
Abstract
Significance:
An optimal supply with oxygen is of high importance during embryogenesis and a prerequisite for proper organ development. Different tissues require varying amounts of oxygen, and even within single organs, different phases of development go alongside with either physiological hypoxia or the need for sufficient oxygen supply.
Recent Advances:
Human induced pluripotent stem cell-derived organoid models are state of the art cell culture platforms for the investigation of developmental processes, disease modeling, and drug testing. Organoids modeling the development of multiple tissues were developed within the past years.
Critical Issues:
Until now, optimization of oxygen supply and its role during organoid growth, differentiation, and maturation have only rarely been addressed. Recent publications indicate that hypoxia-induced processes play an important role in three-dimensional tissue cultures, triggering multilineage communication between mesenchymal cells, the endothelium, as well as organotypic cells. Later in culture, a sufficient supply with oxygen is of high importance to allow larger organoid sizes. Moreover, cellular stress is reduced and tissue maturation is improved. Therefore, a functional blood vessel network is required.
Future Directions:
In this review, we will briefly summarize aspects of the role of oxygen during embryonic development and organogenesis, present an update on novel organoid models with a special focus on organoid vascularization, and discuss the importance of complex organoids involving parenchymal cells, mesenchymal cells, inflammatory cells, and functional blood vessels for the generation of mature and fully functional tissues in vitro. Antioxid. Redox Signal. 35, 217–233.
Introduction
Organoids are artificial three-dimensional (3D) microtissues that can be grown in cell culture and serve as in vitro platform for studying development, disease modeling, toxicity testing, and drug screening. Organoids originate either from pluripotent stem cells (PSCs) or from primary organ-specific progenitors and establish their tissue architecture by self-organization in 3D suspension culture. They recapitulate several aspects of embryonic development and fulfill at least some organ-specific functions.
Within the past years, a broad range of these tissue models was developed, including gastrointestinal (124), brain (66), lung (32), liver (135), or kidney (133) organoids. Culture conditions were optimized to reach higher tissue maturation and organ-specific function. For that reason, complex and well-defined cell culture media supplemented with specific cytokines or small molecules were developed, and the effect of cell numbers per organoid or embedding in different extracellular matrices was explored. However, the role of oxygen has only rarely been addressed so far. With regard to organoid models, the availability of oxygen is interesting in two ways:
During early embryogenesis and fetogenesis, physiological hypoxia is an important driver of development and acts as a developmental morphogen triggering multilineage communication. For that reason, it is likely that similar processes play an important role during proper organoid development in vitro as well. While initial cellular differentiation and morphogenetic processes partially depend on physiological hypoxia, organ growth, maturation, and function at later developmental stages require a proper supply of oxygen via a well-established vascular system. Conventional organoid models lack a functional vasculature, and diffusional supply with nutrients and oxygen is limited to a range of ∼150 μm. For that reason, the size and long-term growth of organoids are limited, and cellular stress as well as substantial cell death in the organoid center are frequently observed. For that reason, strategies that allow for proper organoid vascularization and connection to a microcirculatory system are currently of high interest and will be discussed in detail later in this review article.
This indicates that the in vitro generation of mature organoids depends on a timely controlled optimal oxygen tension that needs to be determined and carefully adjusted for different stages of organoid maturation.
Oxygen Availability in the Adult and Fetal Body
Before we will focus on organoid cultures, we would like to briefly describe oxygen levels in the adult body and during embryonic and fetal development and give an introduction on how oxygen pressure is sensed and translated into cellular action. Moreover, we will give examples on how oxygen pressure influences stem cell behavior. Both embryonic and adult stem cells are the basis of all organoid cultures.
In the adult body, partial oxygen pressure (PO2) within the arterial vascular system ranges from 75 to 100 mm Hg PO2. The PO2 in the tissues (PtO2) depends on oxygen availability and diffusion and is characteristic for different organs (e.g., >100 mm Hg PtO2 in the lung alveolus, 30–48 mm Hg PtO2 in the brain, or 30 mm Hg PtO2 in the cornea) (93). Many stem cell niches within the adult body are characterized by hypoxic conditions. Hypoxia is discussed to induce signaling pathways (e.g., Wnt signaling) (83), which regulate long-term stem cell maintenance and quiescence (37). However, in vitro experiments suggest that different oxygen levels promote and direct cellular differentiation (42, 47, 103, 118).
In the developing conceptus, the level of available oxygen varies between different tissues and among different stages of embryonic and fetal development. The first weeks of human pregnancy occur under hypoxic conditions until the placenta and the embryonic cardiovascular system are fully established (92). At later stages, the PO2 of oxygenized blood coming from the placenta to the fetus via the umbilical vein is about 35–45 mm Hg depending on the gestation stage, which is considerably less than in maternal blood (90). The oxygenized blood is preferentially delivered to the coronary arteries and the brain via a system of shunts and returns to the placenta via the umbilical arteries with a PO2 of 24–29 mm Hg (2).
Even at these later developmental stages, distinct regions with hypoxic environment can still be detected (70). This is due to the generally reduced oxygen availability, the fast expansion of fetal tissue, and the high metabolic rate of proliferating and differentiating progenitor cells. Moreover, vascularization of newly formed tissues mostly occurs with a certain delay and is triggered by hypoxia itself (104, 119).
The limited availability of oxygen has an effect on cellular metabolism and gene expression, which again impacts proliferation, differentiation, migration, and cellular function. For that reason, it is not surprising that the level of available oxygen is discussed as a driving force for several developmental processes.
But how are oxygen levels sensed and translated into cellular action? In 1995, Guang Wang and Gregg Semenza first purified and characterized hypoxia-inducible factor 1 (HIF-1), a protein that was found to induce erythropoietin (Epo) expression in the human hepatoma cell line Hep3B (60) as a reaction to hypoxic conditions (143).
The HIF transcription system is composed of two ubiquitously expressed proteins, HIF-α and HIF-β (aryl hydrocarbon receptor nuclear translocator [ARNT]), which can form a heterodimer that acts as functional transcriptional regulator (76). Three closely related HIF-α paralogs are described within the human genome: HIF-1α, HIF-2α, and HIF-3α. HIF-1α and HIF-2α have a comparable protein structure and are similarly regulated, whereas HIF-3α is less closely related. HIF-α is unstable under normoxic conditions, due to the fact that it is hydroxylated by prolyl- (15) and asparagyl-hydroxylases (67).
The hydroxylated protein can bind the von Hippel–Lindau tumor suppressor protein (VHL), which leads to ubiquitination of HIF-α and subsequent proteasomal degradation (82). The hydroxylases utilize oxygen as a co-substrate, and therefore, a reduced availability of oxygen leads to a decreased HIF-α degradation and expression of HIF target genes. Around 100 target genes were identified, for example, vascular endothelial growth factor (VEGF) (75), Epo (115), inducible NO synthase (iNOS) (95), or mir-210, a microRNA that regulates normoxic gene expression (51).
Oxygen availability can also change the epigenetic signature of a cell [for review, see Camuzi et al. (18)]. On the one hand, this effect can be mediated indirectly by HIFs, which induce the expression of epigenetic modifiers such as the enzymes methionine adenosyltransferase 2A (MAT2A) (74), DNA (cytosine-5) methyltransferase (DNMT1 and DNMT3B) (145), or ten-eleven translocation methylcytosine dioxygenase (TET) (78). On the other hand, the effect can be also directly driven by oxygen-sensitive proteins such as TETs. TETs induce DNA demethylation and their activity is reduced by low oxygen levels (140).
Moreover, oxygen availability regulates cellular metabolism, which in turn has an impact on the epigenome, for example, via intermediate metabolites of the tricarboxylic cycle, such as α-ketoglutarate or NAD+ [for comprehensive review, see Etchegaray and Mostoslavsky (33)].
The Role of Hypoxia During Tissue Development
As discussed above, hypoxic regions can be identified throughout the early developing embryo, for example, in the neural tube, the developing heart, and the intersomitic mesenchyme. The hypoxic tissues show high protein levels of HIFs and expression of VEGF. Moreover, strong angiogenic activity can be observed (70). This is of relevance as a proper endothelial network has been reported to promote organogenesis in vivo (65, 81).
HIF-1α-deficient mice die around embryonic day (ED) 11 and show neural tube defects, heart malformation, and cell death in the cephalic mesenchyme (53). Moreover, a lack of cephalic vascularization and a reduced number of somites was observed (110). A similar phenotype was also described in mice deficient for the HIF-1β subunit protein ARNT (76). The basic helix-loop-helix (bHLH) transcription factor ARNT forms a heterodimer with HIF-1α in response to oxygen deprivation.
Tissue-specific deletion of HIF-1α using different Cre (causes recombination)-driver mice demonstrated the essential requirement of this protein and the impact of hypoxia on normal development. In the following, a few examples are given:
Lung development
A deletion of HIF-1α in the lung by human surfactant protein C (SP-C) promoter-driven Cre expression resulted in defective maturation of the lung epithelium leading to symptoms similar to fetal respiratory distress syndrome (111). The same finding had been reported before in mice lacking HIF-2α and VEGF treatment successfully rescued the defect (22).
Mammary gland development
During pregnancy, the mammary tissue is remodeled and the mammary glands rapidly expand and mature to prepare breast milk. Mice with a HIF-1α deficiency specifically in mammary gland epithelium demonstrate that HIF-1α is essentially required for this process. The genetically altered animals show impaired mammary gland differentiation resulting in lactation failure and altered milk composition (114).
Heart development
The development of the mouse heart between days 8 and 10 occurs within a hypoxic environment and is characterized by high levels of HIF activity in cardiomyocytes. Intriguingly, mice deficient for HIF-1α in cardiomyocytes die around ED 11–12 and reveal defects in looping morphogenesis (63), a defect that has been previously described in the global HIF-1α knockout (23, 53, 110).
Neural crest delamination and migration
The cells of the neural crest are a highly migratory cell population deriving from the neural plate border. At the time point of neural tube closure, these cells delaminate and undergo an epithelial-to-mesenchymal transition (EMT). The resulting migratory neural crest cells start moving to different sites of the body building the peripheral nervous system and differentiate into, for example, the cranial mesectoderm or skin melanocytes. Different publications link hypoxia and HIF activity to neural crest EMT (7, 113), migration (7, 23), and peripheral nervous system development (12).
These data clearly show the relevance of hypoxic niches and HIFs during embryonic and fetal development.
Multilineage Interaction
In most cases, although only specific cell types react to low oxygen pressure, the response to that stimulus is happening on the tissue level. It provokes a series of multilineage interactions, often between stromal components, such as blood vessels and mesenchymal cells and the tissue-specific cell type. Mostly, these are epithelial/mesenchymal interactions.
In many cases, a mutual interaction between cells of the vascular wall (endothelial cells or pericytes) and organ-specific epithelial cells is the driving force of tissue maturation.
A good example is the development of the lung. Lung development happens in a hypoxic environment (70) and requires the presence of HIF-1α (111), which is mostly expressed by the epithelial cells (44). Hypoxia stabilizes HIF-1α, which induces the expression of HIF target genes such as VEGF by cells of the epithelium. The VEGF receptor (VEGFR) is expressed on endothelial cells and in the mesenchyme, and VEGF stimulates blood vessel growth and branching (28) (Fig. 1). In turn, endothelial cells secrete factors such as thrombospondin 1 (TSP1) that induce differentiation of lung epithelial stem cells promoting alveolarization (69) (Fig. 1).
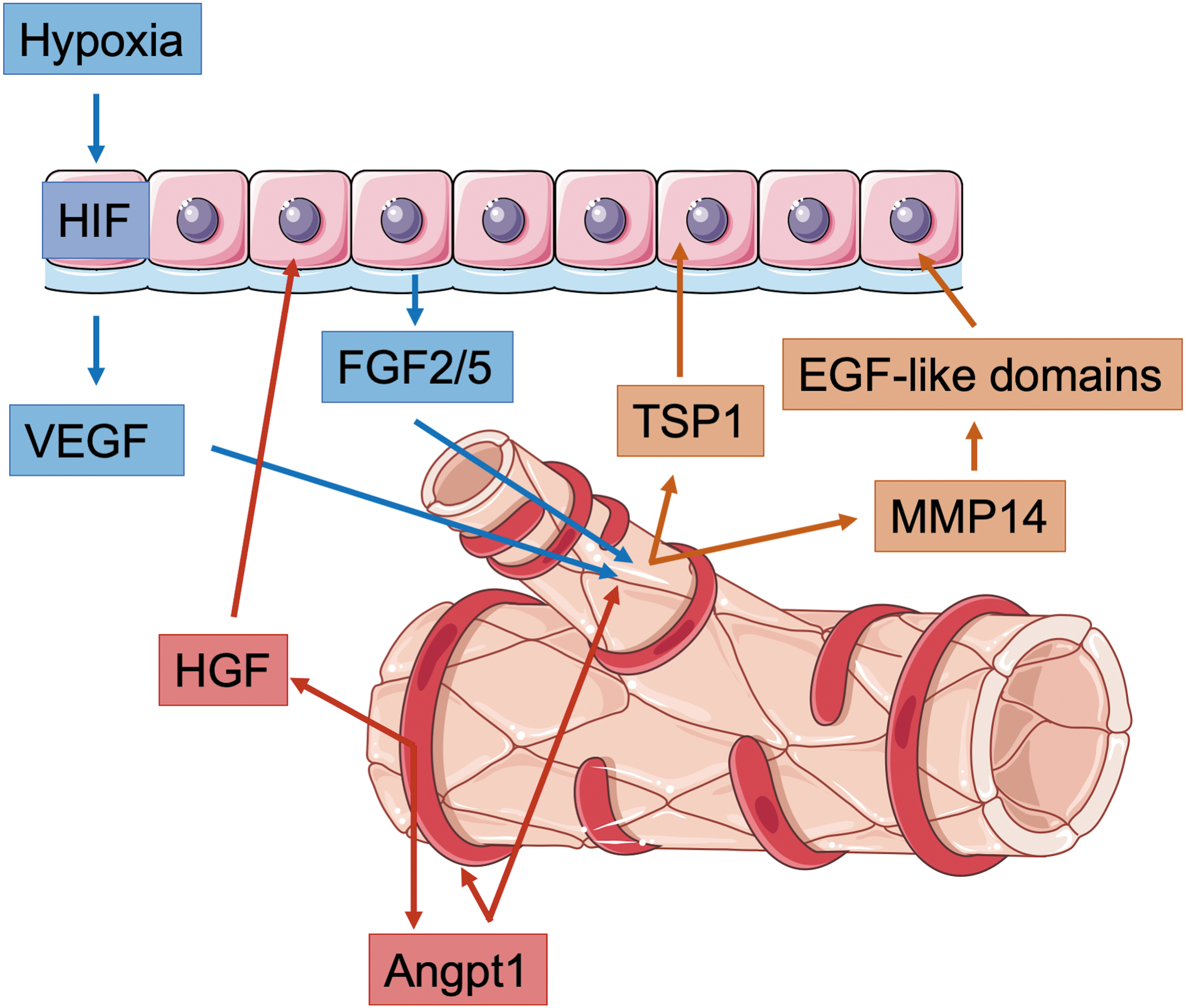
In a model for lung regeneration, it was shown that VEGF triggers endothelial cells to produce matrix metalloproteinase 14 (MMP14) (30). MMP14 expression in endothelial cells is further stimulated by fibroblast growth factor (FGF) 2 and FGF5 secretion from basal cells within the lung epithelium (29). MMP14 activity unmasks cryptic epidermal growth factor (EGF)-like domains stimulating the EGF receptor on epithelial progenitor cells, which lead to an expansion of the epithelial cell pool, a prerequisite for restoration of alveolar structure and pulmonary function (Fig. 1).
Another example that demonstrates the importance of interlineage cross talk in the lung is the finding that a depletion of matrix GLA protein (MGP) in lung endothelial cells during early development instructs the pulmonary epithelium to undergo hepatic differentiation. The absence of MGP allows bone morphogenetic protein (BMP) 4 to act on the epithelial and endothelial cells. BMP4 induces VEGF in the epithelium and VEGFR1 in the endothelium. VEGF then induces high levels of hepatocyte growth factor (HGF) expression by the endothelium driving hepatic differentiation in lung epithelial cells (153).
Besides endothelial cells, pericytes also play an important role during the vascular/epithelial cross talk regulating alveolar development. Lung pericytes release angiopoietin, which acts on endothelial cells and regulates angiogenesis and vascular integrity as well as HGF, which acts on the lung epithelium to drive alveologenesis (55) (Fig. 2).
The Impact of Oxygen on Preimplantation Development In Vivo and PSCs In Vitro
In the following paragraphs, we would like to summarize the current knowledge about the role of oxygen on the earliest stages of embryonic development before implantation of the conceptus into the uterine decidua. Moreover, we will introduce PSCs, a cell type classically derived from the preimplantation embryo, and discuss the impact of oxygen pressure on PSC derivation and culture. These clinically highly relevant cells can differentiate into every cell type of the body and serve as the basis of many currently used organoid models.
Human embryonic development begins with the fertilized egg. During that developmental phase, the resulting totipotent zygote is covered by the zona pellucida. The cells of the early embryo divide and start to form intercellular contacts from the 8-cell stage on, a process called compaction. The cells of the resulting morula partially undergo polarization and differentiate into two populations: the trophoblast and the embryoblast. Moreover, a fluid-filled cavity, the so-called blastocoel, forms and the cells of the embryoblast undergo a second lineage segregation forming the epiblast and the hypoblast. The embryo is now referred to as blastocyst.
At ED 5.5, the blastocyst hatches from the surrounding zona pellucida and eventually implants into the uterine decidua, which marks the endpoint of preimplantation development. Such early developmental steps take place in the oviduct as well as from approximately day 3 on in the uterine cavity. The oxygen tension within the female reproductive tract is very low [11–14 mm Hg O2 in the rhesus monkey (36); 10–15 mm Hg O2 in human (94, 155)] indicating that early development takes place under hypoxic conditions. Indeed, the first study that reported growth of human blastocysts in culture used reduced oxygen conditions (5% O2) in the incubator (127).
Today, several in vitro fertilization laboratories use 5% O2 tension and a lowering to 2% after day 3 is discussed to be beneficial (86). Different studies investigated the impact of oxygen on the in vitro growth and development of human preimplantation embryos (14, 56, 89, 100, 152). Some of these studies demonstrate beneficial effects, others do only observe minor differences. The question if switching culture conditions to low oxygen concentrations significantly improves the clinical outcome of assisted reproduction is still a controversially debated issue.
Human PSCs, also referred to as embryonic stem cells (ESCs), are derived from the inner cell mass of the blastocyst-stage embryo (141). They represent an epiblast-like pluripotent cell type (139) that harbors an enormous potential for several clinical applications and can be used as a basis for disease modeling, toxicity testing, and drug screening (109). Moreover, these cells are a promising cell source for bioprinting and tissue engineering purposes (68) and represent the basis for many organoid models (136). This is due to the fact that PSCs can self-renew and proliferate indefinitely and can be differentiated into any cell type of the body.
Moreover, when cultured as 3D spheres in suspension culture and stimulated with the right cytokines, PSCs differentiate and self-organize into different types of functional microtissues, the so-called organoids, which recapitulate aspects of embryonic development. Several protocols have been developed within the past decade that allow for the generation of, for example, gastrointestinal (124), brain (66), lung (32), liver (135), or kidney (133) organoids. They represent a powerful cell culture platform for the investigation of human development as well as the origin and progression of disease.
Recently published protocols even enable the modeling of early stages of pre- and post-implantation development such as the blastocyst, the egg cylinder, and the gastrulating embryo in cell culture allowing insights into the earliest mechanisms of embryonic development (8, 48, 106, 144, 121, 160, 161).
The advent of induced pluripotent stem cells (iPSCs) made human PSCs available to almost any laboratory, solved many ethical issues regarding their derivation and use, and allowed for the generation of patient-specific PSC lines carrying disease-associated mutations from skin biopsies or blood samples (130, 131, 158).
Although PSCs have been successfully cultured under environmental 21% oxygen tension (160 mm Hg O2) in the incubator, it is clear that from a developmental point of view, this is not their physiological environment. Indeed, culturing PSCs at lower oxygen tensions reduces spontaneous differentiation and increases proliferation and expression of pluripotency markers (34, 39, 101).
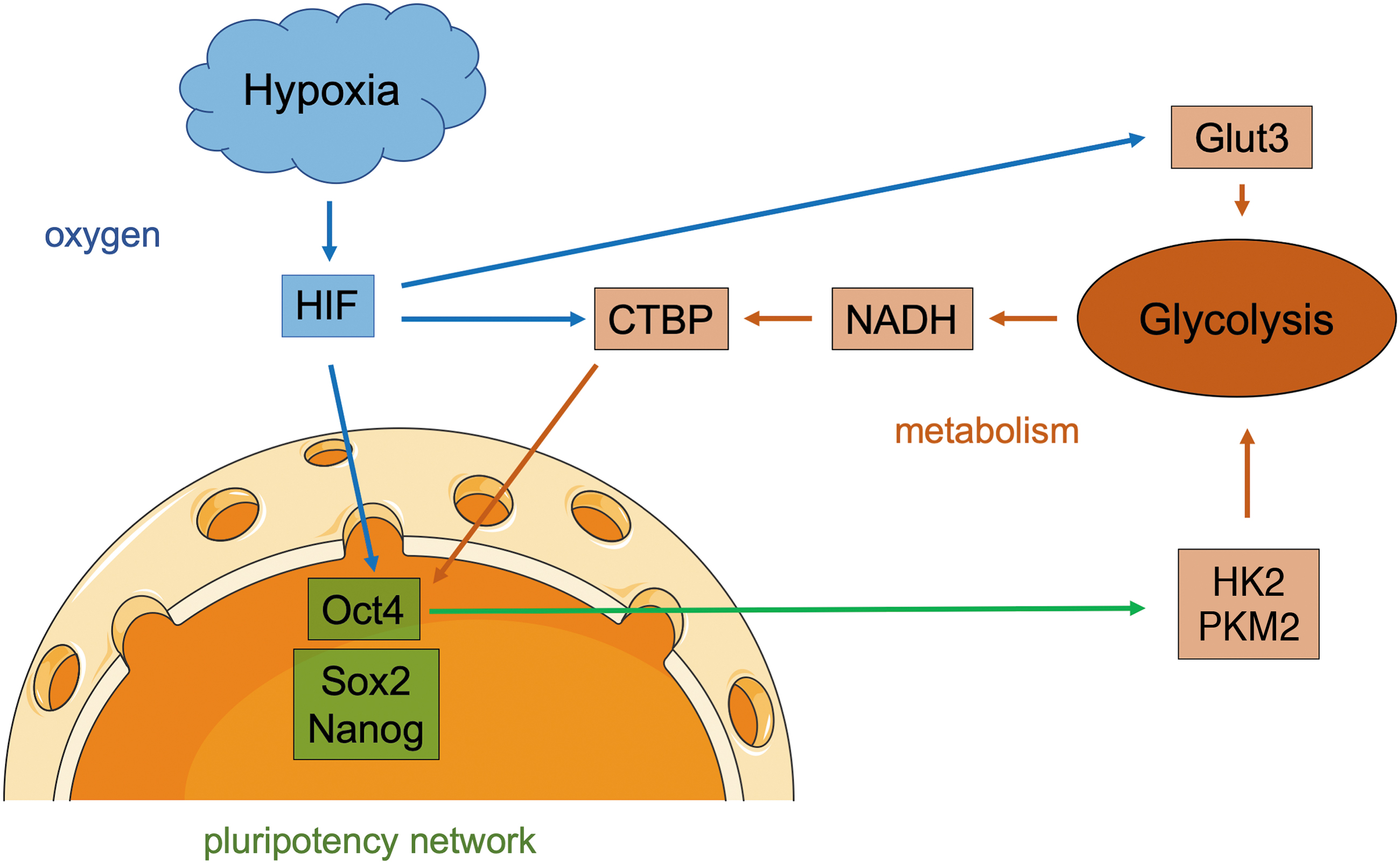
The hypoxia-induced factor HIF-2α was shown to directly bind the octamer-binding transcription factor 4 (Oct4) promotor and HIF-2α deficiency led to altered stem cell function in vitro and embryonic development in vivo (24) (Fig. 2). Moreover, hypoxia upregulates the glucose transporter Glut3 and alters the metabolism in PSCs, in particular by increasing glycolytic activity resulting in higher glucose consumption rates and more lactate production (38) (Fig. 2).
A recent publication indicates that hypoxia-induced HIF-2α expression upregulates target genes such as the pluripotency markers Oct4, SRY (sex determining region y)-box 2 (Sox2), and Nanog as well as C-terminal binding proteins (CTBP). These glycolytic sensors are activated by cytosolic nicotinamide adenine dinucleotide (NADH) (64), which is strongly produced during glycolysis. CTBP dimerizes NADH dependent and acts as transcriptional regulator contributing, for example, to a further increase in pluripotency marker expression (3) and therefore directly links metabolic activity to gene expression (Fig. 2). In turn, core pluripotency transcription factors, in particular Oct4, positively regulate glycolysis directly by upregulation of glycolytic enzymes such as hexokinase 2 (HK2) and pyruvate kinase M2 (PKM2) and forced overexpression of these enzymes in PSCs hampers their differentiation preserving the pluripotent state (58) (Fig. 2).
Interestingly, hypoxia can even drive the re-entry of already committed cells back into the pluripotent state by inducing de-differentiation processes (79).
Besides maintaining the pluripotent state, hypoxia has been further shown to enhance the induction of PSCs from somatic cell types (157). This could be partially explained by the fact that reprogramming cells to pluripotency requires a shift from oxidative to glycolytic metabolism and that HIF activity is required for that process (80).
Cellular reprogramming can be also induced in vivo by direct delivery of reprogramming factors into the adult organism, for example, via viral particles (85) or nanotransfection (40). Interestingly, in vivo reprogramming appears to be very efficient and in vivo reprogrammed iPSCs are closer to ESCs than in vitro produced ones. In vivo reprogramming even results in the generation of totipotent cells (1). Tissue damage and senescence have been discussed as important drivers of in vivo reprogramming (88); however, physiological oxygen levels or even hypoxic tissue niches might play an important role as well.
Hypoxia in the Adult Stem Cell Niche
Organoid models can be either derived from PSCs or directly from primary organ-specific adult stem cells. In the last paragraph, we described the impact of hypoxia on PSCs. In this paragraph, we would like to make a small excursion to adult stem cells.
Knowledge gained within the past years demonstrated that precisely regulated oxygen availability is not only relevant for the preservation of the pluripotent state but also stem cell maintenance in the adult stem cell niche as, for example, demonstrated for neural and myogenic progenitors. An effect that appears to be Notch-dependent (46). This fits to the observation that different adult stem cell niches show hypoxic conditions, for example, the hematopoietic stem cell niche in the bone marrow (125) or the neural stem cell (NSC) niche in the subventricular zone (25).
Data measuring HIF-1α levels in different adult stem cells reveal HIF stabilization in mesenchymal stem cells (MSCs) (96), NSCs (83, 108), and hematopoietic stem cells (91). Interestingly, HIF-1α is stabilized in some stem cells also under aerobic conditions (91) probably due to high messenger ribonucleic acid (mRNA) expression levels (96) or protective mechanisms that prevent their degradation (108). In this context, different cytokines, for example, stem cell factor (SCF) or thrombopoietin, have been shown to stabilize HIF-1α even under normoxia (99, 156), which leads to activation of HIF target genes (59). In NSCs, HIF-1α is essentially required for cellular maintenance and moreover promotes proangiogenic signaling, maintaining the integrity of their vascular niche (71). The endothelial cells in term stimulate NSC self-renewal (117).
Studies investigating hematopoietic stem cells suggest that HIF signaling, mediated either by the hypoxic niche (125) or by cell intrinsic mechanisms (91), seems to be required to keep stem cells in an undifferentiated and quiescent state (137) and to program their metabolism in a way that these cells mainly rely on glycolysis, which reduces intracellular reactive oxygen species (ROS). Low ROS levels prevent stem cells from DNA and protein damage and mitochondrial dysfunction, which leads to cellular aging and senescence. Moreover, ROS can directly alter transcription via Nrf2, which impacts cellular proliferation and differentiation (57). Additionally, cellular metabolism directly modulates the epigenome, for example, via intermediate metabolites of the tricarboxylic cycle such as α-ketoglutarate (19) [for review, see Lisowski et al. (73)].
Intriguingly, increasing the glycolytic flux has been demonstrated to maintain hematopoietic stem cells in an undifferentiated state (45). For that reason, a hypoxic environment as well as a precisely controlled metabolic program could be the key for efficient long-term maintenance of hematopoietic stem cells in vitro (61).
Besides maintaining stemness, modulation of oxygen availability and activation of HIFs also context dependently favors specific differentiation routes among others, for example, neural progenitor cell differentiation is directed more to the glial than the neuronal lineage under hypoxic conditions (151, 154).
These examples show the direct effect of hypoxia and/or HIFs on gene expression and stem cell behavior. Similar mechanisms are also playing an important role in tissue development during embryogenesis.
The Impact of Oxygen Availability on Organoid Models
Human embryologic events can be modeled in vitro using the so-called organoid cultures. These can be generated by growing either pluripotent or adult stem cells in 3D suspension cultures supplemented with instructive cytokines or small molecules and embedded into extracellular matrix. Several model systems such as intestinal, brain, liver, lung, and blood vessel organoids were described within the past decade. Morphogenetic events occurring within these microtissues closely mimic several aspects of real embryonic development. For that reason, organoids are promising platforms to study the effect of hypoxia on human cellular differentiation and morphogenesis in a tissue context.
Within the past decade, organoid cultures have been optimized with regard to culture media composition and matrix embedding; however, the impact of oxygen availability has been only rarely addressed so far.
In the following paragraphs, we would like to:
- give some examples showing the impact of oxygen on organoid cultures
- describe hypoxia-induced multilineage communication using the example of liver organoids
- introduce strategies for organoid vascularization to assure a sufficient oxygen supply and reduce cellular stress and enable long-term growth of larger organoids
- briefly discuss studies using organoids to study the detrimental effects of pathological hypoxia on tissue development.
Oxygen Availability Has a Significant Impact on Organoid Development—Some Examples
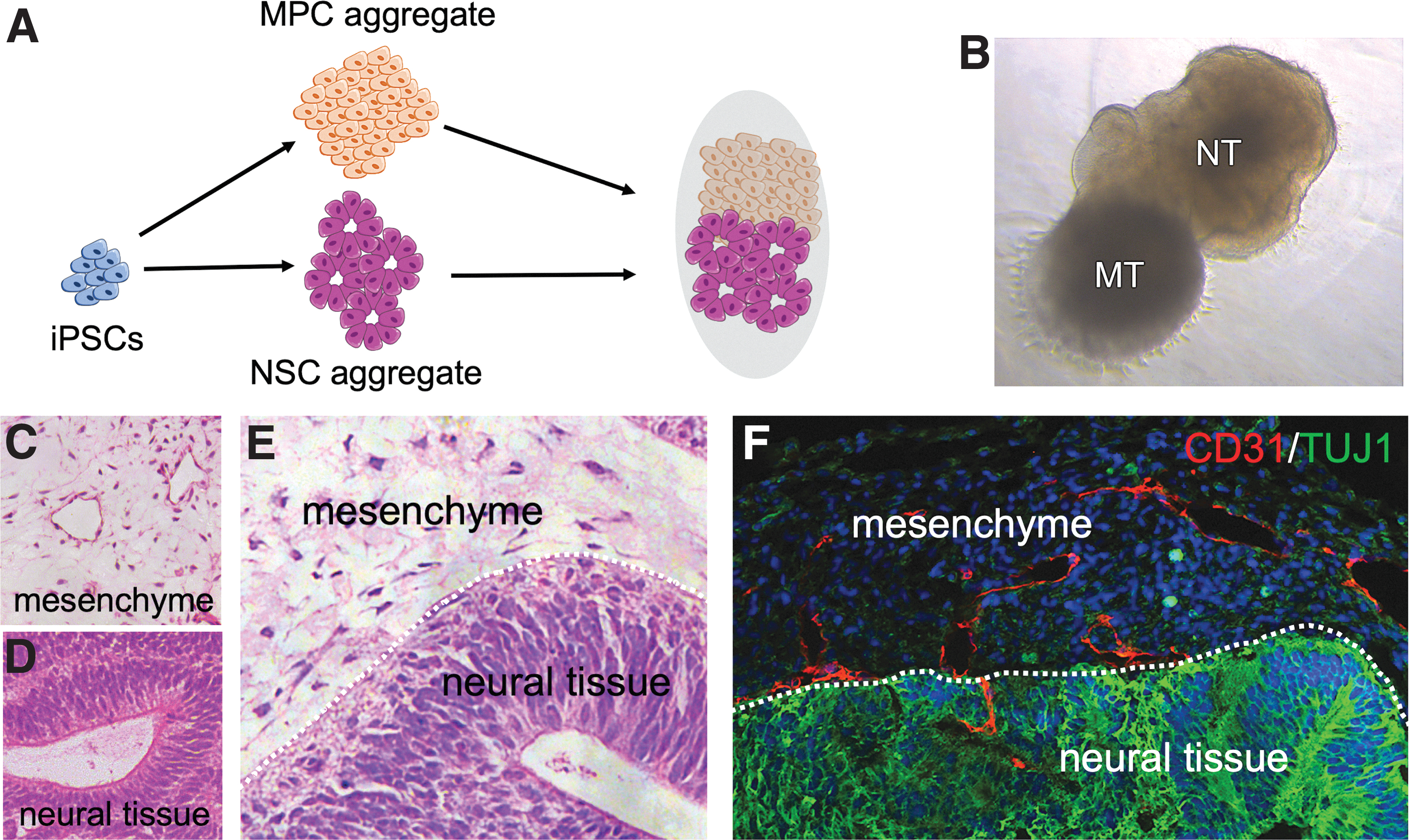
We recently published a study describing the generation of complex human organoids including vascular networks (148, 150). For that purpose, we differentiated mesodermal progenitor cells (MPCs) from human iPSCs by activating Wnt and BMP signaling. The resulting Brachyury (T) positive progenitor cells were cocultured with either tumor cells or neuroectodermal progenitor cells to generate vascularized tumor or neural organoids. Interestingly, we observed that proper vascularization in tumor organoids essentially requires hypoxic conditions. Similar to the tumor environment in vivo, hypoxia in organoids probably stabilizes HIF-1α resulting in VEGF expression by tumor and stroma cells, which induce vasculogenesis and angiogenesis (84, 116) finally leading to organoid vascularization.
Interestingly, a need for hypoxic conditions to induce proper vascularization was not required in coculture with neuroepithelial progenitor cells (148). This might be due to the fact that HIF-1α is stabilized in NSCs also under aerobic conditions leading to a continuous VEGF expression that induces and orchestrates vascular network formation (107, 108) independent of hypoxia.
A second example of how oxygen influences organoid differentiation comes from studies investigating the generation of retinal organoids. The authors show that optimization of oxygen tension accelerates and improves the generation of photoreceptors (20, 21). Interestingly, within the first week of differentiation, hypoxic conditions strongly support the differentiation outcome. However, in later phases of organoid growth, a sufficient supply with oxygen is required to support proliferation and final differentiation/maturation (31, 41). This example well demonstrates that oxygen tension has to be carefully adjusted for different stages of organoid maturation. This can be achieved in specially designed rotating bioreactors (31). Moreover, oxygen-releasing biomaterials could be an interesting option as well (129).
The third example comes from a recent publication on artificial embryo-like structures. As described above, early embryo-like entities can be derived from PSCs in vitro. One way to achieve this is the coculture of epiblast stem cells and trophoblast stem cells (138), which results in the formation of blastocyst-like structures, the so-called extended pluripotent stem cells (EPS)-blastoids (122). As already mentioned before, early embryonic development occurs under hypoxic conditions (92). For that reason, the influence of oxygen pressure on EPS-blastoid development was tested, and indeed, switching from 20% to 5% oxygen increased the frequency of cystic structure formation from 2.5% to 15.17% (122). This observation is in line with results from mouse preimplantation embryos cultured under different oxygen tensions in vitro (142).
Another example is intestinal organoids. A recent study describes the injection of Escherichia coli bacteria into the organoid lumen, which resulted in hypoxic conditions within the organoid. The authors observed bacterial contact dependent but also hypoxia-induced processes, which resulted in increased antimicrobial peptide production, maturation of the mucus layer, and improved barrier function (49).
The last example is hepatic organoids. These will be discussed in more detail in the next paragraph. Different recent studies describe that timely controlled oxygen delivery supports liver organoid formation and maturation (5, 35).
Hypoxia Induced Multilineage Communication Between Vascular and Epithelial Compartments in Liver Organoids
A recent study demonstrates that optimal hypoxia within a defined time window regulates differentiation of human iPSC-derived liver organoids via HIF activation and transforming growth factor (TGF) β signaling (5).
HIF activity is found in the developing liver bud in vivo, before the establishment of fetal circulation, and therefore, early processes of hepatoblast differentiation, proliferation, and delamination might depend on hypoxic conditions during that time window (72). The same is also true for other tissues, for example, the kidney (9).
In the complex organoid model, hypoxia upregulates VEGF expression in endothelial, mesenchymal, and hepatic cell types. However, only endothelial cells express VEGFR2 (KDR) and can respond to the signal, which results in increased vascular sprouting (17) (Fig. 3). This is of importance as a proper endothelial network has been reported to promote organogenesis in the liver in vivo (81). Moreover, moderate hypoxia in liver organoids suppresses TGFβ1 and TGFβ2 signaling via HIF-1α and HIF-2α stabilization, promoting hepatoblast differentiation toward hepatocytes (Fig. 3). In contrast, extensive hypoxia increases TGFβ signaling triggering cholangiocyte differentiation (Fig. 3). TGFβ is released by mesenchymal cells and endothelial cells and acts on the hepatoblasts indicating an essential need for a multilineage intercellular cross talk (5) (Fig. 3).
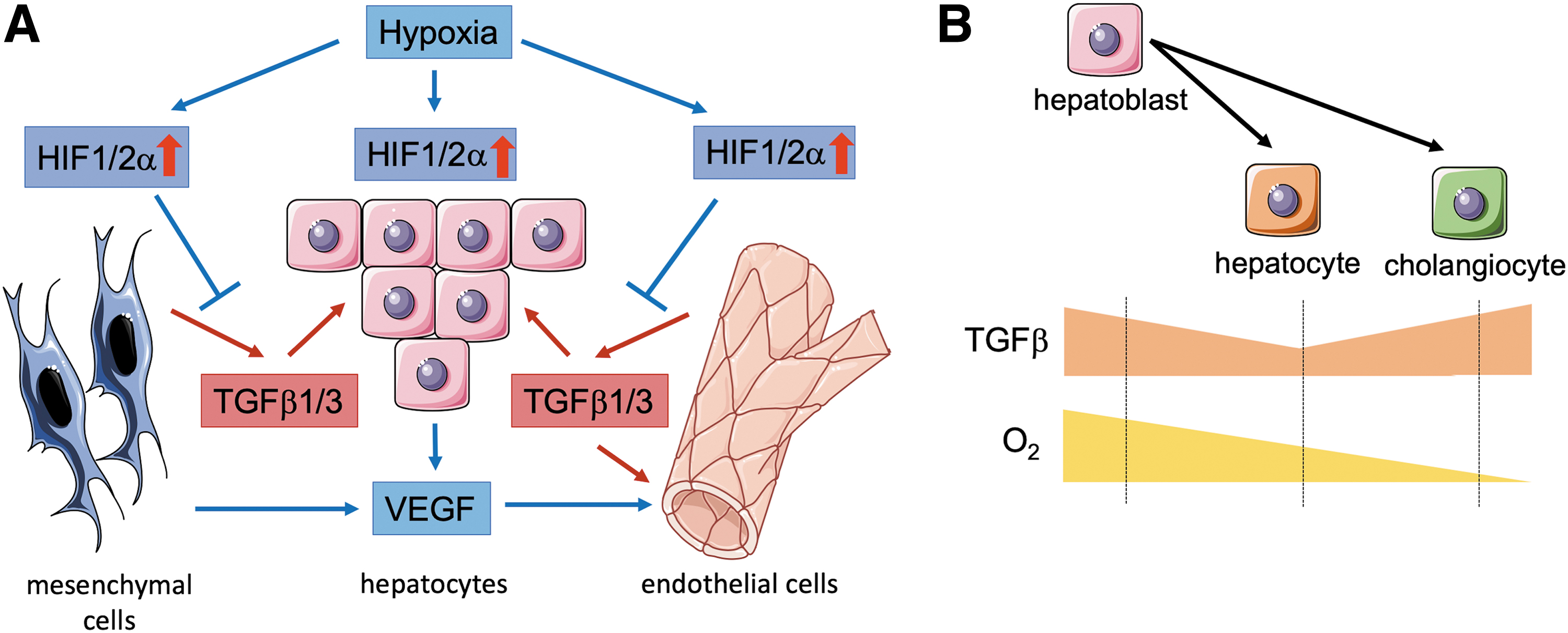
Complementary studies demonstrate that hepatocytes differentiated from iPSCs in two-dimensional culture clearly differ from hepatocytes differentiated in 3D organoid culture, which more closely resembles the in vivo situation (4, 17). The 3D organoid cultures contained hepatoblasts, MSCs, as well as endothelial cells (human umbilical vein endothelial cells). The authors show that hypoxia-driven activation of several signaling pathways (e.g., tumor necrosis factor [TNF], FGF, JAK/STAT, NF-κB, HIF, and VEGF signaling) as well as the production of extracellular matrix components and matrix remodeling enzymes (MMP2/4/19) in the organoid microenvironment enable multilineage communication, which is an essential driver of hepatocyte maturation (17).
A similar study confirms the beneficial effect of multicellular organoids on hepatocyte maturation and suggests that reduced Wnt and TGFβ signaling caused by the presence of the nonparenchymal cells is responsible for this effect (43).
These data well demonstrate that a complex cross talk between different cell types or tissue compartments is required to generate mature and functional organoids. During embryonic development, such inductive processes do also happen on a larger scale. A recent publication demonstrates that direct coculture of hind and foregut organoids enables the induction of the hepato-biliary-pancreatic anlage, an assembly of cells that arises during development at the hind and foregut boundary and can give rise to a set of different organs, namely the liver, the pancreas, and the biliary tract (62). This demonstrates that increasing organoid complexity by assembling different types of organoids might enable us to model the development of whole organ systems. A technique that has been used before to create neural circuits by coculturing different neural organoids (6, 11). Such cocultures have been referred to as assembloids (98).
Engineering Organoid Vascularization Results in Larger and More Mature Organoids, Reduced Cellular Stress Levels, and Long-Term Growth
As described above, organogenesis and tissue development are influenced by oxygen availability. The orchestration of early development often relies on physiological hypoxia. In contrast, finally established and expanding tissues react very sensitive to hypoxic conditions. Blood vessels are essentially required within growing tissues to allow rapid cellular proliferation and prevent cell death by sufficient supply with oxygen and nutrients.
The consequences of a lacking vascular system can be observed in cerebral organoids, which show large necrotic/apoptotic areas in their center as the organoid exceeds a certain size (>500 μm in diameter) limiting the diffusional supply of the cells with nutrients and oxygen (66). This does not only negatively impact size and long-term growth of organoids but also impairs cellular subtype specification and maturation as, for example, demonstrated in cortical organoids (10). If a sufficient supply of oxygen and nutrients is granted, for example, in cortical organoid slice cultures (105) or in vivo transplanted neural organoids (77), more mature cell types can be observed. Interestingly, a beneficial effect can be even detected in organoids containing a primitive vascular plexus, which is not connected to a circulatory system (16).
For that reason, strategies enabling the robust vascularization of organoids are currently of high interest and several studies regarding this issue have been recently published by us and others.
Takebe et al. demonstrated that self-condensation of adult or embryonic endothelial cells, mesenchymal cells, and organ-specific epithelial cell types results in the formation of vascularized organ buds, which can connect to a circulatory system following transplantation in the mouse (134), a protocol that could be also transferred to the human system (132). More recent studies demonstrate that multilayered human blood vessels can self-organize from human iPSC-derived early MPCs (146, 148) and can be grown in 3D mesenchymal spheres, the so-called blood vessel organoids, in vitro (146).
Different protocols were established to achieve cerebral organoid vascularization by either incorporation of iPSC-derived endothelial cells (102) (Fig. 4A) or mesodermal progenitors (148) (Figs. 4B and 5). An additional study made use of genetically altered iPSCs ectopically expressing human ETS variant 2 (ETV2), a transcription factor that plays an essential role during endothelial cell differentiation (128) and has been shown to drive the conversion of fibroblasts into endothelial cells (87). ETV2 iPSCs were mixed with unaltered iPSCs and cerebral organoids were induced. Due to the expressed transgene, ETV2 iPSCs differentiate toward the endothelial lineage, even under neurogenic conditions, which resulted in vascularized cortical organoids showing reduced hypoxic regions as determined by HIF-1a activity and significantly less cell death in core regions of the organoid (16) (Fig. 4C).
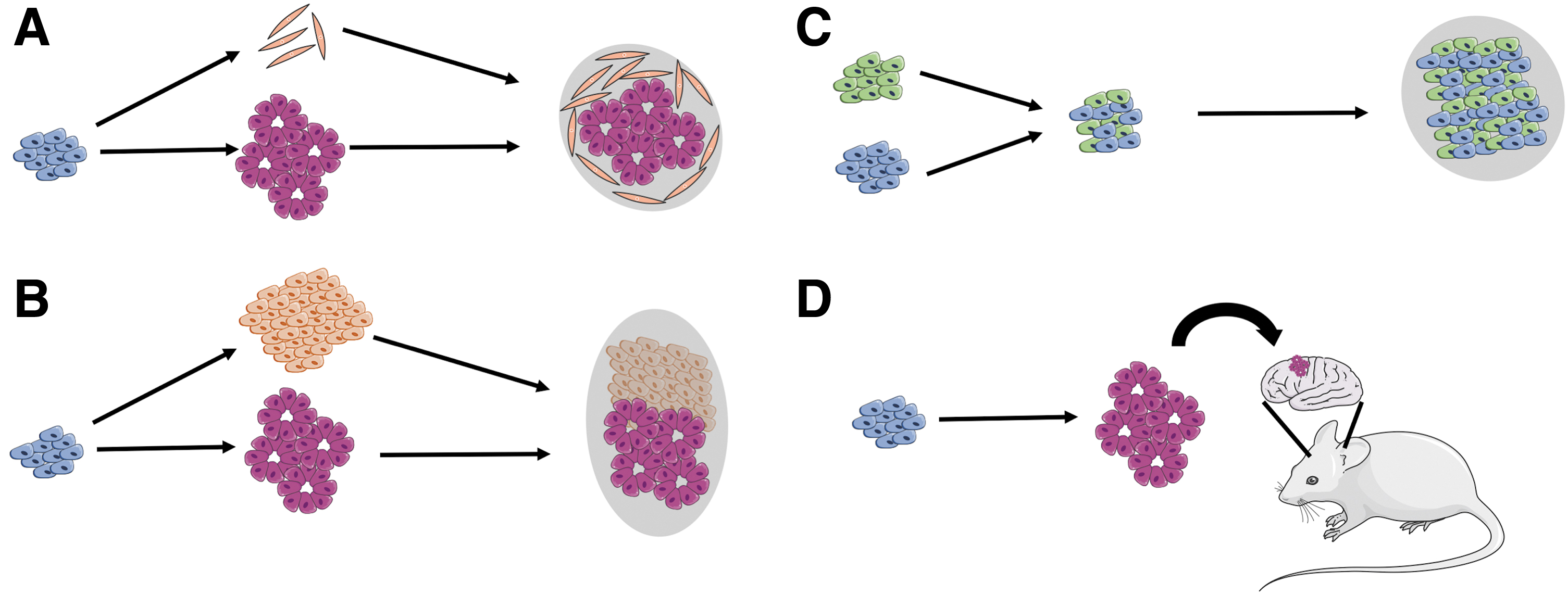
In this context, nanotransfection of reprogramming factors known to induce an endothelial cell fate such as ETV2, FOXC2, and FLI1 into organoids might represent an interesting strategy as well (40).
However, it needs to be mentioned that blood vessels are more complex than an endothelial tube as they consist of an intimal layer, a tunica media, as well as the adventitial connective tissue, which harbors a versatile stem cell niche (159). These stem and progenitor cells are important for vascular integrity and new vessel formation [for review, see Worsdorfer et al. (149)]. For that reason, we expect approaches utilizing mesodermal progenitors (146, 148) to be more suitable to achieve a bona fide vascular network than approaches using endothelial cells alone (16, 102).
In a different approach, cerebral organoids were transplanted into the brain of a mouse, which led to vascularization by the host and nutrition and oxygen supply of the grafted tissue via the mouse circulation (27, 77) (Fig. 4D). This strategy provides a functional vascular network and allows for the long-term culture (>10 months) of organoids within a living animal. Moreover, the functional vascular system significantly decreased cell death in the grafts.
However, the vessels (and also invading microglia cells) are mouse derived. For that reason, transplantation of prevascularized organoids generated by incorporation of human mesodermal progenitors (148) could provide a stably integrated and functionally connected all-human graft, which might be a good strategy for future studies.
An alternative approach that can also result in complex vascularized neural organoids is the use of primary human brain tissue.
A recent publication describing the establishment of patient-derived glioblastoma organoids (GBOs) well demonstrates that concept (54). The authors grow organoids from pieces of dissected brain/glioblastoma tissue. These tissue pieces can be long-term grown and expanded in cell culture and maintain the parental cellular heterogeneity. Interestingly, GBOs display a hypoxic gradient, resulting in different tissue niches with actively dividing tumor cells at the surface and quiescent tumor stem cells in deeper layers close to the hypoxic core of the organoid (52). Moreover, primary cell-derived organoids contain different tumor cell types, but in some cases, an endothelial cell network, microglia, and other brain cell types are found.
The main limitation of the method is that it requires access to human brain tissue. In the case of, for example, glioblastoma, this access is granted during surgical removal of the tumor. However, this method is not suitable for the investigation of most hereditary diseases or developmental processes. In those cases, iPSC-derived organoids are superior as iPSCs can be easily derived from urine (162), blood (126), or dermal fibroblasts (130) with minimal harm for the patient. Moreover, iPSCs can be further manipulated in cell culture, for example, by clustered regularly interspaced short palindromic repeats/CRISPR-associated 9 (CRISPR/Cas9) mediated gene editing.
The above-described technologies are not only suitable for neural organoids but could be applied to all organoid models, such as liver, gastrointestinal, or lung organoids. In contrast, kidney organoids derived from human iPSCs intrinsically develop a rudimentary blood vessel network. This is because such organoids are derived from mesodermal progenitors and during that process angioblasts are concurrently induced with nephron progenitor cells (133).
Interestingly, a culture under flow on a millifluidic chip expands the pool of endothelial progenitors and results in perfusable vascular networks surrounded by mural cells (50). This indicates that connecting prevascularized organoids to millifluidic chips (50) or microfluidic systems (120) could greatly enhance vascular network formation and maybe even more important maturation of the vessel wall initiated by flow-induced shear stress that is sensed by the cells of the endothelium and triggers multiple biological responses in endothelial cells as well as surrounding smooth muscle cells and pericytes.
Organoids as a Platform to Study the Detrimental Effects of Hypoxia
The data presented above show that physiological hypoxia is an important driver of several developmental processes. However, pathological hypoxia can cause severe damage during fetal development as well as in the adult body. Placental insufficiency during fetal development or lung immaturity in premature infants can cause such conditions. Other examples in the adult organism are, for example, chronic hypoxia in the ischemic limb or in the retina of diabetic patients.
As organoids represent a model for developing or immature tissues, they are currently best suited to study the effect of pathological hypoxia during fetal development. Organoids representing tissues known to be negatively affected by hypoxic conditions such as the kidney (123) or the brain (112) have been established within the past years.
Some recent studies investigate this issue focusing on the effect of pathological hypoxia on the developing human brain. An insufficient supply of brain tissue with oxygen can occur during embryonic development or in premature born infants due to lung immaturity. This leads to hypoxic episodes bearing a high risk of developing a critical condition termed encephalopathy of prematurity. So far, the susceptible cell types and the molecular mechanisms underlying this disease were not clear because access to human fetal brain tissue is difficult and animal models are inadequate to study human cortical development.
A recent study used cerebral organoids and exposed them to hypoxic conditions. The authors found that intermediate progenitors in the developing cerebral cortex are especially affected by hypoxia and that the effect was related to an unfolded protein response. Moreover, they identified a small-molecule inhibitor of unfolded protein response that partially prevented the detrimental effect (97). A similar study demonstrated that outer radial glia cells and differentiating neuroblasts/immature neurons were sensitive to hypoxia in a model of prenatal hypoxia injury, while ventricular zone stem cells remained unaffected (26). A third study cultured cerebral organoids under hypoxic conditions and revealed that hypoxia repressed gene markers for forebrain, oligodendrocytes, glial cells, and cortical layers, as well as genes important for the migration of cortical neurons. Moreover, they demonstrate that the neuroprotective drug minocycline mitigates the detrimental effect (13).
These studies show that organoids are an attractive platform to study the detrimental effects caused by pathological hypoxia and test pharmacological substances, which might mitigate tissue damage.
Summary and Conclusion
Hypoxia has the potential to modulate the activity of HIF, the epigenome, and the metabolic state of a cell. These processes influence stem and progenitor cell behavior by modulating proliferation and gene expression and therefore impact on cellular differentiation and stem cell quiescence. This is true in the developing embryo and also in the adult body, in which several stem cell niches exist, which enable the regeneration of various tissues either in a regular manner or after wounding and disease.
In cell culture, organ development, disease progression, and tissue regeneration can be studied in the so-called organoids, miniature versions of developing organs, grown from human stem cells. The influence of oxygen availability on the maturation of organoids is getting more and more into the focus of research. The published data suggest that a complex interplay between different cell types induces a cascade of signaling events finally resulting in fully matured tissues. Oxygen availability appears to be one of the factors triggering these events.
Therefore, a precisely regulated oxygen pressure during organoid culture might be beneficial to achieve proper tissue development and maturation. Moreover, a complex tissue context enabling the interplay between the stromal compartment (blood vessels and mesenchymal cells) and the parenchyma, the “functional” cells of the organ (e.g., lung or liver epithelial cells), is essentially required, which underlines the need for more complex organoid models including not only parenchymal but also stromal cells.
Besides helping to understand the beneficial effects of physiological hypoxia, organoids can also be used to shed light on the mechanistic processes causing the detrimental effects of pathological hypoxia leading to tissue damage and disease, especially during development. Moreover, a limited availability of oxygen and nutrients is the major hurdle restricting the growth of in vitro-generated microtissues such as organoids or tissue patches. In addition, effective transplantation of tissue patches requires a fast connection to the host circulatory system. For that reason, proper prevascularization appears to be mandatory to avoid cell death or rejection of the graft. Consequently, robust strategies for the integration of functional blood vessels into different kinds of organoids are currently of high interest.
More research is needed to (i) understand the impact of oxygen availability on organoid development, (ii) develop strategies for the generation of complex organoid models including the organ-specific parenchymal cells, a stromal compartment, blood vessels, tissue-resident immune cells, and probably also a peripheral nervous system (147), and (iii) understand the complex interplay between all these cell types and how this finally boosts tissue maturation.
Footnotes
Authors' Contributions
P.W. and S.E. conceptualized the content of this work. P.W. reviewed the literature and drafted the article and figures. P.W. and S.E. discussed and edited the review.
Acknowledgments
We thank all members of the Stem Cell Biology and Regenerative Medicine laboratory in Würzburg for useful comments and discussion. Figures 1–![]() are composed of graphical elements taken from the image bank from Servier Medical Art licensed under a Creative Commons Attribution 3.0 Unported License (
are composed of graphical elements taken from the image bank from Servier Medical Art licensed under a Creative Commons Attribution 3.0 Unported License (
Author Disclosure Statement
The authors declare no competing or personal financial interests.
Funding Information
This work was supported by the German Research Foundation (DFG, TR225-B04) and the IZKF-Würzburg (Interdisziplinäres Zentrum für Klinische Forschung der Universität Würzburg, E-D-410).