
Abstract
Significance:
S-nitrosylation, the post-translational modification by nitric oxide (NO) to form S-nitrosothiols (SNOs), regulates diverse aspects of cellular function, and aberrant S-nitrosylation (nitrosative stress) is implicated in disease, from neurodegeneration to cancer. Essential roles for S-nitrosylation have been demonstrated in microbes, plants, and animals; notably, bacteria have often served as model systems for elucidation of general principles.
Recent Advances:
Recent conceptual advances include the idea of a molecular code through which proteins sense and differentiate S-nitrosothiol (SNO) from alternative oxidative modifications, providing the basis for specificity in SNO signaling. In Escherichia coli, S-nitrosylation relies on an enzymatic cascade that regulates, and is regulated by, the transcription factor OxyR under anaerobic conditions. S-nitrosylated OxyR activates an anaerobic regulon of >100 genes that encode for enzymes that both mediate S-nitrosylation and protect against nitrosative stress.
Critical Issues:
Mitochondria originated from endosymbiotic bacteria and generate NO under hypoxic conditions, analogous to conditions in E. coli. Nitrosative stress in mitochondria has been implicated in Alzheimer's and Parkinson's disease, among others. Many proteins that are S-nitrosylated in mitochondria are also S-nitrosylated in E. coli. Insights into enzymatic regulation of S-nitrosylation in E. coli may inform the identification of disease-relevant regulatory machinery in mammalian systems.
Future Directions:
Using E. coli as a model system, in-depth analysis of the anaerobic response controlled by OxyR may lead to the identification of enzymatic mechanisms regulating S-nitrosylation in particular, and hypoxic signaling more generally, providing novel insights into analogous mechanisms in mammalian cells and within dysfunctional mitochondria that characterize neurodegenerative diseases.
Introduction
Bacteria perpetually encounter and respond to redox stresses in their environment, particularly oxidative and nitrosative stresses encountered during infection. Facultative bacteria such as Escherichia coli also experience endogenous redox stress: oxidative stress when growing aerobically in an oxygen-rich environment and nitrosative stress when growing anaerobically in the presence of nitrate. The bacterial transcription factor OxyR acts as a regulator of both stresses, undergoing oxidative or nitrosative modifications of a single regulatory Cys (S-sulfenylation/disulfide or S-nitrosylation, respectively) to direct distinct transcriptional responses. The OxyR oxidative stress regulon of ∼30 genes is well characterized, and includes genes with anti-oxidative functions. Similarly, nitrosative stress promotes the OxyR-dependent activation of a much larger, but less well-characterized regulon of ∼100 genes, including the gene for a master regulator of S-nitrosylation, the S-nitrosylase hybrid cluster protein (Hcp). More specifically, unlike oxidation of OxyR, which is a consequence of high levels of oxidants acting relatively nonspecifically, S-nitrosylation is mediated enzymatically. Targeted S-nitrosylation of proteins (including of OxyR itself) by Hcp confers a survival advantage. Thus, remarkably, targeted S-nitrosylation of specific proteins protects cells from the effects of aberrant and nontargeted S-nitrosylation due to external nitrosative stress. Here, we review this central function for OxyR—the classic aerobic regulator of oxidative stress—in protecting against nitrosative stress under anaerobic conditions, and reveal parallels between regulation of S-nitrosylation in E. coli and hypoxic signaling by nitric oxide (NO) in mammalian cells, and in mitochondria in particular.
Bacteria serve as model organisms that allow for identification and characterization of redox stress responses due to the ease of genetic manipulation coupled with the flexibility in growth conditions. Facultative bacteria such as E. coli can be grown aerobically or anaerobically; in the absence of oxygen, E. coli can extract energy through respiration by using any of a number of electron acceptors, with different metabolic outcomes, or they can even utilize fermentation in the absence of electron acceptors (105). Bacteria can encounter both oxidative and nitrosative stresses resulting from their environment or their own metabolism, and studies in bacteria have led to identification and/or better understanding of key enzymes that counter these stresses (9, 61).
During evolution, mitochondria in eukaryotic cells originated from endosymbiotic bacteria (84). Bacteria have been used as model systems to study mitochondrial functions, especially for studies involving respiratory complexes (80), and it is highly likely that at least some mechanisms of generation of and protection from redox stress are conserved. Mitochondria have long been known to be the source of damaging reactive oxygen species (ROS), especially superoxide (O2 −) produced by the one-electron reduction of oxygen (67). Moreover, dismutation of superoxide leads to the formation of hydrogen peroxide (H2O2) that can reversibly oxidize proteins and can lead to redox signaling (67). More recently, it has been appreciated that mitochondria are also a source of NO, primarily under hypoxic conditions (54), and that S-nitrosothiol (SNO)-based signaling is prevalent within mitochondria. Accumulating evidence suggests that aberrant S-nitrosylation of mitochondrial proteins may play an important role in neurodegenerative diseases. In this review, we focus on the consequences of NO generation by bacteria under hypoxia, and by analogy in mitochondria (54).
Respiration-Mediated Generation of NO and ROS
In all organisms, aerobic growth is associated with the obligate production of reactive oxygen intermediates from the incomplete reduction of oxygen during metabolism (32, 44). The production of oxygen radicals and reactive species, including O2
−, H2O2, and hydroxyl radicals (HO•), leads to endogenous oxidative stress and to consequent induction of enzymatic defense mechanisms (32, 43). Early studies in bacteria demonstrated the essential importance of these protective enzymes for cellular function. Complete lack of superoxide dismutase activity in E. coli inhibits growth in air; exogenous amino acids rescue growth by overcoming the oxidative inhibition of enzymes involved in amino acid biosynthesis (9). Similarly, bacteria in which H2O2-consuming activities, catalase (KatG) and alkyl hydroperoxide reductase (AhpC), are absent have a growth defect during aerobic conditions (89). In this case, growth inhibition is a result of oxidative damage at iron
In the absence of oxygen, facultative anaerobes such as E. coli preferentially utilize nitrate as the electron acceptor for respiration (105). During anaerobic respiration on nitrate (ARN), the nitrate reductase NarGHI is induced (92), which serves as a source of NO by reducing nitrite (a reaction secondary to its function in nitrate reduction) (45, 79). NO produced during ARN exerts an endogenous nitrosative stress (90, 112) and, consequently, leads to the induction of genes previously shown to be activated by toxic amounts of NO, including hmp, hcp-hcr, yeaR-yoaG, and ytfE (27, 42, 71, 75). Notably, nitrosative stress by NO also disrupts iron
Redox-Based Sensors and the Transcription Factor OxyR
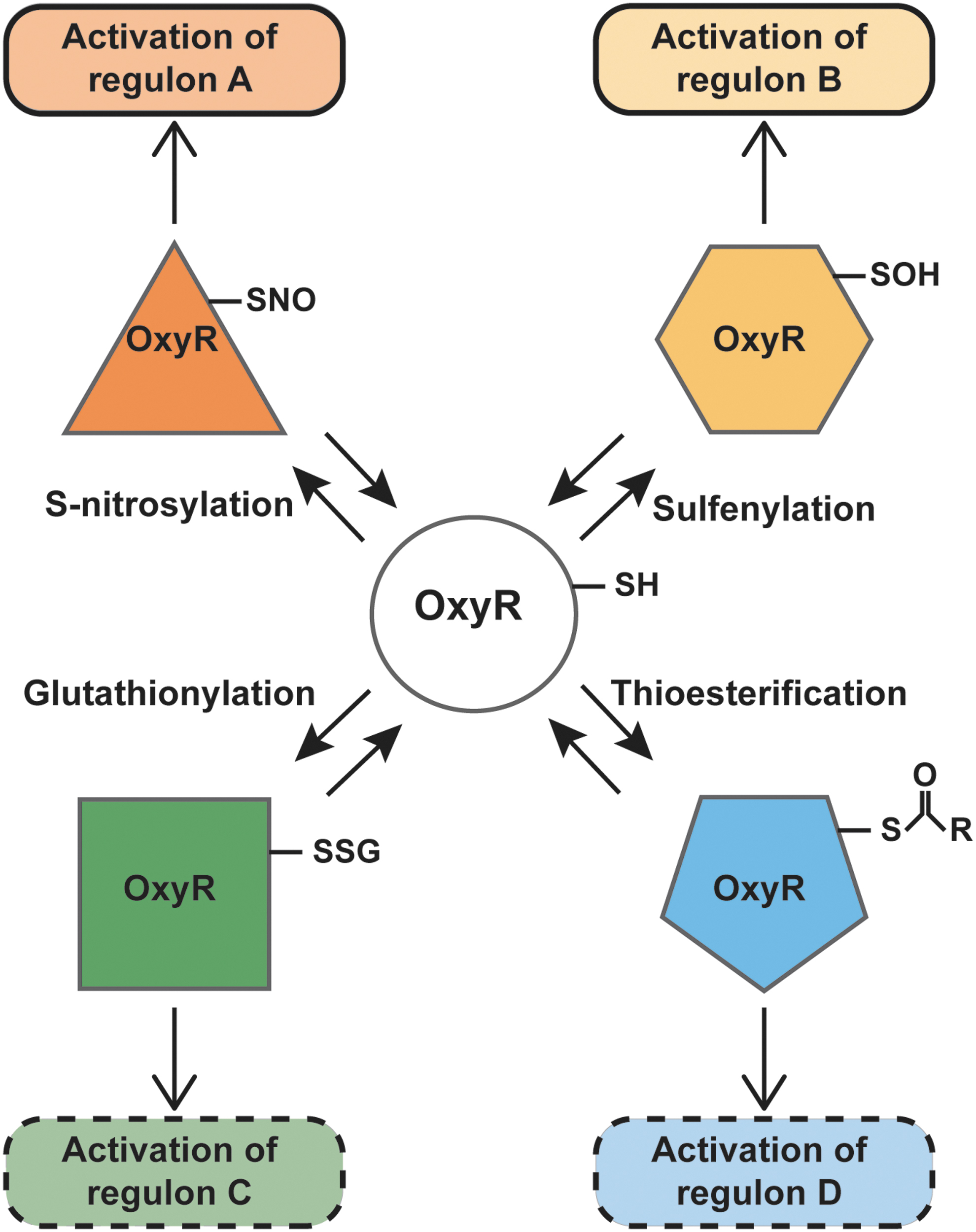
The analogy to oxidative stress centers on thiol groups, which may undergo several redox-based modifications in the context of redox-based stress—some specific to the nature of the stress and some shared across stressors. Reactive cysteines generally have a low pKa and are ionized at physiological pH; the thiolate is often stabilized by hydrogen bonding, by proximity to positively charged residues or by location at the N-terminus of an α-helix (85). The bacterial transcription factor OxyR, a member of the LysR family of transcription factors (88), possesses one such reactive cysteine, Cys199. Based on the crystal structure of OxyR, it has been suggested that interaction with a positively charged arginine residue contributes to the lowering of the pKa of Cys199 (12) to make it particularly susceptible to redox modifications. It has been further proposed that alternative redox modifications of OxyR (52) can lead to distinct transcriptional responses, thereby forming the basis of a molecular redox code (Fig. 1). We review here new evidence in favor of the concept that OxyR senses oxidative and nitrosative stresses through alternative modifications of Cys199, leading to distinct transcriptional consequences. We further discuss the protective function of an OxyR-regulated gene, hcp, in endogenous S-nitrosylation, and hypothesize that similar enzymatic mechanisms may function in mitochondria.
E. coli as a Model for Nitrosative Stress
Nitrosative stress is characterized by excessive S-nitrosylation of proteins. Intracellular protein
Endogenous Oxidative and Nitrosative Stress
Bacteria are exposed to exogenous redox stresses in their environment, and even more so during infection where the host innate immune system targets invaders by producing ROS and reactive nitrogen species (RNS) via NADPH oxidases and inducible nitric oxide synthase (iNOS), respectively [reviewed in Nathan and Shiloh (68)]. However, as discussed earlier, growth in an aerobic environment also leads to endogenous ROS-mediated oxidative stress, and thus bacteria have evolved shared mechanisms to protect against both endogenous and exogenous ROS stresses. Similarly, anaerobic growth in the presence of nitrate (the preferred electron acceptor for respiration in the absence of oxygen) leads to the production of NO/RNS, and hence endogenous nitrosative stress. Although the endogenous levels of RNS are far lower than those generated by the host immune system, defenses against constitutive RNS also help protect bacteria during infection (3). For example, bacterial flavohemoglobin (Hmp), an enzyme that consumes NO under both aerobic and anaerobic conditions (37), is essential for growth on nitrate (17) and is protective against host NO in infection (100). Indeed, it seems likely that bacteria lacking these endogenous responses would be more sensitive to exogenous nitrosative stress-induced killing. Thus, understanding how bacteria cope with endogenous oxidative and nitrosative stresses may allow insights into their ability to survive in a host during infection.
There are several similarities between the endogenous oxidative stress generated in bacteria during growth in oxygen and the endogenous nitrosative stress derived from anaerobic growth on nitrate, which we will draw attention to in this review. First, just as oxidative stress is an inevitable consequence of aerobic growth, endogenously generated NO can be considered an inevitable consequence of ARN. Second, nitrosative stress manifests as high levels of protein S-nitrosylation (39, 90), just as oxidative stress manifests as high levels of S-oxidation (26). Third, E. coli respond to these stresses by inducing genes that encode for proteins that scavenge ROS or NO or otherwise counter their effects. Fourth, the transcription factor OxyR is induced by both oxidative and nitrosative stresses, under either aerobic or anaerobic conditions, and confers protection against each, although through distinct responses.
Notably, OxyR is essential for bacterial survival in air (under certain conditions) (24, 36), and also for anaerobic growth on nitrate (90), indicative of the central role played by this sensor/transcription factor in protecting against both stresses. It is perhaps not surprising, therefore, that OxyR controls many of the genes previously identified with protection from hydrogen peroxide and from NO.
OxyR as a Sensor of Oxidative and Nitrosative Stress
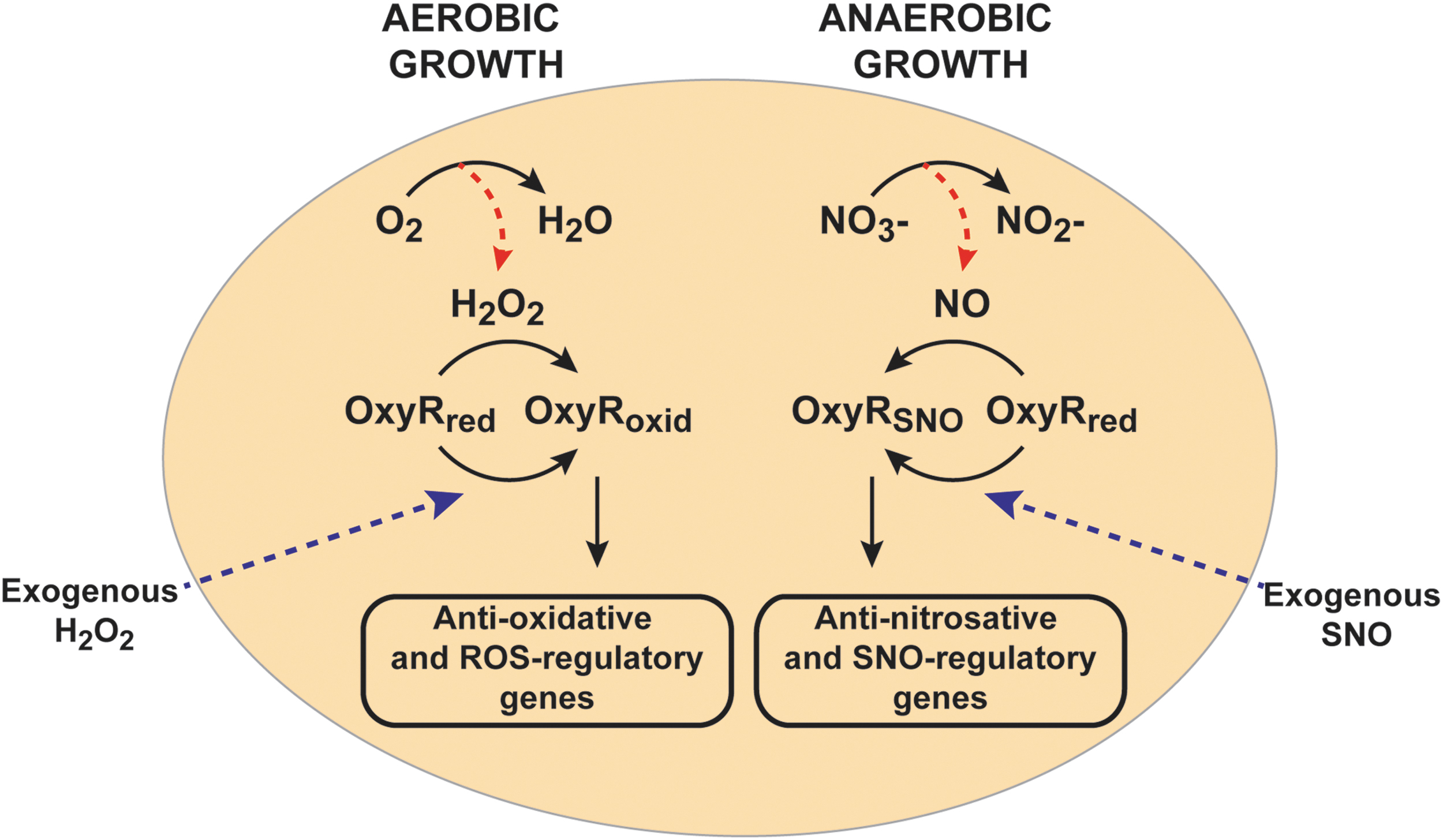
First identified in Salmonella typhimurium and E. coli, the transcriptional factor OxyR is the primary sensor for hydrogen peroxide in bacteria (14). Reduced OxyR is inactive as a transcription factor, whereas oxidation of OxyR at the active site cysteine residue, Cys199, leads to transcription of the OxyR regulon of ∼30 genes, which includes the anti-oxidative genes catalase (katG), alkyl hydroperoxide reductase (ahpCF), and glutathione reductase (96) (Figs. 1 and 2). E. coli lacking OxyR are highly sensitive to hydrogen peroxide and other oxidative stress-inducing agents. Importantly, not only does the OxyR regulon ensure direct scavenging and removal of hydrogen peroxide (via katG) and organic hydroperoxides (via ahpC), but it also counters consequences of oxidative injury, including through dps, which encodes an iron (Fe2+)-sequestering protein that protects DNA from Fenton chemistry, and glutaredoxin and trxA (thioredoxin), which remove oxidative modifications from proteins (96). Surprisingly, OxyR functions in anaerobes as well, including Porphyromonas gingivalis, Bacteroides fragilis, and Bacteroides thetaiotaomicron (22, 65, 83). At first glance, the role and function of OxyR in oxidative stress would appear to preclude a direct role under anaerobic conditions, so it has been assumed that OxyR must protect against adventitious oxygen or H2O2 in these anaerobes.
Biology is nothing if not innovative in reusing mechanisms that are efficient, especially in bacteria, and OxyR is a prime example. This transcription factor is regulated by a sensor thiol Cys199 that is subject to sulfenylation (S-OH) and consequent disulfide formation (52, 96, 119). But thiols may undergo multiple modifications, including S-nitrosylation, S-glutathionylation, and thioesterification, which may form the basis of a molecular code through which alternative Cys modifications produce different functional responses (15, 113). It has, thus, been proposed that OxyR may be capable of transducing distinct transcriptional responses (35, 52, 90). Although distinct S-sulfenylation OxyR and SNO-OxyR regulons have been identified (90, 117, 120), glutathionylated- and thioesterified-OxyR regulons are still hypothetical (Fig. 1).
S-nitrosylation of OxyR, first demonstrated in anaerobically growing E. coli treated with the SNO donor S-nitrosocysteine (39), has served as proof-of-concept. S-nitrosylated OxyR (SNO-OxyR) is in an activated state that is able to promote transcription at target genes (39, 52, 90) (Fig. 2). Initial studies presumed that SNO-OxyR activated transcription of the same well-characterized oxidative stress-induced regulon of OxyR, including, for example, katG and oxyS, although to a lesser degree than hydrogen peroxide. However, although NO donors induced transcription of katG and oxyS (52, 119), S-nitrosylated OxyR was quite different from S-oxidized OxyR in its affinity for previously identified DNA target sites (52). Moreover, under truly physiological conditions where endogenously generated NO leads to S-nitrosylation, SNO-OxyR actually promotes transcription of a unique set of genes, almost entirely nonoverlapping with the previously known regulon induced by oxidized OxyR (90). This nitrosative stress-specific OxyR regulon of >100 genes includes numerous enzymes that are protective against nitrosative stress and/or well known to be induced by NO, including hcp-hcr (27, 42, 71, 75, 90). Figure 2 shows these distinct functions of OxyR under aerobic (or oxidative stress) conditions versus anaerobic (or nitrosative stress) conditions, which result in oxidized and S-nitrosylated forms of OxyR, respectively. This largely nonoverlapping nature of the oxidized and S-nitrosylated OxyR regulons explains why an earlier study that looked at NO-dependent gene expression of oxidized-OxyR genes concluded that OxyR is not a primary regulator of the bacterial response to nitrosative stress (66).
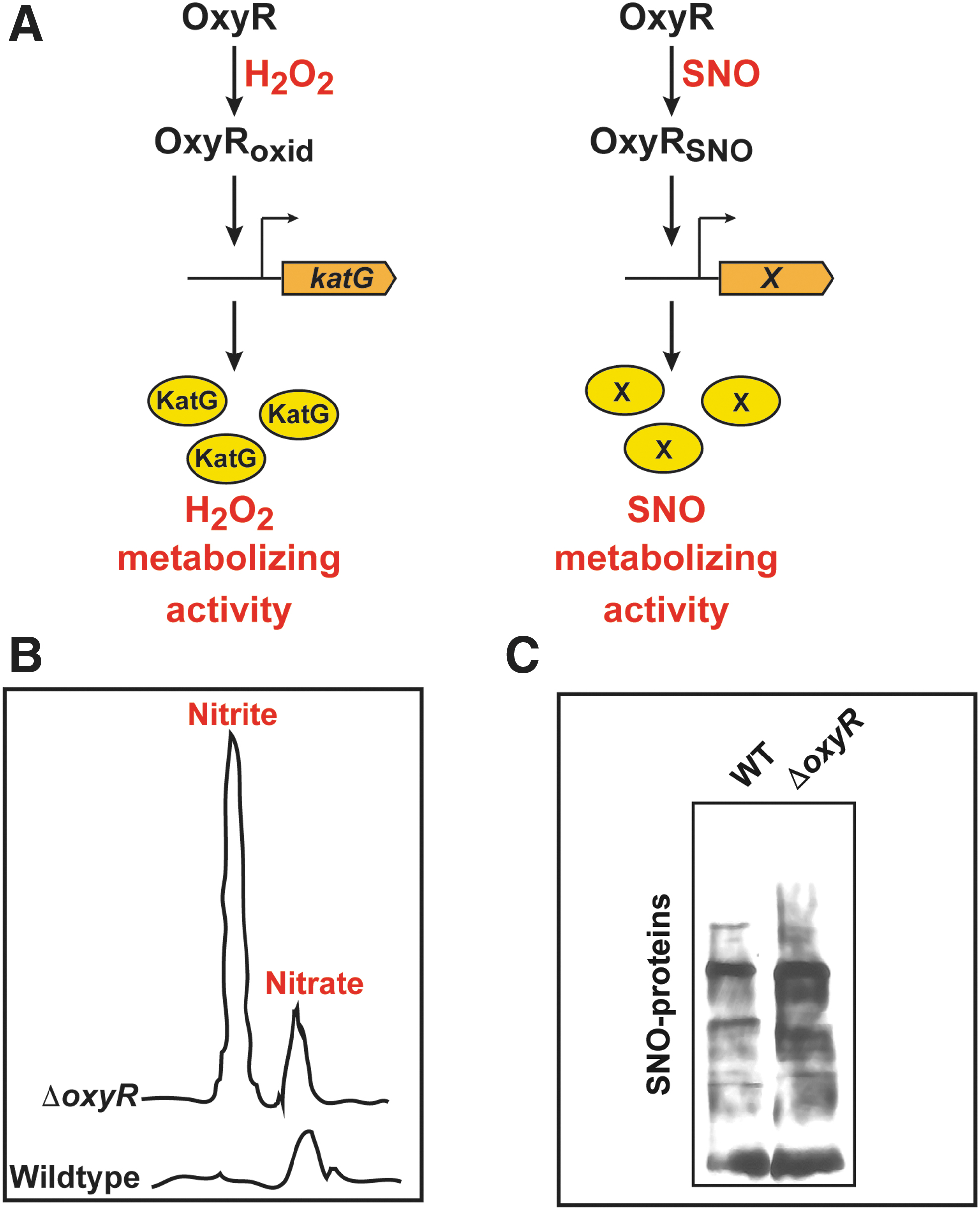
Thus, OxyR acts as a redox sensor and switch: It senses both ROS and RNS, and it transduces distinct transcriptional responses that are designed to remove the specific stresses that modified and activated OxyR in the first place. For example, oxidation of OxyR by H2O2 induces the OxyR-regulated gene katG, which encodes for catalase that effectively removes H2O2, whereas S-nitrosylation of OxyR induces an unknown denitrosylase enzyme that removes SNOs (Fig. 3A). More specifically, OxyR binds DNA as a dimer of dimers, using a helix-turn-helix motif for binding, with each monomer binding on an adjacent part of the DNA duplex (12, 102). On oxidative activation, OxyR recruits RNA polymerase to specific promoters (55, 90). Alternative post-translational modifications of OxyR at Cys199 (oxidation vs. S-nitrosylation) lead to different conformations (52) that bind to different DNA sequences and activate transcription at distinct promoters (Fig. 1). Oxidation of OxyR Cys199 to sulfenic acid is, in fact, sufficient for transcriptional activation (52), but structural studies have shown further transition of this Cys199 sulfenic acid to a disulfide bond between Cys199 and Cys208 (12). S-nitrosylation and S-sulfenylation of Cys199 lead to different structural changes in OxyR, as revealed by circular dichroism and by differences in DNA-binding affinity (52). Thus, these different modifications may alter the interactions between OxyR monomers, or the OxyR interaction with specific DNA sequences or with RNA polymerase. A careful structural analysis of each of these modified forms will be required to reveal exactly how the distinct redox-based modifications of a single cysteine in OxyR lead to differential transcriptional activity.
Notably, the OxyR gene itself is not transcriptionally induced by either oxidative or nitrosative stress (90, 97), although its transcription has been shown to be regulated by the growth phase (34). The induction of OxyR-regulated genes is solely a result of OxyR activation by post-translational modification, which is designed for rapid responses.
Role of OxyR in SNO Metabolism Under Aerobic Conditions
OxyR regulates the bacterial response to nitrosative stress not only under anaerobic conditions but also under aerobic conditions. E. coli rapidly metabolizes S-nitrosothiols to nitrate and nitrite aerobically, and it is notable that in the absence of OxyR there is an aberrant accumulation of nitrite when compared with the wild type strain (39) (Fig. 3B). Nitrite is known to be toxic to bacteria (10) and hence may be a factor causing the growth inhibitory effects of SNOs on ΔoxyR strains (39, 90). The exact mechanism by which OxyR influences metabolism of SNOs remains to be elucidated.
Role of OxyR in Regulating S-Nitrosylation During Anaerobic Growth
OxyR-deficient bacteria are sensitized to the growth inhibitory effects of NO/RNS. An E. coli oxyR knockout strain demonstrates growth inhibition during ARN (as well as after exposure to NO/SNO under aerobic conditions) (39, 90). When grown under ARN in the absence of OxyR, E. coli show overall increases in protein S-nitrosylation (SNO
Enzymatic S-Nitrosylation in E. coli
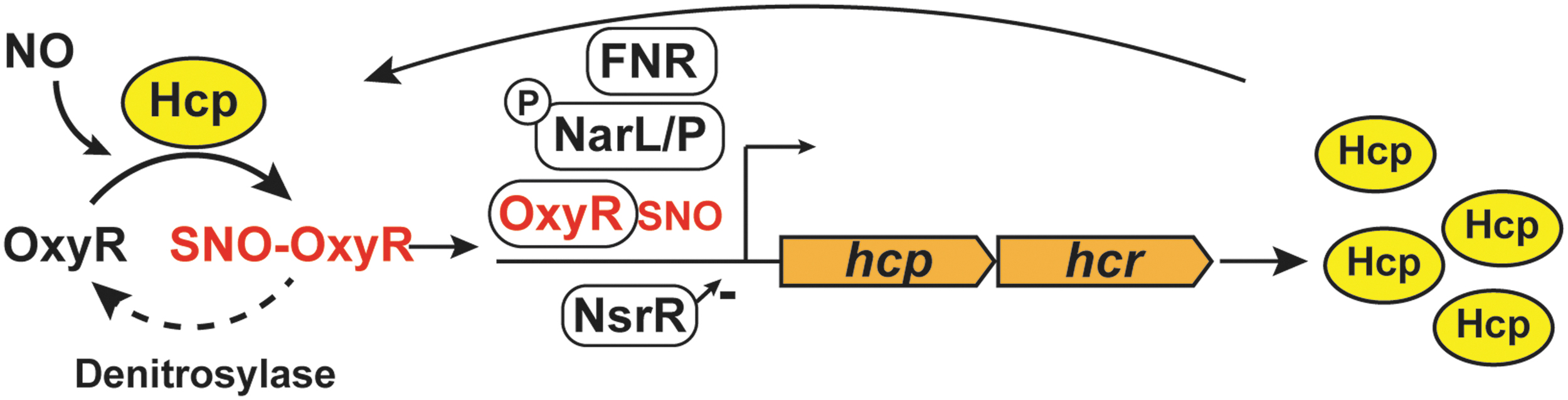
One member of the OxyR regulon, the hybrid cluster protein Hcp, is a critical element of a three-component enzymatic machinery that is responsible for specific and regulated S-nitrosylation of proteins during ARN (91). Hcp is one of the maximally induced genes when E. coli are grown on nitrate, and this induction is abrogated in the absence of OxyR. Hcp functions as a protein S-nitrosylase that mediates de novo synthesis of SNO from NO produced by nitrate reductase (NarG) (91). Hcp in E. coli contains a [2Fe-2S] cluster as well as a [4Fe-2S-2O] “hybrid” cluster (106), which are essential for its SNO-synthase activity (91). In fact, prior treatment of cells with the divalent metal cation chelator 1,10-phenanthroline decreases the formation of overall SNOs during ARN (90). The iron clusters can serve as electron acceptors for the conversion of NO to NO+, and dinitrosyl iron complexes (DNICs) have been shown to support the oxidative requirements for S-nitrosylation by NO (7, 57, 95, 107). Mechanistically, Hcp S-nitrosylates itself and then dimerizes in the presence of NO. The Hcp dimer serves as a nucleus for a large interactome; SNO-Hcp is then able to S-nitrosylate multiple partner proteins, including OxyR. The Hcp interactome also includes a class of enzymes known as transnitrosylases: proteins such as glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and TrxA that can transfer their own SNO to additional substrates through direct interactions. Thus, under anaerobic growth conditions that generate endogenous NO, Hcp promotes the S-nitrosylation of OxyR, which is, in turn, activated to protect against nitrosative stress, as well as S-nitrosylation of many other proteins required for growth (Fig. 4). For example, Hcp-dependent S-nitrosylation regulates the activity of key metabolic enzymes, including dihydrolipoamide dehydrogenase (LpdA). Lack of Hcp also profoundly inhibits swimming motility during ARN (91).

Even though the induction of Hcp in ARN is eightfold lower in the oxyR knockout strain, Hcp remains basally transcribed in the presence of nitrate independent of OxyR, ensuring that OxyR can become nitrosylated rapidly when NO is first present (90). The major transcription factors regulating gene expression during ARN include fumarate-nitrate regulation (FNR) and nitrate/nitrite response regulator proteins (NarL/NarP). FNR activates transcription of genes required during anaerobiosis and inhibits transcription of genes required for aerobic growth (49, 87), whereas NarL/NarP respond to levels of available nitrate to induce genes required for nitrate metabolism such as nitrate reductase, NarG (78). It is interesting to note that besides OxyR, transcription of hcp is regulated by FNR and NarL/NarP, in response to anaerobiosis and nitrite/nitrate, respectively (28). In addition, in the presence of NO, repression of hcp by the transcriptional repressor HTH-type transcriptional repressor (NsrR) is relieved (27). Besides hcp, the NsrR regulon also includes several other genes that protect against NO and RNS, such as hmp (27). In sum, hcp is part of the protective machinery against NO: basal levels of Hcp are controlled during ARN by FNR, NarL, NarP, and NsrR; whereas OxyR regulates further induction of Hcp to create a positive feedback loop (Fig. 5).
Until recently, S-nitrosylation in bacteria had been studied in the context of nitrosative stress, with no consideration for any role in signaling. Bacterial responses to NO/SNO were viewed as aimed at removing NO or SNO and mitigating cellular injury. These responses involved enzymes that directly consume NO, namely flavohemoglobin (Hmp) (38) and flavorubredoxin (NorV) (33), or that metabolize SNOs (GSNOR) (61). Our new data suggest a different perspective: The growth advantage conferred by Hcp is mediated not only by the removal of NO/SNO but also by promoting S-nitrosylation of specific proteins that confer benefit on metabolism and growth (Fig. 4), reflective of a primary signaling function for NO that is analogous to its role in yeast, plants, and mammals (1, 41, 116). Further, it should be noted that mitochondria are, of course, major players in cellular metabolism. Thus, endogenous signaling in bacteria draws analogy to the metabolic function of NO in mammalian cells. Inasmuch as signaling by S-nitrosylation is enzymatic in E. coli, one might anticipate the same in mammalian cells. Indeed, although Hcp does not have homologues in higher organisms, Fe-S cluster center-containing enzymes are commonplace in mitochondria, as discussed in the Regulation of OxyR S-Nitrosylation section.
Regulation of OxyR S-Nitrosylation
As noted earlier, S-nitrosylation of OxyR is enzymatically regulated by Hcp. However, SNO-OxyR levels are also regulated by an unidentified mechanism of denitrosylation, consistent with an enzymatic process. That is, after E. coli treatment with an exogenous NO donor, S-nitrosylated OxyR is rapidly denitrosylated (90). Additional studies will be required to determine whether this denitrosylation is due to known denitrosylases, including the thioredoxin system [reviewed in Benhar et al. (6)] or SNO reductases (1, 61), or to novel enzymes.
Role of the OxyR-Dependent Anaerobic Regulon
A gene ontology pathway analysis of the anaerobic OxyR regulon using STRING (101) revealed that these genes belong to pathways ranging from cellular respiration and amino acid biosynthesis to stress responses (Table 1). Many of these genes are metabolic enzymes involved in respiration, and they may be upregulated to maintain normal metabolism and growth. For example, NO inhibits branched-chain amino acid synthesis by inhibiting the enzyme IlvD, leading to auxotrophy (42). Upregulation of multiple genes in the branched-chain amino acid synthetic pathway (ilvA, ilvD, ilvE, ilvM, ilvN, leuB, and leuD) (Table 1) may, therefore, compensate for this inhibition.
Gene Ontology Pathway Enrichment Analysis of OxyR Regulon Genes
The OxyR-dependent genes induced under anaerobic growth on nitrate were grouped by biological processes by using STRING.
Interestingly, the SNO-OxyR regulon also includes genes with anti-oxidative functions (katG, catalase; msrB, methionine sulfoxide reductase; lpdA, lipoamide dehydrogenase; tpx, thiol peroxidase) (Table 1). One possibility is that these genes have yet-to-be discovered anti-nitrosative functions. This idea is supported by the fact that strains deficient in tpx or lpd have significant decreases in SNO levels during ARN (91). Another possibility is that the regulon is generally aimed at counteracting host defenses during infection, and it therefore provides coverage against both nitrosative and oxidative stresses.
OxyR-Dependent Regulons Under Different Growth Conditions
Facultative bacteria such as E. coli precisely and specifically regulate gene expression depending on growth conditions (aerobic vs. anaerobic) and availability of electron acceptors (nitrate, fumarate, dimethyl sulfoxide, or trimethylamine oxide) (105). The transcription factor FNR is required for the expression of many genes that are necessary for anaerobic metabolism, whereas aerobic respiration control protein regulates genes for aerobic metabolism. Other transcription factor systems such as NarL/P and nitrate/nitrite sensor proteins activate gene transcription in the presence of nitrate. Many bacterial genes have binding sites for multiple transcription factors, allowing complex regulation by growth conditions.
It is highly likely that the complete OxyR regulons, both oxidative and nitrosative stress specific, are much larger than those currently identified because the bacterial transcriptome can be very different under different growth conditions. One such example has been reported for the cytochrome c peroxidase (ccp) gene, which is only expressed under anaerobic conditions (because it requires FNR for expression), but it is further induced by H2O2 in an OxyR-dependent manner (50). Transcriptome analysis of H2O2-treated wild type and ΔoxyR E. coli growing anaerobically will undoubtedly identify additional genes that are regulated such as ccp. By the same token, the nitrosative stress-specific OxyR regulon was identified under anaerobic conditions in E. coli growing on nitrate (91). We expect exogenous NO to lead to a distinct transcriptional response, and additional SNO-OxyR targets likely will be identified in cells growing anaerobically with fumarate or DMSO as electron acceptors. Also, different sources of nitrosative stress, endogenous versus exogenous (NO donors or SNO donors), will likely generate varied transcriptional responses (48, 66, 90). Further, host innate immune responses can generate both oxidative and nitrosative stress at the same time, which should activate both oxidized-OxyR and SNO-OxyR regulons. Since OxyR operates as a multimer, combinatorial effects of SNO and SOH modifications of OxyR might activate additional sets of genes.
Role of OxyR Under Anaerobic/Microaerobic Conditions Within the Host
During infection of multicellular hosts with low oxygen/anaerobic microenvironments, facultative bacteria (e.g., E. coli) can transition from aerobic to anaerobic respiration. For example, during urinary tract infections (UTI), the concentration of oxygen within the bladder lumen is insufficient for aerobic respiration (62, 86), requiring invading bacteria traveling up the urethra to switch from aerobic to anaerobic metabolism. Various studies have shown that hmp (flavohemoglobin), norV (flavorubredoxin), and hcp (hybrid cluster protein), genes that are known to protect against nitrosative stress (33, 38, 90), are induced in uropathogenic strains during UTI (64, 86, 100), implying that bacteria are exposed to sufficient NO during UTI to induce SNO-OxyR-dependent transcription. Despite the pathophysiological relevance of low tissue oxygen concentrations in the presence of nitrate, this feature of infection is frequently neglected.
In the large bowel, which is generally believed to be anaerobic, it has been shown that the ability to respire on nitrate provides facultative anaerobes with an advantage compared with strictly aerobic or strictly anaerobic species, or to facultative anaerobes in which nitrate reductase is not induced (110). These conditions of anaerobic growth in the presence of nitrate are analogous to the conditions under which the induction of a large regulon by SNO-OxyR has been documented (90). Further, OxyR has been shown to be essential for growth in various mouse models of infection that are characterized by anaerobic/microaerobic conditions that are likely rich in nitrate (derived from host RNS), including intra-abdominal abscess, UTI, and sepsis (47, 99). Although OxyR function has been ascribed to protection from host-derived ROS, deletion of the neutrophil phagocyte oxidase, a major source of host-derived ROS, did not rescue the ΔoxyR phenotype in UTI models (46). Thus, it is likely that OxyR is also serving to protect against nitrosative stress under those conditions.
NO Production in Mitochondria Parallels Anaerobic NO Generation in E. coli
Recent studies suggest many parallels between anaerobic NO generation in bacteria and NO production by mitochondria, including the potential for enzymatic generation of SNOs. By way of reminder, nitric oxide synthases (NOSs) are the major source of aerobic NO generation in mammalian cells (30). A mitochondrial nitric oxide synthase (NOS) has also been described in the inner mitochondrial membrane (5), but studies regarding its activity have led to conflicting results and its existence has been questioned (56). In 1999, Kozlov et al. reported on an NOS-independent source of mitochondria-generated NO (54) under hypoxic conditions, whereby nitrite undergoes a one-electron reduction by the respiratory chain (11, 54). Different proteins in the respiratory chain have been proposed to be the site of NO production, including ubiquinone/cytochrome bc1 (69) and cytochrome c oxidase (11). In such cases, NO generation was observed under hypoxic conditions when the respective enzymes utilize nitrite instead of oxygen as an electron acceptor (11, 69). Other mitochondrial proteins with nitrite reductase activity include cytochrome c (4) and mitochondrial amidoxime reducing component proteins (94). Generation of NO in mitochondria is, thus, remarkably similar to anaerobic respiration in E. coli, where NOS-independent production of NO occurs due to a secondary activity of nitrate reductase under anaerobic conditions, as described earlier.
S-Nitrosylation in Mitochondria and Its Role in Regulating Mitophagy
In the earlier days of NO biology, S-nitrosylation was believed to require oxygen. Studies of the mitochondrion were the first to establish otherwise: Proteins were found to become S-nitrosylated in anoxic mitochondria after addition of NO (31). Many mitochondrial proteins are now known to be regulated by S-nitrosylation (13, 23, 74). These include enzymes involved in the tricarboxylic acid cycle, fatty acid synthesis, and the electron transport chain (13, 23). How proteins are S-nitrosylated in the mitochondria, however, is not well understood. One model is that protein-bound DNICs, formed from nitrite-derived NO, could act as SNO synthases in a manner similar to Hcp in E. coli.
Mitochondrial DNA (mtDNA) encodes for 13 subunits of the respiratory chain, 2 rRNAs and 22 tRNAs, independently of nuclear DNA (2), and therefore mtDNA transcription will regulate mitochondrial oxidative phosphorylation. Two mitochondrial transcription factors encoded by nuclear DNA, transcription factor A, mitochondrial (TFAM) and dimethyladenosine transferase 2, mitochondrial, as well as an RNA polymerase, DNA-directed RNA polymerase, mitochondrial are known to subserve mitochondrial gene expression [reviewed in D'Souza and Minczuk (20)]. Although S-nitrosylation of these proteins has not been studied, Hsp60, a chaperone required for the transport of TFAM to the mitochondria, is S-nitrosylated by iNOS, thereby facilitating its binding to TFAM and its transport to mitochondria (98). mtDNA transcription is also regulated by additional SNO-proteins, including cAMP response element-binding protein (21, 93) and signal transducer and activator of transcription 3 (51, 63), although the exact effect of S-nitrosylation on mitochondrial transcription is unclear. It is quite possible that akin to OxyR, a mitochondrial protein(s) could act as a sensor of both SNO and S-oxidation to regulate transcriptional responses. This is an especially tantalizing idea given that mitochondria generate superoxide/H2O2 versus NO under high and low oxygen tension, respectively.
Although there is no mitochondrial protein homologous to Hcp, the features and components of endogenous enzymatic S-nitrosylation observed in E. coli are present in mitochondria. A hypothetical enzymatic cascade in mitochondria akin to the one described in E. coli is shown in Figure 6. In this scenario, NO is generated in mitochondria by respiratory chain enzymes under hypoxic conditions (11, 54). Further, experiments done in mice demonstrated that the NO produced in mitochondria from dietary nitrite is associated with the formation of DNICs (25, 103) and SNOs (103). Also, GAPDH has been demonstrated within mitochondria of mouse hearts and exhibits transnitrosylase activity there (53). SNO-GAPDH may be transported into mitochondria (53), or possibly S-nitrosylated in situ by a DNIC-containing protein acting as an “SNO-synthase.” Thus, all three components required for de novo S-nitrosylation, namely an enzymatic NO source, a potential SNO-synthase, and a transnitrosylase, are present within mitochondria, as they are in E. coli.
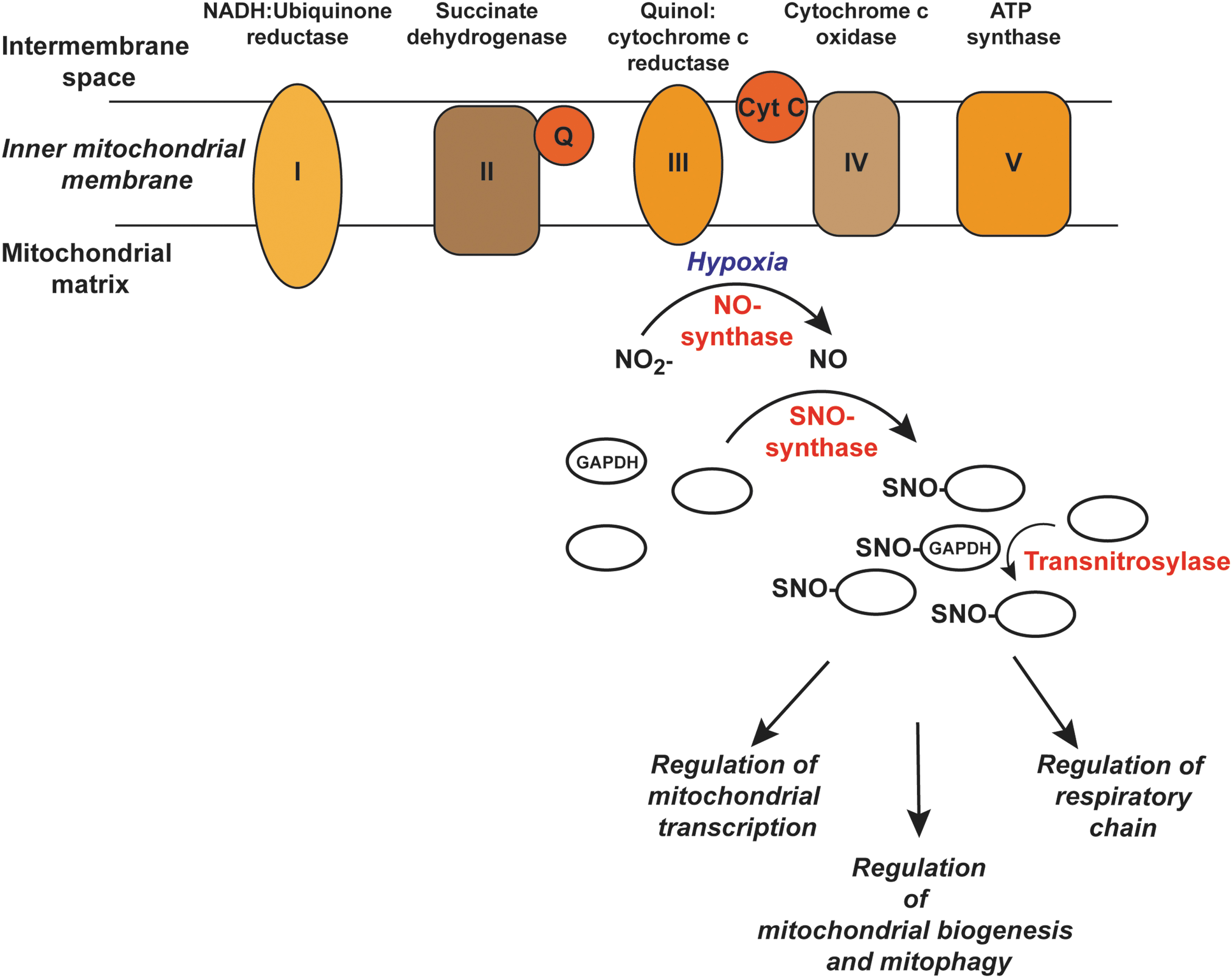
Mitochondria are dynamic, and they can undergo regulated proliferation, fission, fusion, and degradation (mitophagy) (104). Mitophagy refers to the selective autophagic removal of mitochondria (59), and regulated mitophagy is essential for normal cellular functioning whereas its dysregulation can lead to pathophysiology (73). One important pathway that mediates mitophagy is the PTEN-inducible putative kinase 1 (PINK1)-Parkin pathway whereby accumulation of PINK1 in damaged mitochondria leads to the phosphorylation and recruitment of the ubiquitin E3 ligase Parkin to the mitochondrial outer membrane to promote the recruitment of autophagy receptors (58, 73, 115). S-nitrosylation of Parkin was first reported in neurons and was associated with Parkinson's disease (16, 114). One study reported that Parkin S-nitrosylation inhibits ubiquitin ligase activity (16), whereas another showed that S-nitrosylation of Parkin at an alternate site (Cys323) activates the E3 ligase leading to enhanced mitophagy (72). PINK1 is also S-nitrosylated and this inhibits its protein kinase activity, leading to impaired mitophagy (70). Studies with mice lacking the GSNOR enzyme and in aging humans in whom GSNOR levels decline show higher SNO-levels and increased S-nitrosylation of Parkin (81) that is associated with mitochondrial dysfunction. Overall, these data suggest that excessive S-nitrosylation inhibits mitophagy, which may have implications not only for aging and Parkinson's but also for Alzheimer's disease where aberrant S-nitrosylation (e.g., of dynamin-related protein-1) has also been implicated in mitochondrial dysfunction. These interesting data notwithstanding, the key SNO-players in mitophagy and mitochondrial dysfunction remain to be identified. Such an understanding should lead to a more targeted approach to regulating mitochondrial S-nitrosylation therapeutically.
Conclusion
Although OxyR has served as a classic model for understanding the cellular response to oxidative stress, the physiological role of OxyR in situ also includes protection against nitrosative stress. Thus, E. coli has evolved a common transcription element in OxyR to protect itself against both aerobic and anaerobic stressors. In effect, facultative anaerobes such as E. coli can sense different stresses through alternative modifications in OxyR and tailor their genetic responses accordingly. This newly discovered role for OxyR in regulating anaerobic stress has also revealed new physiological functions mediated by S-nitrosylation, as well as enzymatic machinery subserving endogenous S-nitrosylation. Most notably, OxyR-regulated, Hcp-mediated S-nitrosylation is required for bacterial motility and metabolism under anaerobic conditions. Further examination of genes in the nitrate-dependent OxyR regulon may reveal targetable vulnerabilities in these strategies employed by bacteria to survive infections on the one hand and loci of dysfunction that underlie human disease on the other hand. Enzymes first characterized in bacteria that regulate S-nitrosylation may serve as models for identifying and understanding homologous or comparable mechanisms present in mammalian systems. Given that dysregulated S-nitrosylation of specific SNO-proteins is now recognized as a factor in many diseases, including neurological disorders mediated by mitochondrial dysfunction, elucidation of enzymatic mechanisms involved in S-nitrosylation/denitrosylation may suggest new therapeutic approaches to human disease.
Footnotes
Funding Information
This work was supported by the National Institutes of Health under grants GM099921, HL075443, HL128192, HL126900, and DK119506.