O2) leads to emergence of drug-tolerant nonreplicating persistence (NRP) bacteria (148, 150). Interestingly, Mtb isolated from the caseum of rabbit lungs is nonreplicating and tolerant to first- and second-line TB drugs (115). Consequently, use of biosensor pimonidazole confirmed the hypoxic nature of Mtb-laden granulomas from guinea pigs, rabbits, and nonhuman primates (140). Moreover, the accumulation of NADH within Mtb recovered from mouse lungs was observed utilizing 14C nicotinamide incorporation assays (26). Likewise, metabolic profiling using 1H magic angle spinning nuclear magnetic resonance studies demonstrated the presence of increased concentrations of GSH inside Mtb-laden granulomas inside the lungs of guinea pigs (126). Interestingly, GSH is inhibitory to Mtb growth (139) and mycobacterial mutants defective in the uptake of GSH grow better inside macrophages (138). These observations strongly suggest that Mtb is exposed to reductive stress within the host. In response to redox stress, Mtb modulates its metabolic plasticity for surviving and persisting inside the host (133). Metabolic pathways of Mtb are tightly regulated by the redox state of NADH/NAD+, NADPH/NADP, and mycothiol (MSH)/mycothiol (oxidized) (MSSM) (summarized in Fig. 1). Understanding the metabolic pathways and redox homeostatic pathways utilized by Mtb to survive inside the host could pave the way for developing anti-Mtb drugs. An earlier review by Steyn and coworkers has shed light on the effect of reductive stress in microbes (42). In this review, we summarize the current understanding of sensing and neutralization of reductive stress in mycobacteria.
Thiol Redox Balance in Mycobacterium
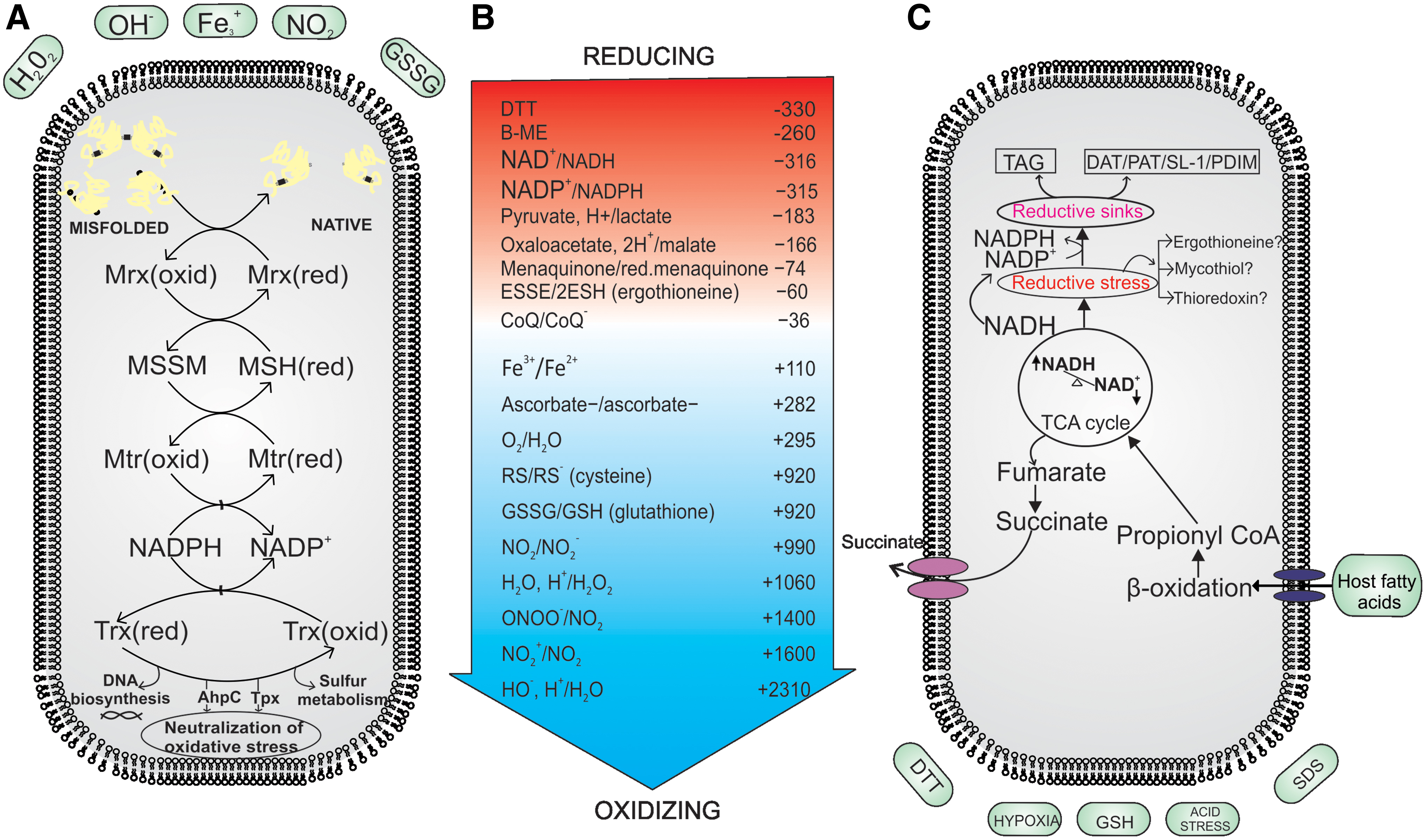
Most eukaryotes and a large number of prokaryotes utilize GSH to maintain redox balance (41, 98). GSH is a tripeptide (γ-glutamyl cysteinylglycine) that acts as the major nonprotein thiol in cells. It is primarily involved in redox homeostasis, but also has a variety of other functions in the cell (98). Interestingly, genus Mycobacterium is devoid of GSH and produces two different types of low-molecular-weight thiols, that is, MSH (being the reduced form) and MSSM (being the oxidized form) and ergothioneine (EGT). In addition to these thiols, Mtb utilizes thioredoxins (Trxs, Trx for singular) to maintain thiol redox homeostasis (depicted in Fig. 2). Here, we explain the molecular functioning of these buffering systems and elaborate their importance in mycobacterial survival under reductive stress.
Mycothiol in thiol redox homeostasis of Mycobacterium
MSH is formed through a series of enzymatic reactions that lead to conjugation of cysteine residues with the pseudodisaccharides of myo-inositol and N-glucosamine (4, 88). This conjugation results in the stabilization of cysteine against autoxidation. Being the dominant species among nonprotein thiols, MSH is produced in millimolar concentrations in cells (87). The role of MSH in protecting against oxidative stress is well documented. MSH can reduce the oxidized cysteine residues of a protein through mycoredoxin, generating MSSM in the process (137). MSSM can then be reduced by mycothione reductase (95). MSH mutants are more susceptible toward nitrosative and oxidative stresses, suggesting an important role for MSH in protection against ROS and reactive nitrogen intermediates (RNI). These mutants have decreased survival compared with wild-type bacilli after treatment with hydrogen peroxide (H2O2) and plumbagin, a superoxide generator (89, 105, 106). In Mtb, mshB and mshD mutants have increased sensitivity to cumene hydroperoxide and H2O2, respectively (27, 28). These data clearly established a protective role of MSH against oxidative stress. However, its role in reductive stress is poorly defined.
While the basal level of MSH:MSSM in fast-growing Mycobacterium smegmatis (Msm) mc2155 is 200:1 (higher levels of reduced thiols), in slow-growing Mycobacterium bovis bacillus Calmette-Guerin (BCG), it is estimated to be 50:1 (134). This observation suggests different regulatory mechanisms for maintaining cellular thiols in fast- and slow-growing mycobacterial species. In addition, while high concentrations of GSH (4–8 mM) inhibit the growth of slow-growing BCG, such concentrations do not affect the growth of Msm (96). These observations favor the hypothesis that different mechanisms are present for dissipating reductive stress in fast- and slow-growing mycobacterial species.
Measurements of MSH:MSSM have tremendously benefited from the development of a genetically encoded green fluorescent protein (GFP)-based sensor (18), in which mycoredoxin is fused with redox-sensitive GFP, enabling real-time measurement of MSH:MSSM in mycobacterial cells. Using this probe, Singh and coworkers revealed that intracellular Mtb experiences reductive stress inside the acidified phagosomes of macrophages (84). In another study, Abramovitch and coworkers utilized redox sensor R0GFP and demonstrated that Mtb faces reductive thiol environment upon mycobacterial growth on reduced carbon sources in the presence of acidic pH (9). They further demonstrated that thiol reductive stress (TRS) initiates growth arrest in Mtb cells similar to the NRP-like phenomenon. Importantly, this growth arrest was observed only in slow-growing Mtb H37Rv and was not observed in fast-growing Msm. Together, these observations indicate a unique biological response of slow-growing mycobacterial species to TRS and therefore require further exploration.
EGT in thiol redox homeostasis of Mycobacterium
EGT is a cytoprotective antioxidant that utilizes sulfurized histidine residues to exert protection (46). EGT plays an important role in regulating redox homeostasis, microbial physiology, and cellular metabolism in Mtb and several other microbes (12, 96, 108, 113). In mycobacteria, enzymes involved in EGT biosynthesis are encoded by a single genetic locus (egtA through egtE) as originally identified in Msm (117). Genetic mutants attenuated for EGT biosynthesis established the role of EGT in redox homeostasis. While Msm mutants lacking either EGT or MSH alone can resist oxidative stresses generated by cumene hydroperoxide and tert-Butyl hydroperoxide (114), mutants lacking both EGT and MSH are hypersusceptible to peroxide stress, suggesting roles of the thiol systems in protecting against oxidative stress (114). The sensitivity of mycobacterial cells lacking EGT to oxidative stress was confirmed by Steyn and coworkers (113). Using the ROS-responsive dye CellROX Green, they showed that mutants attenuated for producing either MSH or EGT have higher ROS levels compared with wild-type bacilli. Furthermore, using colony forming unit-based methods, they demonstrated higher sensitivity of mutants lacking EGT toward oxidative stress and antimycobacterials. Importantly, Steyn and coworkers demonstrated that EGT levels increased in the MSH-deficient Mtb strain, whereas MSH did not accumulate in EGT-deficient mutants (113). Finally, the importance of EGT in Mtb's survival inside hosts was demonstrated using a mouse model (113). It is worth noting that redox and NO sensor WhiB3 and PknD regulated intracellular EGT levels (108, 113). Both WhiB3 and PknD suppressed the biosynthesis of EGT. Interestingly, whiB3 mutant cells accumulate EGT, but still experience oxidative thiol stress at acidic pH (84). The above-cited literature clearly defines the importance of EGT in mycobacterial physiology (depicted in Fig. 3). However, further research is required to understand the exact role of EGT in cellular functions.
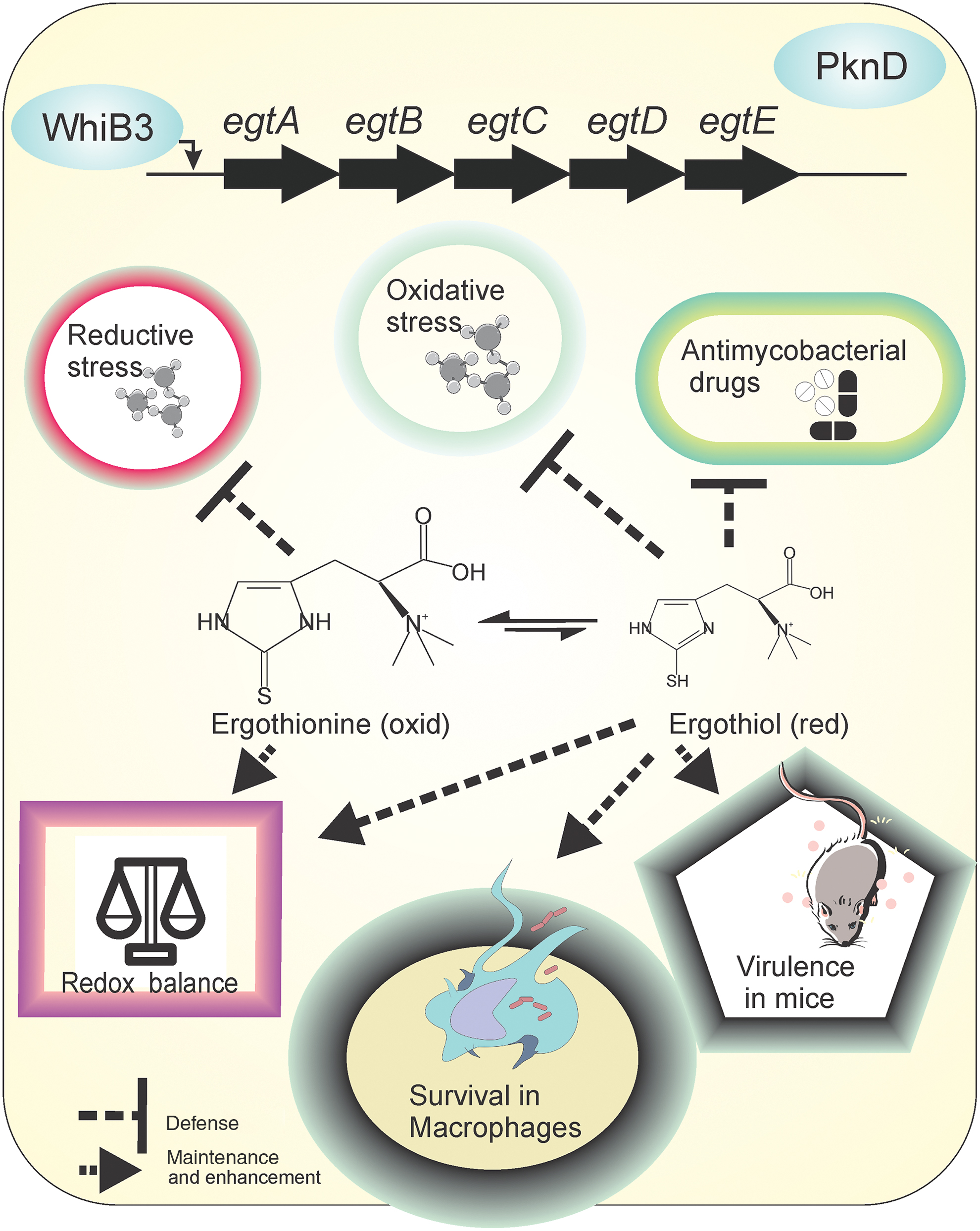
An important property of EGT is its low redox potential of −60 mV compared with other thiols, such as GSH (−240 mV) (99) and MSH (−230 mV) (118). This makes it resistant to autoxidation in comparison with other thiols. Furthermore, at the physiological pH of 7.0, EGT remains in oxidized form rather than in reduced form (29). This resistance to oxidation raises the question of how EGT protects cells from oxidative stress. Possibly, EGT could be a protectant against TRS rather than oxidative stress. This hypothesis is supported by the observation that lipid oxidation and the levels of peroxides are lower in the EGT deletion mutant compared with wild type (123). Such decrease in basal levels of peroxide and oxidized lipids may arise from accumulated reduced thiols inside the EGT mutant. We suggest that the role of EGT in protection against reductive stress shall be further explored.
Trx in redox homeostasis of Mycobacterium
Trxs are small redox active ubiquitous proteins having disulfide reductase activity (75). They play an important role in maintaining a reducing cytoplasmic environment and are capable of harvesting reducing power from NADPH for deoxyribonucleotide synthesis, sulfur metabolism, oxidized protein repair, detoxification of oxidative stress, and so on (50). A conserved WCXXC motif (i.e., the Trx fold) is used as a redox motif and is reduced by thioredoxin reductase (TR) that utilizes FAD as a cofactor for transferring electrons from NADPH to oxidized Trxs (153).
Mtb possesses three Trxs (trxA, trxB, and trxC), an uncharacterized ThiX (Rv0816c) and a TR (Rv3913). All three Trxs possess insulin reductase activity, but TR is capable of reducing only TrxB and TrxC, suggesting that the latter two may be the only functional Trxs (2). Interestingly, TrxB and TrxC are induced in response to oxidative and nitrosative stress, whereas TrxA is not. The Mtb Trx–TR system has been implicated in the detoxification of oxidative stress through alkyl hydroperoxidase AhpC and thiol peroxidase Tpx (65). Whether the Trx–TR system of Mtb also protects Mtb from reductive stress has not been analyzed. Importantly, TR is essential for in vitro growth (55), suggesting a necessary function of TR during normal growth even in the absence of oxidative stress. In agreement with these findings, Ehrt and coworkers utilized a conditional knockdown strain and demonstrated that the depletion of TR leads to Mtb cell lysis (76). They further demonstrated the requirement of TR in establishing and maintaining infection in mice. TR-depleted cells were hypersensitive to the thiol oxidant diamide but resistant to ROS or RNI, indicating an important role of the Trx–TR system in maintaining an intracellular reductive thiol environment. Testing these conditional mutants against the thiol reductant dithiothreitol (DTT) could possibly reveal the function of this system in protecting against TRS. The Trx–TR system is regulated by sigma factor SigH and its cognate anti-sigma factor RshA, which is redox active (102, 127). It would be interesting to determine whether Trx–TR and SigH play critical roles in regulating responses to TRS or thiol oxidizing stress.
Effect of reductive stress on drug sensitivity and tolerance in Mycobacterium
The redox state of mycobacterial cells is closely associated with the activity of antimycobacterial agents. Indeed, many frontline TB drugs, such as isoniazid (INH), ethionamide, and PA-825, are activated by the reducing intracellular environment (primarily through NADH) of the cell. Furthermore, both EGT and MSH are critical for mycobacteria to tolerate antimycobacterials, such as rifampicin (RIF), INH, bedaquiline, and clofazimine. In agreement with this, mutants attenuated in the biosynthesis of MSH and EGT are susceptible to these drugs (113). Importantly, NADH accumulation in response to antimycobacterials was heightened in intracellular bacteria compared with bacteria in culture medium (19). Thus, the implications of NADH accumulation with respect to drug susceptibility and resistance require further analysis.
Effects of TRS have been studied both in gram-positive and gram-negative bacteria. Exposure to thiols leads to inhibition of bacterial growth through the inhibition of DNA, RNA, and protein synthesis (67) and with the unfolding of proteins in the periplasmic space. Similar to gram-negative bacteria, exposure to TRS by GSH leads to inhibition of mycobacterial growth (139). To study the effect of TRS on Mtb, Trivedi et al. exposed the bacterium to varying concentrations of DTT (132). They demonstrated that DTT-induced stress resulted in the formation of biofilms at liquid–air interfaces in shaking cultures and at flask bottoms in standing cultures (132). Intriguingly, these biofilms harbor drug-tolerant, metabolically active bacterial cells. To further understand the TRS-mediated effects, transcriptome analysis was performed after treatment with DTT, which suggested that TRS downregulates DNA synthesis, RNA transcription, and protein translation. TRS induces the genes related to iron metabolism, components of type VII secretion system, and markers of oxidative stress. Further analysis suggested that the TRS response is mediated by sigE, sigB, and whiB3; whether they coordinate mycobacterial response to TRS and play a role in biofilm formation is yet to be determined. The above findings suggest that TRS induces biofilms and thus drug tolerance. Contrary to these findings, Jacobs and coworkers suggested that exposure of persisters to TRS (mediated by cysteine) leads to increased respiration and efficient killing of Mtb by frontline drugs, such as INH (142). These findings suggest that persister cells face reductive stress and further exposure to TRS through high cysteine concentrations (4 mM in this case) leads to enhanced Mtb death. The same group demonstrated that cysteine is converted to cystine within cells exposed to TRS. This conversion is associated with increased levels of H2O2 inside the mycobacterial cells. Importantly, the combination of INH and thiol N-acetyl cysteine killed intracellular mycobacterial cells with higher efficiency than INH alone. These observations also suggest that intracellular Mtb is faced with reductive stress. Exposure to increased reductive stress renders Mtb further susceptible to killing by antibiotics such as INH. Importantly, another study has suggested that prolonged treatment with INH is associated with accumulation of NADH in Mtb cells in vitro and inside macrophages (19). Another antioxidant vitamin C effectively kills mycobacterial cells (141). Jacobs and coworkers demonstrated that vitamin C increases the concentration of iron in Mtb cultures and assists the generation of ROS through the Fenton reaction (141). It was also observed that a combination of vitamin C and INH was more potent at killing Mtb cells than INH alone, suggesting a correlation of redox stress with drug susceptibility. Interestingly, vitamin C is implicated in improved prevention and treatment of TB (58, 80). TB patients have been reported to carry vitamin C deficiency, (5) and host defenses are known to accumulate intracellular ascorbic acid (at millimolar concentrations) in response to infections (13, 73, 146, 151).
Mycobacterial Mechanisms of Sensing Reductive Stress
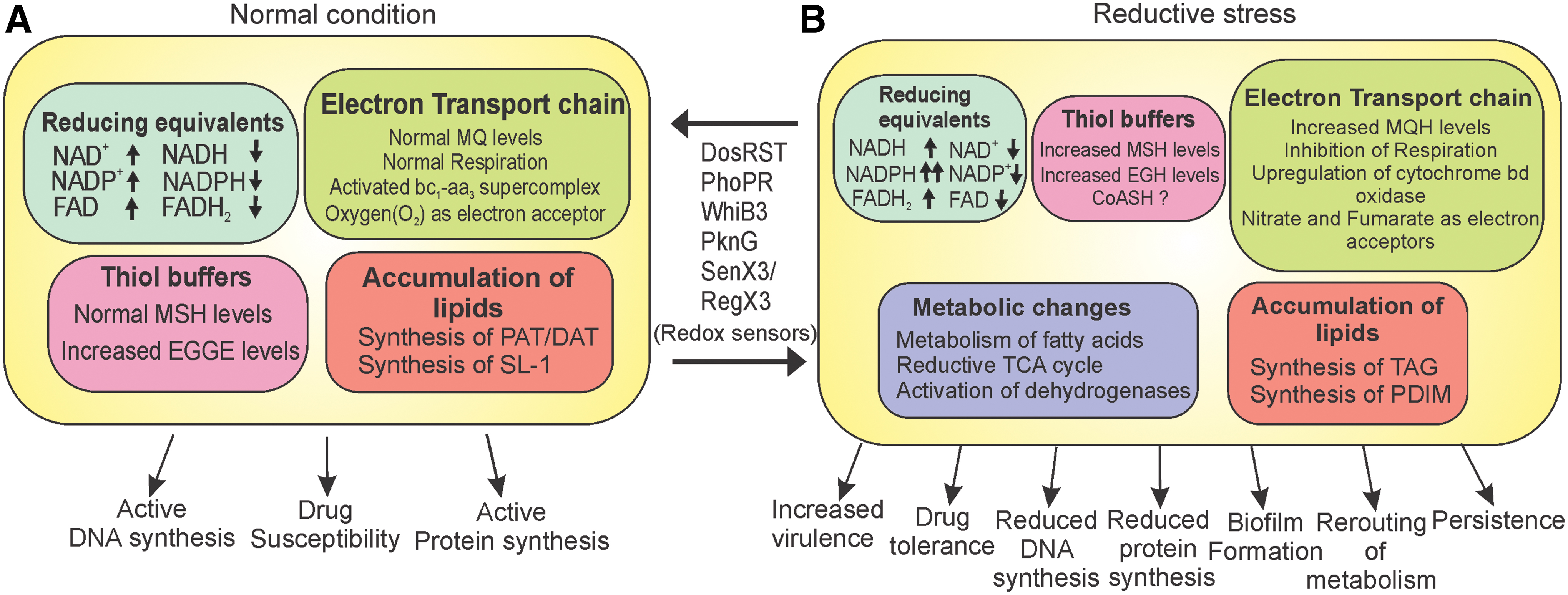
Reductive stress may build up within Mtb cells residing in the host. Hypoxia, one of the important factors in host–pathogen interactions, has been extensively studied as a source of reductive stress (148). Exposure to hypoxia results in inhibition of respiration, decreased ATP levels, and increased NADH levels, leading to reductive stress (104). Importantly, hypoxia-induced reductive stress leads to the inhibition of mycobacterial growth, elongation of cells, and induction of drug tolerance. These cells may survive for years and resume growth upon exposure to ambient O2 levels (148). Thus, the hypoxia-induced NRP state and O2-induced resumption of growth have been extensively studied. The genetic circuits that regulate transition into the NRP state and vice versa have been explored. To respond to these stresses, Mtb uses a diverse machinery of redox sensors that trigger genetic responses capable of modulating metabolism to survive during reductive stress. The sensory array includes heme-based proteins, such as DosS, DosT, and SenX3, and iron–sulfur cluster-based WhiB proteins. Other proteins that play an important role in sensing reductive stress include PknG and PhoPR. In the following section, we describe these sensory modules, their sensing mechanisms, and their contributions to mycobacterial physiology.
The DosRST two-component system: regulation of NRP
The DosRST sensory system consists of heme-based histidine sensor kinases, DosS and DosT, that sense and relay signals to response regulator DosR. DosR regulates the expression of a repertoire of ∼50 genes collectively called the Dos/dormancy regulon. This regulon is highly upregulated under hypoxia and consists of genes encoding for nitrate reductase (narK2X and narX), fumarate reductase (Frd) (frdABCD), formate hydrogen lyase (Rv0081–Rv0088), ferredoxin (fdxA), and chaperone proteins, such as α-crystallin, genes encoding for DNA repair (nrdZ), and triacylglycerol (TAG) biosynthesis (tgs1 and Rv2623). Besides hypoxia, this two-component system (TCS) responds to a variety of stresses, including NO and CO (70, 91, 94, 119, 143). These stresses lead to inhibited respiration and thus are associated with the cellular accumulation of menaquinol, reduced cytochromes, and NADH. The system is thought to play an important role in facilitating the transition of actively replicating cells into a hypoxia-induced NRP state. Ablation of DosRST leads to reduced survival in response to hypoxia; Mtb recovers from hypoxic cultures upon resumption of ambient O2 levels (25). Furthermore, it was observed that the dosR mutant is highly susceptible to acidic pH under anaerobic conditions (107), suggesting its importance in survival under oxygen limiting conditions. Earlier studies using SCID and DBA2 mice have shown hypervirulence in mutant strains, raising concerns on the contribution of this regulon in mycobacterial survival within mice (93). However, recent studies using C3HeB/FeJ mice (45), guinea pigs, rabbits, and nonhuman primates (83) have clearly established a role of this regulon in mycobacterial virulence, survival, and persistence.
The histidine kinases DosS and DosT sense similar redox cues in mycobacterial cells but through different mechanisms (62, 63, 72, 128). Both kinases have two tandem cGMP-specific phosphodiesterases, adenylyl cyclases, and FhlA (GAF) domains. The first GAF domain binds heme through a dative covalent bond. Both proteins exhibit autokinase activity in their ferrous deoxy form; thus, both sensor kinases activate DosR in hypoxia (72, 128). In the presence of O2, DosT binds O2 in a concentration-dependent manner. The binding of O2 with DosT heme leads to a decrease in the autokinase activity (72, 128). However, NO and CO can bind with DosT and lock it in an active state, thereby upregulating dormancy regulon (72). Two mechanisms have been suggested for O2 sensing by DosS. One of the groups suggested that the heme group of DosS is redox sensitive and is rapidly oxidized upon exposure to O2 (31, 32, 72). This oxidation leads to inhibition of the autokinase activity. Such a mechanism would require a redox partner that could reduce DosS when O2 is limiting. FADH has been proposed as a potential donor for the reduction of DosS (32), but the redox active protein that catalyzes the reduction of DosS has not yet been identified. Other studies suggested that DosS is an O2 binding protein but has a higher affinity for O2 compared with DosT (62, 63, 128). Similar to DosT, DosS can be locked in an active state by NO and CO (72). Given that the Dos regulon is upregulated by ascorbate, which is capable of feeding electrons into the electron transport chain (ETC) (59), it is plausible that DosS is indeed a redox sensor. Furthermore, the Dos regulon can be upregulated when long-chain fatty acids are the primary carbon source (111). The Dos regulon is also modulated by the phosphorylation of DosR by the serine/threonine kinase PknH, which has been shown to protect against redox stress (30). In summary, DosS and DosT coordinate the mycobacterial response to a number of reductive signals, including hypoxia, NO, CO, and to inhibited respiration (as depicted in Fig. 4).
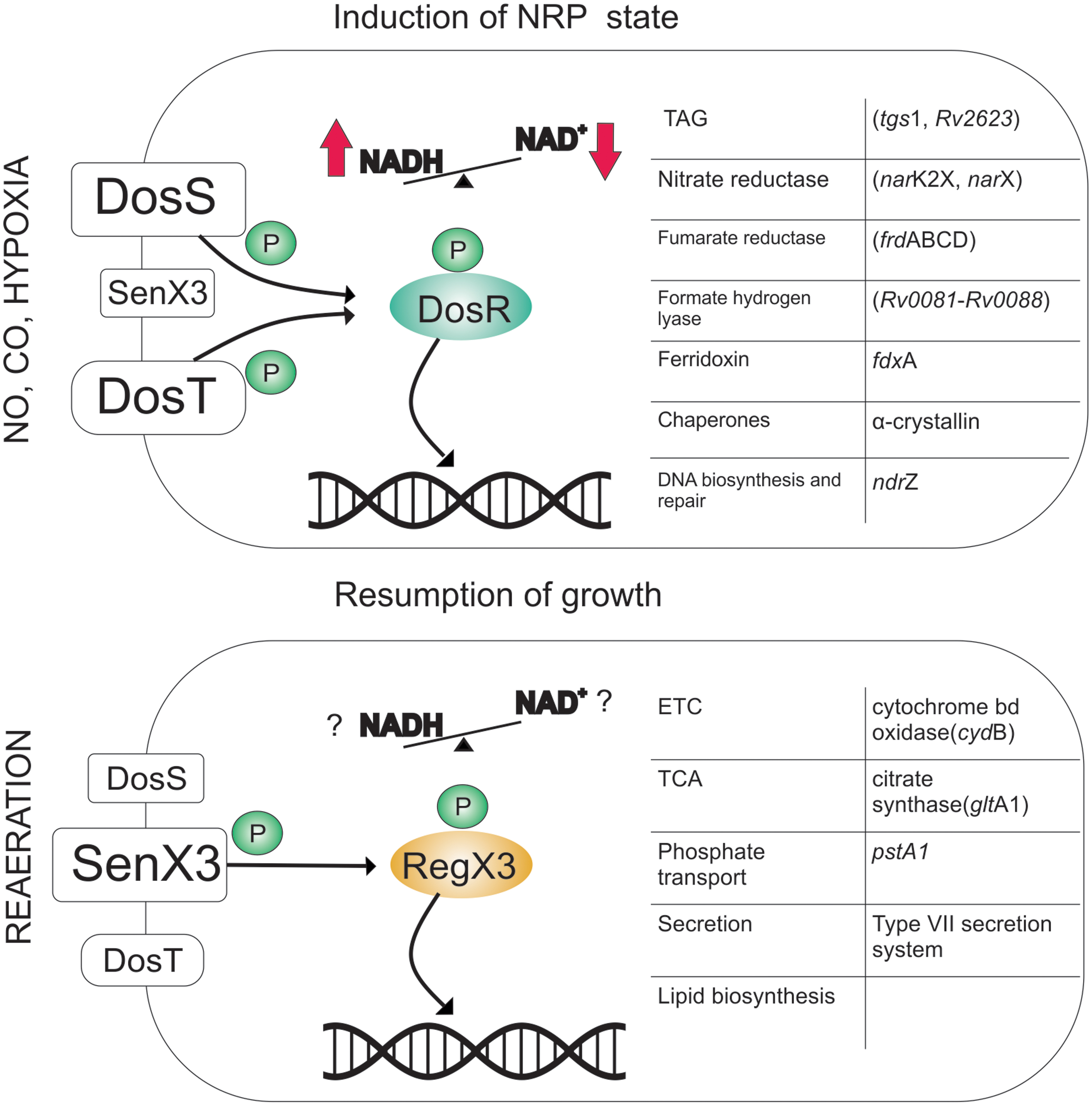
The SenX3/RegX3 TCS: regulation of regrowth upon resumption of ambient oxygen
The SenX3/RegX3 system plays a vital role in resumption of growth in hypoxic cultures upon exposure to ambient O2 (124). In this system, SenX3 is the sensor histidine kinase, whereas RegX3 is the response regulator. This TCS is required for mycobacterial survival inside macrophages and in guinea pigs and mice (93, 109). Interestingly, SenX3 contains a Per–Arnt–Sim domain, which has the potential to bind heme; thus, SenX3 has been predicted to be an O2 sensing protein (109). Singh and Kumar (124) demonstrated that SenX3 is indeed a heme-based sensor, further demonstrating that the autokinase activity of SenX3 is activated upon exposure to O2. They presented evidence to suggest that the SenX3 kinase is a three-way nanoswitch with three levels of activities. Oxidized protein has the maximum activity, whereas the kinase activity is reduced in the ferrous-deoxy form. This autokinase activity is further reduced upon binding with NO or CO. In agreement with biochemical data, a senX3 mutant Mtb strain was attenuated during regrowth in response to the resumption of ambient O2 levels. Furthermore, the regrowth of hypoxic cultures in O2 was inhibited by the presence of NO and CO, and was dependent on SenX3 (124) (Fig. 4). These findings suggest that the SenX3/RegX3 system could regulate the resuscitation of Mtb from the persistent state when infecting humans. Given that RegX3 is a regulator of cytochrome bd oxidase (cydB) of ETC and citrate synthase (gltA1) of tricarboxylic acid (TCA), RegX3 can be expected to be a regulator of mycobacterial growth (110). In addition to regrowth, RegX3 is also believed to regulate phosphate transport, type VII secretion system (39), and mycobacterial persistence (85). Given that SenX3/RegX3 TCS is required for regrowth, its role in lipid biosynthesis and for growth on different carbon sources should be analyzed.
The PhoP/PhoR TCS: a regulator of acid stress response
Upon inhalation of Mtb-laden aerosol by a human host, Mtb is phagocytosed by alveolar macrophages. There, it encounters a pH of ∼6.2–6.4 within phagosomes (78, 130) after arresting phagosome–lysosome fusion. This arrest can be relieved through macrophage activation by cytokines, such as interferon gamma. In activated macrophages, Mtb faces a pH of 4.5–5.4 within acidified vacuoles (78, 116, 130, 140). The acidic pH leads to the arrest of mycobacterial growth despite the availability of nutrients, such as fatty acids, and O2 as a terminal electron acceptor (9). This growth arrest is associated with drug tolerance, variability in acid-fast staining, modulation of lipid biogenesis, and expression of virulence factors (8). Transcriptional response to acidic pH is coordinated by the PhoPR TCS (1, 9, 10). Given that the intraphagosomal transcriptional response is regulated by PhoPR TCS and its deletion mutant is attenuated for survival inside macrophages and in infected animals, PhoPR TCS is considered a virulence factor of Mtb (79, 145). The phoPR mutant Mtb strain has been proposed as a vaccine candidate and is currently in clinical trials (79). Importantly, PhoPR TCS regulates the expression of virulence lipids and the secretion of virulence factor, ESAT-6-CFP10 (6, 43).
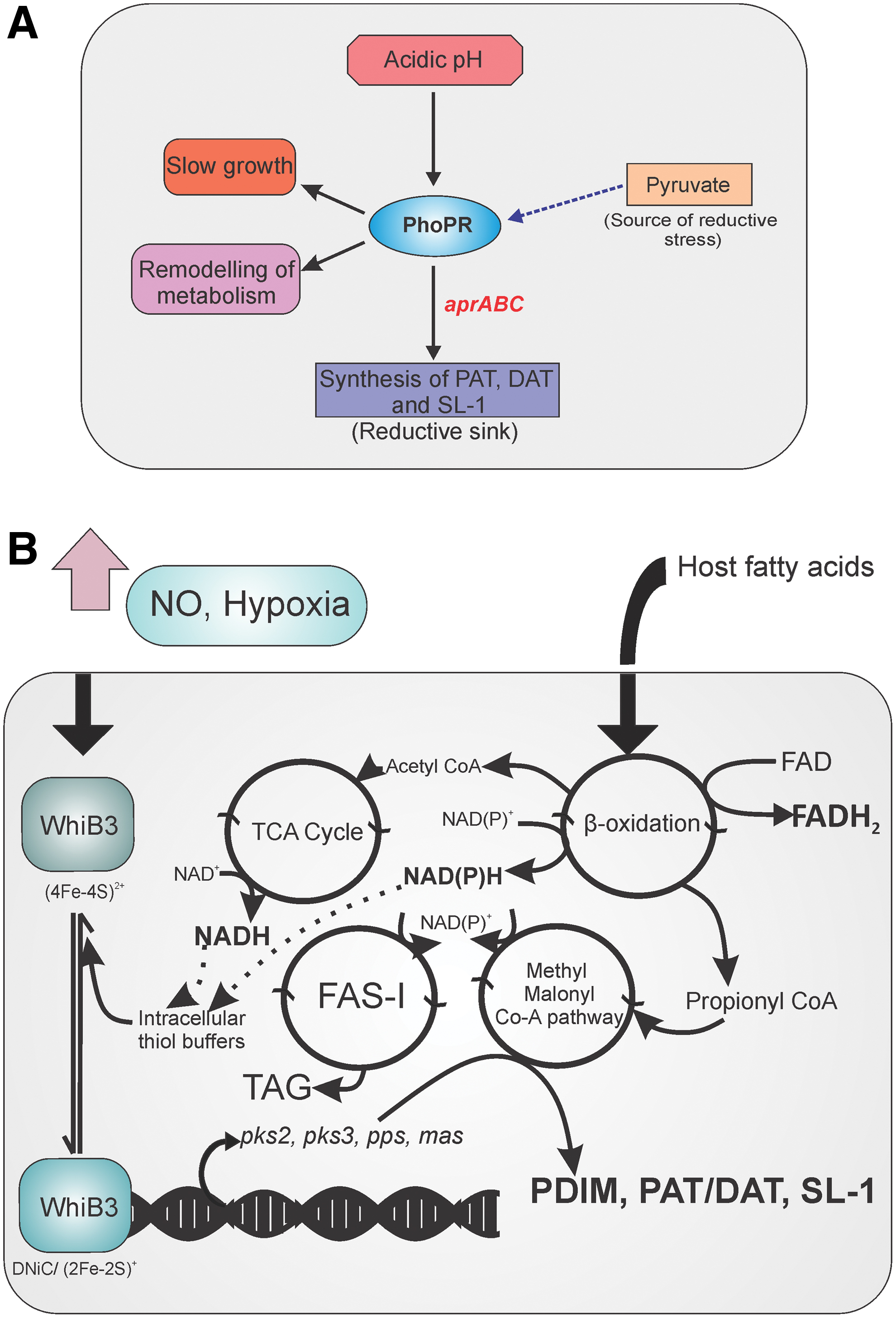
PhoPR TCS acts as a regulatory driver that connects redox homeostasis with central metabolism and virulence in Mtb. Baker et al. established that Mtb cells experience reductive thiol stress at a pH of 5.7 (9). Similar reductive stress has been observed when Mtb was grown on medium containing pyruvate as the carbon source instead of glycerol (9). Such reductive stress could be sensed through the PhoPR TCS to modulate metabolism so that Mtb can utilize host-associated carbon sources, such as cholesterol and long-chain fatty acids. However, the identity of the signaling molecules and the mechanism of signaling remain poorly understood. PhoPR is required for maintaining redox homeostasis for growth on fatty acids. The phoP mutant is compromised in its capability to maintain the cellular thiol redox homeostasis when cultured in acidic pH of 5.7, or upon growth on substrates feeding directly into the TCA cycle. Importantly, PhoP is the regulator of the biosynthesis of lipids, such as sulfolipids, poly-acyltrehalose (PAT), and diacyltrehalose (DAT) (51, 54). Upon accumulation of NADH and NADPH due to the use of fatty acids as the carbon source or due to growth under acidic pH, Mtb has been proposed to synthesize the sulfolipids and acyltrehaloses (PAT and DAT) to oxidize the reduced cofactors (9). PhoP guides the synthesis of these lipids through the aprABC gene locus (1). Sulfolipids, PAT, and DAT are considered virulence lipids that modulate the immune system and help in mycobacterial survival inside the host. The phoP mutant of Mtb is unable to induce the biosynthesis of these virulence lipids in response to acidic pH or reducing carbon sources. Instead, the mutant relies upon TAG biosynthesis for dissipating the reductive stress (9) (Fig. 5A). We suggest that the link between TRS and altered carbon metabolism should be further analyzed.
WhiB3: an integrator of metabolism with the intracellular redox state of the cell
WhiB3 was discovered as a missing piece in the role of principal sigma factor RpoV (SigA) in mycobacterial virulence (129). Steyn et al. demonstrated that WhiB3 interacts with RpoV from virulent M. bovis, but does not interact with the RpoV from avirulent M. bovis (129), which has an arginine to histidine mutation at position 515. This point mutation was shown to be responsible for the loss of virulence in M. bovis (33). Singh et al. demonstrated that WhiB3 is an iron–sulfur {Fe–S} cluster protein that senses intracellular NO and O2 levels (122). Surprisingly, the Mtb whiB3 mutant strain was not attenuated for survival in hypoxia or upon exposure to NO. Interestingly, this mutant was attenuated for growth on medium containing glucose, succinate, fumarate, pyruvate, and citrate as carbon sources. However, this strain was able to grow better than the wild type on medium containing acetate as the sole carbon source. These findings were the first to indicate that Mtb WhiB3 is a sensor of change in the intracellular redox state generated by growth on different carbon sources. The whiB3 mutant strain has an altered cell size and shape, and shows significantly different colony rugosity, suggesting that WhiB3 plays an important role in regulating the cellular physiology of Mtb (121). Elegant studies by Cumming et al. revealed that the Mtb whiB3 mutant has an altered bioenergetic state. Mtb whiB3 mutant experiences reductive stress when grown on medium containing cholesterol, palmitate, or glucose as the carbon source and possesses significantly higher levels of NADH:NAD+. Interestingly, this strain is able to maintain a suitable and comparable NADH:NAD+ when grown on acetate or propionate. These experiments suggest that WhiB3 is utilized by Mtb to regulate central metabolism in response to different carbon sources (35). These differences in the cellular bioenergetics of the whiB3 mutant strain were confirmed through measurements of O2 consumption rate (a measure of ETC flux) and extracellular acidification rate (a measure of carbon catabolism).
To answer how Mtb dissipates accumulated reductive cofactors, such as NADPH, due to growth on fatty acids as a carbon source inside macrophages, Singh et al. (121) performed a series of lipid labeling experiments and demonstrated that Mtb synthesizes polar lipids, such as PAT and TAG, inside macrophages to replenish the depleted NAD+ pool. However, the Mtb whiB3 mutant strain is attenuated in PAT production and thus channels reductive flux through the synthesis of phthiocerol dimycocerosate (PDIM). Interestingly, Mtb synthesizes PAT under growth-favoring conditions or upon thiol oxidative stress caused by diamide. However, upon TRS, Mtb shifts metabolism toward the synthesis of PDIM and TAG (121). In addition, lipid analyses of the whiB3 mutant demonstrated differences among PAT, DAT, sulfolipid-1 (SL-1), PDIM, and TAG. Through a series of DNA binding studies, Singh et al. (121) also demonstrated that WhiB3 can directly bind to the promoter of genes involved in lipid biosynthesis, such as pks2 and pks3. Importantly, the DNA binding was dependent on the redox state of the cysteine thiols, suggesting that WhiB3 can directly regulate the lipid biosynthesis. These studies clearly suggest that Mtb WhiB3 is a regulatory switch that connects the cellular redox homeostasis with lipid anabolism (Fig. 5B). Given that the Mtb whiB3 mutant is attenuated for survival inside macrophages, and because it is induced inside macrophages in a pH-dependent manner (112), Mehta et al. measured the redox potential of MSH (Emsh) in Mtb cells in response to decreasing pH, using a genetically encoded biosensor (84). They observed that acidic pH caused a reductive shift in Mtb Emsh. However, the whiB3 mutant was unable to adjust its Emsh in response to acidic pH and was attenuated for survival at a pH of 4.5. Microarray profiling reaffirmed the various pathways required for redox homeostasis, such as Trxs (trxB1, trXB2, trxC), sodA, and the MSH biosynthesis pathway, controlled by WhiB3 in response to acid stress. These findings clearly linked WhiB3 with Emsh regulation in response to changes in metabolism upon acidic pH stress.
PknG: a regulator of redox homeostatic system
Recently, a novel system of regulated hydrolysis of excess NADH, FAD, and adenine dinucleotide phosphate-ribose was described (155). In this system, excess NADH leads to increased PknG expression, which phosphorylates ribosomal protein L13 and facilitates its interaction with Nudix hydrolase RenU. Nudix hydrolases are ubiquitous hydrolytic enzymes capable of cleaving NUcleoside DIphosphates linked to X (NDP-X), to yield NMP and P-X (82). This association activates the NADH and FAD hydrolytic activity of RenU. Importantly, disruption of components of this redox homeostatic system leads to NADH and FAD accumulation, increased susceptibility to oxidative stress, reduced survival inside hosts, and inability to form biofilms (155). PknG is known to phosphorylate and regulate functioning of the glycogen accumulation regulator GarA (90). Unphosphorylated GarA inhibits α-ketoglutarate decarboxylase; in the TCA cycle, suggesting that PknG could regulate central metabolism through GarA. Furthermore, PknG possesses a rubredoxin (Rbx)-like domain at its N-terminal. This Rbx domain plays a key role in the functioning of PknG and is regulated by the redox state of the cell (49, 154). In agreement with its role in redox homeostasis in mycobacteria, pknG mutant exhibits reduced Emsh and is attenuated for survival under oxidative stress and hypoxia (69). Finally, recent studies have suggested that PknG senses amino acids for regulating metabolism inside mycobacterial cells (22, 90).
Metabolic Reductive Stress and Its Dissipation in Mycobacteria
Similar to other bacteria, Mtb utilizes the NADH:NAD+ redox couple for harvesting the reductive energy and NADPH:NADP+ for using this energy in the synthesis of cellular components. During growth favoring conditions, Mtb maintains 3- to 10-fold higher levels of NAD+ compared with NADH (26, 52, 104). However, inhibition of respiration by hypoxia or NO leads to increased levels of NADH (19, 74, 104). In addition, utilization of fatty acids as carbon source inside macrophages also leads to accumulation of NADH levels, which must be oxidized back to NAD+ for permitting growth. Interestingly, the development of new genetically encoded probes for measuring NADH:NAD+ with spatiotemporal resolution has indicated a metabolic heterogeneity among Mtb populations (19, 20). These reporter strains have confirmed that inhibition of respiration leads to NADH accumulation. Surprisingly, ROS-mediated stress also leads to NADH accumulation within cells (21). These findings are in agreement with a previous study in yeast cells, in which ROS lead to the inhibition of respiration and accumulation of NADH (24). We believe that similar to yeast cells, ROS may inhibit mycobacterial respiration, as reflected in the accumulation of NADH. Intracellular Mtb also faces reductive stress, even more so inside activated macrophages, perhaps due to inhibition of respiration caused by NO generated by inducible nitric oxide synthase (19). This is in agreement with earlier findings, in which Mtb recovered from lungs of infected animals showed accumulation of NADH and NADPH (26). Unfortunately, genetically encoded tools capable of measuring mycobacterial NADPH:NADP+ are currently not available. Establishing such reporter strains of Mtb could play key roles in elucidating the role of NADPH:NADP+ in reductive stress. In the following section, given the critical role of modulating metabolism in circumventing reductive stress, we describe the metabolic pathways utilized by Mtb to replenish the NAD+ pool or for avoiding further NADH generation.
Modulation of the ETC in response to reductive stress
Mtb utilizes a highly flexible and robust ETC to survive inside the host. Given that oxidative phosphorylation is critical for Mycobacterium, ETC is tightly regulated at multiple steps (64). NADH and FADH channel electrons into ETC, NADH dehydrogenase (NDH), and succinate dehydrogenase (SDH). Both NDH and SDH pump electrons into ETC via menaquinone (MK). Mtb possesses two terminal oxidases, namely, a supercomplex of menaquinol-cytochrome c oxidoreductase (bc1-aa3 supercomplex) and a cytochrome bd-type menaquinol oxidase capable of directly transferring electrons from menaquinol to O2 (64). Importantly, the proton pumping bc1-aa3 supercomplex is active under high NAD+ levels of aerobic conditions, whereas cytochrome bd oxidase is utilized under conditions where NADH levels accumulate, such as hypoxia and inside mice lungs (120), suggesting that ETC is regulated in response to reductive stress. Importantly, whereas metabolic reductive stress slows respiration, small thiols, such as DTT (56) and
Reductive TCA cycle
Mtb possesses variants of the TCA cycle (Fig. 6A) that are quite different from the canonical TCA cycle of Escherichia coli. The flexibility in choosing variants of TCA allows mycobacteria to survive various redox stresses. Interestingly, Mtb also modulates its TCA cycle for survival during hypoxia-induced reductive stress by downregulating the genes encoding for oxidizing enzymes of the TCA cycle, such as citrate synthase and isocitrate dehydrogenase, with concurrent increases in the expression of malate dehydrogenase and Frd (26, 147). In an elegant study, Watanabe et al. utilized 13C labeling and concomitant analyses of metabolites in aerobic and anaerobic conditions to demonstrate that Mtb cells divert carbon from glycolysis for the production and secretion of succinate through the reductive branch of the TCA cycle (147) (as depicted in Fig. 6A).
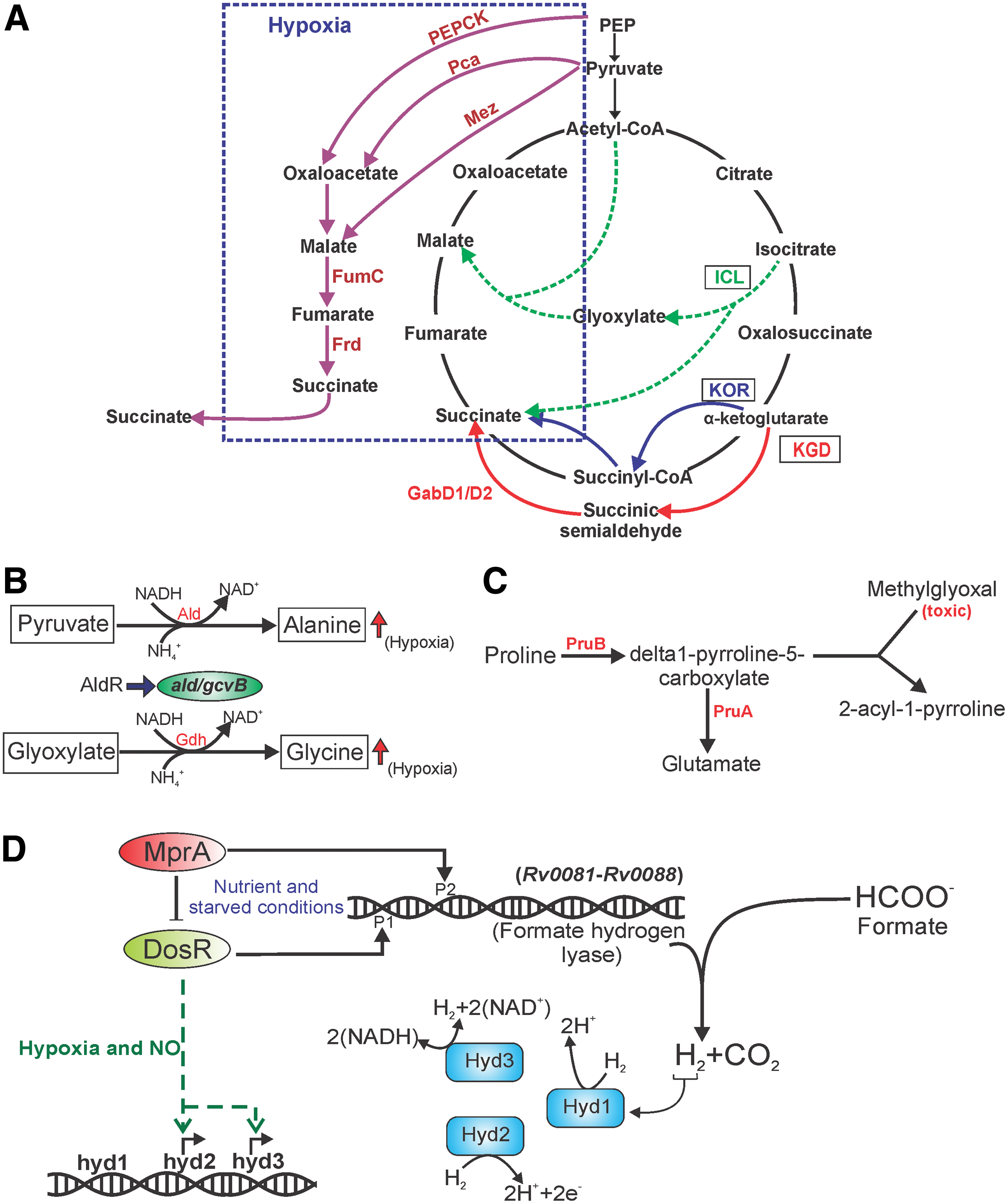
The importance of the reductive TCA cycle during hypoxia was validated by Eoh and Rhee through a 13C-based metabolic profiling of Mtb under hypoxic conditions (40) by demonstrating that the reductive TCA cycle facilitates the survival of the bacterium upon using fatty acids as the carbon source. They also demonstrated that isocitrate lyase (ICL) activity increased during hypoxia, a condition that induces reductive stress. Strains of Mtb deficient for ICL were attenuated for survival during hypoxic growth. Simultaneously, SDH has been suggested to play an important role during mycobacterial survival under hypoxia. Given that SDH connects TCA (and reductive TCA) with ETC, succinate can provide the electron flow necessary to generate sufficient membrane potential to sustain survival; excess succinate can be secreted. Interestingly, Mtb slows down respiration in response to antimycobacterials (INH, RIF, and streptomycin) through its reductive TCA cycle; inhibition of ICL activity leads to susceptibility toward these drugs (86).
Role of dehydrogenases in the dissipation of reductive stress
Mtb utilizes a number of dehydrogenases to dissipate excessive NADH and to replenish NAD+. Alanine dehydrogenase converts pyruvate into
In addition to alanine, Mtb is also capable of using proline as a carbon source. Proline metabolism is dependent on pyrroline-5-carboxylate dehydrogenase (PruA) and proline dehydrogenase (PruB), which are under the transcriptional control of membrane-anchored DNA binding protein PruC (17) (Fig. 6C). Notably, proline metabolism can help in the survival of mycobacteria on glycerol or on fatty acids and protect the bacteria from methylglyoxal, which is produced during the metabolism of fatty acids and a few carbohydrates (17). Evidently, genes involved in proline metabolism are induced upon exposure of mycobacterial cells to hypoxia (14).
Bacterial cells such as E. coli often produce formate from glucose through mixed acid fermentation. Formate is then oxidized to CO2 and hydrogen (H2) by formate hydrogen lyase (81). Mtb also encodes for a putative formate hydrogen lyase (Rv0081–Rv0088), which is under the control of redox sensory modules DosRST and MprAB. The lyase is induced in response to hypoxia and nutrient starvation (57). Mycobacteria also contain three {NiFe}-hydrogenases that can oxidize and produce H2, namely, Hyd1, Hyd2, and Hyd3 (16). Importantly, DosR induces Hyd3 (MSMEG_3931–3928) and Hyd2 (MSMEG_2719–2718) in response to hypoxia and NO. These hydrogenases lead to the generation of H2 upon O2 depletion (Fig. 6D). Furthermore, H2 production and oxidation are important for the survival of Mtb under hypoxia/anoxia because Mtb strains mutated for hydrogenases are attenuated in survival. Analyses of NADH/NAD+ levels suggest that these mutants are unable to maintain an adequate redox balance, indicating that hydrogenases contribute to survival in hypoxia/anoxia-induced reductive stress (15).
Accumulation of lipids in response to reductive stress
Mtb can also dissipate reductive energy (accumulated NADH and NADPH) through the synthesis of storage lipids such as TAG, or cell wall lipids such as PDIM and SL-1. Syntheses of these lipids consume NADH and thus relieve the bacterium from reductive stress. TAG serves as a reserved substrate and is composed of a water-insoluble triester of glycerol with a fatty acid, which produces a significant level of energy upon oxidization (3, 42, 144). Furthermore, as TAG is charge neutral, large quantities of TAG can be accumulated inside mycobacterial cells in the form of lipid droplets. Evidently, physiological conditions associated with intracellular reductive stress are associated with TAG accumulation, including growth on fatty acids under acidic pH, NO, CO, and hypoxic conditions (37, 70, 91, 119, 143). Importantly, Mtb cells require TAG synthesis when entering an NRP state and during regrowth (77). Mtb contains nearly 15 genes encoding TAG synthases (TGS). Of these, Rv3130 (tgs1) has the highest TGS activity and is under the control of a DosRST TCS (36). Given that tgs1 is under the control of the DosRST system, TGS biosynthesis is regulated by NO, CO, and hypoxia (36). TAG synthesis transfers the metabolic flux away from the TCA cycle to assist Mtb cell survival during hypoxic stress (7). In the absence of tgs1, Mtb is unable to stall its growth in response to hypoxia and is attenuated for survival under hypoxia (7). The significance of TAG accumulation in TB pathogenesis is reflected from the fact that the W-Beijing lineage of Mtb constitutively upregulates the DosRST operon and overproduces triglycerides. What remains unknown is whether the strains belonging to this lineage have increased resistance toward reductive stress. Another interesting study showed that bacteria recovered from the sputum of TB patients have TAG lipid droplets and overexpress tgs (44). These findings suggest an important role of TAG as a redox sink for dissipating reductive stress.
In addition to TAG, Mtb can also accumulate polyketide lipids, such as PDIM and SL-1, which not only dissipate reductive stress but also subvert the immune system (92, 100). Singh et al. (121) demonstrated that exposure of Mtb cells to DTT induces PDIM production along with some amount of TAG. Incidentally, a concomitant decrease in PAT was observed upon DTT treatment. PAT is a virulence lipid located in the outer layer of the cell wall and is capable of inhibiting the uptake of Mtb cells by macrophages and activation of T cells (100). In another study, long-chain fatty acids were used as carbon sources, and the lipid transcriptome was analyzed. The analysis revealed that in response to reductive stress generated due to the metabolism of long-chain fatty acids, lipid biosynthesis (especially of PDIM and TAG) was highly induced along with whiB3, dosR, and rv0081 (111). Furthermore, using a mass spectrometry-based lipidomics approach, Cox and colleagues (66) demonstrated that mycobacterial cells recovered from mice accumulate PDIM and SL-1. The metabolism of odd chain fatty acids leads to the generation of propionate, which ultimately leads to the generation of toxic methylmalonyl coenzyme A (MMCoA). Likewise, perturbation of intracellular levels of MMCoA modulates PDIM and SL-1 biosynthesis (66). Besides hypoxia and fatty acids, mycobacterial growth under acidic pH is also associated with reductive stress (summarized in Fig. 7). Interestingly, the exposure of Mtb cells to acidic pH induces the production of SL-1 (9).

Concluding Remarks
Research in the last few years has established the concept of reductive stress. NADH, NADPH, and MSH are the predominant reducing equivalents inside mycobacterial cells. Importantly, cellular accumulation of these reducing agents leads to reductive stress and growth inhibition (summarized in Fig. 8). Interestingly, the effects of hypoxia, NO, growth on fatty acids, and exposure to acidic pH on mycobacterial physiology have been studied for quite some time, but their relationship increased NADH and/or MSH levels, and thus, reductive stress has been appreciated only recently. Since the redox state of the cell is entwined with the cellular growth, mycobacteria utilize a number of sensors to monitor fluctuations in the redox state. These sensors include heme-based sensor kinases (DosS, DosT, and SenX3), iron–sulfur cluster proteins such as WhiB3, and serine/threonine kinases such as PknG. Upon detecting the reductive stress, Mycobacterium modulates its metabolism and utilizes its branched respiratory chain, reductive TCA cycle, plethora of dehydrogenases, and lipid biosynthesis for dissipating the reductive stress. Importantly, reductive stress is used by Mycobacterium as a cue for regulation of virulence (via synthesis of SL-1, PAT, and DAT) and survival inside host (via synthesis of PDIM and TAG). While on one hand, the accumulation of NADH/NADPH inside cells induces growth arrest and drug-tolerant phenotype, on the other hand, increased thiols inside mycobacterial cells are associated with biofilm formation. These observations underline the importance of reductive stress in mycobacterial physiology.
Future Directions
Despite recent recognition of the importance of reductive stress in the physiology of Mycobacterium, the field of reductive stress is still in its infancy and the opportunity to learn is bigger than ever. New tools capable of monitoring the metabolic and redox state have been instrumental in elucidating the current understanding of reductive stress in mycobacteria. We still do not clearly understand how DTT or GSH inhibits mycobacterial growth. The relationship of excess NAD(P)H and TRS with respiration, central metabolism, and protein and DNA synthesis needs to be extensively studied. To understand these fundamental questions, reporter strains in which methionine reductase (catalyzing the reduction of MSSM into MSH) is overexpressed or hyperactive need to be developed. Such strains will possess higher MSH:MSSM ratios and thus will conclusively prove the relevance of higher levels of MSH on mycobacterial physiology. Along similar lines, the effect of accumulation of NADH and NADPH on lipid metabolism and mycobacterial physiology needs to be explored. Also, reporter strains possessing higher levels of NADH and NADPH should be created to study their effects on mycobacterial survival and growth. Furthermore, reporter strains monitoring the cellular levels of oxidized/reduced EGT will be valuable. Another important aspect that needs to be examined is how reductive stress in the infected host cells (such as macrophages) affects mycobacterial survival and trafficking. We need to understand the molecular and cellular biology of TRS-induced changes in mycobacterial physiology, such as decrease in cell size, intracellular protein aggregation, cell aggregation, and development of biofilms. We still do not understand the underlying genetic circuit that leads to these effects. Furthermore, the molecular machinery that is involved in induction of persister phenotype in response to reductive stress needs to identified and characterized. We look forward to an exciting future that will provide answers to these pertinent questions.
Footnotes
Acknowledgments
We are thankful to Dr. Dibyendu Sarkar for scientific and language editing of the article. We are thankful to Mr. Rajkumar, CSIR-Institute of Microbial Technology, for the management of laboratory requirements.
Funding Information
A.K. is supported through the “Swarnajayanti Fellowship (Year 2016–2017) (DST/SJF/LSA-02/2016-17)” and “National Bioscience Award for Career Development 2017/2018 (BT/HRD-NBA-NWB/37/01/2018)” by the Department of Science and Technology and the Department of Biotechnology, respectively. P.S.M. and S.S. are supported by a Senior Research Fellowship and Junior Research Fellowship from the Council of Scientific and Industrial Research.