
Abstract
Significance:
Bioactive sulfur species such as hydrogen sulfide (H2S), persulfide species (R-SnSH, n ≥ 1), hydrogen polysulfide (H2Sn, n ≥ 2), sulfur dioxide (SO2), and carbon disulfide (CS2) participate in various physiological and/or pathological pathways such as vasodilation, apoptosis, inflammation, and energy metabolism regulation. The oxidation state of the individual sulfur species endows them unique biological activities.
Recent Advances:
There have been great strides made in achieving molecular understanding of the sulfur-signaling processes.
Critical Issues:
The development of various chemical tools that deliver reactive sulfur species in a controllable manner has played an important role in understanding the different roles of various sulfur species. In this review, we focus on three types of sulfur species, including persulfide, SO2, and CS2. Starting with a brief introduction of their physiological functions, we will then assess the various drug delivery strategies to generate persulfide species, SO2, and CS2 as research tools and potentially as therapeutic agents.
Future Directions:
Development of donors of various sulfur species that respond to distinct stimulus is critical for this field. Another key to the long-term success of this field is the identification of an area of unmet medical need that can be addressed with these sulfur species.
Introduction
Sulfur signaling is implicated in a large number of physiological and pathological processes (27, 34, 40 –43, 64, 68, 69, 73, 92, 93). Early on, attention in the field of sulfur signaling was largely focused on hydrogen sulfide (H2S), which is often referred to as the third gasotransmitter with some disagreement (85, 89). Gasotransmitter is generally defined as endogenously produced gaseous signaling molecules, although with some debate (62). Previously, donors of various sulfur species at different oxidation states were sometimes collectively referred to as precursors of H2S (41). However, in recent years, researchers have increasingly realized that sulfur species at different oxidation states might have their distinct biological functions due to their difference in chemical properties (6, 60, 64). For example, H2S and RSSH are excellent reducing agents and play important roles in maintaining the intracellular redox balance (32). Further, sulfur species are known to chelate transition metals, and play roles in regulating enzyme activities and functions (6). In addition, persulfides are known to react with the cysteine residue in a protein, leading to persulfidation, which can in turn regulate enzyme functions and/or offer protection against oxidative damage (32, 60).
The involvement of multiple species and the diversity of the chemistry in sulfur signaling are keys to its pleiotropic effects in anti-inflammation, vasodilation, cytoprotection, metabolic rate regulation, among others. As if the involvement of multiple species is not complex enough, an added difficulty in studying sulfur signaling is the rapid interconversion of the various sulfur species under physiological conditions. Such conversions probably are also dependent on the redox state of the intracellular environment because many such reactions are redox reactions (83). All these factors convolute the study of the detailed mechanistic issues in sulfur signaling, complicate the interpretation of results, and lead to sometimes seemingly contradictory results and conclusions. All these complicating factors also mean that donors of various species with well-defined chemistry, stability, release rates, and triggering mechanisms are needed for further studies and for “apple to apple” comparisons of data from various donors (7, 30, 58, 71, 78, 98). One indeed needs to examine the specific sulfur species involved, the cellular conditions, and the release kinetics in interpreting the mechanistic meaning of the relevant biological responses. Another factor that needs to be considered is the fast exchange among various sulfur species at different oxidation states. For example, H2S (oxidation state: −2) can be oxidized to persulfide (oxidation state: −1), then to sulfite (oxidation state: +4), and finally to sulfate (oxidation state: +6) (26). Reduction of some sulfur species such as disulfide can also happen very quickly (1, 91). In this special issue, there are other manuscripts that examine various aspects of sulfur signaling and redox chemistry. This review specifically focuses on donors of persulfide, sulfur dioxide (SO2), and carbon disulfide (CS2).
Persulfide Species: Endogenous Production, Physiological Activities, and Their Donors
Persulfide species R-SSnH (n ≥ 1) play very important physiological roles (20). These persulfide species can be formed through enzyme catalysis or chemical reactions. For example, from cysteine, Cys-S-S-H can be formed by cystathionine-β-synthase or cystathionine-γ-lyase catalysis. Cys-S-S-H can go through disproportionation reaction to form Cys-S-S-S-Cys and H2S, which could generate more complex persulfide species and hydrogen polysulfide through thiol exchange. Glutathione (GSH) can also react with Cys-S-S-H to produce glutathione persulfide (GSSH), which can go through the same process as Cys-S-S-H. Glutathione reductase can also participate in this process by reducing G-S-S-Sn-G (n ≥ 1) to G-S-Sn-H. 3-Mercaptopyruvate sulfurtransferase is also found to be able to generate persulfide species (35). In mouse organs such as liver and heart, the concentration of Cys-S-S-H and GSSH has reported to be in the range 50–100 and 1–5 μM, respectively (20).
On appearance, sometimes hydrogen polysulfides and persulfide (RSnH, n ≥ 2) seem to have similar physiological effects to H2S. For example, donors that deliver pure persulfide species and pure H2S both were reported to exhibit cytoprotective effects from oxidative damage and cardioprotective effects in a murine model of myocardial ischemia-reperfusion (MI/R) (41, 99). However, often hydrogen polysulfides or persulfides exhibit greater potency than H2S. For example, hydrogen polysulfides were found to be able to induce Ca2+ influx by activating transient receptor potential A1 channels in rat astrocytes and are 320 times more potent than H2S (36). H2S2 and GSSH have been reported to directly modify glyceraldehyde 3-phosphate dehydrogenase (GAPDH) activity through cysteine persulfidation while H2S itself incapable of doing so (22, 94). Such results are understandable because the oxidation state of H2S does not allow it to directly persulfidate a thiol group; H2S would have to be oxidized first before doing so. Therefore, in a persulfidation-related signaling process, H2S might simply act as a precursor molecule. All these findings further state the importance of examining the roles of oxidation state of sulfur species in sulfur signaling. As a result, delivering donors at the right oxidation state for mechanistic studies is an important issue. Below, we discuss the various available donors of persulfide in detail.
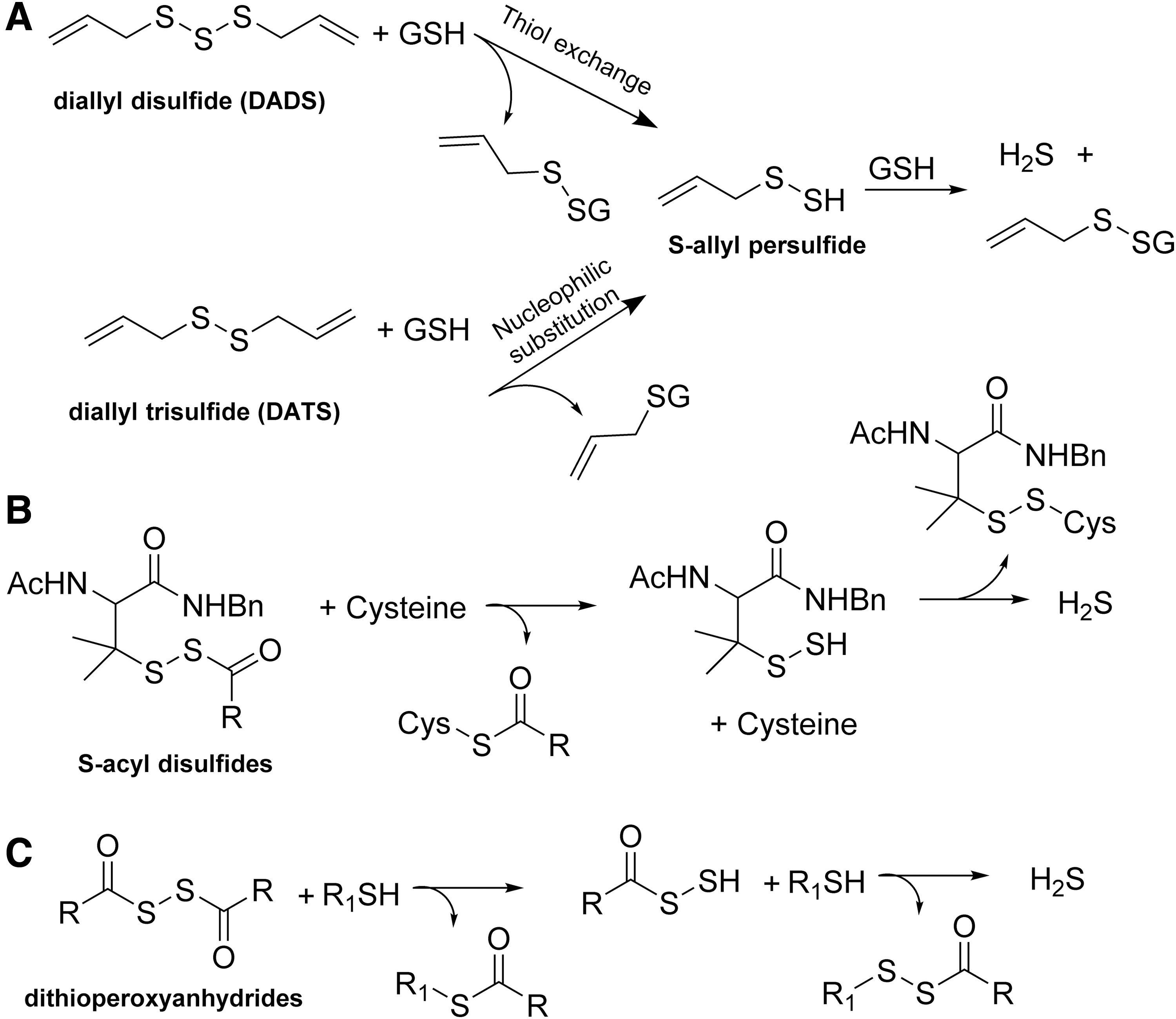
Model persulfide substrates have been prepared for the study of the fundamental chemistry of persulfides such as their stability and degradation product(s) (2, 3, 16, 18). However, those persulfide substrates are not generated under physiological conditions and are not the focus of this review. There are various thiol-activated H2S donors, which would generate a persulfide intermediate and release H2S in the presence of other thiol species such as cysteine. For example, diallyl disulfide and diallyl trisulfide have been shown to release H2S via S-allyl persulfide intermediate in the presence of cellular thiols (Fig. 1A) (51). A series of S-acyl disulfide derivatives developed by Xian's laboratory (Fig. 1B) and dithioperoxyanhydrides developed by Galardon's laboratory (Fig. 1C) have been also used as H2S donors via a persulfide intermediate (29, 72, 96, 97). Although those thiol-activated donors can generate persulfide, these persulfide species would quickly exchange with the thiol present in the system to generate H2S. Thus, they are often viewed as H2S donors instead of persulfide donors, although these donors can probably play the role of both.
Our laboratory has developed a series of esterase-sensitive persulfide prodrugs (99). In the first example reported in 2017, the design took advantage of a masked “hydroxymethyl disulfide” intermediate for stability and esterase sensitivity. Specifically, a hydroxyl group was protected through ester formation (Fig. 2). Esterase-mediated hydrolysis of the ester would lead to a “hemithioacetal-like” intermediate
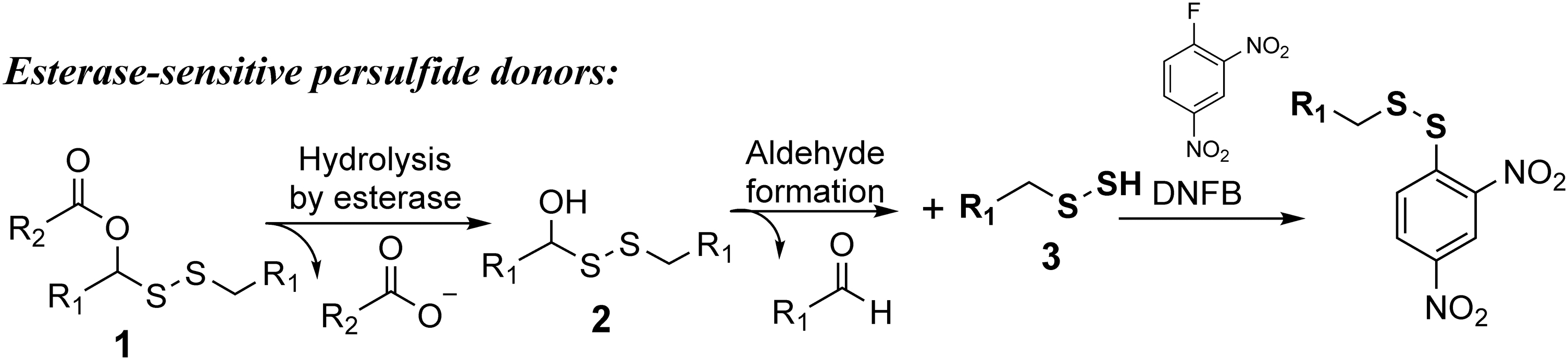
The prodrugs were used in clarifying one chemistry question and in demonstrating the protective effect of persulfide in MI/R injury. Specifically, there had been a debate in the literature as to whether S-methyl methanethiosulfonate (MMTS) would only selectively react with thiol, but not persulfide groups. This is an important question because MMTS had been used to study intracellular thiol contents. The selectivity of MMTS between thiol and persulfide affects the interpretation of such results. Using those prodrugs, it was unequivocally shown that both benzyl thiol and benzyl persulfide were able to react with MMTS, leading to trapped products as confirmed by liquid chromatography–mass spectrometry (LC-MS). As a result, one can unequivocally say that MMTS does not have sufficient selectivity for thiol over persulfide to be used as a thiol-selective probe. It was found that 50 or 100 μg/kg of the prodrug was able to effectively protect mice from MI/R injury. Either lower (12.5 μg/kg) or higher dosage (500 μg/kg) did not exhibit any protective effects, demonstrating dose dependence. The amount of sulfane sulfur was also quantified in the plasma after the administration of the prodrugs. The sulfane sulfur level in plasma was elevated to ∼0.6 μM after treatment with 100 μg/kg of the prodrug. Sulfane sulfur refers to “bound sulfur” that can be converted to H2S by reducing reagents such as dithiothreitol (39). No obvious cytotoxicity was observed in rat myoblast H9c2 cells after 24 h of incubation with up to 100 μM of these esterase-sensitive persulfide prodrugs.
Besides delivering general persulfide species, our laboratory took advantages of a “trimethyl lock”-facilitated lactonization reaction to deliver some endogenous persulfide species such as hydrogen persulfide (H2S2) and GSSH (Fig. 3A, B) (94, 95). The “trimethyl lock” concept was first reported by Cohen and Borchardt in the 1970s to offer entropic advantages in promoting lactonization through the introduction of three pendant methyl groups (labeled with red in compound
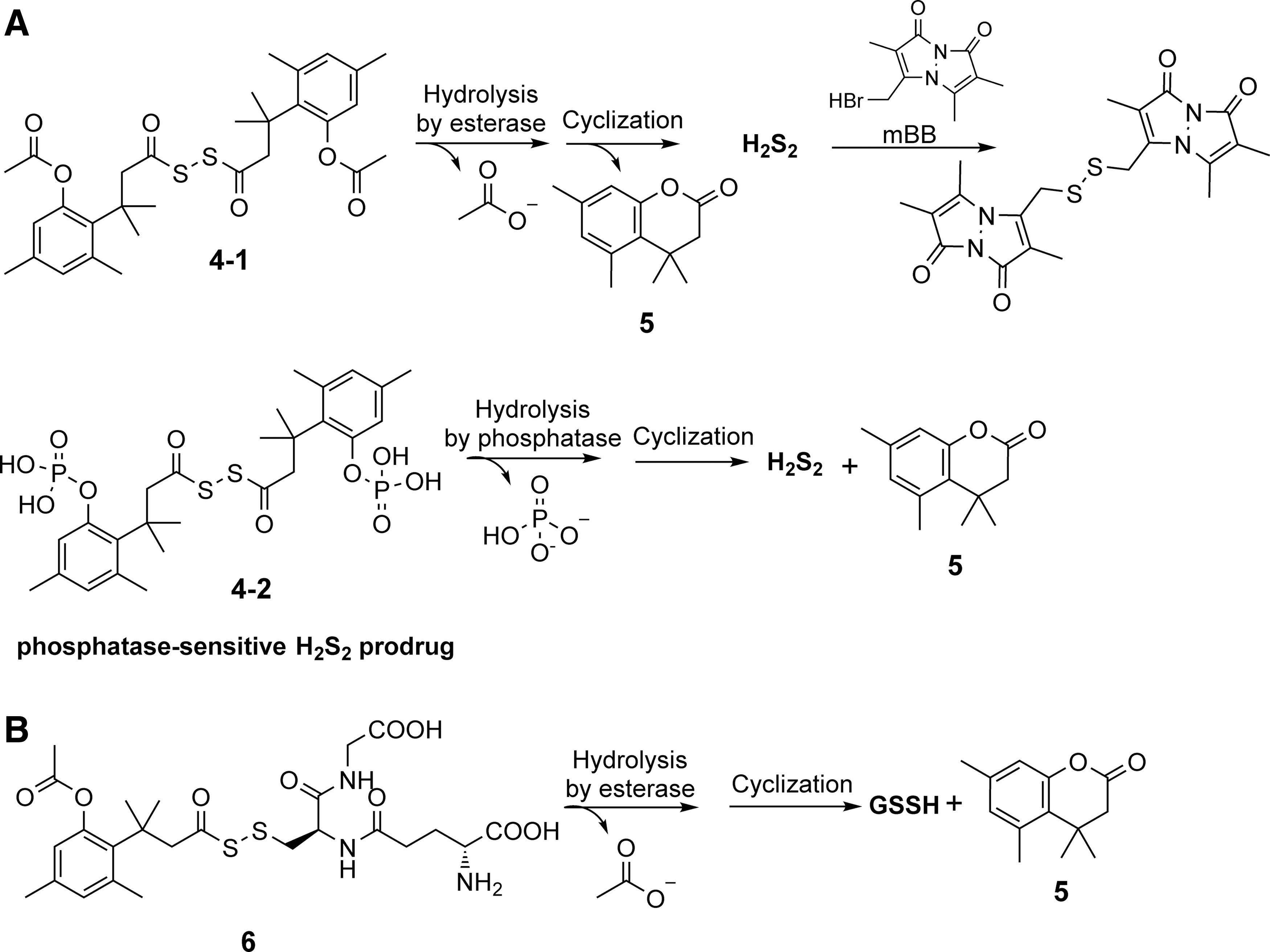
To study the release kinetics, high-performance liquid chromatography (HPLC) was used to measure the generation of byproduct lactone
Our laboratory also utilized the “trimethyl lock” system to build a GSSH donor (Fig. 3B). The GSSH released from the prodrug was trapped by DNFB for the purpose of detection and quantification. However, the trapping yield was only ∼10%, which might indicate the unstable nature of GSSH. Further mass spectrometric studies showed that GSSH itself quickly underwent disproportionation to give GSSSG and H2S. In this study, the GSSH prodrug showed protective effect on H2O2-induced toxicity on rat myoblast H9c2 cells, while H2S, GSH, and the byproduct lactone failed to rescue the cells from the same kind of damage. No obvious toxicity was observed on H9c2 cells at 200 μM after 24 h of incubation with the GSSH donor.
Matson and colleagues reported an ROS-sensitive N-acetyl cysteine (NAC) persulfide donor (70). The design used a boronic acid moiety, which is sensitive to peroxide-mediated oxidation. As shown in Figure 4A, removal of the aryl boronic ester by H2O2 would lead to the formation of an unstable phenol
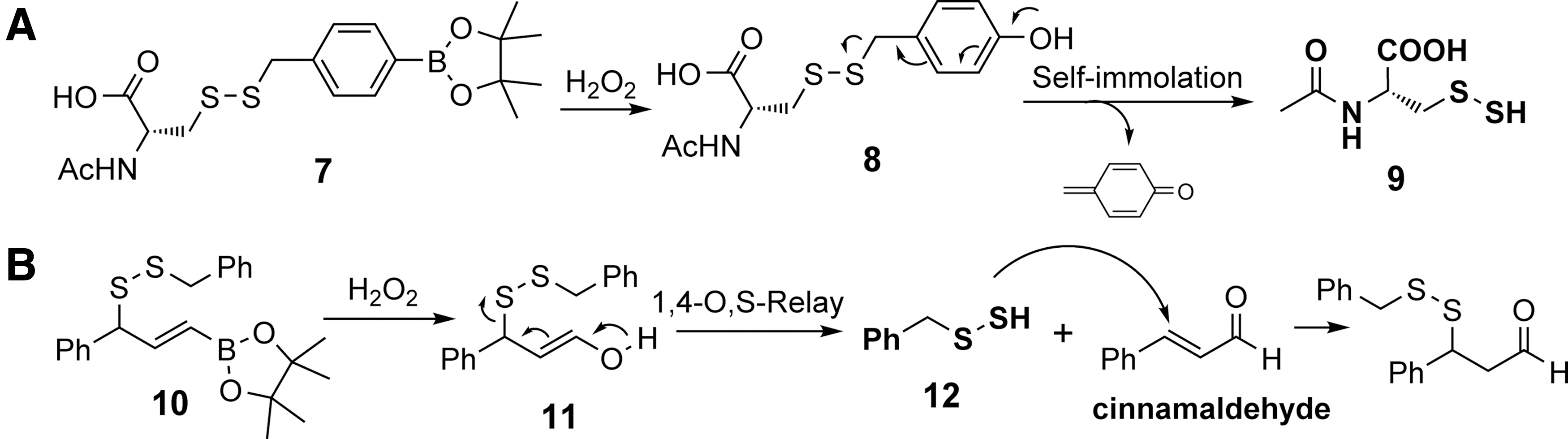
Chakrapani and coworkers also reported an ROS-triggered benzyl persulfide donor. This design relied on a retro Michael addition reaction (4). They also utilized a vinyl boronate ester as the ROS-sensitive moiety (Fig. 4B). In the presence of H2O2, the oxidation would yield an enol intermediate
Khodade and Toscano reported S-substituted thiosiothioureas as persulfide donors under physiological conditions (Fig. 5A) (33). In this design, the terminal sulfhydryl of the persulfide moiety was protected by an S-alkylthioisothiourea moiety. Under near physiological conditions (pH = 7.4), the neutral intermediate
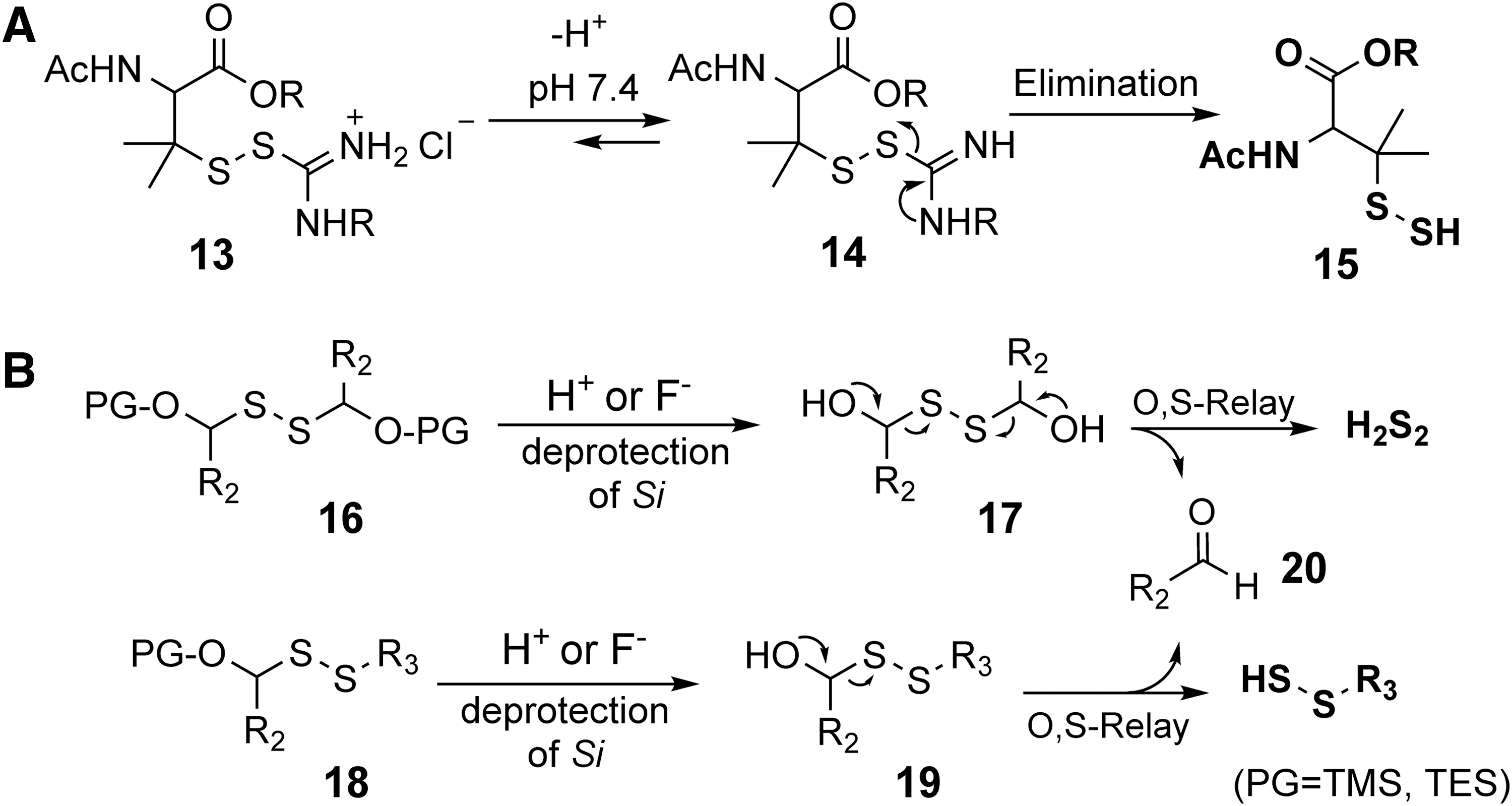
Xian and coworkers developed several persulfide donors based on an O → S relay protection and deprotection mechanism (Fig. 5B) (31). Different protecting groups including trimethylsilyl and triethylsilyl were used for acid- or fluoride-triggered release. Upon desilylation, the unstable intermediate
SO2: Endogenous Production, Physiological Roles, and Prodrugs
Although traditionally viewed as a toxic gas to human body and environment, SO2 has been reported to have various pharmacological effects, especially in the cardiovascular system (14). Both SO2 gas and its derivatives (Fig. 7) induce concentration-dependent relaxation of isolated rat aortic rings with similar potency, suggesting SO2's role as a potential vasoactive molecule (14, 59). SO2 exposure was reported to cause a consistent decrease in blood pressure in salt-induced hypertension rat model. Further in spontaneously hypertensive rats, a significant decrease in SO2 concentration and aspartate aminotransferase (AAT) activity in the plasma was observed, suggesting SO2's role in blood pressure regulation (13, 19). It has been shown that SO2 provides protective effects in monocrotaline-induced pulmonary hypertension, MI/R injury, and atherosclerosis (25, 49, 88). Endogenous SO2 has also been shown to offer protection against oleic acid-induced acute lung injury by suppressing oxidative stress (8). The fact that SO2 has its own enzymatic production pathway and plays various roles under physiological/pathological conditions has led to the proposal that SO2 is a new gasotransmitter (19).
Although the story of SO2 as an endogenous signaling molecule is not without controversy, one thing is for sure: SO2 has been shown to have pharmacological effect and is produced endogenously through the metabolism of sulfur-containing amino acids or through oxidation of H2S (Fig. 6) (28, 75). Some of these reactions are catalyzed by enzymes (26). For example, cysteine dioxygenase is known to oxidize

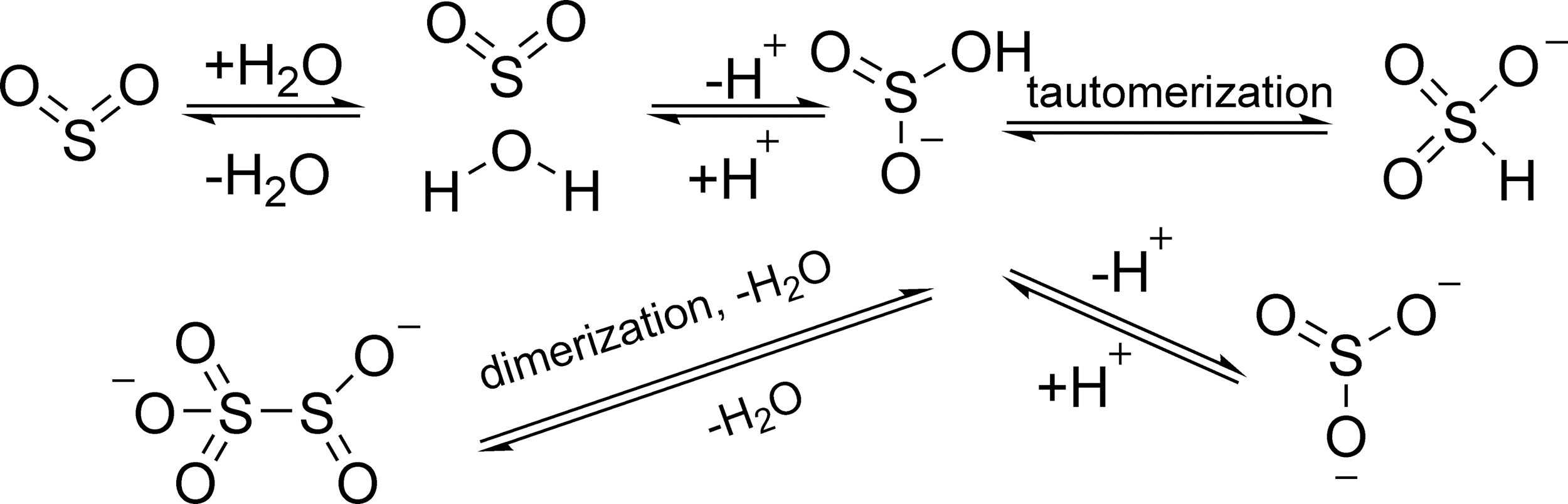
SO2 can readily dissolve in water and form a hydrated SO2 complex (SO2·H2O). This complex would afford bisulfite (HOSO2 − or HSO3 −) and sulfite (SO3 2−) ions depending on the pH. Bisulfite has two tautomers in aqueous solution (Fig. 7) (15, 63). The first and second pKa of SO2·H2O are 1.81 and 6.97, respectively. Therefore, under physiological conditions (pH = 7.4), the molar ratio of bisulfite to sulfite is ∼1:3 (80 –82). Disulfite (S2O5 2−) can be formed by the dimerization of bisulfite. These species exist in equilibrium in the biological system. It is important to recognize that studying SO2 under physiological conditions is actually the examination of various SO2 derivatives as a whole, not a single species (Fig. 7).
Despite the understanding of SO2 chemistry and its known pharmacological roles, the mechanism of action for SO2 is far from clear yet. Whether SO2 has biological targets is still a question. How SO2 might bind and/or modify possible targets has not been well studied. Further, the role of SO2 in affecting redox balance is not clear. All these unanswered questions are more of a reason to develop tools that can deliver SO2 in a controllable fashion to enable detailed mechanistic studies. The section below describes the recent development of SO2 prodrugs and donors.
In 2012, Chakrapani's laboratory reported a series of thiol-sensitive SO2 donors as antimycobacterial agents (Fig. 8) (57). This was the first reported effort in making SO2 prodrugs. The design was based on the 2,4-dinitrophenylsulfonamide scaffold, which was previously used as a thiol probe. The SO2-releasing mechanism involved ipso-nucleophilic substitution on an electron-deficient aromatic ring (Fig. 8). Various analogs with different R1 and R2 groups were synthesized to evaluate the effects of the substituent on SO2 production yields and rates. In the presence of 10 equivalents of cysteine, 100 μM of donors were able to release SO2 in 24%–100% yield within 30 min in PBS (pH = 7.4). The release half-life ranged from 2 to 64 min, depending on the substituents on the sulfonamide nitrogen. One of the SO2 donors was found to inhibit Mycobacterium tuberculosis (Mtb) with a minimum inhibitory concentration (MIC) of 0.05 μM, while its IC50 on human embryonic kidney HEK293 cells was 7 μM. The inhibitory effects on M. tuberculosis were said to possibly involve the depletion of thiol species during the activation stage and the ability of SO2 to induce oxidative stress and to damage biomacromolecules such as lipids, proteins, and DNA. Later, the Chakrapani laboratory reported the synthesis of additional 2,4-dinitrophenylsulfonamide analogs as SO2 donors and antimicrobial agents against M. tuberculosis (56). The antimicrobial activity of those 2,4-dinitrophenylsulfonamide SO2 prodrugs was also tested on various Gram-positive bacteria such as methicillin-resistant Staphylococcus aureus (MRSA) (66). Among these compounds, the most potent one has a MIC of 2 μg/mL. However, these compounds showed no activity against Gram-negative Escherichia coli at up to 32 μg/mL, probably because of permeability issues for the prodrug. Evidence showed that the SO2 release yield is not the only factor that affects the antimicrobial potency. Incidentally, these 2,4-dinitrophenylsulfonamide SO2 prodrugs have also been widely used as a source of SO2 to validate SO2 detection methods (47, 52).

In 2018, Chen's group used this thiol-responsive sulfur dioxide prodrug to form nanocarriers for doxorubicin (74). The released SO2 showed a synergistic effect with doxorubicin toward drug-resistant cancer cells in cell culture studies.
These thiol-triggered 2,4-dinitrophenylsulfonamide SO2 prodrugs have a well-defined release mechanism, half-life, and yields, and are great research tools. However, the use of thiol as a trigger may complicate mechanistic studies of sulfur signaling, and the dinitrophenyl byproduct may also limit its application in therapeutic application due to its potential metabolic toxicity.
In 2013, the Chakrapani laboratory reported a 1-phenyl-benzosultine scaffold for the design of thermally responsive SO2 donors (Fig. 9) (55). Such a structure can undergo a thermal retro-Diels-Alder reaction, resulting in the release of SO2, which can further react with 1,3-dienes to form benzosulfone. Specifically, in acetonitrile (ACN) at 37°C, 1-phenyl-benzosultine would decompose to produce the more stable 1-phenyl-benzosulfone in 92% yield. Such results were interpreted as to suggest that the SO2 released from 1-phenyl-benzosultine further reacted with the 1,3-dienes to generate 1-phenyl-benzosulfone. However, in phosphate buffer at 37°C and pH 7.4, the yield of SO2 from 1-phenyl-benzosultine was ∼89% as calculated based on the formation of SO2 as sulfite. Such results suggest that solvent plays an important role in determining whether the released SO2 would undergo further reactions with a 1,3-diene. Although the mechanistic aspect of this solvent effect was not further explored, it is possible that the enhanced solubility of SO2 and the conversion of SO2 to sulfite species in aqueous medium could help shift the equilibrium in favor of SO2 generation. Through the modification of electron density on the phenyl ring, various SO2 donors were designed with half-life (calculated based on sulfite formation) ranging from 10 to 68 min in phosphate buffer containing 30%–40% ACN, at 37°C and pH 7.4. Compound

In 2015, the Chakrapani group reported a series of photochemically activated SO2 donors based on a benzosulfone scaffold (Fig. 10A) (54). Various benzosulfone compounds

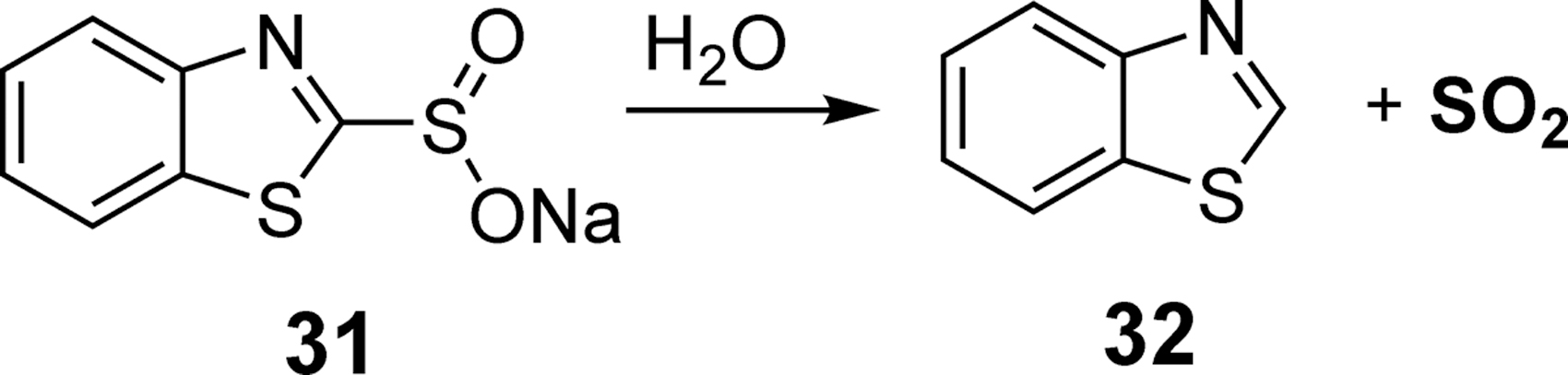
In 2016, Xian's laboratory reported a SO2 donor utilizing a benzothiazole sulfinate (BTS) moiety (Fig. 11) (10). In the presence of water, BTS
In 2017, our laboratory designed a series of SO2 donors via a click-release reaction between thiophene dioxide and a strained alkyne (Fig. 12) (86). This strategy was inspired by our previous work on click-release CO prodrugs (24, 84). As a proof of concept, tetrachlorothiophene dioxide

In 2017, our laboratory reported a class of esterase-sensitive SO2 donors based on a modified Julia olefination reaction (87). As shown in Figure 13A, after esterase-catalyzed hydrolysis of the ester bond, a hydroxyl group was revealed to form intermediate
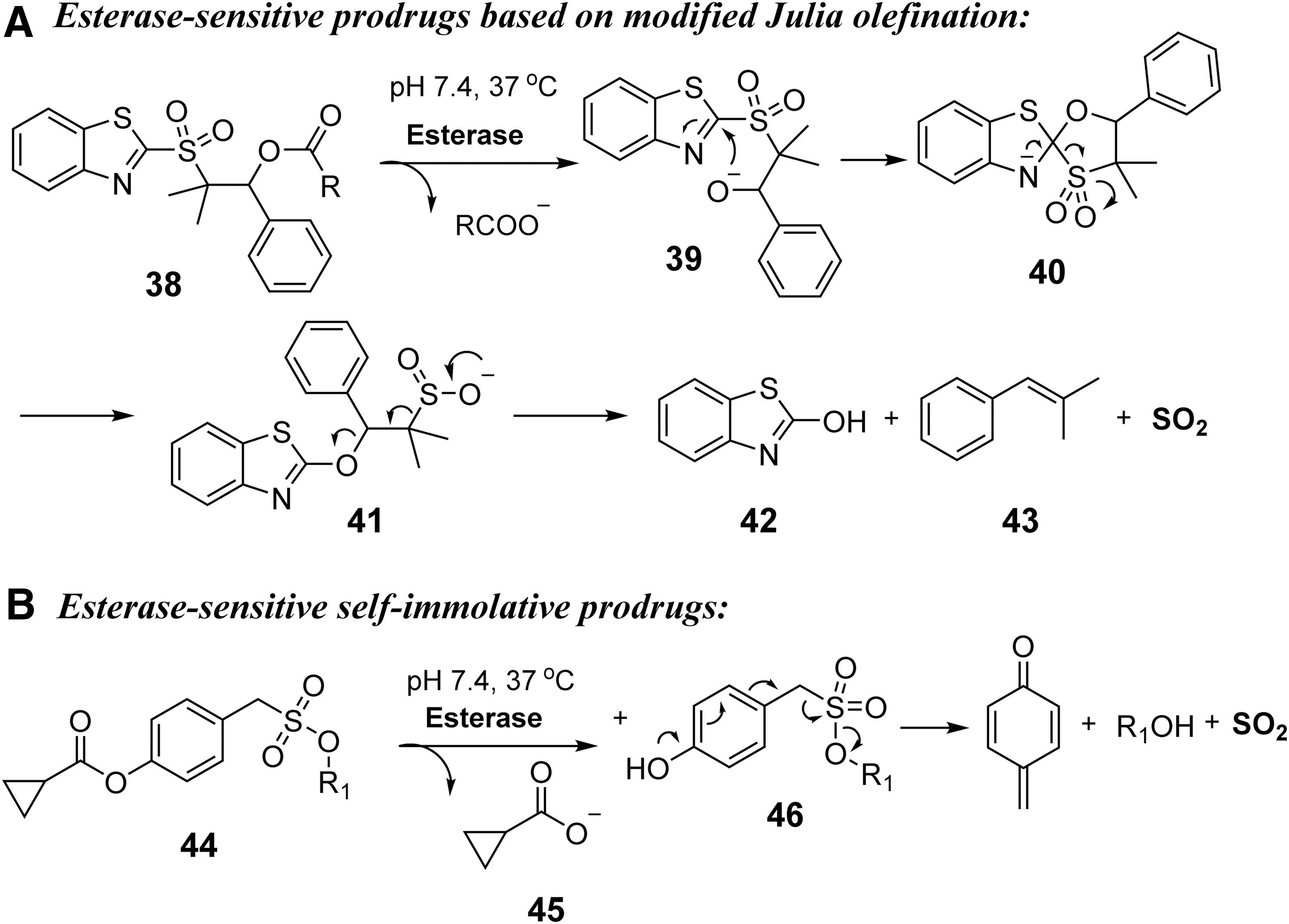
In 2018, the Chakrapani group reported another class of esterase-sensitive SO2 donors (67). As shown in Figure 13B, after esterase-catalyzed hydrolysis, the ionization of the unmasked hydroxyl group would lead to an elimination reaction and C-S bond cleavage to release SO2 from the sulfonate moiety. As a proof of concept, compound
Based on the currently available data, it seems that one of the major areas of application of SO2 donors is in the treatment of cardiovascular disease. However, the application of SO2 in the treatment of bacterial infection has also been explored (9). Thiol-activated SO2 prodrugs developed by Chakrapani's laboratory have shown promising results against M. tuberculosis and MRSA, probably by disrupting redox balance and damaging biomacromolecules such as lipids, proteins, and DNA. However, those thiol-activated prodrugs face the same dilemma as many narrow-spectrum antibiotics: they are far more effective against Gram-positive bacteria than Gram-negative bacteria, possibly due to the difficulty for the donors to penetrate the outer membrane of Gram-negative bacteria (38). Furthermore, translating antimicrobial effect in bacterial culture to in vivo applications faces the same hurdle as many other known antimicrobials; that is, they have to deal with issues of toxicity, selectivity, potency, and drug delivery and other pharmacokinetic challenges.
CS2: A Precursor to H2S or an Active Compound by Itself?
CS2 has been implicated in various biological processes on its own, although CS2 is only produced naturally by soil-based microorganisms, and there is currently no evidence of its enzymatic production in mammalian cells (90). CS2 is relatively inert to hydration to form H2S at biologically relevant pH, and the approximate half-life was estimated to be 1.1 years at pH 9 (12). CS2 is also inert to carbonic anhydrase (CA), which catalyzes the hydrolysis of CO2. When hydrolyzing CS2, CA II is about nine orders of magnitude less effective compared with the hydrolysis of CO2 (11, 12, 17, 77). However, free thiol such as cysteine and GSH can react with CS2 to form trithiocarbonates, which might be a form of post-translational cysteine residue modification to modulate protein function much the same way as protein S-persulfidation. Free amines can also react with CS2 to form dithiocarbamates (DTCs). For example, the free amino group on the dopamine molecule can form DTC with CS2. This DTC formed from dopamine and CS2 is known to chelate the copper center of dopamine β-hydroxylase and inhibit its activity (53). CS2 can also bind and inhibit proteins with the Ni/Fe–S cluster in a reversible manner; this can also be another way of CS2 signaling (44). More comprehensive and detailed discussions can be found in a recent review (12). There have been efforts to build CS2-releasing molecules to further probe CS2's biological effect. Along this line, the Ford group reported a series of DTCs derived from

Conclusions
One of the difficulties in studying sulfur signaling is the rapid interconversion of various sulfur species through redox reactions, which leads to scrambling among the various species. Furthermore, the reciprocal effects of cellular oxidative stress and availability of various sulfur species convolute the interpretation of results from mechanistic studies. The availability of a range of donors of sulfur species at various oxidation states hopefully would help deciphering results from mechanistic studies. It is our hope that this review article will stimulate additional work to further improve the tools available, and more discussions of the challenging issues facing sulfur-signaling studies, especially in the context of redox chemistry, with the ultimate goal of achieving a thorough understanding of the complex chemistry and biology issues in sulfur-mediated biological processes.
With the many demonstrated pharmacological effects of various sulfur species, there is always the question of whether any of these pharmacological effects would ever translate into successful development of therapeutic agents. This is truly a billion-dollar question, and it is a long-term issue. Intuitively, the demonstrated pharmacological effects would surely suggest a high degree of likelihood for further pharmaceutical development of these donors of sulfur species. However, the reality might be very different. It is our opinion that a key to success in this area is to identify an area of unmet medical need that can tolerate the complication of the rapid interconversions among the various sulfur species. Further, one has to put the question of clinical indication in the context of desired pharmacokinetic profiles, the route of delivery, and the possibility of feedback controls in sulfur signaling and thus development of drug resistance. The last point would require in-depth understanding of sulfur-signaling biology and much more data from animal models. We might still be a long way from realizing the therapeutic potential of sulfur signaling. Only in the context of a specific clinical indication, one can truly assess the possibility for a particular sulfur donor to be developed for further clinical application.
Footnotes
Acknowledgments
We gratefully acknowledge the Brains and Behavior program at Georgia State University for a predoctoral fellowship to Z.Y. and the financial support of the Georgia Research Alliance to B.W. in the form of a GRA Eminent Scholar endowment. We thank Dr. Gabriel Gojon for the invitation for this review.
Funding Information
We gratefully acknowledge the Brains and Behavior program at Georgia State University for a predoctoral fellowship to Z.Y., and financial support from Georgia State University internal resources and Georgia Research Alliance Eminent Scholar endowment fund to B.W.