
Abstract
Aims:
In hemolysis, which is accompanied by increased levels of labile redox-active heme and is often associated with hemostatic abnormalities, a decreased activity of activated protein C (APC) is routinely detected. APC is a versatile enzyme that exerts its anticoagulant function through inactivation of clotting factors Va and VIIIa. APC has not been demonstrated to be affected by heme as described for other clotting factors and, thus, is a subject of investigation.
Results:
We report the interaction of heme with APC and its impact on the protein function by employing spectroscopic and physiologically relevant methods. Binding of heme to APC results in inhibition of its amidolytic and anticoagulant activity, increase of the peroxidase-like activity of heme, and protection of human umbilical vein endothelial cells from heme-induced hyperpermeability. To define the sites that are responsible for heme binding, we mapped the surface of APC for potential heme-binding motifs. T285GWGYHSSR293 and W387IHGHIRDK395, both located on the basic exosite, turned out as potential heme-binding sites. Molecular docking employing a homology model of full-length APC indicated Tyr289 and His391 as the Fe(III)-coordinating amino acids.
Innovation:
The results strongly suggest that hemolysis-derived heme may directly influence the protein C pathway through binding to APC, conceivably explaining the decreased activity of APC under hemolytic conditions. Further, these results extend our understanding of heme as a multifaceted effector molecule within coagulation and may allow for an improved understanding of disease development in hemostasis under hemolytic conditions.
Conclusion:
Our study identifies APC as a heme-binding protein and provides insights into the functional consequences.
Introduction
The redox active molecule heme is known for its essential role as the prosthetic group in a number of hemoproteins, such as hemoglobin and cytochromes. Each of up to 25 trillion circulating red blood cells (RBCs) in the human body is considered to contain 2.5 × 108 hemoglobin molecules with 5 × 109 molecules of heme bound, which roughly corresponds to 80 mM heme (9, 10). Under hemolytic conditions, heme is released as a result of erythrocyte lysis and subsequent degradation of hemoglobin, which, in turn, may lead to repletion of the heme scavenger hemopexin (57). Heme may also adhere to other plasma proteins, such as albumin and α1-microglobulin, resulting in a pool of so-called “labile heme” (63).
Labile heme is well known as a regulator of various proteins in a broad range of biochemical and pathophysiological processes, such as transcription, signal transduction, inflammation, or hemostasis, but it is still unresolved as to what extent it might influence coagulation processes, for example, in hemolytic states (19, 37, 57).
Intravascular hemolysis has been reported to increase the thrombotic risk (1, 44). This can be partially explained by hemolysis-induced coagulation activation through exposure of negatively charged phospholipids from ruptured RBCs, induction of tissue factor expression, and stimulation of von Willebrand factor release from endothelial cells (37, 57). However, the underlying mechanisms of these events have not yet been elucidated (1, 16, 54).
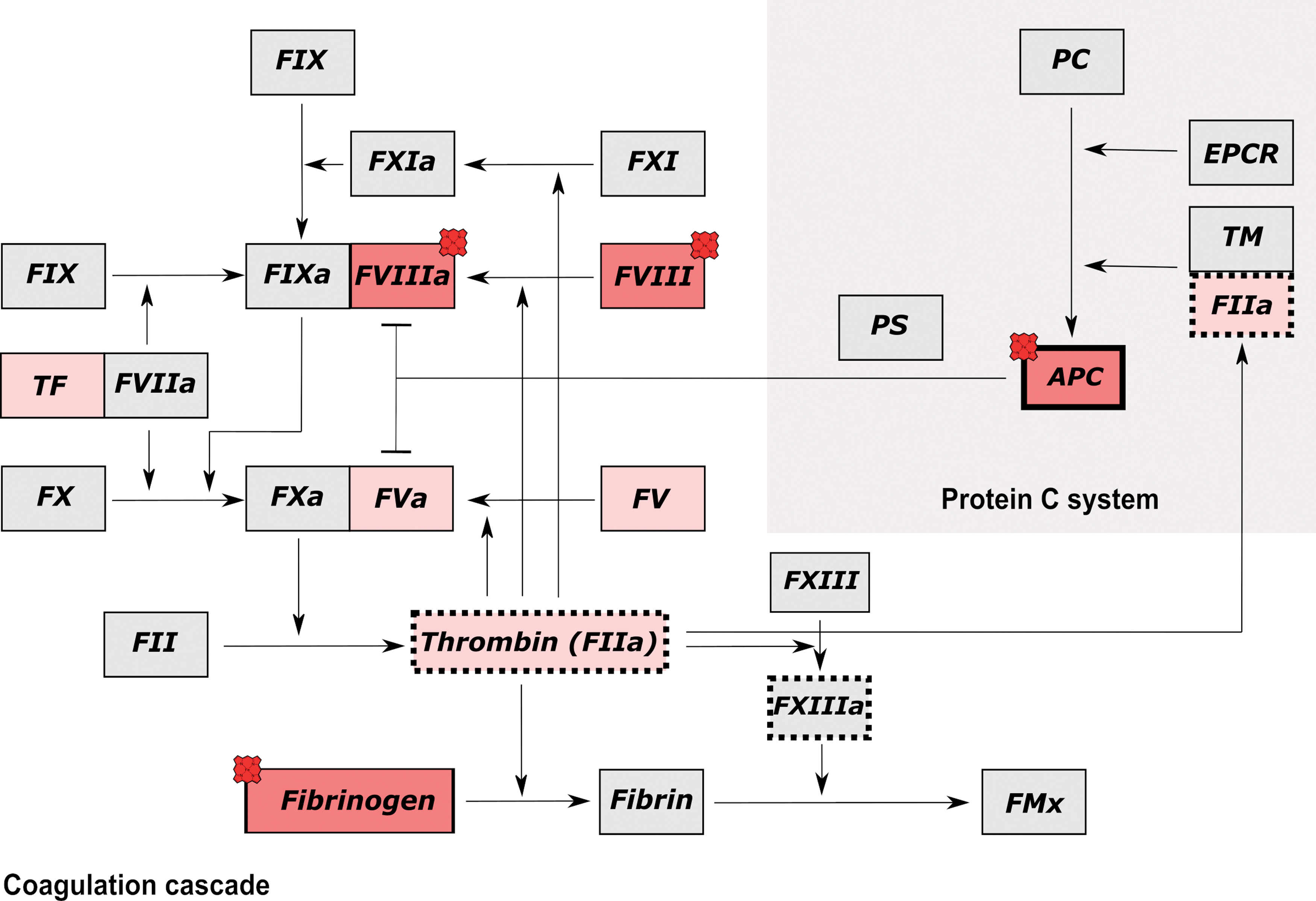
Interference of heme with endogenous inhibiting systems such as the protein C (PC) pathway, might contribute to the hemolysis-induced hypercoagulable state. PC is the zymogen of the serine protease activated PC (APC) that is synthesized in a vitamin K-dependent manner in the liver and circulates at a concentration of 70 nM in human blood (23). PC consists of a heavy and a light chain connected by a disulfide bond (32). The light chain is composed of a Gla domain and two epidermal growth factor (EGF)-like domains, whereas the heavy chain comprises the serine protease domain. Depending on the glycosylation pattern of the heavy chain, different molecular forms (α, β, and γ) of PC exist (20).
PC becomes activated to APC through limited proteolysis and cleavage of the activation peptide by the thrombin-thrombomodulin complex on the luminal surface of endothelial cells. Once formed, APC downregulates further thrombin formation by proteolytic inactivation of the procoagulant cofactors activated factor VIII (FVIIIa) and activated factor V (FVa) (56). In addition, APC can exert pleiotropic cytoprotective effects, including the activation of antiapoptotic and anti inflammatory pathways enhancing, for example, endothelial barrier integrity. These effects are induced by APC-mediated activation of the G-protein coupled protease-activated receptors 1/3 (PAR1/3) and involve binding of APC to the endothelial protein C receptor (EPCR) (21).
Innovation
Several coagulation factors have been earlier described to be affected by heme, which, together with the occurrence of coagulopathies during hemolytic events, inevitably suggests a link between hemolysis and coagulation. For the first time, we demonstrate transient heme binding to activated protein C (APC), an anticoagulant serine protease, and identify two potential binding sites. Further, our findings on the functional consequences for APC on heme binding led us to assume that a heme overload under hemolytic conditions might directly affect the protein C pathway and, consequently, exerts an effect on clotting processes.
The increased thrombotic risk of inherited PC deficiency, inherited protein S (PS) deficiency, and APC resistance emphasize the importance of this anticoagulant pathway (3, 43, 66). In addition, Gabre et al. reported that APC can induce the expression of heme oxygenase 1 (HO-1), an enzyme that catalyzes the degradation of heme (17). Earlier, however, an upregulation of PC and thrombomodulin by HO-1 was shown by Fei et al. (13). Continuing the line, in patients with sickle cell disease (SCD) accompanied by severe hemolysis, decreased levels of APC were partially reported based on the amidolytic activity of the enzyme, however, without describing the underlying mechanism (60, 67).
In previous studies, we identified potential heme-regulated proteins such as interleukin-36α and cystathionine-β-synthase, which were unknown to bind heme (6, 39, 49, 61, 70). Herein, we provide evidence that human APC is a heme-binding protein and thus might represent a heme-regulated protein, as well.
Surface plasmon resonance (SPR) spectroscopy and in silico studies were used to characterize the respective interaction. Potential heme-binding sites on the surface of APC were identified and analyzed by ultraviolet/visible (UV/vis) spectroscopy. To visualize heme binding to APC, we employed molecular modeling and docking studies by using an in-house generated homology model. Moreover, we demonstrated an impact of heme on the amidolytic and anticoagulant function of APC and observed an increased peroxidase-like activity of the heme-APC complex. Apart from that, the cytoprotective functions of APC were not influenced by heme binding as human umbilical vein endothelial cells (HUVECs) were protected from heme-induced cytotoxicity in the presence of APC.
Taken together, our data provide novel valuable insights into the regulation of the anticoagulant protein APC by heme, which may contribute to our understanding of the physiological functions of APC as well as the development of hemolytic diseases associated with hemostatic abnormalities.
Results
Heme binds to human APC
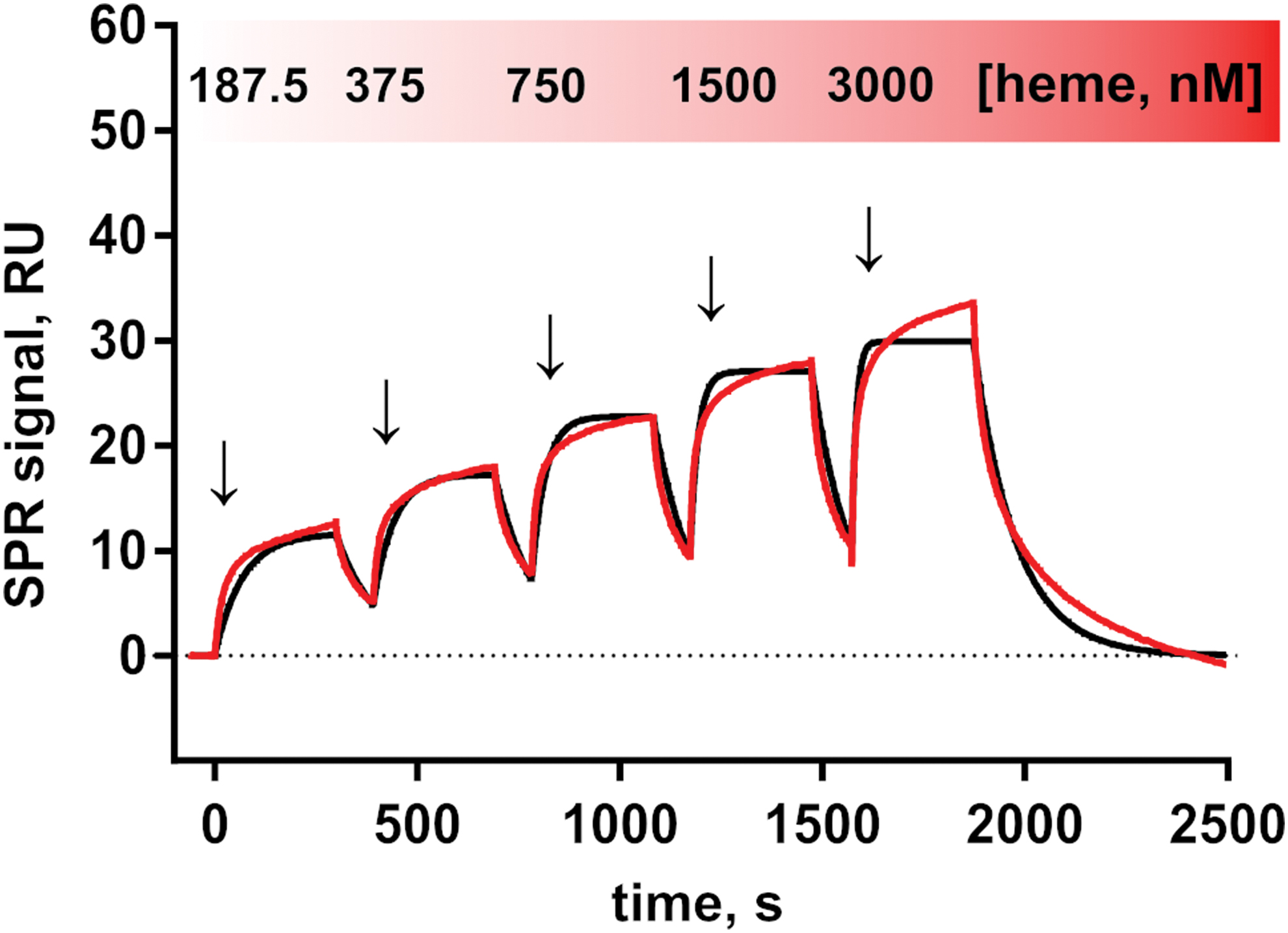
To demonstrate and characterize the binding of heme to APC, SPR measurements of the heme-APC interaction were performed. The analysis of the association of heme with APC by using a global 1:1 kinetic model revealed a equilibrium dissociation constant (K D) value in the nanomolar range (400 ± 47 nM) according to the best fit (Fig. 1). The binding of heme to APC is transient, which is characterized by a rapid association (association rate constant [k a] = 2.06 × 104 m−1·s−1) and dissociation process (dissociation rate constant [k d] = 8.08 × 10−3·s−1). A stoichiometry of ∼1:1.3 (protein:heme) was determined, which indicates either overlaying binding events of heme to different binding sites, nonspecific binding, or both.
Heme binds to APC-derived peptides
Previously, it was shown that nonapeptides represent suitable models for analyzing heme-regulatory/binding motifs (HRMs/HBMs) on the surface of proteins (39, 40, 49). Four APC-derived nonapeptides were included in this study (peptides
Heme binding was observed by a Soret band shift to ∼420 nm for all peptides (Fig. 2 and Supplementary Fig. S1). It was previously demonstrated by different spectroscopic methods, including resonance Raman spectroscopy, continuous-wave electron spin resonance spectroscopy, and two-dimensional nuclear magnetic resonance spectroscopy, that a shift to ∼420 nm is typical for a mixture of a hexa- and pentacoordination or hexacoordination of a heme-peptide/protein complex (5, 7, 39, 61). According to the best fit, a stoichiometry of 1:1 was determined for all heme-peptide complexes investigated herein. The highest heme-binding affinities were observed for peptides
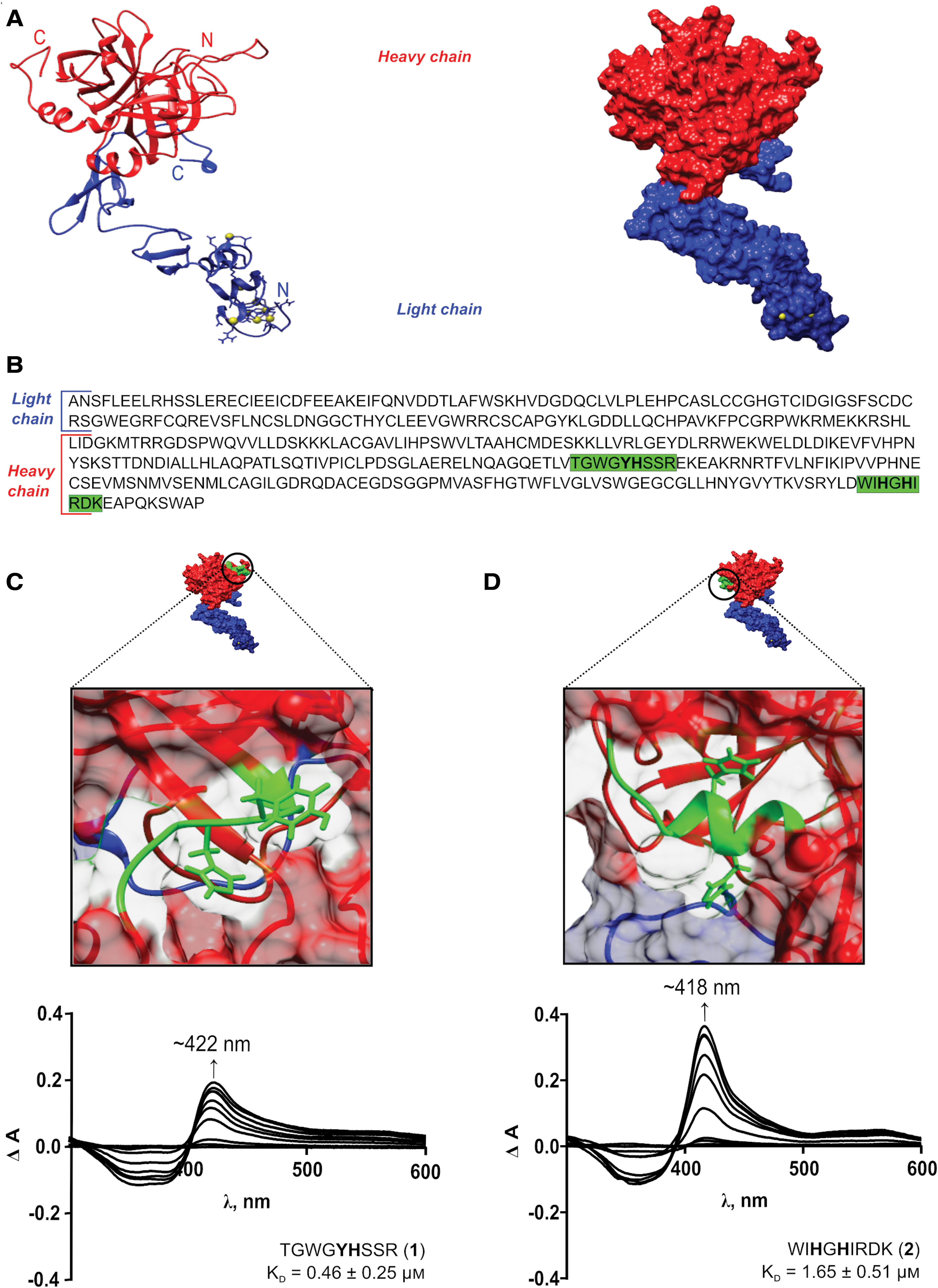
The sequence stretch of peptide
In silico study of heme-APC complex via molecular docking and molecular dynamics simulations
In silico structural modeling of APC was employed in this study, because an experimentally derived structure of the full-length APC has not been available so far (45). Earlier reported homology models of APC were performed for either the serine protease domain or the zymogen form of PC (14, 51).
In our work, the five top-ranked templates used to build the initial homology models of APC were PDB ID: 1AUT, 3F6U, 2AER, 1W0Y, 3HPT, and they had an alignment target coverage of 83%, 83%, 94%, 93%, and 80%, respectively. The final hybrid model (Fig. 2A) consisted of 405 residues [full coverage of APC(1–405), i.e., without activation peptide] and included contributions from the following initial models: 1AUT-∼(39–393), 1W0Y-∼05(1–52), 1W0Y-∼02(68–76), and 2AER-∼01(394–406) (Supplementary Table S2). To evaluate the quality of the model, Z-scores, which indicate the total deviation in energy against energy distributions derived from entirely random conformations, were used as a metric. In an ideal situation, a value of 0 is expected but in most real-world cases values close to 0 are acceptable (68). Accordingly, the overall Z-score obtained as a weighted average of the individual Z-scores of three distinct components, namely, dihedrals, one-dimensional packing, and three-dimensional (3D) packing, was found to be −1.407. Ramachandran plot analysis of the structure showed that 87.2% of the models' residues were in the most favored regions, 12.2% of residues in the allowed region, and a meagre 0.6% of the residues in the disallowed region, indicating that the final model obtained was of good quality.
As a result of focused molecular docking experiments on the HRMs
The stability of the heme-motif interaction for the motifs
With the individual motifs investigated, a 50 nanoseconds molecular dynamics (MD) simulation of heme bound to both motif
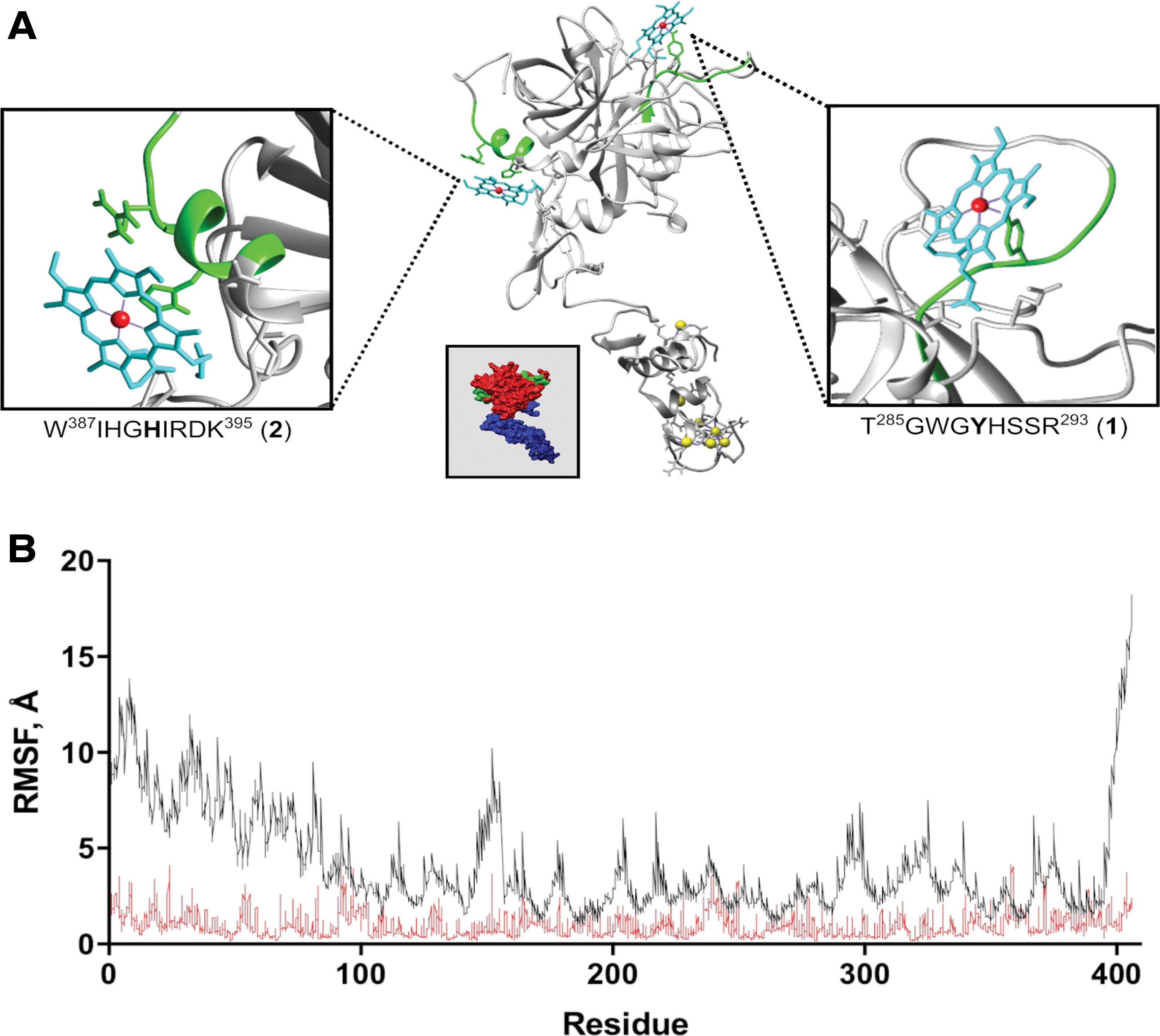
The structural consequences of heme binding to APC were assessed via two separate 300 nanoseconds long MD simulations conducted on both the heme-bound and -unbound form of the protein. We observed that the association of the two heme molecules with APC had a marked impact on the dynamics of APC by inducing a significant reduction of the inherent flexibility/mobility of the protein. The per residue root mean square fluctuation (RMSF) of the residues was used as an MD-derived measure to compare the dynamics of the heme-bound and -unbound form of APC. The RMSF values of the two simulations clearly indicate that heme binding to APC has a dampening effect on the natural dynamics of the protein (Fig. 3B). This structural impact can be hypothesized to have a functional consequence on the protein.
Heme inhibits the amidolytic activity of APC
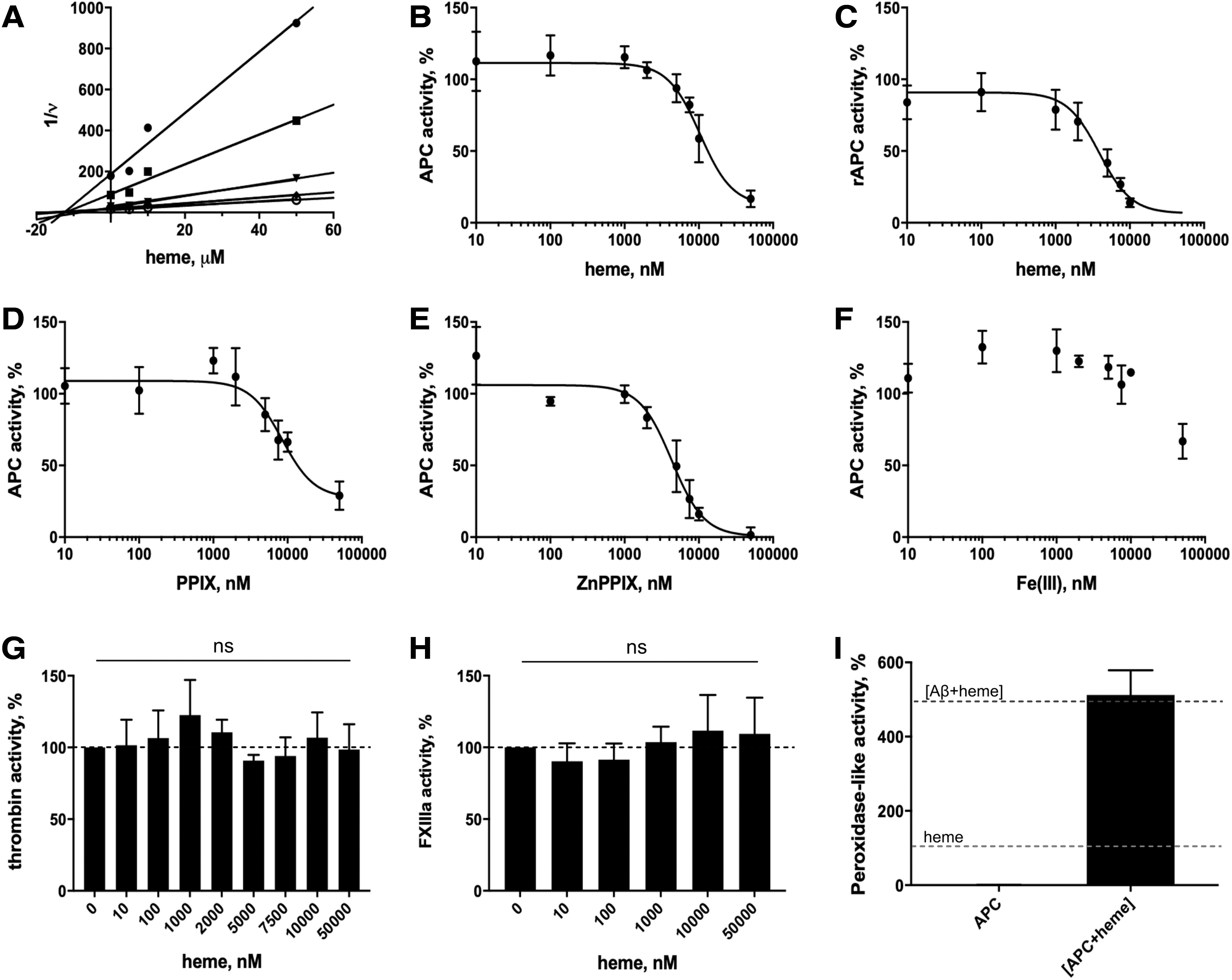
The amidolytic activity of APC was analyzed in the presence of different concentrations of p-Glu-Pro-Arg-MNA as the substrate, revealing a Michaelis-Menten constant (KM) value of 0.42 ± 0.07 mM, which is in the same range as reported for other chromogenic peptide substrates of APC (Supplementary Fig. S3) (59). In the presence of heme, the inhibition of the APC activity by heme as a competitive inhibitor revealed an inhibitor constant (Ki) value of 12.56 ± 2.31 μM (Fig. 4A). In comparison, the amidolytic activity of APC was also studied in the presence of two other porphyrin analogs, namely zinc protoporphyrin IX (ZnPPIX) and protoporphyrin IX (PPIX) as well as iron(III) chloride at concentrations ranging from 0 to 50 μM. Heme inhibited the activity of plasma-derived APC (pdAPC) and recombinant APC (rAPC) in a dose-dependent manner with a half maximal inhibitory concentration (IC50) value of 10.41 ± 1.24 μM and 3.88 ± 0.87 μM, respectively, according to the best fit (Fig. 4B, C). At the highest concentration of heme (50 μM) applied, an inhibition of 80% of pdAPC's activity was observed. A Hill slope of 2.0 suggests two binding sites for heme with positive cooperativity (n = 2). A similar effect toward pdAPC was observed after incubation with PPIX, revealing an IC50 value of 8.46 ± 1.68 μM, n = 2 (Fig. 4D). ZnPPIX had an even stronger effect by decreasing the pdAPC activity to 1.42% (IC50 = 4.33 ± 0.87 μM, n = 2) (Fig. 4E), which has been shown for zinc(II) ions as well by Zhu et al. (77). Iron (III) ions reduced the pdAPC activity only to 66.83% at a concentration of 50 μM (Fig. 4F). To compare the effect of heme on APC's activity with that on other enzymes of the blood coagulation cascade, the enzymatic activities of the serine protease thrombin and the transglutaminase factor XIIIa (activated factor XIII [FXIIIa]) were observed in the presence of heme. Both thrombin and FXIIIa were not affected by heme in their enzymatic activities (Fig. 4G, H). Indeed, these proteins were not detected as heme-binding proteins with the publicly available webserver HeMoQuest (48), indicating reliability of the program and the underlying search criteria regarding HBMs.
Heme-APC complex shows peroxidase-like activity
The peroxidase-like activity of heme-protein complexes has been reported earlier, with amyloid beta being the most prominent protein representative evoking a significantly increased activity (∼500%) compared with uncomplexed heme (2, 71). The activity of the heme-APC complex was determined by using an experimental set-up established earlier (2, 64, 71). Normalized against the basal heme activity, the heme-APC complex showed a more than fivefold higher activity (512.33 ± 66.50%; Fig. 4I) and, thus, is one of the most efficient molecules regarding this effect compared with the peptides and proteins that have been studied so far (28, 29, 64, 71).
Heme is able to inhibit the anticoagulant activity of APC
To investigate the impact of heme on the anticoagulant function of APC, the conventional activated partial thromboplastin time (aPTT)-based clotting assay was performed with plasma samples. Heme alone had no significant effect on aPPT, whereas APC (5 nM) prolonged the clotting time of normal plasma by approximately twofold (Fig. 5A). An incubation of APC with 1 μM heme did not change this effect. In contrast, the preincubation of APC with heme at 10 μM and higher concentrations caused a marked reduction of the clotting time, counteracting the anticoagulant function of APC. Thus, the clotting time was effectively shortened to that of normal plasma without addition of APC.
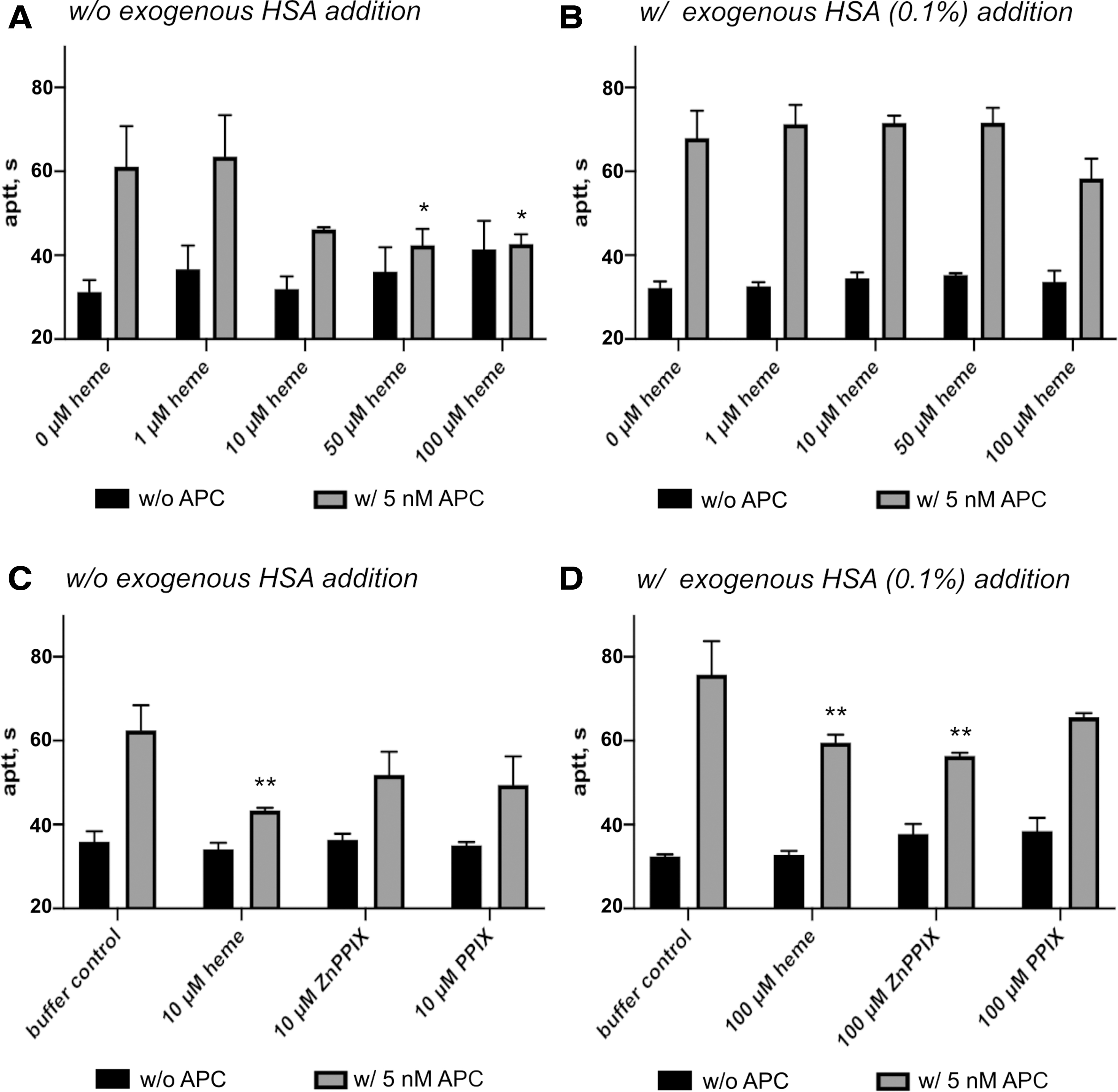
On preincubation with heme (up to 50 μM), the addition of exogenous albumin (0.1%) completely restored the clotting time to the level when only APC was applied (Fig. 5B). At 100 μM heme, however, a tendency toward inhibition of the anticoagulant function of APC was observed again (not statistically significant). Due to the solubility and high aggregation potential of heme in correlation with the required assay conditions, the effect of higher concentration could not be investigated despite the pathological relevance. Further studies demonstrated that heme could already induce a slight inhibition of APC's anticoagulant without any preincubation, which is more emphasized after a preincubation time of 15 min or longer (Supplementary Fig. S4).
Since the heme analogs ZnPPIX and PPIX were potent to inhibit the amidolytic activity of APC similar to heme, their impact on the anticoagulant function was investigated as well. Similar to heme, none of the analogs had a direct influence on aPTT. However, after preincubation with APC and the addition of this mixture to plasma, aPTT was shortened (Fig. 5C, D). In the presence of additional human serum albumin (HSA), an effect comparable to heme was observed (Fig. 5D). The lowest concentration of heme still exerts activity in the absence of HSA (i.e., 10 μM) and it seemed to be insufficient for the heme analogs (Fig. 5C). To achieve the same effect as heme in this approach, an increase of their concentration to 100 μM was required (Supplementary Fig. S5).
APC protects HUVECs from heme-mediated hyperpermeability
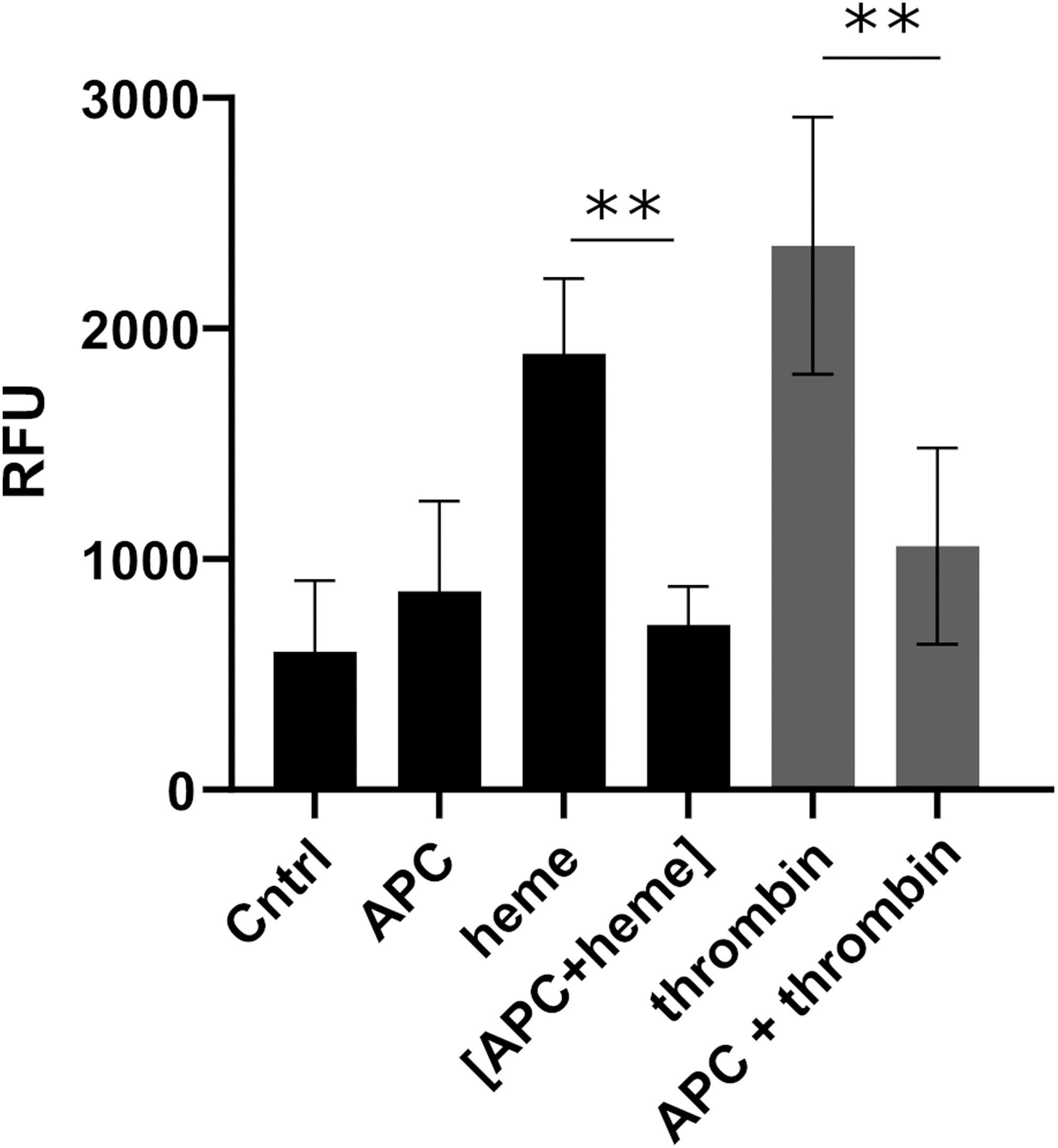
To study the influence of heme on the cytoprotective function of APC, an endothelial barrier protection assay with HUVECs was performed. Heme exposure of the HUVECs resulted in a concentration-dependent increase of fluorescein isothiocyanate (FITC)-dextran flux through the cell monolayer (Supplementary Fig. S6). In the presence of APC, the HUVECs were protected from heme-induced hyperpermeability. Therefore, the cytoprotective function of APC was not influenced by heme (Fig. 6).
Discussion
This study provides evidence that heme binds to APC and thereby impacts its function (Fig. 7). We demonstrate a heme-binding affinity for APC in the nanomolar range, which is typically found for heme-regulated proteins, as can be exemplified with the transcription repressor Bach1 (K
D = 140 nM) or heme oxygenase 2 (K
D = 33 nM [dithiol state], K
D = 350 nM [disulfide state]) (47, 76). The transient manner of heme binding to APC is illustrated by both rapid association and dissociation of the heme molecule. A stoichiometry of 1:1.3–1:2 suggested one to two binding sites for heme with either overlaying binding events, different binding affinities of the two motifs, or a mixture of specific and unspecific binding events. pdAPC represents a mixture of different glycosylated forms of APC (α, β, and γ). Different accessibility of potential binding sites through a missing glycosylation may result in different binding properties, which, in turn, may potentially affect the stoichiometry. This is further evidenced by the computational studies using a combination of molecular docking and MD simulations, suggesting heme binding to both motifs
We identified T285GWG
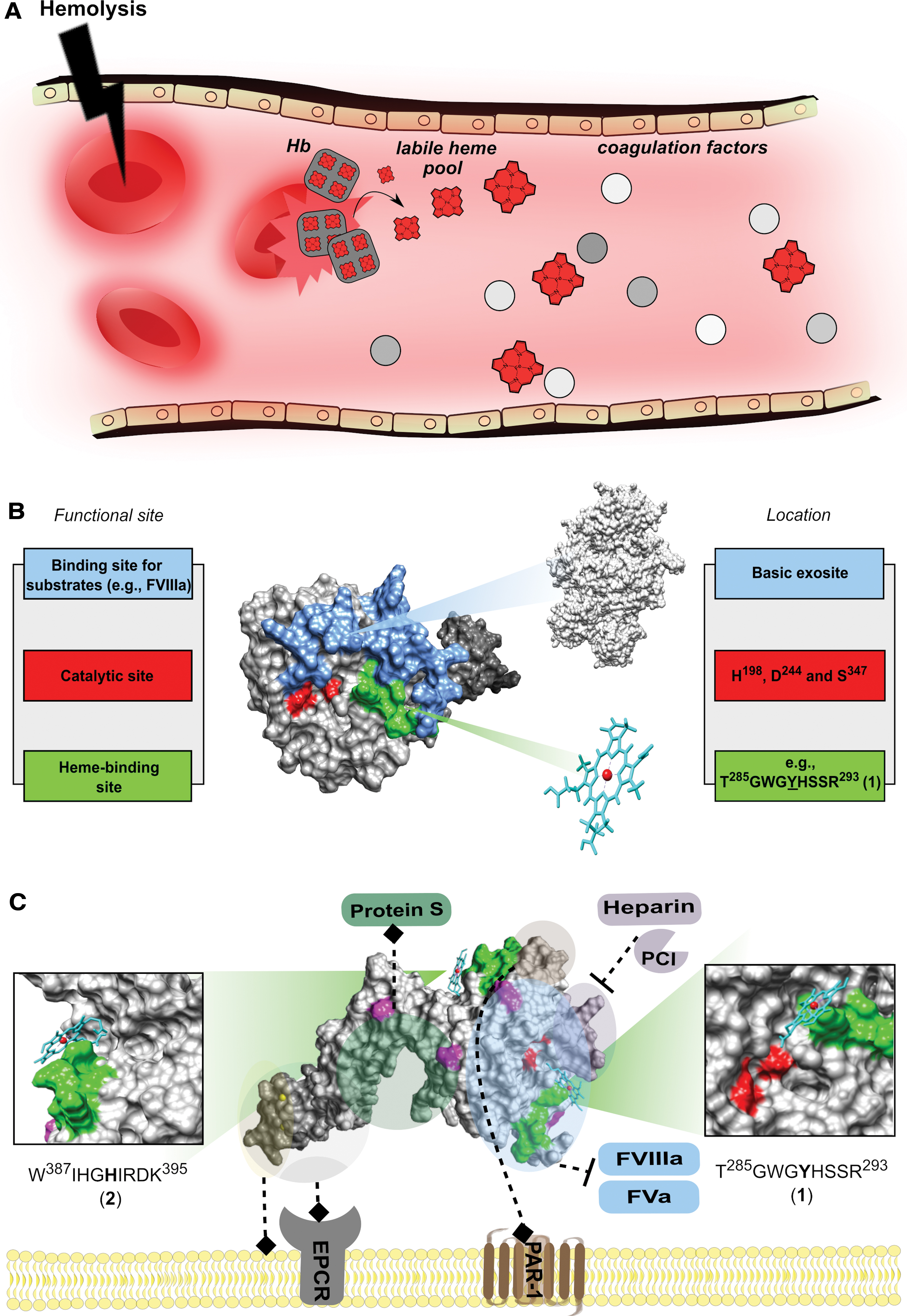
Molecular docking studies allowed for the assumption that heme binding to both motifs is possible, which fits to the aforementioned results. Moreover, two other motifs showed moderate heme-binding properties based on experimental and computational approaches. Therefore, a recruiting binding process could be conceivable. The proposed binding sites (motif
However, heme is, indeed, able to inhibit the amidolytic activity of rAPC and pdAPC with an IC50 value of 3.88 ± 0.87 μM and 10.41 ± 1.24 μM, respectively. The recombinant form merely differs from the pdAPC in the glycosylation pattern (75). rAPC often shows a higher content of the highly active β-APC (diglycosylated heavy chain) and only 50%–60% of the α-form (triglycosylated heavy chain), whereas the pdAPC contains 70%–80% of α-APC (22). In addition, the oligosaccharides in rAPC are described to be less branched, since they are only diantennary (74). Thus, the higher impact of heme on the recombinant form might be due to less hidden binding sites and an improved access of heme to these sites.
Nevertheless, the fact that heme is able to impair the conversion of a small peptide substrate by APC might be an indication for the lower APC activity measured during hemolytic crisis in SCD patients (60, 67). The APC preincubation with PPIX resulted in a similar effect regarding the amidolytic activity, which demonstrates the importance of the protoporphyrin ring system for the interaction. Interestingly, ZnPPIX, which is naturally formed in iron deficiency (41), had an even stronger effect on APC. The amidolytic activity was almost completely suppressed.
By now, ZnPPIX itself was only reported to be a potent inhibitor of HO-1 reducing APC levels in septic mice (13). However, zinc(II) ions were shown to inhibit the enzymatic activity of APC toward a peptide substrate and FVa with a binding affinity in the micromolar range (K D = 7.4 μM) (77). In the present experimental set-up, ZnPPIX seems to have an even higher effect, which again would confirm the significance of the porphyrin ring for interaction. Physiologically, increased levels of ZnPPIX accompanying iron deficiency states were earlier associated with malaria, which in the course of disease development results in hemolysis (27). Under these conditions, ZnPPIX could potentially contribute to decreased APC amidolytic activity. Our results demonstrate that the inhibitory effect is provoked not only by heme but also by other PPIX derivatives, which may lead to an even stronger effect under physiological conditions.
In further support of our hypotheses, heme and its analogs were capable of inhibiting APC's anticoagulant function in an aPTT-based clotting assay. Therefore, heme even enabled the complete elimination of the anticoagulant effect of APC. As one of the classical heme scavenger proteins, albumin binds heme and captures it under hemolytic conditions (57, 63). Thus, on addition of exogenous HSA, we could observe that APC is protected from inhibition by heme to a certain extent, fully recovering the anticoagulant function of APC. However, heme applied in the highest possible concentration with respect to the assay conditions, that is, 100 μM, again induced a reduction of APC's anticoagulant function. It can, thus, be assumed that massive local increase of heme, as occurring during hemolytic events, leads to an inhibition of the anticoagulant properties of APC (Fig. 8). This scenario is conceivable at least in case of a complete saturation of the heme scavengers, since already a comparably low heme concentration of 10 μM could induce the inhibition of APC in our approach.
Further, the heme-APC complex was found to exert a much higher peroxidase-like activity (∼512.33%) than heme only (100%), an effect that was also demonstrated earlier for the respective complexes with amyloid beta or fibrinogen (2, 28, 71). Interestingly, we recently reported a peroxidase-like activity of >360% for the complex of heme with the APC-derived peptide WIHGHIRDK (motif
This function might be particularly important during hemolytic events in sepsis, which are associated with the formation of reactive oxygen species such as H2O2 (11, 33). Further, heme itself is well known for its cytotoxic effect through induction of oxidative stress, which, in turn, has been suggested to promote coagulation (18, 42). Again, this would support the procoagulant effect of heme. In addition, APC exerts cytoprotective functions through interaction of its acidic exosite with the receptor PAR1 (21, 73). Interestingly, the potential HBM
As reported earlier (62), heme exposure of endothelial cells resulted in a concentration-dependent barrier dysfunction of the endothelial monolayer. Our results obtained using an HUVEC-based barrier assay support these findings. In the presence of APC, however, the cytotoxic effect of heme was partly neutralized, indicating that the cytoprotective functions of APC are not inhibited by heme binding. Thus, the interaction between heme and APC seems to be of bilateral nature. Although heme partially inhibits the anticoagulant functions of APC, the cytoprotective functions remain intact.
Herein, we report for the first time a direct interaction of heme with the serine protease APC, providing a first evidence for specific binding motifs as well as for functional consequences resulting from heme binding to APC. To what extent heme might influence the PC system through interaction with APC depends on time and spatial hierarchy of the simultaneously occurring reactions as well as on concentration of other heme-binding proteins (apart from heme-scavenging proteins) and their respective heme-binding affinity (Figs. 7 and 8A). However, it is tempting to speculate that heme-induced inactivation of APC contributes to the hypercoagulable state associated with severe hemolysis.
Materials and Methods
Reagents
Hemin [Fe(III) PPIX], later designated as heme, was obtained from Frontier Scientific (Logan, UT). Stock solutions of 1 mM heme were freshly prepared as described earlier (5, 7, 38, 39, 71). The stock solutions were diluted to the desired concentration in the respective assay buffer directly before use. Stock solutions of PPIX and ZnPPIX (Frontier Scientific) were prepared identically to heme. Ferric chloride (FeCl3, anhydrous; Merck, Darmstadt, Germany) was dissolved 1 mM in water (double-distilled) for stock solution and freshly diluted before use.
Plasma-derived human APC (HCAPC-0080; Hematologic Technologies, Essex Junction, VT), human α-thrombin (T6884; Sigma-Aldrich, St. Louis, MO), recombinant human factor XIIIa (T070; Zedira, Darmstadt, Germany), and HSA (A3782; Sigma-Aldrich) were commercially obtained. The recombinant human APC (Drotrecogin alfa, Xigris®) was obtained from Eli Lilly and Company (Indianapolis, IN). As a drug, Xigris was in use for the treatment of severe sepsis but was withdrawn from the market due to a lack of efficiency. It differs from pdAPC only in its glycosylation pattern, which was demonstrated to result in considerable differences in activity (74, 75). The purity of all proteins was examined by sodium dodecyl sulfate polyacrylamide gel electrophoresis.
SPR-binding studies
SPR-binding studies were performed on a Biacore T200 instrument (GE Healthcare Europe GmbH, Freiburg, Germany) as described earlier (49, 70). APC was dissolved in 10 mM acetate buffer (pH 4.5) at a concentration of 10 μg/mL and immobilized by amine coupling on a CM5 sensor chip (GE Healthcare) to a response unit (RU) of 2261 RU. Binding kinetics were determined by a titration of five consecutive injections with different heme concentrations (187.5, 375, 750, 1500, 3000 nM), prepared by dilution of the stock solution (1 mM) with running buffer. Measurements were performed at 25°C in running buffer (20 mM 2-Amino-2-(hydroxymethyl)propane-1,3-diol [Tris]-HCl, 150 mM NaCl, 0.05% Tween 20, pH 7.4) with a flow rate of 30 μL/min in the standard single-cycle kinetics method (Biacore T200 control software; GE Healthcare). Regeneration of the sensor chip surface was achieved by injection of 10 mM NaOH. Binding kinetics was evaluated by global fitting with the Biaevaluation software (GE Healthcare).
Screening and selection of potential heme-binding sites in APC
As described earlier, potential HBMs were selected as nonapeptide sequences by using the sequence-based detection of heme binding motifs (SeqD-HBM) search algorithm, which was recently manifested with the webserver HeMoQuest (5, 7, 38, 39, 48, 72). HeMoQuest is a computational approach for the prediction of transient heme binding to proteins (48). After entering the sequence of APC, 10 motifs were shortlisted based on the search criteria (a potential coordinating site [Cys, Tyr, or His], a positive net charge, and surface-exposed). Five cysteine-based motifs were excluded since the cysteine residues are involved in disulfide bridges (45), and one motif included a glycosylation site (Asn235) (22), hindering heme binding. Finally, four motifs were synthesized as nonapeptides for studying their heme-binding properties (see Synthesis, Purification, and Amino Acid Analysis of APC-Derived Peptides section) (Supplementary Table S1).
Synthesis, purification, and amino acid analysis of APC-derived peptides
Automated solid-phase peptide synthesis of APC-derived peptides
Analysis of heme binding to APC-derived peptides by UV/vis spectroscopy
Heme binding to peptides
Molecular dynamics and docking simulation studies of the heme-APC complex
First, homology modeling was employed to construct the full-length 3D structure of APC in YASARA structure (version 18.2.7) (34, 35). The program used five initial structures (PDB ID: 1AUT, 3F6U, 2AER, 1W0Y, 3HPT) as templates in the construction of 17 different initial models based on different alignment variants. A hybrid model was built by combining the best parts of the 17 initial models. Finally, the Gla domain of the hybrid model was manually replaced by the Gla domain obtained from an earlier study that contained the calcium ions at their precise locations (51). This model was energy minimized by using the steepest descents and stimulated annealing protocol as implemented by the YASARA macro
Using selected structures obtained from the production phase of the MD simulation trajectory of the model, we performed focused molecular docking studies of heme to potential HRMs of APC. The snapshots were selected arbitrarily on visual examination of the trajectory, specifically choosing ones where the coordinating residue of (H or Y) of the potential HRM was found with good surface exposure. The heme molecule was downloaded from the ChemSpider database (ChemSpider ID 16739951) (50). For each potential HRM, a cubic simulation cell (15 × 15 × 15 Å) was built around the expected coordinating residue.
The AutoDock Vina algorithm was used to run docking simulations via YASARA (35, 65). The ensemble docking approach was used by creating a protein receptor ensemble of five high-scoring solutions of the side chain rotamer network at a temperature of 298 K (65). To each rotamer, heme was docked 20 times, resulting in a hundred docking poses per run. The resulting poses were clustered to produce a list of ligand-receptor complex conformations, sorted by predicted binding energies and a 5 Å (default value used by Vina) heavy ligand atom RMSD threshold to separate the clusters.
The MD simulations of selected heme-APC docked complexes were conducted on the YASARA version 19.1.27 platform to assess the stability of heme binding to the specific HRMs, thereby validating the docking hit that was found (8, 36). Heme docking to APC was considered successful in poses where the distance between the Fe3+ ion of heme and the non-protonated nitrogen atom of histidine or the hydroxyl oxygen of tyrosine is ≤3 Å. First, selected docked complexes for each HRM were loaded into YASARA as starting structures for the MD simulations. All MD simulations were conducted in the NPT ensemble with a 2 fs time step, at 298 K and a pressure maintained at 1 bar. Periodic boundary conditions were used on the simulation cell, which was constructed to have a minimum distance of 10 Å from all protein atoms. Long-range forces were computed with a cutoff of 8 Å, and the particle-mesh Ewald method was used (12).
Three rounds of simulations were conducted for each complex, the first of which was run for 20 nanoseconds. Further, two independent validation runs for 50 nanoseconds each were conducted by creating mildly altered initial configurations of the system via energy minimizations (Supplementary Fig. S7). The first 20 nanoseconds run was used to verify whether the docked complex was stable in the first place and the subsequent two 50 nanoseconds validation runs were done to be sure that the binding persists even when the initial coordinates of the complex are altered due to the fact that MD simulations are highly sensitive to the initial configuration of the system (4). Analyses of the MD trajectories were done in both YASARA and VMD (version 1.9.3) by first converting the trajectory into the GROMACS XTC format (26, 30). Molecular graphics were created from UCSF Chimera (1.13.1) (52), and plots were generated by using GraphPad prism 8.0 software. To enable a qualitative comparison of the heme-protein interaction between various motifs, the binding energy computed by the Poisson-Boltzmann method as implemented by the
Amidolytic activity of human APC, thrombin, and factor XIIIa (FXIIIa) in the presence of heme
The impact of heme on the catalytic activity of APC was spectrophotometrically analyzed on a Multiskan GO microplate spectrophotometer (Thermo Fisher Scientific) based on a diagnostic assay, which is commonly used for the quantification of functionally active PC (Berichrom® Protein C). APC (100 nM) was preincubated with different concentrations of heme (0–50 μM) for 15 min at room temperature in the dark. Ninety microliters of the heme-APC complex was placed into a 96-well microplate (Greiner Bio-One GmbH, Frickenhausen, Germany), and 10 μL of the chromogenic substrate p-Glu-Pro-Arg-MNA (Berichrom Protein C kit; Siemens Healthcare Diagnostics, Erlangen, Germany) was added in a concentration varying from 0 to 2.5 mM. The assay was performed in HEPES buffer (100 mM, pH 7.4).
KM was obtained by nonlinear regression fitting using the Michaelis-Menten equation, whereas the inhibition constant Ki was graphically analyzed by using Dixon plot (GraphPad prism 8.0 software). Performing the assay with a constant substrate concentration of 300 μM allowed for the comparison of the effect of heme with that of other porphyrin analogs (ZnPPIX and PPIX) and iron(III) chloride. Further, the impact of heme on rAPC (Xigris) was investigated. The concentration of rAPC needed to be adapted to 50 nM. A range of heme concentrations (0–10 μM) was applied.
The same procedure was used for thrombin, but with a final enzyme concentration of 25 nM and heme in concentrations of 0–50 μM. Again, p-Glu-Pro-Arg-MNA served as the substrate. The activity of FXIIIa was monitored in Tris buffer (0.05 M Tris, 0.154 M NaCl, 0.01 M CaCl2, pH 7.5) by using H-Tyr-Glu(pNA)-Val-Lys-Val-Ile-Gly-NH2 (100 μM) as the substrate (25). The final concentration of FXIIIa was 400 nM. Heme was added in final concentrations of 0–50 μM. The increase of absorbance due to substrate conversion was monitored every 10 s for 20 min at 405 nm. The remaining activity of the enzymes as the change of absorbance per second was normalized against the basal activity without any potential ligand. Data were analyzed as the mean of triplicate, and the IC50 value was determined by fitting the curves with nonlinear regression (GraphPad prism 8.0 software).
Monitoring the peroxidase-like activity of the heme-APC complex
As described earlier (64, 71), 42 μM of heme was preincubated with 42 μM of pdAPC for 30 min in phosphate buffered saline (PBS) buffer (pH 7.4). A mixture of 3,3′,5,5′-tetramethylbenzidine (TMB; 1.66 mM) and H2O2 (163.2 mM) served as the substrate. After transfer of 10 μL of the heme-APC complex (final concentration 1 μM) to 200 μL of the substrate, the absorbance was detected at 652 nm. For comparison, the same experiment was performed with heme as well as with APC only as controls. Data were normalized against the activity of heme and evaluated as the mean of triplicates.
Anticoagulant activity of APC in the presence of heme
The impact of heme on the anticoagulant activity of pdAPC was analyzed by using an aPTT-based clotting assay, performed with the semi-automatic ball coagulometer KC10 (Amelung, Lemgo, Germany) (24). APC (5 nM) was incubated with different concentrations of heme (0–100 μM) for 1 h in Dulbecco's phosphate buffered saline (DPBS) buffer (with Ca2+ and Mg2+) or in DPBS with 0.1% HSA (with Ca2+ and Mg2+). After addition to the 10-fold volume of citrated pool plasma (in-house preparation), 100 μL of the respective mixture was immediately incubated with 100 μL of the contact phase activator Actin FS (Siemens Healthcare Diagnostics) at 37°C for 3 min followed by the addition of 100 μL of 30 mM CaCl2 solution (Siemens Healthcare Diagnostics), as described earlier (24).
As a control, pool plasma containing the same amount of heme without addition of exogenous APC was subjected to the assay system. Clotting times, given in seconds, were automatically determined by the coagulometer. Data were evaluated as the mean of triplicates (GraphPad prism 8.0 software). In a second approach, the impact of heme with the lowest potent concentration was compared with that of its analogs ZnPPIX and PPIX (10 μM in the absence and 100 μM in the presence of 0.1% HSA).
Evaluation of the cytoprotective function of APC in the presence of heme
The endothelial barrier permeability assay (Merck) was used to determine the cytoprotective function of pdAPC after preincubation with heme. As described earlier (58), HUVECs were seeded (2.5 × 104 cells/well) onto collagen-coated culture inserts of a 96-well plate and subsequently cultured in endothelial cell growth medium (ECGM, C-22010; PromoCell, Heidelberg, Germany) for 4 days to reach maximum confluency. The upper chamber contained 125 μL and the lower chamber contained 250 μL of ECGM. One hour before starting the experiment, ECGM was exchanged with phenol red-free endothelial cell basal medium (ECBM, C-22215; PromoCell). APC (20 nM) was preincubated with heme (120 μM) for 30 min in phenol red-free ECBM. Subsequently, the mixture was applied to the upper chamber and the cells were incubated for 3 h at 37°C, 5% CO2.
As controls, buffer, APC, and heme were separately incubated with the cell monolayer. After washing the cells three times with PBS, the APC/heme mixture was replaced by either thrombin (20 nM) or ECBM as a control for 30 min at 37°C. Later, the content of the upper and lower chamber was replaced by FITC-labeled dextran (75 μL of 1:40 dilution in phenol red-free ECBM) and 250 μL phenol red-free ECBM, respectively, and incubated for 20 min at room temperature. Finally, the determination of the fluorescence intensity (λex = 485 nm, λem = 535 nm) with a Synergy 2 microplate reader allowed for the quantitation of the flux through the endothelial cell monolayer at a rate proportional to the permeability. Data are presented as the mean of quadruplicates.
Statistical analysis
Ordinary one-way analysis of variance with post hoc Tukey's multiple-comparisons test was used to calculate the indicated p values. A p value of 0.05 was assumed to be significant. Calculations were performed by using GraphPad prism 8.0 software.
Footnotes
Authors' Contributions
M.-T.H., N.S.H., B.P., and D.I. conceived and designed the study. M.-T.H. performed the experiments and collected the data. N.S.H. performed the barrier protection assay. N.A., A.A.P.G., and A.B. carried out the computational studies. M.-T.H., N.S.H., B.P., A.A.P.G., A.B., and D.I. analyzed the data. B.P. and J.O. contributed analysis tools. The article was written through the contributions of all authors.
Acknowledgments
The authors like to thank Kornelia Hampel and Bastian Zimmermann (Biaffin GmbH Kassel) for SPR measurements. In addition, the authors thank Amelie Wißbrock (University of Bonn) and Sneha Singh (University Hospital Bonn) for useful scientific discussions. They are also grateful to Charlotte A. Bäuml and Sabrina Linden (University of Bonn) for technical assistance.
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
Financial support of M.-T. H. by the University of Bonn is gratefully acknowledged.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Video S1
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.