
Abstract
Aims:
Delphinidin (DEL) is a plant-derived antioxidant with clinical potential to treat inflammatory pain but suffers from poor solubility and low bioavailability. The aim of the study was to develop a well-tolerated cyclodextrin (CD)–DEL complex with enhanced bioavailability and to investigate the mechanisms behind its antinociceptive effects in a preclinical model of inflammatory pain.
Results:
CD–DEL was highly soluble and stable in aqueous solution, and was nontoxic. Systemic administration of CD–DEL reversed mechanical and heat hyperalgesia, while its local application into the complete Freund's adjuvant (CFA)-induced inflamed paw dose-dependently reduced mechanical hyperalgesia, paw volume, formation of the lipid peroxidation product 4-hydroxy-2-nonenal (4-HNE), and tissue migration of CD68+ macrophages. CD–DEL also directly prevented 4-HNE-induced mechanical hyperalgesia, cold allodynia, and an increase in the intracellular calcium concentration into transient receptor potential ankyrin 1 expressing cells. Both 4-HNE- and CFA-induced reactive oxygen species (ROS) levels were sensitive to CD–DEL, while its capacity to scavenge superoxide anion radicals (inhibitory concentration 50 [IC50]: 70 ± 5 μM) was higher than that observed for hydroxyl radicals (IC50: 600 ± 50 μM). Finally, CD–DEL upregulated heme oxygenase 1 that was prevented by HMOX-1 siRNA in vitro.
Innovation:
In vivo application of DEL to treat inflammatory pain is facilitated by complexation with CD. Apart from its antioxidant effects, the CD–DEL has a unique second antioxidative mechanism involving capturing of 4-HNE into the CD cavity followed by displacement and release of the ROS scavenger DEL.
Conclusion:
CD–DEL has antinociceptive, antioxidative, and anti-inflammatory effects making it a promising formulation for the local treatment of inflammatory pain.
Color images are available online.
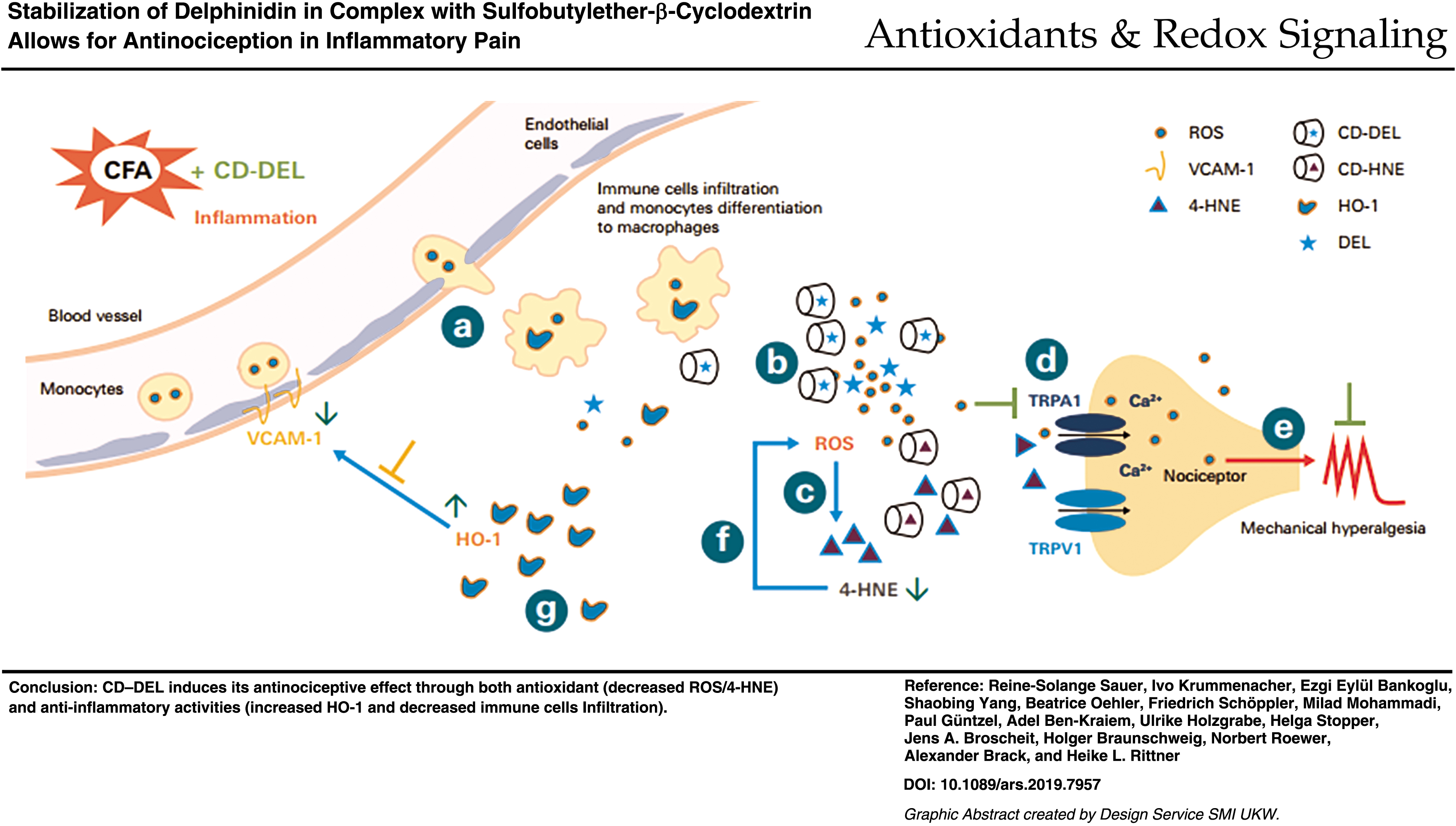
Innovation
Delphinidin (DEL) is a plant pigment from anthocyanin family, which has been associated with different beneficial properties on health and diseases. Its clinical use is limited by its poor bioavailability and lack of solubility in aqueous solution, hence its antinociceptive effect has not been investigated. We have developed a complex of DEL and cyclodextrin (CD) to overcome those problems. For the first time, we demonstrate that CD–DEL complex decreases hyperalgesia in rats with complete Freund's adjuvant-induced inflammatory pain through both antioxidative and anti-inflammatory pathways. The mechanism of action involved was the displacement and release of DEL from the CD cavity by 4-hydroxy-2-nonenal.
Introduction
Acute and chronic pain conditions adversely affect quality of life and significantly burden health care systems. Inflammatory pain arises from proalgesic mediators including reactive oxygen species (ROS) (e.g., superoxide [O2
ROS, 4-HNE as well as exogenous irritants such as allyl isothiocyanate (AITC) are well-known agonists of the transient receptor potential ankyrin 1 (TRPA1) expressed on peripheral nociceptors. Their activation increases the intracellular calcium concentration (49, 50) and thereby pain sensitivity. TRPA1 is implicated in mechanical and cold hypersensitivity observed in a variety of models of inflammatory, neuropathic, and cancer pain (10, 55). Better management of oxidative stress by supplementation of antioxidants has the potential to decrease ROS/4-HNE and activation of nociceptors via calcium influx to relieve pain.
Anthocyanins are a group of plant-derived compounds that are low in cost and nontoxic. They possess chemotherapeutic, cardio-, neuro-, hepatoprotective, and antidiabetic properties (4). Because of their diverse mechanisms of action, anthocyanins might be suitable therapeutic agents against various inflammatory conditions. The polyphenolic compound delphinidin (DEL) is found in pigmented fruits and vegetables, and is a potent antioxidant (38). It is one of the most abundant dietary anthocyanins, and possesses strong anti-inflammatory and anticancer properties (1, 23, 30).
The solubility, chemical stability, and bioavailability of poorly soluble drugs like DEL can be significantly improved by cyclodextrins (CDs) and their derivatives, which are widely used in drug formulation and development (19). CDs have no obvious effects on the cardiovascular or respiratory system, nor on function of the autonomic and somatic nervous systems in animals complete Freund's adjuvant (CFA)-induced inflammation (62). In this study, DEL was complexed with various CDs (e.g., sulfobutylether [SEB] β-CD) to identify the most soluble complex. However, the molecular mechanisms of the antinociceptive effects of CD–DEL have not yet been characterized.
During oxidative stress, cells develop adaptive protection mechanisms to maintain homeostasis. Heme oxygenase 1 (HO-1) is an oxidative-sensor protein induced by ROS generation (31) and tightly regulates intracellular ROS levels (42). However, the accumulation of ROS can have harmful effects when they exceed endogenous antioxidant defense capacity. Under these conditions, exogenous control is required.
Phytochemicals recruit the antioxidative stress pathways. This involves the synthesis of HO-1 (2, 33, 68, 72, 73), which plays an active role in the resolution of inflammation (13) and peripheral antinociception (52, 57) as well as facilitating antioxidant activity (7, 34). Moreover, the protective effects of DEL against oxidative stress have been demonstrated (2, 29, 47). Therefore, we hypothesized that the CD–DEL complex induces peripheral antinociceptive effects through both its antioxidant and anti-inflammatory activities by decreasing ROS/4-HNE and increasing HO-1 levels.
Results
Solubility and stability of CD–DEL
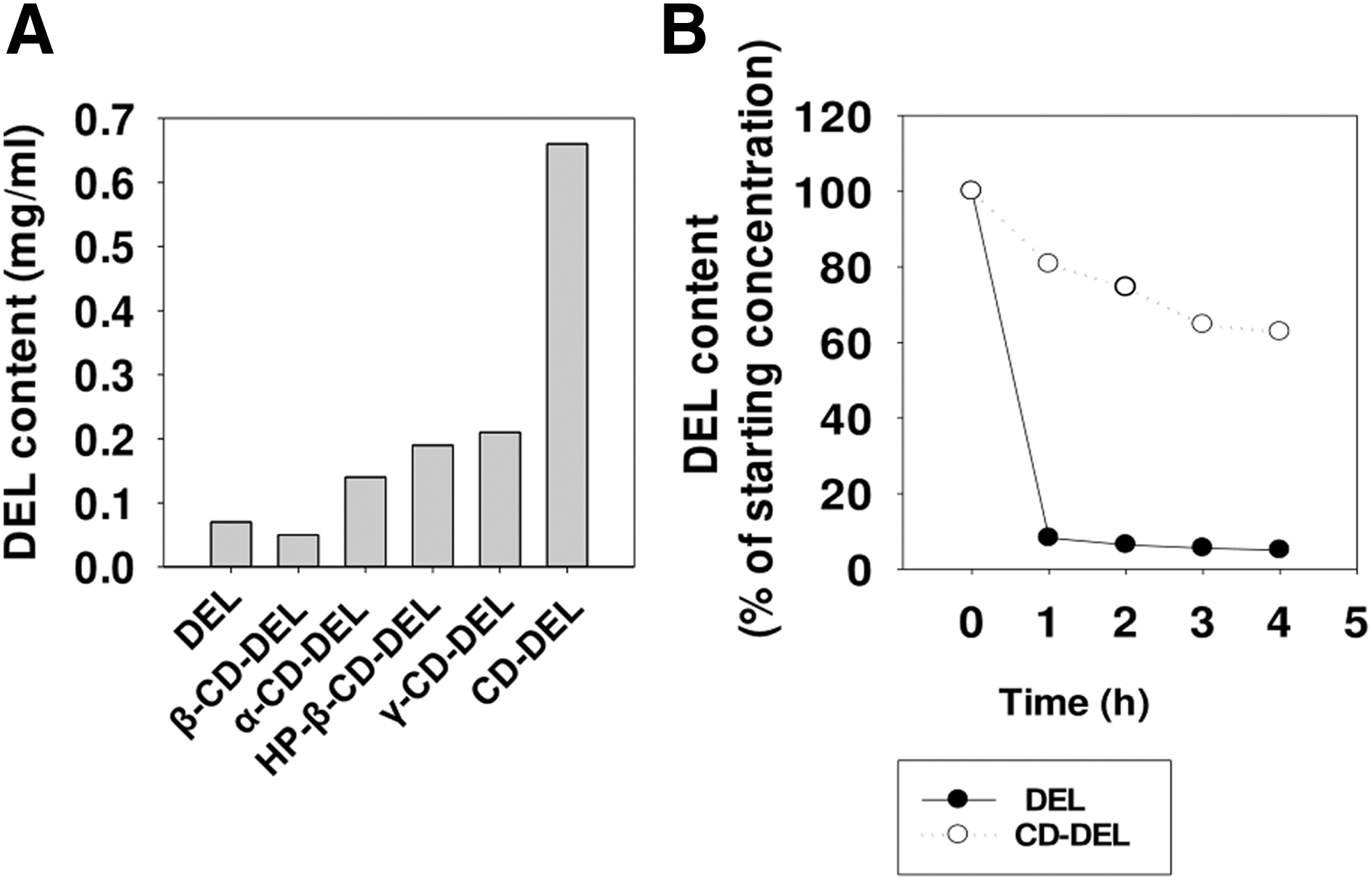
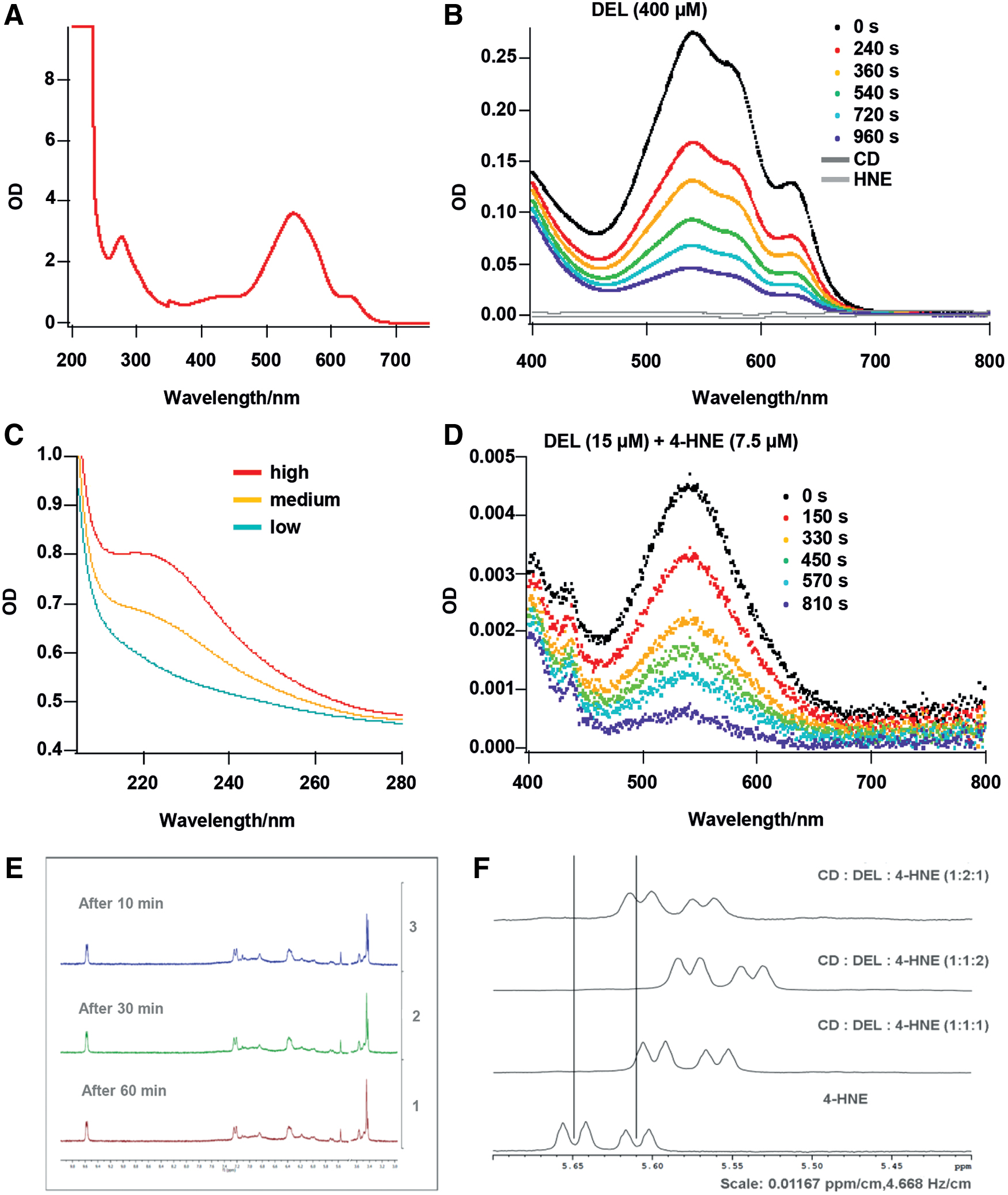
Complexation of DEL with several CDs at different pHs revealed that at pH 4–5, a higher concentration of the complex in aqueous solution could be established. Figure 1A shows the very low solubility of DEL in water alone. Therefore, the solubility of DEL in aqueous CD solutions containing β-CD, α-CD, hydroxypropyl (HP)-β-CD, γ-CD, and SEB-β-CD (here CD) was assessed. Among the different CDs investigated, the highest solubility was found with CD. Furthermore, the stability of DEL with CD in aqueous solution and DEL content were determined after 1, 2, 3, and 4 h. Figure 1B displays the calculated content as the percentage of the starting concentration. CD–DEL was more stable in aqueous solution compared with uncomplexed DEL. Based on the improved solubility of CD–DEL and the safety profile of CD (32 ,44, 53), the CD–DEL complex was selected for further experiments.
Antinociceptive properties and time-dependent concentration of CD–DEL in inflamed paw and plasma
Baseline paw pressure thresholds were obtained before CFA injection in each group. At 4 days CFA inflammation, intraplantar (
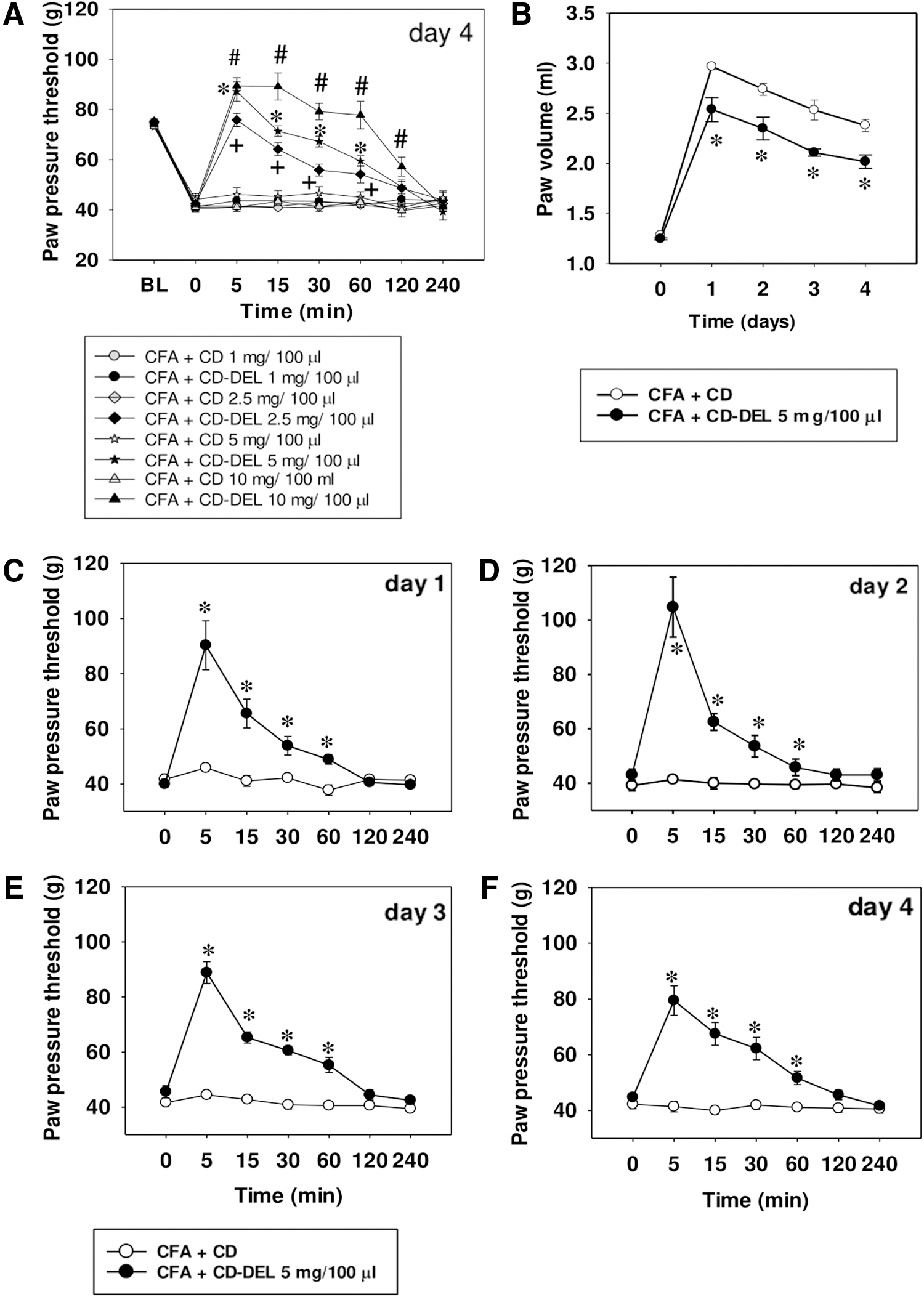
This dose time-dependently decreased paw volume by 15.18% ([(paw volume CFA + CD − paw volume CFA CD–DEL × 100)/paw volume CFA-CD]; CD: 2.37 ± 0.06 and CD–DEL: 2.02 mL ±0.07 p = 0.001) (Fig. 2B), suggesting an anti-inflammatory effect. By daily application, CD–DEL effectively reversed mechanical hyperalgesia each day to the same extent without altering baseline hyperalgesia (Fig. 2C). However, CD–DEL did not affect the heat hyperalgesia after daily injection from days 1 to 4 (Supplementary Fig. S1A–D). No differences in motor function as indicated by the Rota-Rod test were observed (Supplementary Fig. S1E). Furthermore, cold allodynia was significantly delayed after administration of CD–DEL in 4 days CFA-induced inflammation (Supplementary Fig. S2A). Both groups had a similar prolongation of the time spent on the Rota-Rod as a training effect. These results suggest that even local CD–DEL treatment can induce potent antinociception without the development of any tolerance.
To specifically activate TRPA1 activator, the irritant AITC, the main component of mustard oil, was investigated for a peripheral antinociceptive effect of CD–DEL 5 mg. Baseline before injection was 75.5 g ± 0.56. Intraplantar injection of AITC caused mechanical hyperalgesia that was time-dependently reversed by CD–DEL. Significant analgesia started from 1 h (87.0 g ± 3.03, p < 0.001) and persisted for 6 h (69.2 g ± 1.3, p < 0.001) (Supplementary Fig. S2B). In contrast, CD–DEL did not affect heat paw withdrawal latencies in AITC-induced hyperalgesia (Supplementary Fig. S2C
We further investigated whether the effect of CD–DEL is site specific. Systemic application of 12.5, 25, and 50 mg, but not 5 mg CD–DEL, dose-dependently reduced mechanical hyperalgesia after 5 min (Fig. 3A): (5 mg vs. 12.5 mg, p < 0.001; 12.5 mg vs. 25 mg, p < 0.001; 25 mg vs. 50 mg, p = 0.002). A significant difference was observed between all tested doses and the control groups until 60 min. A small increase in paw pressure thresholds was still observed after 240 min with the highest dose.
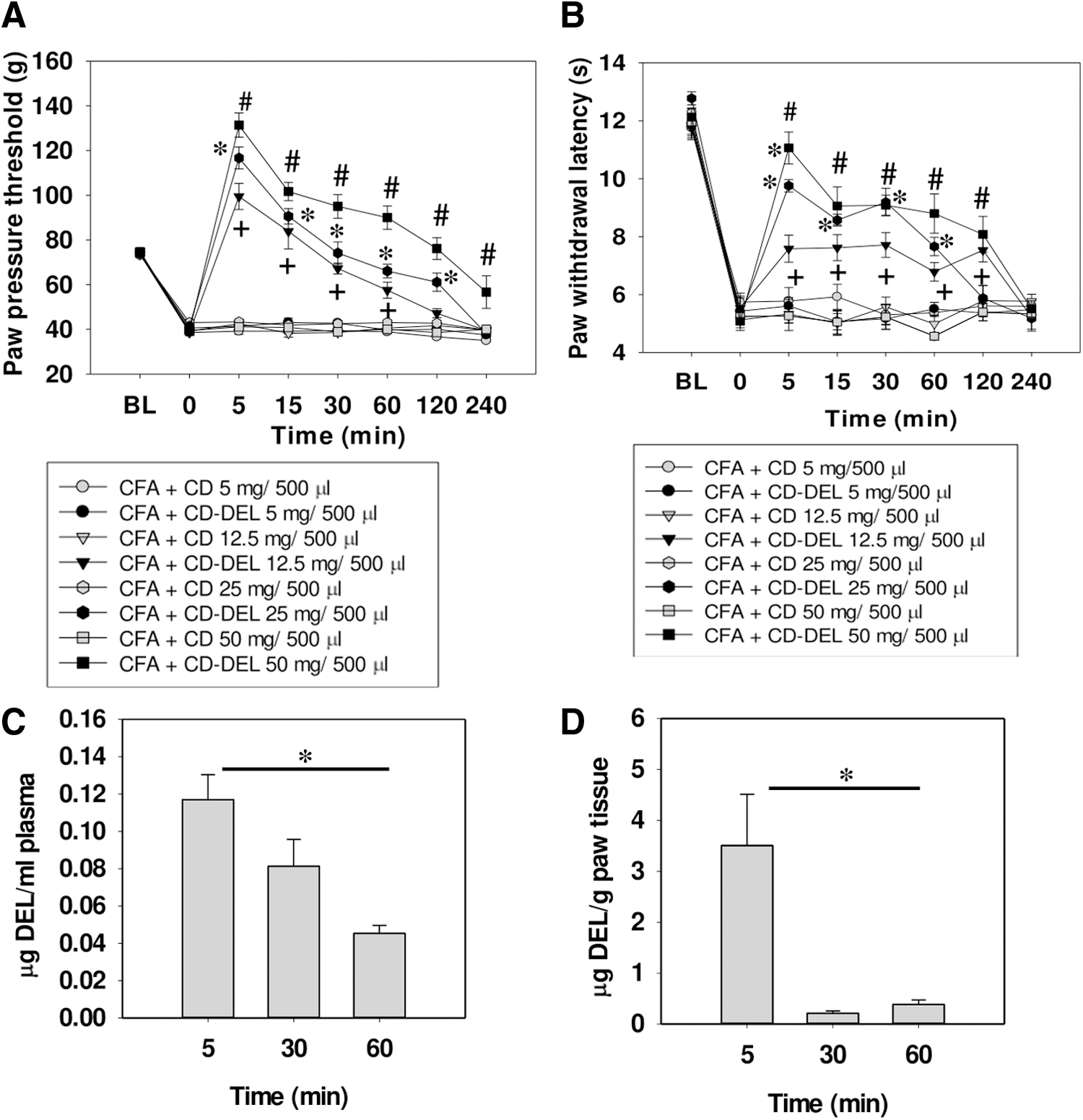
Systemic application of CD–DEL also dose-dependently decreased heat hyperalgesia after 5 min (Fig. 3B
To evaluate the pharmacokinetics, the concentration of DEL was measured at time points with significant antinociceptive effects, in the inflamed plasma and paw, respectively, after systemic and local administration of CD–DEL. After intraperitoneal (i.p.) application of CD–DEL, with a five times higher dose (25 mg), only very low plasma concentration levels of DEL were reached after 5 min (0.1 2 μg/mL ±0.01) (Fig. 3C). In contrast, when the animals were treated intraplantarly, the concentration of DEL in the paw reached the maximum level of 4.99 μg/g ± 1.75 at 5 min, and then significantly decreased after 60 min (0.28 μg/g ± 0.09, p = 0.002) (Fig. 3D). We therefore explored the local mechanisms in more detail.
Decreased 4-HNE in macrophages after local administration of CD–DEL
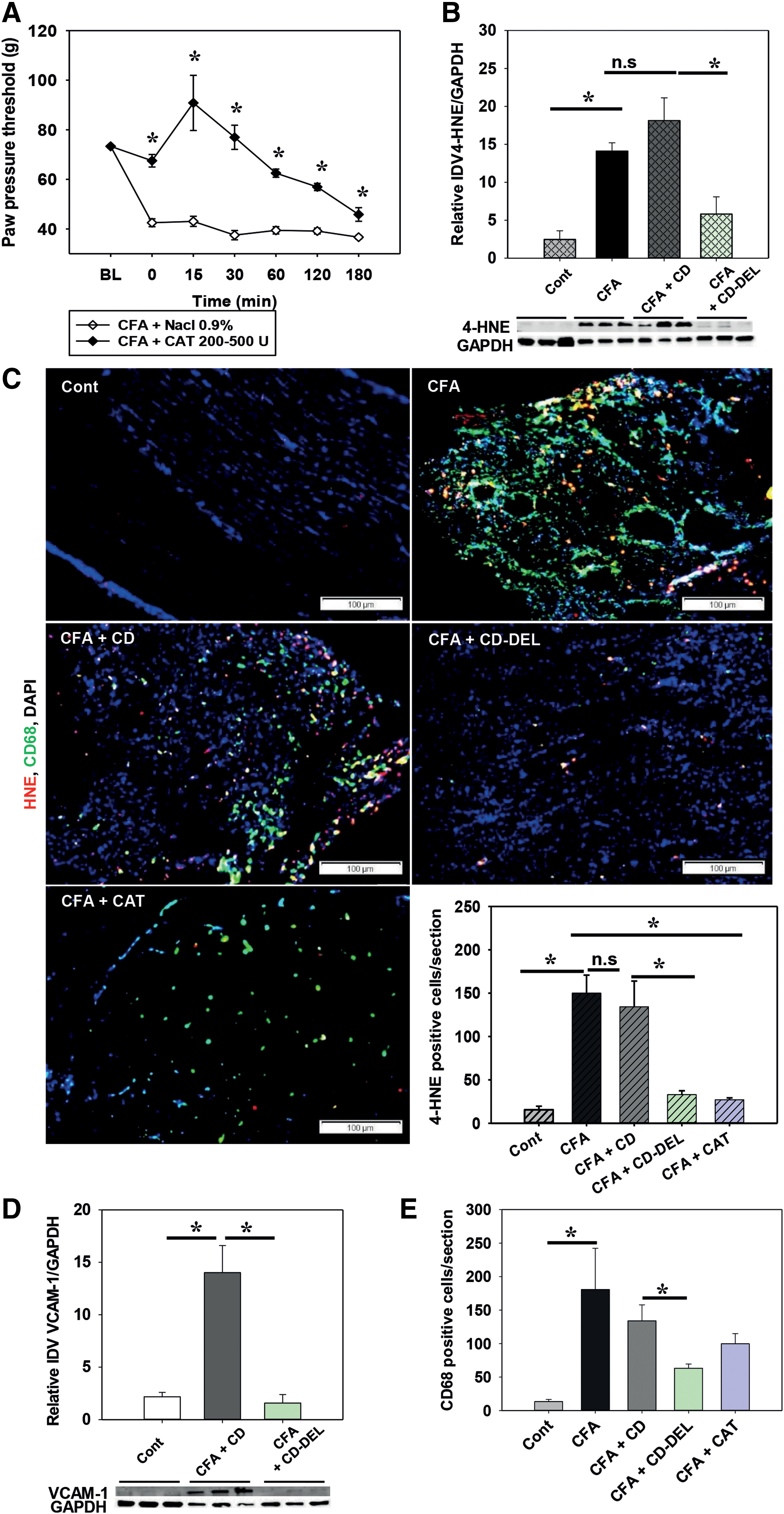
DEL is known for its antioxidant property. Next, we investigated whether hyperalgesia in CFA-induced inflammation is dependent on ROS, and therefore antioxidants, for example, catalase (CAT), relieve hyperalgesia. CAT caused a time-dependent antinociceptive effect and was used as positive control (Fig. 4A, C, and E). This suggested that the antinociceptive effect of CD–DEL could be related to its antioxidant property. Since ROS are unstable and difficult to measure directly, we assessed the level of the lipid peroxidation end-product 4-HNE after CD–DEL in the inflamed paw on day 1 CFA after two CD–DEL injections. This time point after CFA was chosen because (1) the antinociceptive profile was similar from days 1 to 4, and (2) we wanted to analyze effects on early immune cell migration and protein expression. The formation of 4-HNE protein adducts was increased in CFA-induced inflamed paw (integrated density value [IDV] control: 2.5 ± 1.1; CFA + CD: 18.1 ± 3.0, p < 0.001) compared with naïve control and decreased after treatment with CD–DEL (IDV CFA + CD: 18.1 ± 3.0 and CFA + CD–DEL: 5.8 ± 2.3, p = 0.001) but not CD alone (Fig. 4B). Similarly, immunostaining of 4-HNE protein adducts revealed an increase of 4-HNE immunoreactive (IR) cells on day 1 after CFA and a significant decrease in animals treated with CD–DEL (CFA + CD: 134.2 ± 29.7 and CFA + CD–DEL: 33.0 ± 4.4, p < 0.001) (Fig. 4C). A similar effect was seen by CAT treatment used as positive control (CFA: 150.0 ± 20.8 and CFA + CAT: 27.0 ± 2.3, p = 0.020). Since ROS and downstream products are known to be produced by macrophages, we investigated whether adhesion molecules such as vascular cell adhesion molecule 1 (VCAM-1) and immune cell infiltration were affected by CD–DEL. VCAM-1 was increased in the CFA-induced inflamed paw compared with control and decreased by 88.5% after treatment with CD–DEL (IDV CFA + CD: 14.0 ± 2.6 and CFA + CD–DEL: 1.6 ± 0.8, p < 0.001) but not CD alone (Fig. 4D). Similarly, CD–DEL, but not CD, significantly decreased the number of clusters of differentiation 68 (CD68) IR cells after CFA (CFA + CD: 135.0 ± 23.8 and CFA + CD–DEL: 63.6 ± 6.3, p = 0.009) (Fig. 4E). The majority of CD68 IR cells were colocalized with 4-HNE IR cells. Therefore, CD–DEL also reduced the number of CD68/4-HNE IR cells in the inflamed paw by 46.36% (Supplementary Fig. S1F). In summary, CD–DEL reduces monocyte recruitment as well as the formation of 4-HNE in macrophages.
Blockade of 4-HNE-induced hyperalgesia and increased intracellular calcium by CD–DEL
4-HNE is a known TRPA1 activator that induces inflammatory mechanical hyperalgesia and cold allodynia. Both CD–DEL and TRPA1 antagonist HC-030031 time-dependently reversed 4-HNE-induced mechanical hyperalgesia (Fig. 5A): baseline before injection was 82.2 g ± 1.5. Intraplantar injection of 4-HNE induced mechanical hyperalgesia that was time-dependently reversed by CD–DEL. Significant mechanical antinociception by CD–DEL started from 1 h (paw pressure threshold [PPT] 4-HNE + CD: 70.0 g ± 1.7; 4-HNE + CD–DEL: 85.6 g ± 3.3, p < 0.001) and persisted for up to 6 h (PPT 4-HNE + CD: 60 g ± 1.5; 4-HNE + CD–DEL: 75.0 g ± 1.4, p < 0.001). HC-030031 showed a similar effect after 1 h (PPT 4-HNE + Veh: 70 g ± 1.721; 4-HNE + HC-030031: 83.611 g ± 2.17, p < 0.001) and 6 h (PPT 4-HNE + Veh: 54.722 g ± 2.803; 4-HNE + HC-030031: 67.5.0 g ± 1.273, p < 0.001). Interestingly, CD–DEL and HC-030031 also decreased cold allodynia induced by 4-HNE at 1, 3, and 6 h (all p < 0.001, Fig. 5B). No effect of CD–DEL was observed on 4-HNE-induced heat hyperalgesia (Supplementary Fig. S4).
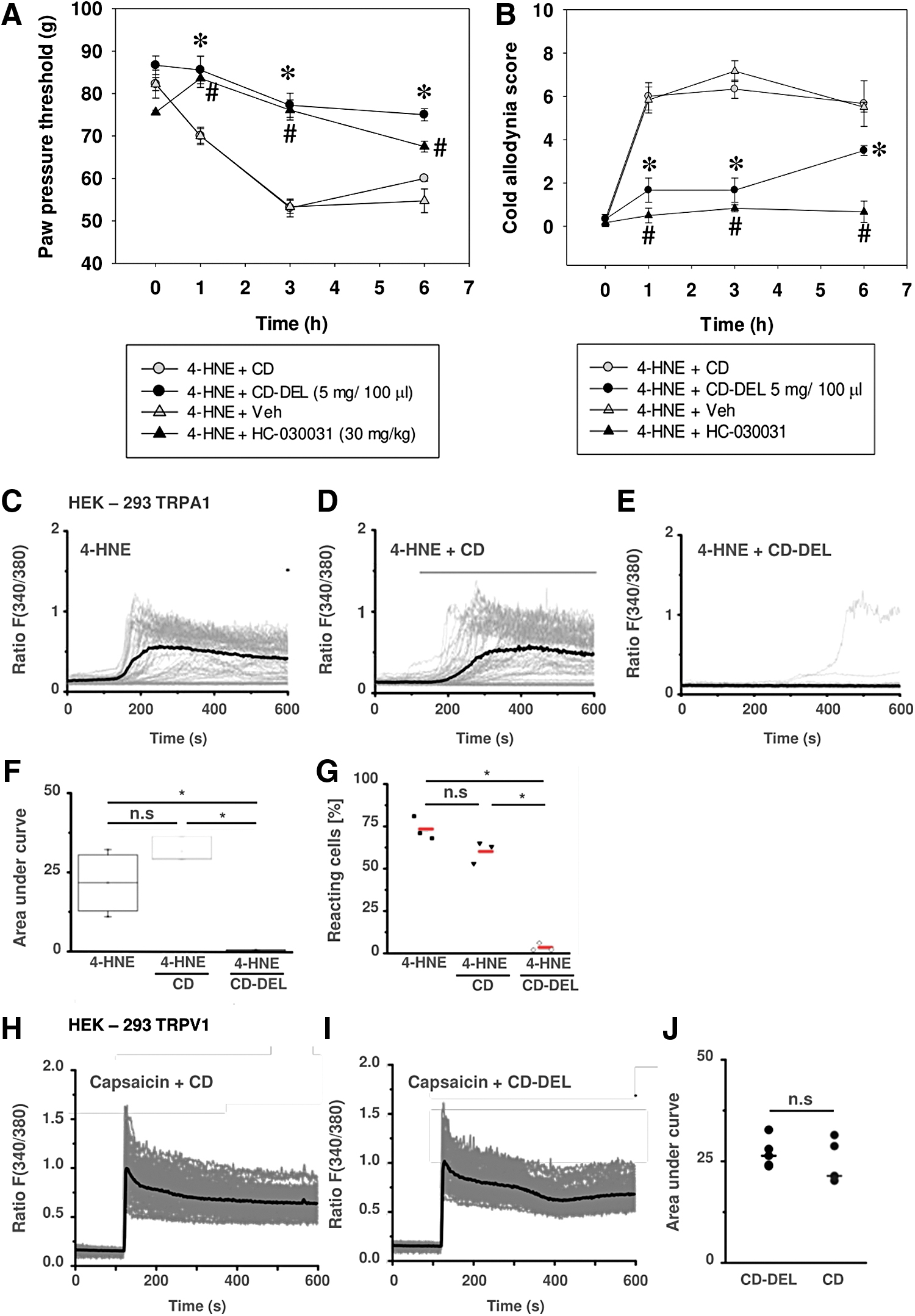
Since 4-HNE activates TRPA1 (68), we wanted to confirm the interaction in vitro. In human embryonic kidney 293 cells (HEK-293) TRPA1, 4-HNE elicited Ca2+ influx that was completely blocked by simultaneous incubation with CD–DEL (Fig. 5C–G). To investigate whether other irritants can also be scavenged, HEK-293 TRPV1 were activated by capsaicin in the presence of CD–DEL (Fig. 4H–J). No block of capsaicin-induced TRPV1 activation by CD–DEL was observed. These data suggest that CD–DEL might specifically interact with 4-HNE or downstream products elicited by 4-HNE.
No direct binding of CD–DEL with 4-HNE but possible competitive action of 4-HNE and DEL for CD cavity
To analyze the interaction between 4-HNE and CD–DEL, we measured absorption spectra at different reaction times. The absorption spectrum of CD–DEL in the ultraviolet (UV)/visible (vis) range is shown in Figure 6A. In addition to the transitions in the range from 450 to 650 nm, a very strong transition in the UV range <250 nm was observed. The absorption spectra of the pure CD–DEL solution showed a time-dependent decrease of the absorption bands in the detectable range of 400–800 nm indicating a decrease in CD–DEL concentration (Fig. 6B). CD and 4-HNE showed no absorption bands in this wavelength range. Next, the time-dependent decrease in absorption of CD–DEL after addition of 4-HNE was measured (Fig. 6D). No significant difference between CD–DEL and CD–DEL +4-HNE was observed (compare Fig. 6B and Fig. 6D). Figure 6C shows a 4-HNE absorption band in a different wavelength range (∼230 nm) that—in principle—could be used to monitor the 4-HNE concentration decrease directly. Unfortunately, due to the weak absorption feature of the 4-HNE, the 4-HNE concentrations used for the 4-HNE + CD–DEL experiment were about a factor of 1000 too low to be detectable. Therefore, we found that the method of UV/vis absorption spectroscopy was not suitable to evaluate the CD–DEL +4-HNE interaction. Although nuclear magnetic resonance (NMR) could then detect time-dependent spectra of 4-HNE, no change in any spectrum was observed when 4-HNE was combined with CD–DEL (Fig. 6E). However, when 4-HNE, CD, and DEL were combined individually, a twofold excess of 4-HNE (in relation to DEL) led to a change in the color of the solution from purple to brownish, which indicates that 4-HNE displaces DEL from the cavity. Interestingly, with a twofold excess or equal molar ratio of DEL, the color remained purple, indicating that DEL stayed in the cavity. The competitive NMR measurement also indicates that 4-HNE and DEL can displace each other in the CD cavity (Fig. 6F). The comparison of the 4-HNE spectrum with the equal molar ratio spectrum (CD:DEL:4-HNE) revealed a shift of the 4-HNE signals to the high field because of the incorporation of 4-HNE into the CD cavity. A twofold excess of 4-HNE shifted the signals even further to the higher field, a twofold excess of DEL instead led to a less pronounced shift to the higher field, which indicates a displacement of 4-HNE from the CD cavity.
Scavenging effect of CD–DEL for superoxide radicals and to a lesser extent to hydroxyl radicals
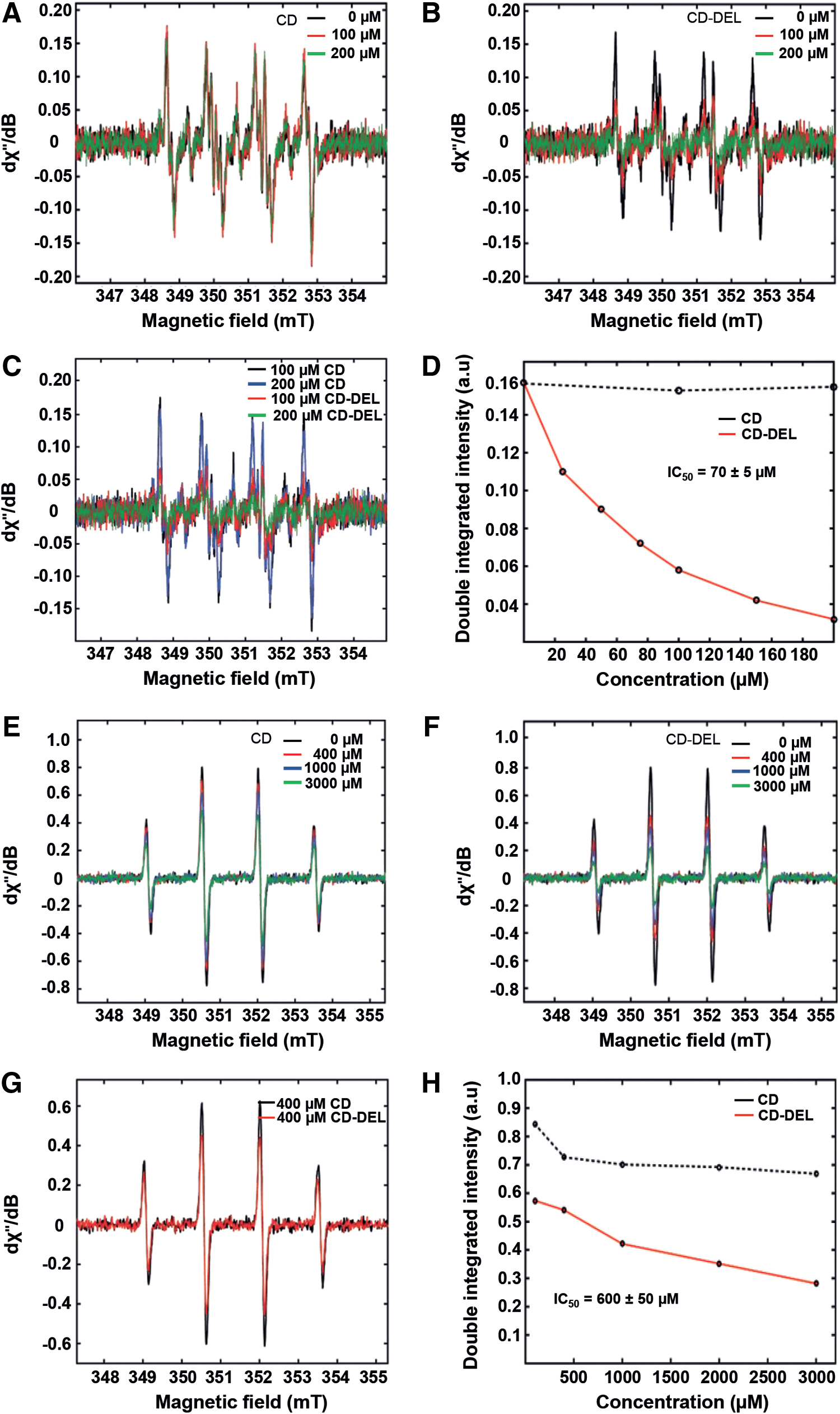
Since 4-HNE (conjugated with glutathione) also elicits the production of ROS, we next investigated the antioxidant activity of CD–DEL by electron paramagnetic resonance (EPR) spin trap. The ability of CD–DEL to compete with the spin trap 5,5-dimethyl-1-pyrroline N-oxide (DMPO) for superoxide and hydroxyl radicals was examined by determining the substrate concentration, which causes a 50% decrease in the EPR signal intensity of the corresponding spin adducts (DMPO-
Antioxidative effect of CD–DEL and upregulation of HO-1 in vitro and in vivo
To validate the antioxidant capacity in vitro, HEK-293 cells, known to increase ROS after activation by 4-HNE, were stained with oxidation-sensitive dye, dihydroethidium (DHE). The production of intracellular ROS after 4-HNE treatment was prevented by pretreatment with CD–DEL (IDV 4-HNE: 96.4 ± 18.6 and 4-HNE + CD–DEL 10 μM: 34.9 ± 0.3; p = 0.003), but not CD (Fig. 8A, B). In vivo, only CD–DEL reduced ROS formation in inflamed paw tissues by 26.3% (Fig. 8C, D). CD alone did not lead to a significant reduction in ROS levels. Since DEL has been shown to increase the expression of HO-1 in vitro (2), we investigated whether this signaling pathway was involved in the analgesic effect of CD–DEL. In vivo, CD–DEL but not CD treatment significantly increased the HO-1 (HMOX1) mRNA CD (IDV CFA + CD: 0.7 ± 0.05 and CFA + CD–DEL: 1.1 ± 0.03; p = 0.002; Fig. 8E) and protein levels of HO-1 (CFA + CD = 7.2 ± 0.5 and CFA + CD–DEL: 10.7 ± 0.4; p = 0.001; Fig. 8F). In vitro, hemin was used as positive control for the expression of HO-1, and cells were treated either with control siRNA or with HMOX1 siRNA. The latter prevented the increase of HO-1 protein expression compared with control siRNA (Fig. 8G). Similarly, CD–DEL significantly increased HO-1 expression in HEK-293 cells treated with control siRNA (IDV 1.2 ± 0.40) compared with control group treated only with CD (IDV 0.1 ± 0.03), but the densitometric analysis of the ratio of HO-1/glyceraldehyde 3-phosphate dehydrogenase (GAPDH) showed a 10-fold decrease in HO-1 protein expression after incubation of cells with CD–DEL and HMOX1 siRNA (IDV 0.1 ± 0.05, p = 0.013, Fig. 8G). No change was observed in naïve control cells treated with either control siRNA or CD.

Discussion
This study demonstrated that complexation of DEL with CD yields a potent local antinociceptive substance interfering with the proalgesic mediators ROS and 4-HNE in vitro and in vivo. This prevents TRPA1 activation leading to mechanical hyperalgesia and cold allodynia in preclinical models of inflammatory pain.
DEL is known for its antioxidant and anti-inflammatory properties, but its poor aqueous solubility and rate of dissolution are the two critical factors considered an obstacle for its therapeutic use. Kumoro et al. have suggested that DEL is soluble in water (28). Referring to European Pharmacopoeia 10, which states that “soluble” in a solvent refers to 1 g solute dissolved in 10–30 mL solvent,” the solubility observed by Kumoro et al. is still poor.
Corroborating data of this study, Goszcz et al. also found that DEL was unstable in tissue culture medium and the degradation process started immediately (20). Approximately 80% of DEL was lost in the first hour. To overcome such limitations, CDs have been employed as drug enhancers α-CD and β-CD, and a number of alkylated CDs are renally toxic and disruptive of biological membranes but γ-CD and some of its derivatives, as well as HP-β-CD and CD, are safer (67). Among the different CDs investigated, the highest solubility of DEL was found for CD. CDs are often used as drug enhancers for analgesics in clinical and preclinical studies to enhance the solubilization of drugs, reduce drug irritation at the site of administration, and stabilize unstable drugs in aqueous environment [reviewed in ref. (5)]. Among the different CDs investigated, the highest solubility of DEL was found for CD. The threefold increased solubility of the CD–DEL complex could be explained by the fact that the negatively charged sulfobutyl units interact electrostatically with the positively charged DEL and, of the alkyl groups; the butyl group possesses the optimal length for sterically permitting a corresponding interaction.
Local CD–DEL reversed selectively mechanical hyperalgesia, while systemic CD–DEL ameliorated mechanical and heat hyperalgesia. Systemic CD–DEL would not only scavenge ROS in peripheral tissue but also in the spinal cord or the central nervous system as part of the pain pathway. This process is subject to signal facilitation or inhibition in the spinal cord. In this context, spinal and supraspinal circuits facilitated by ROS could be more important for heat hyperalgesia (25, 35, 40)—suggesting a site-specific action of CD–DEL.
ROS are proalgesic via TRPA1; its function in pain has been demonstrated in many studies via TRPA1 knockout (KO) or antagonists in CFA-induced mechanical, as well as cold allodynia and 4-HNE-induced mechanical or nocifensive behavior (12, 49, 70). CD–DEL decreases 4-HNE expression leading to reduced (1) calcium influx in HEK-293 TRPA1, and (2) mechanical hyperalgesia and cold allodynia in vivo—presumably by preventing TRPA1 activation. The selective TRPA1 antagonist HC-030031 completely abolished 4-HNE-induced mechanical hyperalgesia and cold allodynia. Together, our results corroborate previous studies with TRPA1 antagonists inhibiting 4-HNE- induced hyperalgesia (70), and support a parallel mechanism of CD–DEL and TRPA1 antagonists. A direct link was supported by the in vitro experiments. 4-HNE-induced heat hyperalgesia seems to act through TRPA1-independent mechanism in line with previous studies in KO mice (12).
Besides its antioxidant activity, several intracellular targets for the anti-inflammatory effects of DEL have been reported. Anthocyanins and DEL inhibit NF-kappaB and JNK activation, and protein-tyrosine phosphatase 1B overexpression in a model of the adverse effects of a Western diet in mice, including obesity and type 2 diabetes (9). In our study, we observed antinociception within minutes after CD–DEL application. We, therefore, postulated that fast ROS scavenging would more likely explain these effects. However, it is important to notice that inflammatory mediators at day 1 after CFA injection are not necessarily the same at days 2, 3, or 4, although mechanical and/or heat hypersensitivity is always present. ROS-induced oxidative stress during inflammation results in 4-HNE protein adduct formation. These are also found in the shorter lasting Carrageenan inflammation models (48, 69). 4-HNE levels were high in inflamed paws and lowered by CD–DEL. In addition, CD–DEL reduced the number of infiltrating macrophages after only two injections. ROS can promote immune cell migration (11, 22). Taken together, these data indicate that ROS plays a role as a chemoattractant, so that the decreased CD68 IR cells after CD–DEL treatment would be due to its ROS scavenging activity. In response to chemoattractants, endothelial cells produce adhesion molecules causing flowing leukocytes to then tether, roll, adhere, and exit the blood vessel. The expression of the cell adhesion molecule VCAM-1 in the inflamed paws increased, and was downregulated by CD–DEL. Similar mechanisms have been observed by Huang et al., demonstrating that blueberry anthocyanins inhibit tumor necrosis factor alpha (TNF-α)-induced increases of VCAM-1 in endothelial cells in vitro (24).
Since CD–DEL decreases HNE-induced hyperalgesia, we first postulated an interaction of 4-HNE, as an aldehyde, with DEL, a polyphenolic compound. Possible reactions include nucleophilic addition, or radical substitution or additions. A nucleophilic addition is theoretically conceivable but not at physiological pH. We therefore hypothesized that DEL itself would be a radical and could then “radically” attack the Michael system of 4-HNE, whereupon 4-HNE itself would become a radical. However, UV–Vis absorption was unable to detect an interaction because the absorption spectrum of 4-HNE laid exactly within the wavelength range at ∼230 nm, where the absorption of CD–DEL was very strong. In addition, NMR analysis could not demonstrate a visible interaction between the two compounds, when 4-HNE was added to the complex at equal ratio as no shift of the spectrum was observed. We concluded that there is no reaction between both compounds and that the decreased 4-HNE level might be an indirect scavenging effect of ROS because DEL is known for its antioxidative properties. However, competitive experiments using NMR revealed that 4-HNE and DEL could displace each other in the cavity of the CD as an excess of 4-HNE repressed DEL from the cavity. These results indicate that the decrease of 4-HNE in vivo might also be due to HNE captured in the CD cavity. The displacement of DEL through 4-HNE from the cavity allows an oxidation (through light) of DEL, which results in a color change from purple to brownish (14). Further competitive assays are needed for validation. In summary, CD–DEL probably exerts a dual effect: ROS scavenging by DEL and HNE capturing by CD.
Electron spin (paramagnetic) resonance is a reliable method for the detection and quantification of free radicals. Due to the high reactivity and the associated low concentration of hydroxyl and hyperoxidanion radicals, one can use for their determination the so-called spin-trapping method, in which the short-lived radicals are converted by means of spin traps (spin traps) into stable spin adducts, which can be easily detected by EPR spectroscopy. CD–DEL was more powerful in scavenging superoxide than hydroxyl radicals as shown by other anthocyanins before (18, 45), as DEL has the highest activity against superoxide radicals in the group of anthocyanins (54). For DEL derivative present in eggplant (nasunin), the hydroxyl radicals scavenging activity were not due to direct scavenging, but rather to inhibition of their generating system by chelating ferrous ion. However, pelargonidin-3-glucoside, which has neither an orthodihydroxy substitution in the B ring nor a metal chelating capacity, exhibited similar or even higher activity than delphinidin-3-glucoside and cyanidin-3-glucoside, which possesses gallocatechol and catechol structures, and thus chelates iron (17, 18).
Given the 2% DEL content in CD–DEL, the results of this study are in good agreement with the reported value for uncomplexed DEL for superoxide radicals scavenging (IC50 = 1.6 ± 0.1 μM). CD–DEL seems more effective for the scavenging hydroxyl radicals than DEL alone (DEL: IC50 = 34.1 ± 2.5 μM) (71). This might be due to some scavenging activity of pure CD for hydroxyl radicals in EPR. Hydroxyl and superoxide radicals scavenging mediates the reported antioxidant effect with reduced ROS and 4-HNE and antinociceptive activities of CD–DEL. Similar to the approach used in this study, curcumin, a polyphenol with antioxidant and anti-inflammatory properties, has been complexed with β-CD. This improved the antioxidant activity in vitro due to the considerable increase in its solubility [reviewed in ref. (59)]. Furthermore, clinical trials with repeated administration of CDs supported a wide therapeutic window of up to 144 mg/kg every 12 h (8, 53). A previous study demonstrated that doses of CD up to 1500 mg/kg produced no histopathological evidence of toxicity in dog kidneys and no deleterious effects on renal function of healthy volunteers (32). In agreement with that study, the European Medicines Agency revealed in the background review for CDs used as excipients that low doses of CD are not expected to cause adverse effects (European Medicines Agency [2014] Background review for CDs used as excipient. EMA/CHMP/333892/2013). Regarding DEL, the European Food Safety Authority analyzed Wistar rats fed with the anthocyanins extract (cyanidin, petunidin, and DEL) at levels equivalent to 3000 or 6000 mg/day for a period of 90 days. No differences were observed between the test animals and controls in survival, growth, or histopathology of the principal tissues at the termination of the study (Scientific Opinion on the re-evaluation of anthocyanins [E 163] as a food additive, DOI: 10.2903/j.efsa.2013). Moreover, DEL (208-237-0) is approved as food additive, and belongs to anthocyanins used as pigments as food colorants and additives E163.
HO-1 was increased in inflamed paw after treatment with CD–DEL as well as in cells after stimulation with 4-HNE. Anthocyanins and other phenolics from bilberry upregulate the oxidative stress defense enzymes such as HO-1 (41). HO-1 levels are increased in inflammation (56), while HO-1 overexpression or induction is associated with anti-inflammatory pathways (3, 51, 58). Upregulation of HO-1 reduces irritant-induced pain (46), or inflammatory pain by interleukin-1β or CFA (26, 66). In accordance with these results, HO-1 overexpression in endothelial cells inhibits TNF-α-mediated VCAM-1, but not intercellular adhesion molecule 1 expression (64). Thus, in addition to ROS scavenging, CD–DEL seems to elicit its analgesic effect via upregulation of HO-1. Pharmacologic or gene therapy modulation of HO-1 has beneficial immunoregulatory and cytoprotective properties (35, 63). To address the significance of upregulation of HO-1 after CD–DEL treatment, cells were treated with siRNA of HMOX1 to specifically inhibit HMOX1 expression. The expression of HO-1 at the protein level was clearly downregulated after treatment of cells with HMOX1 siRNA. Induction of HO-1 by CD–DEL could therefore be a good strategy to increase individual's self-antioxidant capacity and decrease inflammation.
In summary, CD–DEL is antinociceptive by decreasing ROS and 4-HNE formation as well as infiltration of macrophages and edema formation. CD–DEL exhibits its antinociceptive effects through dual antioxidant and anti-inflammatory activities. Future in vivo experiments are needed to confirm these observations. In this study, CD or CD–DEL demonstrated no effect on the motor activity of rats with CFA-inflammation. Since DEL and CD alone are also nontoxic in humans, the next step would be prolonging the short-lasting antinociceptive effects, for example, using nanocarriers for local application.
Materials and Methods
Electronic laboratory notebook was not used. All animal experiments were performed after the IRB approval by the government of Lower Franconia. Protocol number 108/13.
Drug formulation
Preparation of complexes with different CDs
Complexation of DEL with various CDs and the solubility of the complexes in aqueous solutions were investigated (Patent number 9925274).
Aqueous solutions containing 10% (w/w) of the CDs were prepared, and 5 mL of each solution was added to an excess amount of solid delphinidin chloride (DEL). This excess amount was found with 10 mg DEL in case of α-, β-, and γ-CD, and 15 mg was necessary for HP-β-CD and CD. In the case of β-CD, a concentration of only 2% CD was chosen because of its poor solubility. The suspensions were homogenized by stirring for 20 h at 30°C in the dark to avoid light-induced oxidation and then filtered through a membrane filter of 0.22 μm pore size. The results of solubility test indicated that the strongest interaction was found between CD and DEL, but DEL shows higher chemical stability under acidic conditions. Therefore, the preparation of CD–DEL complex was repeated in media set at different pH values (pH = 6.0, 4.8, and 4.1) in the presence of 10% CD, and the stirring time was reduced to 2.5 h to further reduce the risk of chemical decomposition. The following CDs (obtained from Cyclolab Ltd.) were used: α-CD (ID No: CYL-2322); β-CD (ID No: CYL-3190); γ-CD (ID No: CYL-2323); HP-β-CD (ID No: L-043/07); SEB-β-CD (CD) (ID No: 47K010111). DEL was obtained from Extrasynthese (Batch 49, LOT 11071849).
Stability of DEL in CD complex in aqueous solution
To determine the DEL content for stability in aqueous solutions, CD–DEL complex and DEL were dissolved in 0.9% NaCl solution until starting concentrations of 1.584 mg/mL for CD–DEL and 0.0216 mg/mL for DEL (comparative example) had been established. The solutions were prepared at room temperature (RT), and then stored at 37°C in the dark in closed vials. DEL content was determined after 1, 2, 3, and 4 h.
Quantitative analysis
A reversed-phase high-performance liquid chromatography (HPLC) was used for quantification of DEL in aqueous solution. The following reagents were used: purified water; methanol for chromatography; formic acid, p.a.; 1 M hydrochloric acid as volumetric solution. The column used was a XBridge™ C18, 3.5 μm, 150 mm × 4.6 mm (obtained from company Waters). The mobile phase A consisted of 770 mL water, 230 mL methanol, and 10 mL formic acid, and the mobile phase B (MP-B) consisted of 50 mL water, 950 mL methanol, and 10 mL formic acid. For the HPLC separation, the flow rate was set to 1.0 mL/min, and the gradient was performed as follows: 0% MP-B from 0 to 5 min after injection; 0%–20% MP-B from 5 to 20 min; 20%–100% MP-B from 20 to 25 min; back to 0% MP-B after 25 min. The column was equilibrated for 10 min before next injection. The column heater was set to 30°C ± 2°C, and the injection volume was 20 μL. The UV–Vis detector was set to 530 nm for the assay and to 275 nm for the detection of impurities.
Calibration solution was prepared as follows:
Solutions and sample preparation: Dilution solution 1 consisted of 100 mL methanol and 2.6 mL 1 M HCl; dilution solution 2 consisted of 100 mL 50% methanol and 2.6 mL 1 M HCl. A reference solution of DEL was prepared by weighing 10 mg of delphinidin chloride into a 10 mL volumetric flask and dissolving it in dilution solution 1. After the dissolution, the solution was diluted ∼10-fold with dilution solution 2 to produce an approximate concentration of 0.1 mg/mL. The concentration of DEL in the samples was calculated by Agilent ChemStation software using the external standard calibration method based on the peak area. The calculated content was reported as the percentage of the starting concentration.
Animal models
All animal care and experimental protocols complied with international guidelines for the care and use of laboratory animals (EU Directive 2010/63/EU for animal experiments) and were approved by the Government of Unterfranken (protocol number 108/13). Approved score sheets criteria were defined and animals treated accordingly. Animal studies are reported in compliance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines (27, 39). Male Wistar rats weighing 220–250 g (Janvier Labs, Le Gene st-Saint-Isle, France) were accommodated on dry litter in groups of six under standard conditions (12 h:12 h light/dark cycle, 21°C–25°C, 45%–55% humidity) with food and water ad libitum. All experiments were carried out during the light phase of the cycle. Equal sizes (N = 6) per group were planned. Experimental groups with CFA included six animals/group based on an a priori power analysis (G Power 3) (effect size 2, p < 0.05, t-test) (16). Animals were treated under brief isoflurane anesthesia and were sacrificed at the end of the experiment. Inflammation was induced by unilateral
Nociceptive thresholds were assessed in various groups of rats: Four days after CFA, CD or CD–DEL (Sapiotec GmbH, Würzburg, Germany) was administered
Measurement of nociceptive thresholds and motor function
Determination of heat and mechanical nociceptive thresholds is the standard nociceptive reflexive test in pain research. Mechanical thresholds were measured by paw pressure threshold, a modified Randall Selitto test (Ugo Basile, Gemonio, Italy) as described in previous study (62). Animals were gently restrained under paper wadding. Using a wedge-shaped, blunt piston, incremental pressure was applied on the dorsal surface of the hind paw by means of an automated gauge. The pressure required to elicit paw withdrawal, the PPT (250 g cutoff), was determined by averaging three consecutive trials.
Heat nociceptive thresholds were determined by the Hargreaves test (62). The latency (time; s) required to elicit paw withdrawal was measured with an electronic timer (IITC, Inc./Life Science, Woddland Hills, CA) after application of radiant heat to the plantar surface of a hind paw from underneath the glass floor with a high-intensity light bulb. A decreased threshold was interpreted as hyperalgesia/pain and an increased threshold antinociception/analgesia. Acetone test was performed to assess cold allodynia after 4-HNE or CFA before and after treatment with CD–DEL (5 mg/100 μL) or TRPA1 antagonist HC-030031 [30 mg/kg, i.p. (49) CAS 349085-38-7]. The paw was exposed to a cold stimulus, a drop (100 μL) of acetone, and a four-point scale based on the following graded response: 0 = no response; 1 = quick withdrawal, stamp, or flick of the paw; 2 = prolonged withdrawal or repeated flicking; and 3 = repeated paw flicking with paw licking. The three trial scores were summed to give the final score (from 0 to 9) (65).
An accelerating Rota-Rod (Ugo Basile) (61) was used to determine if local administration of CD–DEL or CD affects the motor function of rats. Rats were placed on a Rota-Rod treadmill and tested after habituation during three training sessions before the data collection. The apparatus was rotating at a gradually increasing speed from 5 to 50 rpm over a cutoff time of 1000 s. Baseline data were measured followed by the application of the CD–DEL or CD. The maximum performance time of five consecutive measurements was analyzed per session, at an intertrial interval of 5 min. Tests were performed before and 60 min after injection.
Measurement of DEL concentration in plasma and paw tissue after local and systemic administration of CD–DEL
Plasma preparation
Rats were coinjected with CFA and CD–DEL at day 0. Blood samples were collected at 1 day using an ethylenediaminetetraaceticacid (EDTA) syringe at 5, 30, 60 min after systemic (i.p.) administration of CD–DEL. After withdrawal of each blood sample, 1 mL of EDTA plasma was immediately stabilized with 0.2 mL of HCl (1 M) and kept on ice. For protein precipitation, 100 μL of this solution was mixed with 500 μL of a solution of methanol:HCl 1 M (90:10), vortexed and centrifuged for 5 min at 15,000 rpm. The supernatant was used for liquid chromatography mass spectrometry (LCMS) analysis.
Paw tissue preparation
Rats were systemically or locally coinjected with CFA and CD–DEL. At 1 day blood (in EDTA tubes) and inflamed paw tissue were collected after 5, 30, 60 min after new application of CD or CD–DEL. One milliliter of a solution of methanol: Triton X-100:HCL 1 M (80:20:1) was added to 1 mL blood and 300–700 mg tissue, homogenized for 5 min in tissue lyser, and kept on ice. After centrifugation for 10 min at 15,000 rpm, 100 μL of supernatant was mixed with 100 μL of a solution of methanol:HCl 1 M (9:1), vortexed, centrifuged for 5 min at 15,000 rpm, and analyzed by LCMS.
Liquid chromatography tandem mass spectrometry
Chromatographic separation was carried out on a Kinetex C18 100 Å (100 mm × 2.1 mm, 2.6 μm) column (Phenomenex) at 40°C with a mobile phase flow rate of 0.40 mL/min. Mobile phases of 0.1% formic acid in water (A) and 0.1% formic acid in methanol (B) were used. Chromatographic separation was performed using a gradient as follows: 0–0.25 min, 10% B; 0.25–1.25 min, 10%–90% B; 1.25–2.20 min, 90% B; 2.20–2.21 min, 90%–10% B; 2.21–3.50 min, 10% B as equilibration time. Injection volume was 4 μL, and the samples were kept at −20°C throughout the analysis.
The mass spectrometric analysis was performed using a Shimadzu 8030+ triple-quadrupole mass spectrometer coupled with an electrospray ionization source in the positive ion mode. Multireaction monitoring mode was applied for quantification using target fragment ions m/z 303.00 → 228.95 for DEL: for calibration, DEL was serially diluted in methanol: HCl 1 M (9:1). Blank-plasma and blank-tissue-extract were spiked with these solutions in a concentration range 0.02 − 10 μg/mL.
Cellular models
HEK-293 cells were either used without transformation or prepared to stably express either recombinant human TRPA1 or rat TRPV1 (34, 40, 46). Human TRPA1 or rat TRPV1 cDNA were cloned to the lentiviral expression vector pCDH1-UBC-MCS-EF1-hygromycin clone 1 (SBI, Mountain View, CA). Lentiviral particles were produced in HEK-293 cells with the pseudotyping vector pMD2.G and the packaging vector pCMVΔR8.91. TRPA1 and TRPV1 positive clones were selected by hygromycin (0.4 mg/mL). The stable cell line was continuously kept under hygromycin selection.
siRNA transfection
HEK-293 cells were seeded in antibiotic-free medium 1 day before the transfection. To knockdown HMOX1 by siRNA, the cells were transfected either with the human HMOX1-specific siRNA or with control siRNA for 48 h using HiPerFect transfection reagent according to the manufacturer's instructions (Qiagen; #301704). After 48 h, the medium was changed, and the transfected HEK-293 cells were treated with CD and CD–DEL (10 μM) for 16 h. Hemin (5 μM, 51280; Sigma) was used as positive control together with a solvent control (0.5% dimethyl sulfoxide [DMSO]).
Western blot
Protein (25 μg) from subcutaneous paw tissue leaving the deep flexor tendon in place was extracted after indicated treatments, separated by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto nitrocellulose membranes as described before (21). HEK-293 cells were treated with CD and CD–DEL for 16 h (2). After the treatment, the medium was discarded, and cells were rinsed with cold phosphate-buffered saline (PBS). Radioimmunoprecipitation assay buffer, which contained freshly added protease and phosphatase inhibitors, was added, and the cells were collected by scraping. Protein isolation was done by physical cell damage (pipetting 10 times through a 1 mL syringe) and centrifuging at 14,000 rpm, 4°C for 30 min. After the centrifugation, the supernatant was decanted, and the protein concentration was measured. Fifty micrograms of protein was separated by 12.5% (w/v) SDS-PAGE, and then was electroblotted to polyvinylidenefluoride membrane.
Membranes from both procedures were blocked with 2.5% nonfat milk and 2.5% bovine serum albumin in 1 × Tris-buffered saline with 0.1% Tween 20 detergent, and then probed with a specific primary antibody at 4°C overnight: polyclonal rabbit anti-4-HNE (1:500, #HNE11-S; Biotrend, Cologne, Germany), mouse monoclonal anti-HO-1 (1:1000, ab68477; Abcam, Berlin, Germany), goat polyclonal anti-VCAM-1 (1:600, sc-1504; Santa Cruz Biothechnology, Dallas, TX), and GAPDH (1:3000 in 5% non-fat milk powder, #3700; Cell Signaling Technology, Frankfurt, Germany) overnight at 4°C. Membranes were further incubated with the corresponding secondary antibodies conjugated to horseradish peroxidase. Protein bands were visualized using enhanced chemiluminescence solution and FluorChemFC2 Multi Image II (Cell BioSciences, Heidelberg, Australia), and the intensity of the bands was quantified using ImageJ software (NIH).
Immunohistochemistry
Subcutaneous tissue with adjacent skin dissected from hind paws was collected, postfixed in the fixative solution, cryoprotected in 10% sucrose solution at 4°C overnight, embedded in tissue-Tek compound (OCT; Miles, Inc., Elkhart, IN), and frozen (22, 60). Sections of 7 μm were fixed with −20°C acetone, permeabilized, blocked, and incubated with the following primary antibodies (1:100) overnight: polyclonal rabbit anti-4-HNE (HNE11-S) in combination with monoclonal mouse anti-ED1 (CD68, MCA341R; Biorad, Puchheim, Germany). This was followed by a sequential incubation with secondary antibodies and 4′,6-diamidino-2-phenylindole. Confocal laser-scanning microscopy was performed (Zeiss LSM 510 META, Jena, Germany), and ImageJ was used for manual counting of positive cells. The number of positive or double positive cells per whole paw section was counted. Then the mean per slide (three sections/slide) was calculated. Three slides were analyzed per animal. The specificity of staining was indicated by the following controls: omission of either the primary antibodies or the secondary antibodies.
Calcium imaging
For ratiometric single cell calcium analysis, either HEK-293 cells were labeled with 6 μM Fura-2/AM for 30 min at 37°C in imaging solution (in mM): 134 NaCl, 6 KCl, 1 MgCl2, 1 CaCl2, 10 HEPES, 5.5 glucose, pH 7.4 adjusted with NaOH. In untransduced cells, no specific changes in intracellular calcium could be measured (37, 43, 49). All measurements were performed at RT using a Nikon TE2000-E microscope. Fura-2/AM is excited for 100 ms with 340/380 nm with a Lambda DG4/17 wavelength switch (Sutter Instruments, Novato, CA). Time-lapse image series were acquired at intervals of 2 s with a cooled EMCCD Andor iXon camera (Andor Technology Ltd., Belfast, United Kingdom) controlled by NIS Elements Software (Nikon, Amsterdam, Netherlands). Objective: CFI S-Fluor 10 × /0.5 (Nikon). Image series were analyzed with ImageJ 1.46r, time series analyzer V2.0 plugin (Rasband, W.S.; ImageJ). The following substances and controls were used: 4-HNE, capsaicin, CD–DEL, and CD used as vehicle. In all statistical analysis, mean of basal fluorescence intensities was determined for each measurement. The number of reacting cells (%) was calculated by 1.5-fold increase of mean basal fluorescent intensity after stimulation. The intracellular calcium concentration of individual cells was recorded over time, and the average trace and the area under curve were calculated from the average. Intervals corresponded to the stimulation period of each substance. Mean of basal fluorescence was set as zero.
UV–vis-absorption spectroscopy and NMR
Absorption spectra of CD–DEL solutions in PBS were recorded in the spectral range from 400 to 800 nm using a setup similar to (36). Stock solutions of CD–DEL at different concentrations CD–DEL (5 mM, 400 μM, 15 μM) were prepared, and subsequent recordings of absorption spectra from a freshly prepared solution were performed for 16 min to monitor the decrease of DEL concentration over time. Separately, 4-HNE (7.5 μM) was added to the freshly prepared 400 μM DEL solutions in PBS and subsequent measurements of absorption spectra allowed for comparison with the DEL depletion kinetics. All absorption measurements were performed using a Cary-5000 UV–vis-NIR spectrometer (Agilent, Santa Clara, CA) in a 1 cm cuvette. Background was corrected by absorption spectra of pure PBS solution.
NMR measurements were performed using a Bruker AV 400 spectrometer (1H 400.131 MHz; Bruker BioSpin, Rheinstetten, Germany) and TopSpin 3.5 software. The signals of the deuterated solvents were used for internal standard (DMSO-d6: 1H 2.50 ppm, number of scans: 16). First, the solvent of the 4-HNE solution (n-hexane) was evaporated under reduced pressure. Eighteen milligrams (0.059 mmol) DEL, 6 mg (0.038 mmol) 4-HNE and 2,2,6,6-tetramethyl-I-piperidinyloxy free radical (TEMPO) were dissolved in DMSO/D2O (ratio 1:5) and incubated (37°C, 400 rpm) under exclusion of light. After 10, 30, and 60 min, NMR measurements were carried out in brown glass NMR tubes to prevent light contact.
For the competitive 1H NMR measurements, identical parameters were used as described above. The spectra were referenced to the external standard of 0.05% sodium trimethylsilylpropionate in D2O (TSP-d4) filled in a coaxial insert tube. For the measurements, three different samples (CD:DEL:4-HNE molar ratio 1:1:1, 1:2:1, 1:1:2) were prepared. As example (molar ratio 1:1:1): 22 mg (0.01 mmol) CD, 3 mg (0.01 mmol) DEL, and 1.6 mg (0.01 mmol) 4-HNE were dissolved in DMSO/D2O (ratio 1:5) and incubated (37°C, 400 rpm) under exclusion of light. After 10 min, NMR measurements were carried out.
EPR to measure superoxide and hydroxyl radicals scavenging activity
For EPR, X-band (9.86 GHz) EPR spectra were recorded at RT on a Bruker ELEXSYS E580 CW/FT EPR spectrometer (Bruker, Rheinstetten, Germany) fitted with an AquaX cell (19-bore version). The following EPR instrumental settings were used to acquire each spectrum: a microwave power of 10 mW, modulation amplitude of 1 Gauss, modulation frequency of 100 kHz, and a conversion time of 5 ms. Values for the double integrated intensity (A) are averages of at least two separate experiments.
The hypoxanthine/xanthine oxidase system generated superoxide radicals (62). For the spin trapping of superoxide radicals with DMPO, the experimental protocol below was followed. PBS buffer (100 mM, pH 7.4) containing diethylenetriaminepentaacetic acid (DTPA, 2 mM) was used to dissolve all reagents. The test compound CD or CD–DEL stock solution (20 mM) was prepared in the above prepared buffer. Solutions of hypoxanthine (1 mM, 400 μL), DMPO (1 M, 80 μL), and the test compound CD or CD–DEL (at various concentrations, 25, 50, 75,100, 200, and 400 μM) were mixed with prepared buffer. Superoxide formation was initiated by the addition of xanthine oxidase (stock 20 U/mL, 4 μL). Reaction mixtures (final volume completed to 764 μL with PBS/DTPA) were transferred to the AquaX cell and the EPR spectra recorded after 90 s.
To generate
The concentrations of the CD–DEL causing a 50% decrease in relative intensity of the DMPO radicals adducts (IC50) were measured by linear interpolation based on experiments in which the CD–DEL was tested at six different concentrations in duplicate.
Microscopic detection of intracellular ROS formation
The cell permeable fluorescent dye DHE was utilized to detect intracellular ROS (2). One day before the experiment, HEK-293 were seeded on cover slips in six-well plates containing 3 mL medium per well. On the day of the experiment, cells were preincubated either with CD–DEL or with CD for 15 min before adding the stress inducer 4-HNE for an additional 30 min. DHE (10 μM) was added for last 20 min during the 30 min treatment with 4-HNE (after pretreatment with CD–DEL or CD at 37°C in the dark). At the end of the incubation time, cells were washed once with PBS, and 5–6 images were taken per treatment group.
Similarly, cryosections from embedded paw tissues (5 μm) were prepared with a Leica CM3050 Scryostat (Leica, Wetzlar, Germany). Sections were stained with 10 μM DHE dye for 30 min in the dark at RT. After staining, sections were washed twice with PBS, mounted and covered with a cover glass. Images of the cells were taken with an Eclipse 55i microscope (Nikon GmbH, Düsseldorf, Germany) and a Fluoro Pro MP 5000 camera (Intas Science Imaging Instruments GmbH, Göttingen, Germany) at 200-fold magnification. Quantification was done by measuring gray values of 200 different locations from 5 to 6 images per animal with ImageJ.
Reverse transcription quantitative polymerase chain reaction
Total RNA from rats' hind paws was extracted using Trizol. Total RNA (1 μg) was transcribed to cDNA using the high-capacity cDNA kit (Applied Biosystems, Thermo Fischer, Waltham, MA). Quantitative polymerase chain reaction (qPCR) analysis was performed with the following primers: HO-1 (Thermo Fisher; cat# Rn00561387_m1). qPCR analysis was carried out using the StepOnePlus Real-Time PCR System (Applied Biosystems) with the following program: fast 40 min for a run, 95°C for 2 min, and 45 cycles at 95°C for 1 s and 60°C for 20 s following TaqMan® fast advanced master mix user guide (Pub. No. 4444605, Rev. D). Samples from animals treated with CD and CD–DEL (six/group) were analyzed in triplicates. Target cDNAs were normalized to endogenous control (GAPDH). Relative quantification of mRNA was calculated using the ΔΔCt method, in which Ct represents the threshold cycle value.
Statistical analysis
One- or two-way repeated measurements analysis of variance (ANOVA) followed by the Student–Newman–Keuls post hoc test was used to analyze the one or more time points and treatment groups when data were normally distributed. The t-test was used to compare two groups. For not normally distributed data ANOVA on ranks was applied. Experiments were performed in triplicates with six animals per group (in vivo) and three independent experiments (in vitro). Differences between groups were considered significant if p < 0.05.
Footnotes
Author Disclosure Statement
N.R. and J.A.B. are owners of Sapiotec GmbH, Würzburg, Germany. No conflict of interest due to other financial support is involved.
Funding Information
This study was supported by internal funds from the University Hospital of Würzburg (R.-S.S., H.L.R., A.B., J.A.B., N.R.), the Chinese Scholarship Council (S.Y.), the Evangelisches Stipendienwerk Villigst, the Interdisciplinary Center for Clinical Research Würzburg (IZKF; N-261 B.O., H.L.R. and Z2/CSP-2 to B.O.), German Ministry for Education and Research (BMBF: N.R., J.A.B.), and the Julius-Maximilians-Universität Würzburg (H.B., H.S., U.H.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.