
Abstract
Aims:
Cytoglobin (CYGB) is a member of the mammalian globin family of respiratory proteins. Despite extensive research efforts, its physiological role remains largely unknown, but potential functions include reactive oxygen species (ROS) detoxification and signaling. Accumulating evidence suggests that ROS play a crucial role in podocyte detachment and apoptosis during diabetic kidney disease. This study aimed to explore the potential antioxidative renal role of CYGB both in vivo and in vitro.
Results:
Using a Cygb-deficient mouse model, we demonstrate a Cygb-dependent reduction in renal function, coinciding with a reduced number of podocytes. To specifically assess the putative antioxidative function of CYGB in podocytes, we first confirmed high endogenous CYGB expression levels in two human podocyte cell lines and subsequently generated short hairpin RNA-mediated stable CYGB knockdown podocyte models. CYGB-deficient podocytes displayed increased cell death and accumulation of ROS as assessed by 2′7′-dichlorodihydrofluorescein diacetate assays and the redox-sensitive probe roGFP2-Orp1. CYGB-deficient cells also exhibited an impaired cellular bioenergetic status. Consistently, analysis of the CYGB-dependent transcriptome identified dysregulation of multiple genes involved in redox balance, apoptosis, as well as in chronic kidney disease (CKD). Finally, genome-wide association studies and expression studies in nephropathy biopsies indicate an association of CYGB with CKD.
Innovation:
This study demonstrates a podocyte-related renal role of Cygb, confirms abundant CYGB expression in human podocyte cell lines, and describes for the first time an association between CYGB and CKD.
Conclusion:
Our results provide evidence for an antioxidative role of CYGB in podocytes.
Introduction
Cytoglobin (Cygb) belongs to the family of mammalian globins in addition to hemoglobin (Hb), myoglobin (Mb), neuroglobin (Ngb), and the recently identified androglobin (Adgb) (6). Most known globins fulfill respiratory functions, supplying the cell with adequate amounts of O2 for aerobic energy production via the respiratory chain in the mitochondria (72). In contrast to Mb and Hb, the more recently identified globins Cygb (5, 7), Ngb (5, 7), as well as Adgb (27) all display hexacoordination of the heme iron atom (i.e., bound by two amino acid residues of the globin fold). While no functional interpretation for this heme hexacoordination currently exists, it is thought to point at a role other than the classical oxygen delivery and supply of Hb and Mb. The tissue expression profile of mammalian Cygb has been extensively studied (25). Cygb is predominantly expressed in fibroblasts and related cell types, and also in distinct nerve cell populations. Its physiological function, however, remains unclear; potentially, Cygb may have a role in oxygen storage, NO metabolism, and ROS protection or signaling.
Several animal model studies assessed the functional role of Cygb. Singh et al. (60) suggested a major role of Cygb in muscle repair and regeneration as myogenic progenitor cells derived from a mouse model, in which Cygb was specifically knocked out in skeletal muscle, were severely deficient in their ability to form myotubes. Moreover, Thuy et al. (66) support the antioxidant role of Cygb using a global knockout mouse model, which displays age-dependent development of multiple organ abnormalities. Using the same Cygb-deficient model, Liu et al. (36) recently provided evidence for a role of Cygb as regulator of NO degradation and cardiovascular tone in the vascular wall, in line with their previous in vitro observations on the NO dioxygenase function of Cygb (37). In addition, Thuy et al. (64) and Yassin et al. (75) both reported an implication of Cygb in tumorigenesis using in vivo models.
Innovation
Cytoglobin (Cygb) represents the fourth recently identified globin in humans. The renal role of Cygb has not been extensively explored. Our findings indicate a previously undiscovered podocyte-related renal role of Cygb, confirm abundant Cygb expression in human podocyte cell lines, provide evidence for an antioxidative role of Cygb in podocytes, and describe for the first time an association between Cygb and chronic kidney disease.
The renal role of Cygb has been underexplored. Nakatani et al. (48) detected expression of Cygb in rat kidney, mainly in fibroblasts, and also in the glomerulus. Upon cyclosporine A-induced nephropathy, Cygb expression was upregulated in fibrotic lesions of the kidney, specifically in interstitial fibroblasts positive for renal fibroblast markers CD73 and αSMA. Thuy et al. (66) also observed substantially increased fibrosis in Cygb KO mice, mostly in the liver, and also in the kidney, suggesting an antifibrotic role of Cygb. Similarly, Mimura et al. (42) and Nishi et al. (49) used Cygb-overexpressing transgenic rats in disease models of remnant kidney and renal ischemia/reperfusion, respectively, to provide evidence for an antifibrotic role of Cygb in kidney fibroblasts, potentially via a reactive oxygen species (ROS) scavenging function. Despite these investigations, the pathophysiological role of Cygb in the kidney, particularly in cell types other than interstitial fibroblasts, remains poorly understood.
Glomerular visceral epithelial cells, namely podocytes, are highly specialized cells in the Bowman's capsule of the kidney that actively participate in the glomerular filtration process due to their foot processes and slit diaphragm. Podocyte injury is the primary cause of impaired glomerular filtration. Accumulating evidence exists that oxidative stress plays a key role in most pathogenic pathways leading to podocyte defects, for example, observed in diabetic nephropathy (DN) (19). Free radicals such as superoxide can induce cell and tissue injuries through lipid peroxidation, activation of NF-κB (23), production of peroxynitrite, and induction of apoptosis. Furthermore, ROS and other free radicals can directly induce cell injury. In vitro studies indicated that Cygb is able to scavenge free radicals, and overexpression of Cygb in renal immortalized fibroblasts (42, 49) as well as in various other cellular models preserves cell viability under conditions of oxidative stress (12, 26, 33, 61, 74), but the role of Cygb in podocytes remains unexplored.
In the current study, we used a Cygb KO model to investigate the renal function of Cygb and observed a podocyte-associated phenotype. Using cellular models of human podocytes for mechanistic studies, we demonstrated abundant CYGB expression in podocytes and investigated the antioxidative role of CYGB in human podocyte cell lines. Podocytes lacking CYGB displayed increased ROS accumulation and cell death, as well as altered expression of genes involved in the antioxidant defense system and apoptosis. Moreover, human renal biopsy expression data from chronic kidney disease (CKD) patients indicated an association between CYGB and advanced-stage renal disease.
Results
Cygb-deficient mice display reduced kidney function and reduced podocyte number
To analyze the renal expression pattern of Cygb, we performed immunofluorescence analysis on mouse kidney tissue. Immunofluorescence revealed that Cygb is expressed in interstitial fibroblasts as well as in the glomerulus (Fig. 1A). To assess a potential Cygb-dependent effect on kidney function, we used a previously established Cygb KO mouse model (65). Reduced Cygb expression in Cygb−/− mice was confirmed in whole-kidney mRNA and protein extracts (Fig. 1B, C) as well as on the immunofluorescence level (Fig. 1A). Compared with wild-type mice, Cygb−/− mice demonstrated a substantial reduction in renal function, as indicated by an increased serum fluorescein isothiocyanate (FITC)-sinistrin concentration and decreased clearance (Fig. 1D). However, analysis of plasma and urine electrolytes did not show significant differences between the two groups (Supplementary Fig. S1A, B). We also did not observe changes in marker genes of fibrosis and vasculature in whole-kidney RNA (Supplementary Fig. S2A). As we detected Cygb expression in the glomerulus, we next investigated a potential role of podocytes in this phenotype. The average number of podocytes per glomerular cross section was determined by counting cells stained with the podocyte-specific marker Wt-1 (20). Although no apparent difference in proteinuria (Supplementary Fig. S1F) nor clear gross morphological kidney changes were observed in the two groups (Supplementary Fig. S1G), podocyte numbers were significantly reduced in Cygb−/− mice compared with Cygb+/+ mice, as assessed by immunohistochemistry (Fig. 1E). Consistent with this observation, Wt-1 mRNA (Fig. 1F) as well as protein levels (Fig. 1G) were downregulated in Cygb−/− mice, confirming a decreased podocyte number.
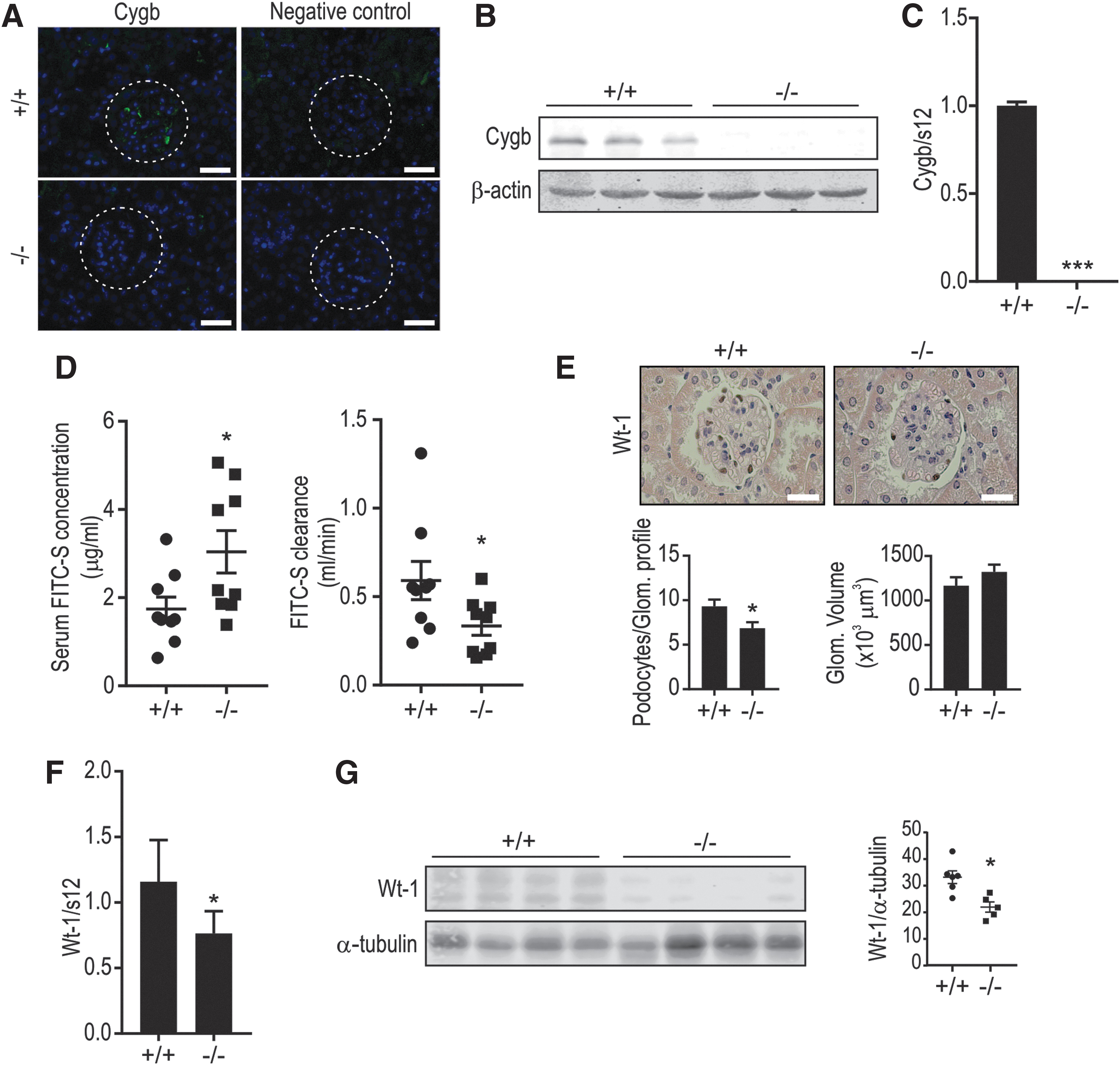
Abundant CYGB expression in podocyte cell models
To mechanistically investigate the role of CYGB in the glomerulus, we explored CYGB levels in human podocyte cell culture models. Compared with various human kidney-derived cell lines, including proximal tubule cells, fibroblasts, and embryonic cells, the most abundant endogenous CYGB mRNA and protein levels were observed in podocytes, particularly in AB8/13 (Fig. 2A), which was used for subsequent investigations. The heat-sensitive AB8/13 cells proliferate at the permissive temperature of 33°C, and stop proliferating and undergo differentiation within 2 weeks at the nonpermissive temperature of 37°C (57). Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) revealed that CYGB mRNA is differently expressed before and after podocyte differentiation (Fig. 2B), and the CYGB protein is localized in both the cytoplasm and nucleus, as assessed by immunoblotting (Fig. 2C).

We established stable AB8/13 knockdown cells using two independent short hairpin RNA (shRNA) sequences targeting CYGB, termed shCYGB-1 and shCYGB-2. CYGB knockdown efficiency was confirmed at the mRNA and protein level (Fig. 2D, E). To obtain an independent podocyte cell culture model, we also generated stably CYGB-depleted shRNA LY cells (Supplementary Fig. S3).
Antiapoptotic role of CYGB in AB8/13
A putative antiapoptotic role of CYGB has been suggested in some cell lines (60, 67), but not in podocytes. This prompted us to investigate cell viability in our cell models. CYGB knockdown podocyte cell lines displayed increased cell death (Fig. 3A), indicating a role of CYGB in cell survival. A similar reduction in cell viability was observed in the independently analyzed LY podocyte cell line (Supplementary Fig. S2). Consistently, we observed substantially increased cleaved-PARP1 levels in shCYGB-1 cells (Fig. 3B), but not in shCYGB-2, possibly due to its more moderate knockdown of CYGB. Moreover, MTT assays demonstrated that shCYGB-1 cells were more susceptible to H2O2 and antimycin A (AMA) compared with WT and shCTR cells (data not shown). Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assays revealed DNA double-strand breaks in shCYGB-1 and shCYGB-2 cells treated with H2O2 for 3 h, but not in shCTR cells (Fig. 3C).

Due to the intimate link between cell death and mitochondrial functions (ATP production and apoptosis), we investigated a potential CYGB-dependent effect on the bioenergetic status in AB8/13 cells. For this purpose, we analyzed mitochondrial respiration, measured as oxygen consumption rate (OCR). Cells were treated sequentially with oligomycin, carbonyl cyanide-4 trifluoromethoxy phenylhydrazone (FCCP), and rotenone/AMA to challenge the mitochondrial electron transport chain and evaluate possible differences in OCR response. CYGB downregulation was associated with decreased OCR already at baseline and also following addition of the various mitochondrial stressors (Fig. 3D). The lower OCR after oligomycin in shCYGB-1 cells is linked to lower ATP production compared with shCTR, consistent with a decreased mitochondrial activity. Subsequent FCCP treatment resulted in an increased maximal OCR in shCTR cells compared with CYGB knockdown cells. Until this stage, shCYGB-2 cells (characterized by a less pronounced knockdown of CYGB than shCYGB-1 cells, Fig. 2E) displayed an intermediate phenotype between shCTR and shCYGB-1. However, after the final treatment with rotenone and AMA, shCYGB-1 and -2 cells reached the same minimal OCR, while shCTR cells displayed higher levels, suggesting a higher nonmitochondrial respiration in control cells compared with CYGB knockdown podocytes (Fig. 3E). Overall, these data indicate that CYGB supports podocyte viability by preserving mitochondrial bioenergetics.
Antioxidative role of CYGB in AB8/13
To further investigate the putative antioxidative role of CYGB in podocytes, we analyzed expression levels of the redox-sensitive genes heme oxygenase 1 (HO-1) and HSP1A1. Basal mRNA levels of both HO-1 and HSP1A1 were increased in shCYGB-1 cells compared with WT and shCTR cells (Supplementary Fig. S4), indicating a CYGB-dependent altered intracellular redox state. Similarly, HO-1 expression levels were increased in CYGB-depleted LY podocytes (Supplementary Fig. S2). To further confirm this observation, WT, shCTR, and shCYGB AB8/13 cells were transiently transfected with the cytoplasmic oxidation-sensitive roGFP2-Orp1 probe. Compared with WT cells, shCYGB-1 and shCYGB-2 cells showed a significant increase in roGFP2-Orp1 oxidation under basal conditions (Fig. 4A), consistent with the gene expression data. Treatment with AMA, a mitochondrial complex III inhibitor, further increased roGFP2-Orp1 oxidation, particularly in the two shCYGB cell models (Fig. 4A). As an independent approach to measure ROS generation in podocytes, we used 2′7′-dichlorodihydrofluorescein diacetate (H2DCFDA), a fluorescent cell-permeable indicator of ROS. Immediately after AMA treatment, 2′,7′-dichlorofluorescein (DCF) fluorescence strongly increased in shCYGB-1 cells compared with WT and shCTR cells (Fig. 4B). To confirm the results with an independent ROS stimulus, cells were subjected to 250 μM H2O2, and DCF fluorescence was measured over a 4-h period. ROS accumulation rapidly increased in shCYGB-1 and shCYGB-2 cells, reaching the highest value after 4 h (Fig. 4C).

Since podocyte loss has been associated with hyperglycemia and subsequent increase in oxidative stress (63), cells were incubated with high glucose (HG) or mannitol (used as iso-osmotic control) for 5 days. Treatment with HG significantly increased ROS accumulation (Fig. 4D) and cell death (Fig. 4E) in shCYGB-1 and shCYGB-2 cells compared with WT cells, indicating a protective role of CYGB in oxidative stress-mediated cell death. On the contrary, overexpression of CYGB reduced ROS accumulation and oxidative stress as assessed by roGFP2-Orp1 oxidation and H2DCFDA-based assays upon different stimuli (Supplementary Fig. S5). Collectively, these data provide evidence for an antioxidative role of CYGB in podocytes.
CYGB deficiency alters expression of multiple genes involved in redox signaling and apoptosis
The CYGB-dependent transcriptome was determined by RNA sequencing of shCTR and shCYGB-1 cells. Three independent samples for each condition were analyzed and the knockdown efficiency of shCYGB-1 was confirmed to be >90% (Supplementary Fig. S6A). Most of the total reads mapped to exons (90%), whereas introns and intergenic regions accounted only for 9% and 1% of the total reads, respectively (Supplementary Fig. S6B), confirming the quality of the multiple mapping. Furthermore, sample-to-sample distance and principal component analysis (PCA) of the RNA-seq data sets indicate clear separation among shCYGB and shCTR cells (Supplementary Fig. S6C, D).
To ascertain the reliability of the sequenced samples, the transcriptomic data were compared with podocyte-specific marker genes based on literature search and single-cell RNA sequencing (scRNA-seq) (39). Genes were considered to be expressed in the shCTR and shCYGB AB8/13 data sets if the mean reads per kilobase per million mapped reads (RPKM) value were ≥0.1 and ≥0.5, respectively (39). In shCTR AB8/13 cells, 83%–95% of the literature-based (RPKM ≥0.5 = 68/82; RPKM ≥0.1 = 72/82) and 76%–82% of the scRNA-seq-based (RPKM ≥0.5 = 58/76; RPKM ≥0.1 = 62/76) essential podocyte genes were expressed (Supplementary Tables S1 and S2), thereby validating that AB8/13 cells represent a genuine podocyte model.
More than 1500 genes were found differentially expressed in shCYGB compared with shCTR cells (Fig. 5A), and among these, the majority (1148) was downregulated. The most significantly upregulated genes included ubiquitin D (UBD), UDP glucuronosyltransferase family 1 member A6 (UGT1A6), and neuropeptide Y receptor Y4 (NPY4R), whereas the most significantly downregulated genes included activity regulated cytoskeleton associated protein (ARC), heparan sulfate 6-O-sulfotransferase 2 (HS6ST2), and MAF BZIP transcription factor B (MAFB). To investigate if the observed differentially expressed genes are associated with common canonical pathways based on established interactions among them, an ingenuity pathway analysis (IPA) was carried out. Interestingly, differentially expressed genes could be attributed to multiple kidney failure-associated pathways (Supplementary Table S3). In addition, using the CLC Genomics Workbench 8.5.1 RT2 Profiler gene lists (Qiagen), we observed an association of the CYGB-dependent transcriptome with apoptosis and oxidative stress pathways (Supplementary Table S4). Using similar RT2 Profiler gene lists as well as Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and IPA tools, we also specifically looked at metabolism-associated genes, but no clear enrichments could be observed (data not shown). Furthermore, analysis of expression levels of candidate metabolism-associated genes in whole-kidney extracts of Cygb+/+ and Cygb−/− mice did not reveal any apparent differential regulation in vivo (Supplementary Fig. S1E).

In line with the increased oxidative stress and apoptosis of CYGB-depleted AB8/13 cells, several antioxidant genes such as DUOX-1, DUOX-2, and GPX-3, as well as some antiapoptotic genes, such as TP73, were found to be downregulated. Furthermore, multiple genes indicating podocyte injury, including SERPINE1 and CTGF, were upregulated (Table 1). To validate the most relevant differentially expressed genes, we performed RT-qPCR on independent RNA samples of shCYGB and shCTR AB8/13 cells (Fig. 5B). A similar CYGB-dependent regulation of DUOX-1, GPX-3, and SOD3 was found in LY cells, validating the results observed in AB8/31 (Fig. 5C). In contrast, a selection of redox-sensitive genes were not differentially regulated in whole-kidney lysates of Cygb+/+ and Cygb−/− mice (Supplementary Fig. S1F). Consistently, levels of protein carbonyls in whole-kidney lysates from Cygb+/+ and Cygb−/− mice were comparable, suggesting the absence of apparent differences in ROS levels in vivo (Supplementary Fig. S1G). Of particular interest is the downregulation of WT-1 and MAFB, both established regulators of podocyte development and viability in shCYGB AB8/13 cells versus shCTR cells. A similar, although less pronounced, downregulation of WT-1 and MAFB could be observed in stably CYGB-depleted LY cells (Fig. 5C). To confirm this CYGB-dependent regulation on protein level, we carried out immunoblotting for both proteins. Immunoblotting experiments clearly recapitulated the obtained RNA results, illustrating reduced protein expression of WT-1 and MAFB in CYGB-deficient AB8/13 cells compared with shCTR cells (Fig. 5D). A comparable trend could be observed in CYGB-deficient LY cells (Fig. 5E). Finally, we also evaluated MafB expression levels in whole-kidney lysates of Cygb+/+ and Cygb−/− mice, demonstrating reduced renal MafB mRNA and protein levels in Cygb-deficient mice (Supplementary Fig. S2B, C). Collectively, analysis of the CYGB-dependent transcriptome in AB8/13 cells revealed altered expression of multiple genes involved in redox signaling and apoptosis as well as essential factors of podocyte biology, consistent with the CYGB-dependent phenotype.
List of Differentially Expressed Genes in shCYGB-1 Compared with shCTR Cells
Genes are divided in four categories: antioxidant, podocyte phenotype, podocyte injury, and apoptosis. p-Values are corrected for multiple statistical testing by FDR.
FDR, false discovery rate.
CYGB is associated with CKD
Genome-wide association studies (GWAS) have proven to be a powerful tool in identifying novel genetic markers involved in CKD (50). By using GWAS databases (3), we identified a single-nucleotide polymorphism (SNP), located in an intergenic region 3′ of the CYGB gene associated with albuminuria (Fig. 6A). University of California Santa Cruz-integrated ENCODE data (10) illustrate that the SNP (rs8082416) is localized in a DNaseI hypersensitivity cluster observed in 115 cell types that reflects open chromatin, and overlaps with strong transcription factor occupancy (Supplementary Fig. S7A). Chromatin immunoprecipitation-sequencing data sets further revealed histone marks for an active enhancer (H3K4Me1) and basal components of the transcriptional machinery (RNA pol2 in cell lines SK-N-MC, IMR90, and HCT-116; as well as p300 in H1-hESC) in this region. In addition, RNA polymerase II-associated chromatin interaction analysis by paired-end tag (ChIA-PET) sequencing data in MCF-7 cells from ENCODE suggested DNA looping of this potential enhancer region to the first intron of the CYGB gene, close to its transcription initiation site. To experimentally validate a putative synergistic action between this potential 3′-enhancer and the CYGB promoter, we performed luciferase reporter gene assays. Whereas a 90 bp fragment of the 3′-enhancer spanning the SNP did not regulate heterologous SV40 promoter-driven luciferase activity, the 3′-enhancer substantially increased endogenous CYGB promoter-driven luciferase activity (Supplementary Fig. S7B), confirming a cooperation between the downstream enhancer and the CYGB promoter. However, the SNP site did not directly alter the transcriptional activity of the CYGB gene under basal conditions as no change in luciferase activity could be observed between the WT and mutated 3′-enhancer-driven construct (Supplementary Fig. S7B).
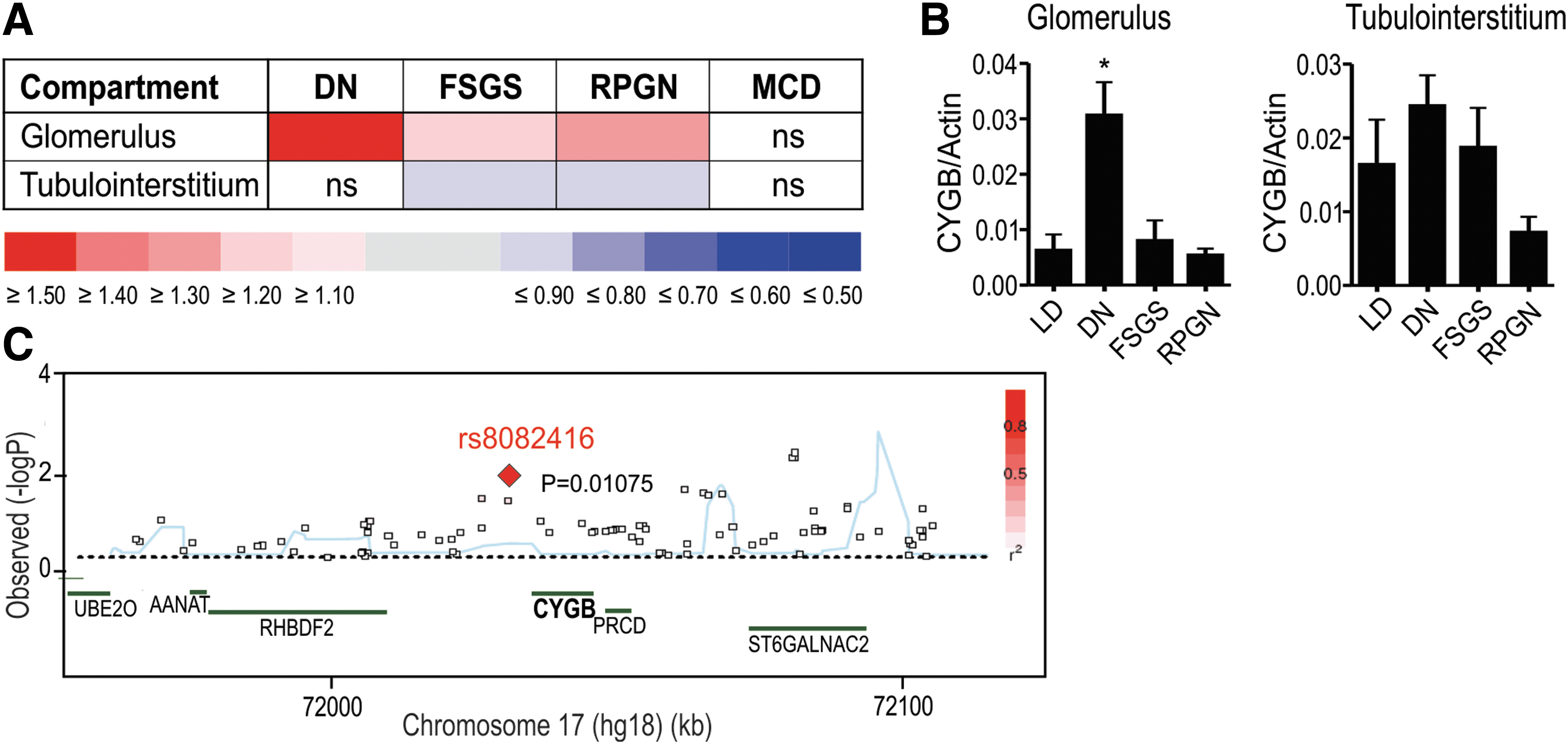
As an independent approach, existing gene array data from human renal biopsy specimens collected from the European Renal cDNA Bank (8) were analyzed. Data were obtained from Affymetrix HG-U133 Plus 2.0 microarrays, hybridized with glomerular and tubulointerstitial complementary DNA (cDNA) procured from different nephropathies as well as pretransplant biopsies from living renal allograft donors as controls. These biopsy expression studies from CKD patients revealed a consistent association of CYGB expression levels with advanced end-stage CKD, including DN (Fig. 6B). CYGB mRNA levels in different kidney diseases were validated by RT-qPCR in an independent patient cohort, supporting the association of increased CYGB mRNA levels with DN (Fig. 6C). In summary, these data suggest an association of CYGB with CKD.
Discussion
Based on in vivo and in vitro data, we propose that Cygb has a previously underappreciated role in podocyte function. Our study demonstrates that Cygb is expressed in mouse glomeruli, is important to preserve basal renal function possibly via a podocyte-related role, and can protect podocytes from oxidative stress and cell death under various experimental conditions in vitro. Specifically, CYGB-deficient podocytes display increased ROS accumulation and apoptosis, as well as dysregulation of multiple genes involved in cellular redox balance, apoptosis, and podocyte integrity. In addition, we provide evidence for an association of CYGB with CKD using patient biopsies.
Whereas the antioxidative function of CYGB has reached a consensus in the globin field, the precise molecular mechanism underlying this function is still poorly understood. Most likely CYGB may scavenge ROS using heme and thiol residues (13, 51), while peroxidase and superoxide dismutase activity has been excluded by the investigations of Trandafir et al. (68). In an attempt to investigate the molecular mechanism in more depth, we explored the function of CYGB in various renal cell models. To the best of our knowledge, a role of CYGB in podocytes has not been investigated so far. Consistent with the RNA-sequencing-based results of microdissected renal tubules from Lee et al. (32), the studies by Geuens et al. (18), and the recent results of Balkawade et al. (1), we detected high levels of Cygb protein in mouse glomeruli (Fig. 1A). Cygb-deficient mice displayed a reduced number of podocytes (Fig. 1E). Consistently, renal function, as measured by FITC-sinistrin clearance, was reduced in Cygb-deficient mice (Fig. 1D). Mechanistic studies in two cellular models of podocytes, which express more abundant endogenous CYGB levels compared with any other kidney-derived cell lines (Fig. 2A), validate an antioxidative function of CYGB. CYGB depletion, using independent shRNA targeting sequences (Fig. 2D, E), induced increased cell death as measured by trypan blue exclusion (Fig. 3A) and cleaved PARP (Fig. 3B), an established apoptosis marker as well as DNA damage (Fig. 3C), corroborating with the previously observed antiapoptotic role of CYGB (14, 31, 60, 61). Using complementary methods, we show that podocytes with attenuated CYGB expression display increased oxidative stress (Fig. 4A–E). In line with this observation, analysis of cellular bioenergetics suggest impaired mitochondrial function in CYGB depleted cells as evidenced by reduced mitochondrial respiratory capacity (Fig. 3D, E). An NO dioxygenase function has been suggested for CYGB (17, 24). As AB8/13 cells only express very low levels of NO synthases, an NO-mediated inhibiting effect of basal OCR, following knockdown of CYGB, is very unlikely and rather argues for an ROS-dependent effect of altered mitochondrial respiratory capacity. CYGB overexpression on the contrary reduced oxidative stress, prevented ROS accumulation in AMA-treated podocytes, and promoted cell viability, in line with previous findings (14, 26, 74). Podocyte apoptosis has been demonstrated as a cause of podocyte depletion (30). Similar to the phenotype of reduced number of podocytes observed in our Cygb−/− mice, multiple pathologies such as DN have been associated with loss of podocytes. Specifically, in DN, hyperglycemia-induced ROS accumulation was shown to contribute to podocyte apoptosis and depletion (35, 63). Upon treatment with HG, CYGB-deficient podocyte cells displayed increased ROS production and more pronounced cell death, compared with controls, strongly suggesting a protective role of CYGB. In contrast to our expectations, we did not observe albuminuria or proteinuria in Cygb−/− mice. Accordingly, MacIsaac et al. reported that the absence of proteinuria does not preclude loss of renal function (41). Alternatively, this lack might be due to the mild nature of the Cygb phenotype that, despite decreased podocyte number, is able to maintain filter barrier integrity or, in case it is not, may rely on enhanced proximal tubule endocytic activity resulting in compensating proximal tubular protein reabsorption. In support of the latter hypothesis, increased megalin expression has recently been described in type 2 diabetes (4). Future studies are required to study if challenging conditions such as induction of DN would lead to proteinuria in Cygb−/− mice. According to the study of Susztak et al. (63), hyperglycemia-induced overproduction of ROS could cause a diminution of the number of podocytes, which could contribute to the development of DN. Although CYGB knockout was sufficient to induce some histological changes such as podocyte loss, it was not sufficient to induce other histological changes as well as functional changes in the glomerulus (albuminuria). These changes may become apparent upon exposure to diabetic conditions and suggest a certain threshold of podocyte injury, which is exacerbated under diabetic conditions (2, 53). On the contrary, we cannot exclude a developmental defect in the Cygb−/− mice.
Transcriptome analysis of CYGB-deficient podocytes and control cells revealed dysregulation of multiple genes involved in redox balance, apoptosis, podocyte function, and podocyte injury (Fig. 5A, B). Most of the genes were downregulated, indicating a general transcriptional repression in the absence of CYGB. As this could potentially hint to a decreased metabolic function, we performed a close inspection of metabolism-associated genes, but only could observe a few moderate changes in gene expression, insufficient for explaining a decreased metabolic function. Similarly, a decrease in general mRNA stability upon lowered CYGB expression cannot be ruled out.
Due to its homology to Mb and comparable O2-binding affinity, CYGB was originally thought to contribute to intracellular O2 supply (69), acting as an O2 reservoir or as a signal transducer in O2 sensing pathways (18, 25). Therefore, CYGB deficiency would decrease oxygen availability and ROS signaling. DUOX-1 and DUOX-2 belong to the NADPH oxidase family and need oxygen as cosubstrate to produce ROS (i.e., H2O2 or O2 −). Both DUOX-1 and DUOX-2 are downregulated in lung cancer (40) and in hepatocellular carcinoma (DUOX-1) (34), in conditions were increased ROS levels were observed. However, their role in podocytes is still unknown. Downregulation of antioxidative genes, including CYGB, SOD3, GPX3, and GPX7, could explain the accumulation of ROS, as previously reported (60). The absence of differential expression of these antioxidative genes in whole-kidney lysates of Cygb+/+ and Cygb−/− mice might be explained by compensation of multiple other surrounding cell types.
Intriguingly, attenuated CYGB expression leads to downregulation of WT-1 and MAFB, essential podocyte transcription factors involved in maintaining cell differentiation (Fig. 5C–E). In line with these in vitro data, both Wt-1 and MafB were downregulated in Cygb−/− mice. WT-1 represents a key regulator of podocyte function and its downregulation leads to glomerulonephritis and mesangial sclerosis (20). Various mutations in WT-1 cause podocytopathies, including Denys–Drash syndrome, Frasier syndrome, and nonsyndromic focal segmental glomerulosclerosis (FSGS) (43). Whereas major efforts have focused on the identification of transcriptional targets of WT-1 in podocytes, the finding of an upstream regulator is of substantial interest as well. Moreover, WT-1 promotes expression of a large subset of the podocyte proteome, probably by activating the transcription of other transcriptional regulators that act either cooperatively with or independently of WT-1 (43). Studies by Dong et al. (11) provided evidence that WT-1 represents a crucial transcription factor for podocyte maturation and maintenance and fulfills its function by regulation of various target genes, including Nphs2, Mafb, and Magi2. In line with the differential regulation of WT-1 in CYGB-depleted podocytes as well as in Cygb−/− mice, we found that these target genes are CYGB dependently regulated in podocytes. Particularly, MAFB, a WT-1 key target gene, has been shown as well to be essential for kidney development (45). Furthermore, its overexpression in podocytes protects against DN, through the regulation of slit diaphragm proteins, antioxidative enzymes, and the Notch pathway (46). In addition, LMX1B, a transcription factor crucial for proper differentiation of podocyte precursors, was shown to be downregulated in CYGB-deficient podocytes. Overall, our RNA-seq-based transcriptome data in an AB8/13 cell model illustrate that CYGB influences multiple crucial transcription factors in podocyte biology, indicate its potential relevance to podocytopathies, and suggest that CYGB might therefore represent a potential therapeutic target.
Using GWAS, we discovered an SNP in the 3′ intergenic region of the CYGB gene, potentially associated with increased albuminuria (Fig. 6A). The SNP coincides with a region displaying several hallmarks of an enhancer and loops back to the first intron of CYGB gene close to the transcription initiation site as indicated by ChIA-PET data. Consistent with the chromatin looping data, reporter gene assays provide evidence for a cooperation between the distal downstream 3′-enhancer and the proximal CYGB promoter in regulating CYGB expression. Although additional studies under experimental conditions would be required to validate a potential direct effect of the SNP on CYGB gene regulation, these experiments indicate a further correlation of the SNP and the CYGB gene locus. Intriguingly, this is the first time that a globin gene was associated with CKD. Simultaneously, the potential role of CYGB in acquired human renal disease was found in an unbiased way by analyzing transcriptomic data of patients with different stages of renal failure (Fig. 6B, C).
In conclusion, our study demonstrates that CYGB (i) is abundantly expressed in human podocyte cell lines, (ii) protects podocytes from oxidative stress and apoptosis, (iii) is required to preserve renal function possibly via a podocyte-associated protective role, and (iv) is implicated in CKD.
Materials and Methods
Reagents
AMA (Sigma-Aldrich, St. Louis, MO) was dissolved in 95% ethanol. H2O2 was purchased from Sigma-Aldrich. H2DCFDA (10 μM; Invitrogen, Thermo Fischer Scientific, Waltham, MA) was dissolved in dimethyl sulfoxide (DMSO). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT; Sigma-Aldrich) for MTT assays was dissolved in phosphate-buffered saline (PBS). For the analysis of human renal biopsies, TaqMan reagents for human CYGB (NM_134268) were purchased from Applied Biosystems (Waltham, MA).
Renal function analysis and tissue sampling of Cygb-deficient mice
C57BL/6 Cygb global knockout mice were generated as described previously (65). To evaluate whether there was a difference in baseline renal function compared with wild-type C57BL/6, glomerular filtration rate (GFR) was determined by measuring FITC-sinistrin clearance. FITC-sinistrin is a fluorescent inulin analog, which is only excreted via glomerular filtration (52). For these experiments, nine male Cygb+/+ and nine male Cygb−/− mice (age 18 weeks) were implanted with two osmotic minipumps (model 1002, release rate of 0.25 μL/h for 14 days; ALZET, Charles River, Beerse, Belgium) filled with 100 mg/mL FITC-sinistrin (Fresenius Kabi Austria GmbH, Linz, Austria), 2 weeks before sacrifice. Mice were anesthetized via intraperitoneal injection with a mixture of ketamine (80 mg/kg) and xylazine (16 mg/kg) in 0.9% NaCl and kept on a heating pad (37°C) during surgery. After midline depilation with Veet cream and skin sterilization with ethanol, a ventrolateral incision (0.5 cm) was made and pumps were placed into the peritoneal cavity. The incision was closed with sterile suture, muscle and skin separately. Mice regained consciousness and mobility under partial exposure to a heating lamp, after which they were put into their housing cages. The two-pump strategy was based on the report of Qi et al. assuring sufficient serum FITC-sinistrin levels for renal function assessment (54). For final GFR calculation, we applied the equation “FITC-sinistrin clearance = FITC-sinistrin pump infusion rate/steady-state blood FITC-sinistrin concentration” (54).
Twenty-four hours before sacrifice, mice were put in metabolic cages with free access to water and food for 24-h urine collection. Urine was stored at −20°C until further analyses. At sacrifice, mice were deeply anesthetized and exsanguinated via the vena cava inferior for blood collection. Serum was extracted and kept at −20°C for further analyses. Kidneys were isolated, decapsulated, weighed, sliced in 1 mm transverse sections, and snap frozen in liquid nitrogen or fixed in neutral buffered formalin. Urine and serum parameters, Na+ and K+ concentrations, were determined using an electrolyte analyzer (IL-943; Instrumentation Laboratories, Bedford, MA). Osmolality was measured using the Fiske One-Ten osmometer (Fiske, Norwood, MA). Albuminuria was assessed by loading 5 μL of urine sample on a polyacrylamide gel and bovine serum albumin was used as positive control.
All experimental procedures were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the University of Antwerp Ethics Committee (permit 2016-47).
Immunohistochemistry
Paraffin-embedded kidney sections were deparaffinized, hydrated, blocked, and incubated overnight with anti-Wt-1 (ab89901; Abcam, Cambridge, United Kingdom). After washing, sections were incubated with a biotinylated goat anti-rabbit immunoglobulin G antibody (Vector Laboratories, Burlingame, CA) and subsequently incubated with avidin and biotinylated horseradish peroxidase (Vector Laboratories). A dark brown color was developed with diaminobenzidine in the presence of 3% H2O2. Sections were counterstained with hematoxylin and eosin and mounted in Eukitt (Sigma-Aldrich). Podocyte counts were assessed by staining kidney sections for Wt-1 and quantification was performed in a blinded manner. Glomerular diameters of 30 glomeruli/section were measured from five mice per experimental group using ImageJ. Glomerular volume was determined as Gv = β/k · (π · r 2)3/2, where β = 1.38 is the shape coefficient for spheres, k = 1.1 is the size distribution coefficient, and (π · r 2) is the glomerular area (58). For immunofluorescence experiments, Cygb staining on Cygb+/+ and Cygb−/− kidneys was performed using anti-Cygb (D-7, sc365246; Santa Cruz Biotechnology, Dallas, TX) and the Vector mouse on mouse immunodetection kit, fluorescein (FMK-2201; Vector Laboratories) according to the manufacturer's instructions. Subsequently, sections were mounted with 4′,6-diamidino-2-phenylindole Fluoromount-G (SouthernBiotech, Birmingham, AL) and visualized using fluorescence microscopy.
Cell culture and treatments
Conditionally immortalized human podocyte cell lines AB8/13 and LY were a kind gift from A. Kistler (57). Cells were cultured in Roswell Park Memorial Institute (RPMI) (Sigma-Aldrich) supplemented with 10% heat-inactivated fetal calf serum (FCS; Gibco, Thermo Fischer Scientific), 50 IU/mL penicillin, 50 μg/mL streptomycin (Invitrogen), 5 μg/mL insulin, 5 μg/mL transferrin, and 5 ng/mL sodium selenite (ITS; Roche, Mannheim, Germany). AB8/13 cells were propagated at 33°C and differentiated for 10–14 days at 37°C in a humidified incubator containing 5% CO2 (Binder, Tuttlingen, Germany). Cell death was measured by the Vi-Cell XR 2.03 Cell Viability Analyzer (Beckman Coulter, Krefeld, Germany) and TC20 automated cell counter (BioRad) using the trypan blue dye exclusion method. HPRT, HEK293T, TK-173, TK-188, and TZ-1 cells were cultured in HG Dulbecco's Modified Eagle Medium (DMEM) (Sigma-Aldrich) supplemented with 10% heat-inactivated FCS, 50 IU/mL penicillin, and 50 μg/mL streptomycin (Invitrogen). HK-2 were cultured in DMEM/F12 (Sigma-Aldrich) supplemented with 10% heat-inactivated FCS, 100 U/mL penicillin, 100 μg/mL streptomycin, 36 ng/mL hydrocortisone, 5 μg/mL insulin, 5 μg/mL transferrin, and 5 ng/mL selenium (ITS) solution (Roche).
mRNA and protein detection and quantification
Total cellular RNA was extracted as previously described (29). Total RNA (2 μg) was reverse transcribed (RT) using the Prime Script RT reagent kit (Takara Bio) and cDNA levels were estimated by qPCR using the primers listed in Supplementary Table S5 and a SYBRGreen qPCR reagent kit (Sigma-Aldrich) in a CFX96 C1000 Thermal Cycler (BioRad). Transcript levels were calculated as described before (28) and displayed as fold change, if not otherwise indicated.
Immunoblotting, signal imaging, and quantification were performed as previously reported (47). Membranes were probed with antibodies against CYGB (13317-AP; Proteintech, Rosemont, IL; or EPR13198; Abcam), synaptopodin (SYNPO) (P-19, sc-21537; Santa Cruz Biotechnology), PARP1 ([E102], ab32138; Abcam), cleaved PARP1 ([E51], ab32064; Abcam), TATA-box binding protein (8515; Cell Signaling Technology, Leiden, The Netherlands), β-actin (A5441; Sigma-Aldrich) and α-tubulin (TU-02, sc-8035; Santa Cruz Biotechnology). Signals from horseradish peroxidase (HRP)-coupled secondary antibodies were detected with ECL substrate (Pierce, Thermo Fisher Scientific) using a luminescent image analyzer (Fusion FX6; Vilber Lourmat, Marne la Vallée, France). Uncropped immunoblots are provided in Supplementary Figure S8. Protein oxidation was detected by the OxyBlot method following the manufacturer's instructions. Briefly, each specimen was divided in two aliquots: one aliquot was subjected to derivatization reaction (2,4-dinitrophenylhydrazine solution), while the other one served as a negative control (derivatization-control solution). The treated samples and the corresponding negative control were loaded into a polyacrylamide gel. After gel electrophoresis, transfer, and blocking, the membrane was incubated with a primary antibody specific to the dinitrophenyl moiety of the proteins, HRP-coupled secondary antibody, and finally exposed on autoradiography film.
Generation of stable knockdown and overexpression cell lines
Expression vectors encoding shRNA sequences targeting human CYGB in a pLKO.1-puro plasmid were purchased from Sigma-Aldrich (shCYGB-1: order number TRCN0000059378; shCYGB-2: order number TRCN0000059381). Control cells (shCTR) were transfected with a nontargeting control shRNA under the control of a U6 promoter in a pKLO.1 puromycin resistance vector (Sigma-Aldrich) as described previously (16). Viral particles were produced in HEK293T cells by cotransfection of the respective transfer vector (3 μg) with the packaging plasmids pLP1 (4.2 μg), pLP2 (2 μg), and pVSV-G (2.8 μg; all from Invitrogen) using polyethylenimine transfection as described before (15). Cells were transduced with lentiviral-pseudotyped particles, and cell pools were cultured with the appropriate antibiotic for selection. For stable overexpression, full-length human CYGB gene and control gene β-glucuronidase (GUS) were cloned into a pLENTI6 plasmid. Viral particles were produced as described above.
H2DCFDA assay
AB8/13 cells were seeded in 96-well plates at 80% confluency and incubated with 10 μM H2DCFDA for 30 min in the dark. Fluorescence was measured using a 96-well fluorometer (Infinite 200Pro; Tecan, Männedorf, Switzerland). Results from kinetic measurements were calculated as described previously (21). Analysis was performed using the internal software i9 control.
roGFP2-Orp1 measurements
AB8/13 cells were grown on glass-bottomed dishes (Ibidi GmbH, Martinsried, Germany) and transduced with the H2O2-sensitive probe roGFP2-Orp1 (22, 44), which was subcloned into the pAd/CMV/V5-DEST adenoviral vector (Invitrogen) using the Gateway technology. After 48 h, the oxidation of the sensor was measured at basal level or upon AMA treatment (50 μg/mL, 30 min). The live measurements of roGFP2 oxidation and calculations of the degree of oxidation were performed as previously described (70).
Cell viability assay
For the MTT assay, AB8/13 cells were seeded in 96-well flat-bottomed plates and then exposed to AMA or H2O2 at the indicated concentrations (final volume 0.1 mL per well). After 6 h, 10 μL of 5 mg/mL MTT solution in PBS was added for 2 h. Following removal of the medium, 100 μL of DMSO was added to dissolve the formazan crystals. The absorbance at 540 nm was determined in triplicate using a plate reader (Infinite 200Pro; Tecan) and normalized by nontreated cells.
TUNEL assay
Cells were seeded on coverslips and treated with 250 μM H2O2 for 3 h. TUNEL of nuclei was performed using the APO-BrdU in situ DNA fragmentation assay kit (K401-60; Biovision, Milpitas, CA) following the manufacturer's protocol.
Determination of cellular bioenergetics
Differentiated human podocytes were seeded on XFe24-well microplates (Agilent Technologies, Santa Clara, CA) at 1.5 × 104 cells per well and incubated at 37°C and 5% CO2 the day before the experiment. The medium was then replaced by RPMI 1640 without bicarbonate, and cells were kept in an incubator without CO2 before transfer to a Seahorse analyzer XFe24 (Agilent Technologies), to measure the OCR. Three measurements of OCR were performed at steady state and after sequential injection of 1 μM oligomycin, 2 μM FCCP, and a mix of 0.5 μM rotenone and 0.5 μM AMA. During the experiment, the cells were kept in a humidified chamber at 37°C, under normal oxygen conditions. At the end of the assay, total protein per well was measured using the Bradford reagent (BioRad, Hercules, CA) and OCR values were normalized to the protein amount.
Analysis of human renal biopsies
Human renal biopsy specimens and Affymetrix microarray expression data (HG-U133 Plus2.0 Array; Affymetrix, Santa Clara, CA) were obtained within the framework of the European Renal cDNA Bank–Kröner-Fresenius Biopsy Bank (8). Diagnostic renal biopsies were obtained from patients after informed consent and with approval of the local ethics committees. Following renal biopsy, the tissue was transferred to RNase inhibitor and microdissected into glomerular (Glom) and tubulointerstitial (Tub) compartments. The microarray expression data used in this study came from individual patients with DN (Glom [n = 7], Tub [n = 7]), FSGS (Glom [n = 16], Tub [n = 7]), rapidly progressive glomerulonephritis (RPGN, Glom [n = 23], Tub [n = 21]), minimal change disease (Glom [n = 5], Tub [n = 5]), as well as pretransplant biopsies from living renal allograft donors as controls (living donors [LD], Glom [n = 18], Tub [n = 18]). Total RNA was isolated from microdissected glomeruli and tubulointerstitium, reverse transcribed, and linearly amplified according to a protocol previously reported (9). Fragmentation, hybridization, staining, and imaging were performed following the Affymetrix expression analysis technical manual (Affymetrix). The raw data were normalized using Robust Multichip Algorithm and annotated by Human Entrez Gene custom CDF annotation version 18. To identify differentially expressed genes, the Significance Analysis of Microarrays method was applied using TiGR (MeV, Version 4.8.1) (71). A q-value below 5% was considered to be statistically significant. RT-qPCR validation of renal biopsies was performed as reported earlier (8, 59). Predeveloped TaqMan reagents were used for human CYGB (NM_134268), and transcript levels were normalized to β-actin levels (LD, Glom [n = 9], Tub [n = 5], DN, Glom [n = 12], Tub [n = 15], FSGS Glom [n = 16], Tub [n = 17]), RPGN Glom [n = 9], Tub [n = 9]).
Genome-wide association study
Genetic association testing for urinary albumin-to-creatinine ratio, and meta-analysis was performed in the CKDGen and CARe cohorts of European ancestry, with further follow-up genetic analysis of significant SNPs in CARe cohorts of African American ancestry and in the Diabetes Control and Complications Trial/Epidemiology of Diabetes Interventions and Complications Study, as previously described (3).
RNA-seq library preparation and transcriptome sequencing
Total RNA from three independent samples of shCTR and shCYGB-1 AB8/13 cells was extracted using the RNeasy mini kit according to the manufacturer's instructions (Qiagen, Hilden, Germany). Before library construction, RNA quality was assessed using an Agilent 2100 Bioanalyzer and the Agilent RNA 6000 Nano Kit (Agilent Technologies). RNA integrity number values ranged from 8.5 to 9.7, indicating high-quality RNA samples. RNA was quantified using the Qubit RNA HS Assay Kit (Invitrogen). Libraries were prepared starting from 800 ng of total RNA using the TruSeq Stranded mRNA HT Sample Prep Kit (Illumina), including a poly-A selection step following the manufacturer's instructions and sequenced as 2 × 150 nt paired-end reads using an Illumina NextSeq 500™. Library preparation and sequencing were performed by StarSEQ®-GmbH (Mainz, Germany). RNA-Seq data are available at the European Nucleotide Archive under accession number PRJEB30641.
Differential gene expression, GO term annotation, and pathway enrichment analyses
Raw sequences were preprocessed to remove low-quality reads and residual Illumina adapter sequences using FASTX-Toolkit 0.0.14. The overall sequencing quality and the absence of adapter contamination were evaluated with FastQC 0.11.2. Preprocessed reads were mapped against the annotated human genome version hg38 with the RNA-seq algorithm of CLC Genomics Workbench 8.5.1 using the following parameters: mismatch cost = 2, insertion cost = 3, deletion cost = 3, length fraction = 0.95, and similarity fraction = 0.95. The mapping step was performed allowing either one (single) or up to 10 (multiple) mappings for each read, and all the downstream statistical and bioinformatic analyses were computed for both mapping outputs. As the results of the downstream analyses for single or multiple mapping converged (data not shown), only the results obtained with multiple mapping (the library of 2 × 150 nt paired-end stranded reads minimizes the occurrence of unspecific read mappings) were included. A summary of the mapping statistics is shown in Supplementary Table S6. PCA was performed and visualized using the plotPCA function of DESeq2 (38) (ntop = 1000) and ggplot2 (73) R packages (Supplementary Fig. 6C, D). Sample-to-sample distance was calculated from the transformed count matrices and visualized as heatmap using the Pheatmap R package. Both approaches indicated a clear separation among the shCTR- and shCYGB-derived data sets. In particular, PC1 accounted for 87% of the variance.
Differentially expressed genes were determined using the EdgeR-based [Empirical analysis of differential gene expression; (55, 56)] statistical tool of CLC Genomics Workbench using default parameters. Genes were considered differentially expressed when presenting |fold change| >2 and false discovery rate-corrected p-value ≤0.05. GO term and KEGG pathway enrichment analyses were performed using WebGestalt (version 2017) using the overrepresentation enrichment analysis method, requiring a Benjamini–Hochberg (BH)-corrected p-value ≤0.05 and a minimum enrichment of four genes for term/pathway. Enrichment in canonical pathways and Tox functions were performed with Qiagen's IPA (Qiagen) Core analysis tool using bias-corrected z-score (when applicable) and BH-corrected p-values ≤0.05.
Luciferase reporter gene assays
A 90 bp oligonucleotide encompassing the rs8082416 SNP was cloned into pGL3prom (Promega) between the MluI and XhoI restriction sites. The CYGB promoter was amplified by PCR from genomic DNA of HEK293T cells and cloned into pGL3basic (Promega). If not otherwise indicated, 3 × 105 HEK293T cells were transiently transfected with 500 ng reporter plasmid in a six-well format using Roti-Fect (Carl Roth, Karlsruhe, Germany). To control for differences in transfection efficiency and extract preparation, 5 ng pRL-SV40 Renilla luciferase reporter vector (Promega) was cotransfected. Cultures were evenly split onto 24-well plates 24 h after transfection. Luciferase activities of triplicate wells were determined using the dual-luciferase reporter assay system (Promega) as described before (62). Reporter activities were expressed as relative firefly/Renilla luciferase activities. All reporter gene assays were performed at least three times independently.
Statistical analyses
If not otherwise indicated, results are presented as mean values ± standard errors of the mean of at least three independent experiments. Statistical analyses were performed using Student's t-test and one-way analysis of variance (ANOVA) or two-way ANOVA, where appropriate. p-Values <0.05 were considered statistically significant. For the human biopsy data, statistical analysis was performed using Kruskal–Wallis and Mann–Whitney tests (SPSS 24.0; SPSS, Inc., Chicago, IL).
Footnotes
Acknowledgments
We thank Gerhard Müller and Michael Zeisberg for providing TK173, TK188, and TZ-1 cells, Moin Saleem for providing two podocyte cell lines, Caroline Fox, Ming-Huei Chen, and the CKDGen Consortium for interrogating urinary albumin-to-creatinine ratio data sets, Patrick D'Haese and Jean-Pierre Montani for advice, Julia Birk and Marcel Saarbeck for cloning assistance, Joey De Backer as well as all Hoogewijs laboratory members for advice and technical support. We also thank the participating centers of the European Renal cDNA Bank-Kröner-Fresenius biopsy bank (ERCB-KFB) and their patients for their cooperation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Swiss National Science Foundation (grant 173000) and the German Research Foundation (grant HO5837/1-1) to D.H., a University Research Priority Program “Integrative Human Physiology” grant to E.B.R., the Biobank ERCB-KFB (Else Kröner-Fresenius-Foundation) to C.D.C., an intramural grant (Center for Computer Sciences, JGU Mainz) to T.H., the International PhD Program (IMB Mainz) to E.P. and T.H., a Grant-in-Aid for Scientific Research from JSPS (No. 25293177 and No. 16H05290) and a Grant for Research Program on Hepatitis from the Japan Agency for Medical Research and Development (18fk0210004h0003) to N.K., and the NCCR
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.