
Abstract
Significance:
Cardiovascular stem cells are important for regeneration and repair of damaged tissue.
Recent Advances:
Pluripotent stem cells have a unique metabolism, which is adopted for their energetic and biosynthetic demand as rapidly proliferating cells. Stem cell differentiation requires an exceptional metabolic flexibility allowing for metabolic remodeling between glycolysis and oxidative phosphorylation.
Critical Issues:
Respiration is associated with the generation of reactive oxygen species (ROS) by the mitochondrial respiratory chain. But also the membrane-bound protein nicotinamide adenine dinucleotide phosphate oxidase (NADPH oxidase, NOX) contributes to ROS levels. ROS not only play a significant role in stem cell differentiation and tissue renewal but also cause senescence and contribute to tissue aging.
Future Directions:
For utilization of stem cells in therapeutic approaches, a deep understanding of the molecular mechanisms how metabolism and the cellular redox state regulate stem cell differentiation is required. Modulating the redox state of stem cells using antioxidative agents may be suitable to enhance activity of endothelial progenitor cells. Antioxid. Redox Signal. 35, 163–181.
Introduction
Stem cells are undifferentiated cells with the capability of unlimited self-renewal and differentiation into specific cell types under adequate conditions (84, 130). They are divided into embryonic stem cells (ESCs) and adult stem cells (ASCs). ESCs are obtained by isolating and cultivating cells from the blastocyst, which forms ∼5 days after fertilization (30). A general feature of ESCs is their unlimited expansion and pluripotency in culture with the ability to differentiate in all cell types from the adult organism. In contrast, ASCs are quiescent cells with a more specified differentiation ability. There are well-defined characteristics for different types of ASCs, including epithelial, hematopoietic, muscular, and neural stem cells (4).
All stem cells are expected to live in so-called stem cell niches, which affect the behavior of the stem cells due to chemical signals and physical contact (39, 97). Mesenchymal stem cells (MSCs) are found in many different tissues, for example, bone marrow (107), adipose tissue (144), and peripheral blood (1). Due to their multipotency, MSCs are able to differentiate in various tissues and therefore contribute to their regeneration. Hematopoietic stem cells (HSCs) give rise to all cells from the blood cell lineage and reside in the bone marrow. Depending on their localization, they are either quiescent in the inner part of the bone, or the vascular niche supports proliferation, differentiation, and migration of the cells (64, 155). Furthermore, it was demonstrated that the vascular adventitia layer serves as a stem cell niche not only for vascular cells but also for nonvascular cells such as macrophages.
Vascular endothelial growth factor (VEGF), produced by these macrophages, has been shown to be important for differentiation of adventitia-derived stem cells (88). Cardiac myocytes are terminally differentiated shortly after birth; therefore, the heart is incapable of regenerating cardiac tissue after any damage (19, 80). Studies reporting that bone marrow-derived cells or cells from the heart itself were able to regenerate cardiac tissue after myocardial infarction (MI) lead to the hypothesis that the human heart contains cardiac stem cells, which are capable of generating smooth muscle cells (SMCs), endothelial cell lineages, and myocytes. However, subsequent studies failed to find any evidence for the regeneration of cardiomyocytes (78, 128).
A vigorous analysis of cardiac regeneration using lineage tracing in fact concluded that not any cell type is capable of producing new cardiomyocytes in the adult heart (76). In an alternative hypothesis, explaining the initial positive results after injecting or infusing progenitor cells is, that stem cells secrete paracrine factors that can mediate a reconditioning effect on the injured heart or trigger an immune response, which mediates a secondary healing process (33, 45, 100).
As will be discussed in further detail below, stem cell proliferation and differentiation is associated with substantial metabolic remodeling. Metabolism involves anabolic reactions allowing for the assembly of biomolecules and catabolic reactions in which complex molecules are degraded for energy conversion. Many different molecules provide energy for the cell, such as amino acids, ketone bodies, fatty acids, and glucose. During glycolysis, glucose is oxidized and broken down to pyruvate, which can subsequently be reduced to lactate, regenerating the reducing equivalents, which have been used in glycolysis (Fig. 1).

During aerobic metabolism, pyruvate is converted into acetyl-CoA (Coenzyme A), which becomes oxidized in the mitochondrial Krebs cycle (tricarboxylic acid cycle). In the Krebs cycle, acetyl-CoA is oxidized to CO2 and at the same time, the reduction equivalents NADH and FADH2 are generated. NADH and FADH2 provide electrons for the mitochondrial respiratory chain, which is located in the inner mitochondrial membrane and transfers electrons onto molecular oxygen (Fig. 1). Simultaneously, protons are transported to the intermembrane space (IMS) to establish a proton gradient. This electrochemical gradient drives the adenosine triphosphate (ATP)-synthase for ATP production.
The oxidation of glucose produces 38 molecules of ATP per glucose. Tightly associated with glycolysis is the pentose phosphate pathway (Fig. 1). Sharing the same first reaction with glycolysis, the intermediate glucose-6-phosphate is converted into ribulose-5-phosphate, which is important for nucleotide synthesis. In addition, the pentose phosphate pathway produces NADPH/H+, which is further used for the reduction of glutathione or synthesis of fatty acids. The glycolytic intermediate 3-phosphoglycerate can be fueled into the “one-carbon metabolism,” which is a cyclic metabolism pathway. The one-carbon metabolism supports several processes, such as purine biosynthesis, amino acid homeostasis, and redox defense.
Fatty acids represent a major energy resource as well. After uptake of fatty acids into the mitochondrial matrix via the acyl-carnitine shuttle, they undergo β-oxidation. With multiple reaction steps, the fatty acids are degraded to acetyl-CoA, generating FADH2 and NADH as electron donors. Subsequent oxidation of acetyl-CoA in the Krebs cycle produces even higher amounts of ATP than glucose (129 molecules ATP for palmitate). However, the ATP/O2 ratio is less favorable for fatty acid—compared with glucose—or ketone oxidation (10).
Metabolic Flexibility in Cell Fate Decisions of Pluripotent Stem Cells
Early mammalian development of pluripotent stem cells is associated with substantial changes in the metabolism. As energy stores of mammalian oocytes predominantly consist of amino acids and proteins, but only low levels of glycogen and fat, the first cells of the fertilized egg use an oxidative metabolism to gain energy (Table 1). In fact, high concentrations of glucose can inhibit early embryogenesis (11). The immature mitochondria, which are inherited from the oocyte, have a spherical morphology and only low numbers of cristae structures. Despite the preference for oxidation, oxygen consumption rates are relatively low. During blastocyst implantation, inherited mitochondria are progressively lost due to rapid cell divisions. As ATP is a potent inhibitor of glycolysis, decreasing ATP/ADP ratios release the break on the rate-limiting enzyme phosphofructokinase 1 (PFK1; Fig. 1) and induce a switch of cellular metabolism toward increased glycolysis (60).
Key Features of Stem Cell Metabolism
ATP, adenosine triphosphate; HIF-1α, hypoxia-inducible factor 1α; PDH, pyruvate dehydrogenase; PPAR peroxisome proliferator-activated receptor; ROS, reactive oxygen species.
The oocyte factors Tcl1 and Tcl1b1 have been recently found to mediate this switch to glycolysis by regulating mitochondrial biogenesis during reprogramming of human-induced pluripotent stem cells (iPSCs) (63). The octamer-binding transcription factor 4 (Oct4) target Tcl1 suppresses mitochondrial targeting of the mitochondrial polynucleotide phosphorylase, causing decreased mitochondrial biogenesis, and contributes to reduced oxidative phosphorylation (63). With the first steps of differentiation of the pluripotent epiblast cell layer in the blastocyst, glucose uptake is strongly increased due to the upregulation of glucose transporters. Upregulation of hexokinase and lactate dehydrogenase, and downregulation of pyruvate dehydrogenase activity, redirect substrate fluxes toward increased glucose metabolism and lactate production (Table 1). Glycolytic intermediates also serve as precursors for amino acid synthesis, lipid synthesis and nucleotide and nicotinamide adenine dinucleotide phosphate (NADPH) synthesis, allowing for rapid proliferation.
The avoidance of an oxidative metabolism and the upregulation of glycolysis are evidence for a more reducing environment of stem cells, which also avoids generation of reactive oxygen species (ROS). Interestingly, a switch from oxidative phosphorylation to glycolysis is observed during reprogramming of differentiated cells to iPSCs. One transcription factor that is required and upregulated during reprogramming is Oct4, which regulates several metabolic genes. Small molecule-mediated iPSC generation was shown to mediate the metabolic switch to glycolysis (161). The establishment of glycolytic metabolism occurs earlier than the expression of pluripotency markers (36). Promotion of glycolysis induces iPSC reprogramming, inhibition of glycolysis or stimulation of oxidative phosphorylation can inhibit reprogramming (112). In summary, the metabolism of stem cells fundamentally differs from differentiated cells by the predominant role of glycolysis in energy conversion.
Molecular Mechanisms of Stem Cell Fate Decisions
Upregulated glycolysis
Metabolic switching in pluripotent stem cells is often compared with the Warburg effect of highly proliferative cancer cells (143), in which glycolysis provides the largest proportion of energy demand. A clear difference between the glycolytic metabolism in cancer cells (Warburg effect) and the metabolism in proliferating stem cells is the refueling of cytosolic metabolites in stem cells. Glucose flux in proliferating cancer cells results in regeneration of the rate-limiting NAD+ by lactate production even under aerobic conditions. In stem cells, pyruvate seems to be required to form the Krebs cycle metabolite citrate, which is subsequently cleaved into acetyl-CoA units by the cytosolic ATP citrate lyase (94). Acetyl-CoA regulates histone acetylation, resulting in an open euchromatin state, which maintains the pluripotent epigenetic state and self-renewal of PSCs (7, 41). Similarly, increasing histone acetylation by inhibition of histone deacetylases promotes reprogramming of somatic cells into iPSCs (53).
The increased glycolytic flux may also have functional implications by changing the glycosylation pattern of key regulatory proteins. The increased glucose flux also enhances the metabolic flux through the pentose phosphate pathway, providing reducing equivalents (such as NADPH) and precursors for nucleotide biosynthesis, which are required for rapid proliferation. Similarly, the hexosamine biosynthetic pathway (HBP) is increased under conditions of high glucose flux. This HBP generates uridine diphosphate N-acetylglucosamine (UDP-GlcNAc), which plays a role in glycosylation and O-GlcNAcylation (14). Many biological processes, including the pluripotency of stem cells, are regulated by O-GlcNAcylation. The transcription factors Oct4 and sex-determining region Y-box 2 (SOX2) activate many pluripotency-related genes and are O-GlcNAcylated in stem cells (59). Blocking O-GlcNAcylation disrupts ESC self-renewal and reprogramming of somatic cells to iPSCs. Upon differentiation, the O-GlcNAc modification is rapidly removed.
Amino acid catabolism
Stem cells show a strong bias to utilize glutamine for energy metabolism. Glutamine is converted to α-ketoglutarate (α-KG), which fuels the Krebs cycle (Fig. 2). α-KG is a cofactor for epigenetic modifier enzymes, like the Jumonji domain containing histone demethylases (JmjC), which are regulating chromatin remodeling and manifesting pluripotency. Supplementation of α-KG to ESC culture promoted pluripotency, whereas the competitive inhibitor succinate induced differentiation (2). A characteristic change in the metabolism of amino acids in stem cells is also manifested in the prominent role of threonine metabolism (121). Upregulation of the threonine dehydrogenase induces the conversion of threonine to glycine and acetyl-CoA. Acetyl-CoA serves as an anaplerotic substrate to replenish Krebs cycle intermediates. Similarly, the catabolism of the amino acids valine, leucine, isoleucine, and lysine is upregulated in undifferentiated stem cells, forming acetyl-CoA and numerous intermediates of the Krebs cycle, including oxaloacetate, α-KG, fumarate, and succinyl-CoA.
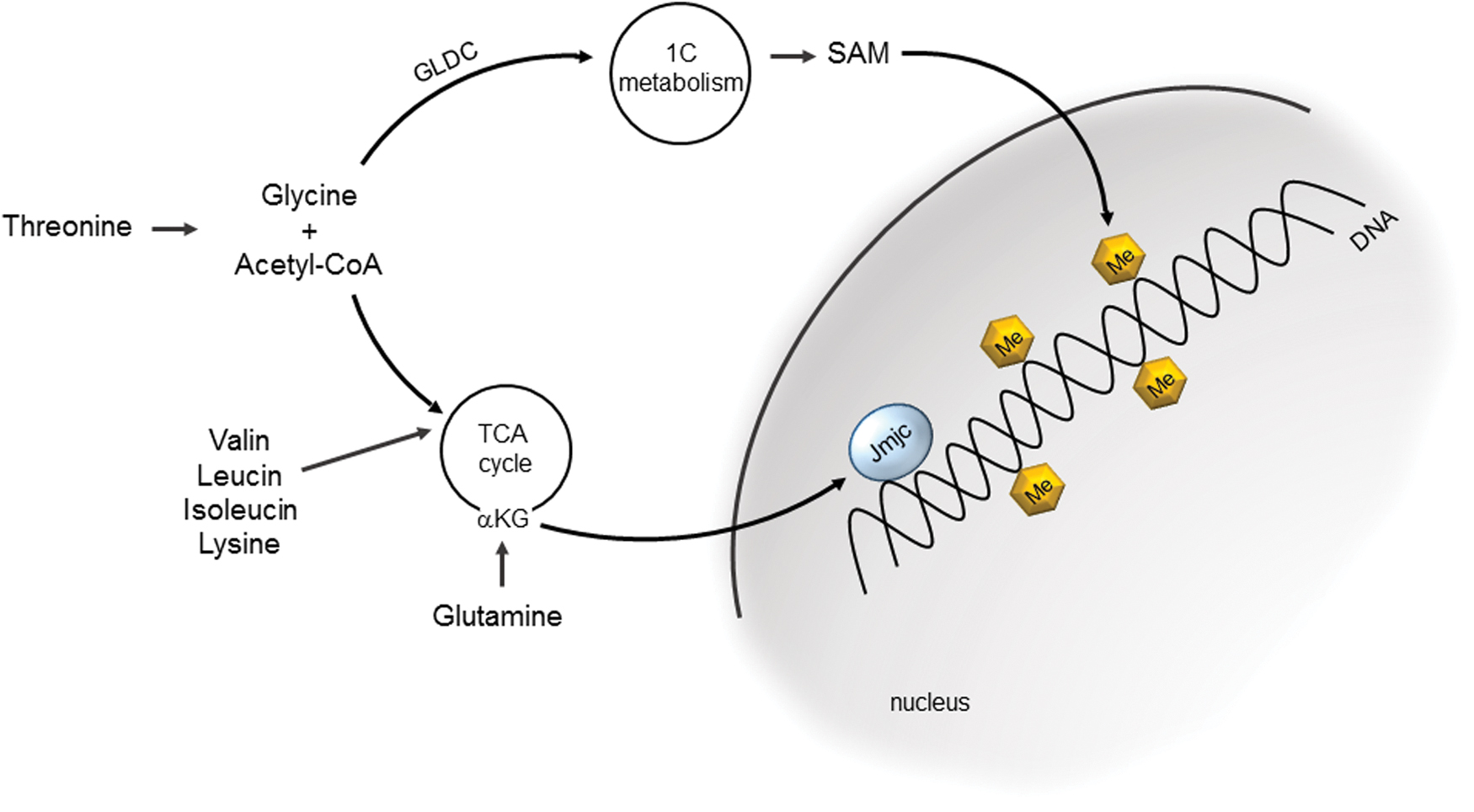
Glycine, the second metabolite in the breakdown of threonine, is decarboxylated by the mitochondrial enzyme glycine decarboxylase to form 5,10-methylene-tetrahydrofolate, which is an intermediate in the one-carbon metabolism (160). One-carbon metabolites are important players in epigenetic modifications of chromatin, as they induce methylation of histones (Fig. 2), which is correlated with a chromatin structure required for the regulation of the self-renewal state of PSCs. Active one-carbon metabolism is typical for undifferentiated ESCs, and genes of the folic acid cycle such as MTHFD2, FOLR1, and PIPOX show high expression levels in ESCs.
Lipid metabolism
Activation of stem cells to enter a differentiation program is often associated with a shift from glycolysis to oxidative phosphorylation. The metabolic shift includes a decrease in glycolytic flux, normalization of threonine metabolism, and an increase in fatty acid oxidation (FAO). The metabolic shift induces a more oxidative state in differentiating stem cells, which becomes an important mediator of the differentiation process. The stem cell metabolome is characterized by a distinct prevalence of unsaturated metabolites. Unsaturated metabolites are known to be more susceptible to oxidation (153). One interesting finding in stem cells is a strong enrichment of unsaturated species of long-chain fatty acids such as arachidonic acid (C20:4), eicosapentaenoic acid (C22:5), and docosahexaenoic acid (C22:6). These fatty acids are well-studied precursors for lipid mediators, such as prostaglandins and thromboxanes. These signaling molecules are released from membrane lipids, such as phosphatidylserine, by Ca2+-dependent phospholipases A2-mediated cleavage and are then subsequently oxidized by cyclo-oxygenases, enzymes of the Cytochrome P450 family, or undergo spontaneous oxidation by singlet oxygen.
Lipid mediators are important signaling molecules. For example, prostaglandin E2, an eicosanoid pathway product, has been demonstrated to promote HSC proliferation by promoting Wnt signaling (44). This suggests that eicosanoid metabolism might be critical in regulating HSC proliferation and differentiation. Free radical oxidation of linoleic acid (C18:2) produces 9- and 13-hydroxyoctadecadienoic acid (9- and 13-HODE), respectively, and their derivatives. These lipid mediators are known to activate the transcription factors peroxisome proliferator-activated receptor (PPAR) β and γ. PPARs regulate transcription of a large panel of genes, involved in lipid and glucose metabolism, inflammation, cell proliferation and differentiation.
9-HODE and 13-HODE are also important for activating TRPV1, a Ca2+ permeable channel in the plasma membrane, which plays a well-studied role in the differentiation of mESCs into cardiomyocytes (104). Moreover, oxidized forms of eicosanoids, such as 11,12-epoxyeicosatienoic acid, play a role in the engraftment of HSCs (70). In the concept of “chemical plasticity,” the high degree of unsaturated long-chain fatty acids in stem cells makes them susceptible to oxygenation, which allows for the production of a highly active pool of lipid mediators. Support for this concept came from a study revealing a transient change in the reduced glutathione (GSH)/oxidized glutathione (GSSG) ratios during differentiation, compensated by an increase in ascorbic acid levels. The change from reductive to oxidative environment might serve as a trigger for the signaling of lipid mediators. Accordingly, preventing the generation of polyunsaturated fatty acids delays differentiation (153).
FAO was recently found to play an important role in the self-renewal of long-term HSCs. The HSC population is maintained by asymmetrical cell divisions, in which the daughter cells either undergo self-renewal or differentiation. Asymmetric stem cell divisions, where one daughter cell differentiates and the other self-renews, preserve the stem cell population and replenish the mature cell pools. Inhibition of FAO enhanced symmetric, differentiating cell division, whereas pharmacologic activation of FAO increased asymmetric division. These data led to the conclusion that FAO is critical for stem cells to divide and have at least one daughter cell with self-renewal capacity.
Mitochondria act as central metabolic hubs in stem cell differentiation
Mitochondria are surrounded by two membranes, the outer membrane (OM) and the inner membrane (IM), which are separated by the IMS. Invaginations of the IM into the matrix compartment (cristae structures) form a functionally distinct compartment containing the respiratory chain (Fig. 3). The IM is composed of a unique lipid composition with a high concentration of the phospholipid cardiolipin (CL), a unique lipid only found in mitochondrial membranes (101a). The CL molecule adopts a cone-shaped structure and is anticipated to support membrane curvature by localizing in the highly bended regions within the cristae structures. A large protein structure, the mitochondrial contact site, and cristae organizing system (MICOS) locate at the cristae junctions and mediate the formation of cristae structures, thereby contributing to the complex mitochondrial morphology (5, 23, 105, 141).

In the majority of differentiated cells, mitochondria form a large tubular network. The reticular morphology is maintained by two opposing processes: fusion and fission. Fusion is the process of merging of two mitochondria and is regulated by a set of dynamin-related GTPases: the mitofusins (Mfn1, Mfn2) in the OM and OPA1 in the IM (Fig. 3) (24, 118, 149). Their activity is counteracted by the segregation of two mitochondria (fission), mediated by DRP1, which also belongs to the family of dynamin-related GTPases. Fission and fusion maintain the homogeneity of mitochondria within a cell (119).
Distinct mitochondrial morphology has been found in stem cells and differentiated cells. Mitochondria in stem cells have a globular shape, poorly formed cristae structures, and are commonly found in a predominantly perinuclear localization (108). During differentiation of ESCs to cardiomyocytes mitochondria elongate, which is also accompanied by an increased expression of Mfn2 and Opa1. Cardiac-specific knockout of the profusion genes MFN1 and MFN2 causes mitochondrial fragmentation and redistribution from the perinuclear localization to the cell membrane. The ability of mitochondria to take up Ca2+ at morphological distinct sites greatly impacts cellular Ca2+ fluxes. As Ca2+ uptake takes place at sites where the OM is in proximity to the endoplasmic reticulum (ER) membrane, the regulated Ca2+ flux critically depends on mitochondrial morphology and the association with the ER membrane. Consequently, changes in the mitochondrial morphology disturb Ca2+ uptake in mitochondria. In cells with an ablation of MFN2 and OPA1, increases in cytosolic Ca2+ levels have been documented. Cytosolic Ca2+ activates calcineurin, which increases Notch signaling, promoting cellular stemness and decreased expression of the differentiation factors GATA4, MEF2C, and Nkx2.5 (62).
A role for mitochondrial morphology also has been observed during the process of iPSC reprogramming. The profission factor DRP1 mediates early fragmentation of mitochondria, which is important for the ability of stem cells to migrate. Fragmentation of the mitochondrial network allows cells to reposition mitochondria, with fragmented mitochondria positioned at the front edge of the migrating cell and long mitochondria at the opposite end of the cell. In vascular SMCs, it was observed that initiation of migration activates DRP1-mediated mitochondrial fission. In this cell type, mitochondrial fragmentation served to increase mitochondrial ATP production at the site of high energy demand. Interestingly, mitochondrial fission also induced emission of ROS, which is also required for cell migration (147).
Mitochondria are the central metabolic hubs in the cell. The reducing equivalents NADH and FADH2, which are generated during glycolysis and in the Krebs cycle, are oxidized by the respiratory chain in the mitochondrial IM. The respiratory chain consists of five complexes (I–V), which mediate electron transport from reducing equivalents to molecular oxygen. Coupling this electron transport with proton export across the IM generates the membrane potential, which serves as an energy source for the production of ATP by the F1Fo-ATPase (complex V). The membrane potential in stem cells is significantly lower compared with differentiated cells. Expression and incorporation of the uncoupling protein 2 into the mitochondrial membrane cause a short circuit of the membrane potential, and limit mitochondrial ATP synthesis in stem cells.
Respiratory chain complexes assemble into large supercomplexes, formed by complex I, a dimer of complex III and several copies of complex IV (72a, 149a). Supercomplex formation allows efficient electron transfer between these complexes and regulates generation of ROS by the respiratory chain (117a). The mitochondria-specific phospholipid CL is required for the structural integrity of supercomplexes (8a, 28a, 52a, 91a, 105a, 159a). During cardiac development, the embryonic metabolism switches to the more efficient oxidative metabolism in cardiac myocytes. Transcriptome analyses indeed showed reprogramming of the transcriptome encoding components not only of glycolysis, FAO, and the Krebs cycle but also of the electron transport chain. Interfering with respiratory chain function using specific inhibitors causes deficient assembly of sarcomeres, dysfunction in contraction, and prevented differentiation of stem cells into functional cardiomyocytes (22, 127). Studies in HSCs showed that changing mitochondrial membrane lipid composition by a defect in the lipid biosynthesis enzyme PTPMT1 caused a severe impact on mitochondrial energy metabolism and led to a large expansion of the HSC pool but prevented differentiation into downstream lineages (156, 159). These data clearly indicate the role of mitochondrial energy metabolism in the differentiation of stem cells.
The Role of Oxygen Levels and ROS in Stem Cell Differentiation
Oxygen level and stem cell quiescence
Early embryogenesis is adapted to a low-oxygen environment of <5% and low levels of ROS. Fetal cells, including cardiomyocytes, are highly dependent on glycolysis for energy production. Experiments in human ESCs (hESCs) have shown that the optimum oxygen concentration for mammalian embryonic development is low. hESCs cultured at oxygen concentration of 1%–4% preserve their proliferative ability and form embryoid bodies (35). At higher oxygen concentrations, hESCs began to differentiate spontaneously (157). Proliferative progenitor cells use a unique combination of glycolysis and oxidative phosphorylation for energy conversion. The availability of lactate also induces lactate oxidation for energy production. In the in vitro differentiation of iPSC or ESC into cardiomyocytes, the preference of cardiac progenitor cells for lactate is used in the selection step enriching the amount of cardiomyocytes and the elimination of noncardiomyocytes.
Similar to the fetal, also the neonatal heart uses glucose and lactate for ATP generation immediately after birth. Glycolysis rapidly decreases a few days after birth and provides <10% of total ATP production similar to the adult heart. The transition is induced by an increase in workload and growth, which causes energy demands that cannot be met by glucose and lactate consumption alone. Dietary changes, induced by maternal milk, increase the availability for free fatty acids and elevations in O2 and ROS levels, mediating a switch from glycolysis dependence to oxidative metabolism in mature cardiomyocytes. The metabolic switch to oxidative metabolism is associated with the loss of the regenerative phenotype.
Neonatal mouse hearts can regenerate in the first postnatal week, but this ability is lost after day 7. Studies exposing neonatal mice to hyperoxia (100% oxygen) found that hyperoxia induced cardiomyocyte cell cycle arrest after birth and further reduced the regenerative potential, whereas mild hypoxia (15% oxygen) prolonged the regenerative window (110). Administration of antioxidant agents resulted in a significant increase in mitotic cells after birth, indicating that increased levels of ROS under normoxic conditions induced cell cycle arrest and the terminal differentiation program (13). The adult heart is metabolically flexible and is capable of oxidizing many energy sources, such as lactate, which is produced by skeletal muscles during extensive exercise conditions, or ketone bodies. FAO predominates over other nutrients during physiological conditions (10).
The well-studied HSCs niche resides in the endosteal zone of bone marrow. Here, oxygen tension is reduced to values as low as 1% due to the limited blood perfusion (93). Under these hypoxic conditions, HSCs show a slow proliferation rate, which protects them from oxidative stress and increases their self-renewal activity. The transcription factor hypoxia-inducible factor 1α (HIF-1α) is an important mediator of metabolism under low oxygen concentrations. An oxygen-dependent protein modification targets the HIF-1α protein to the ubiquitin–proteasome system (Fig. 4). Therefore, HIF-1α is unstable and rapidly degraded under normoxic conditions.
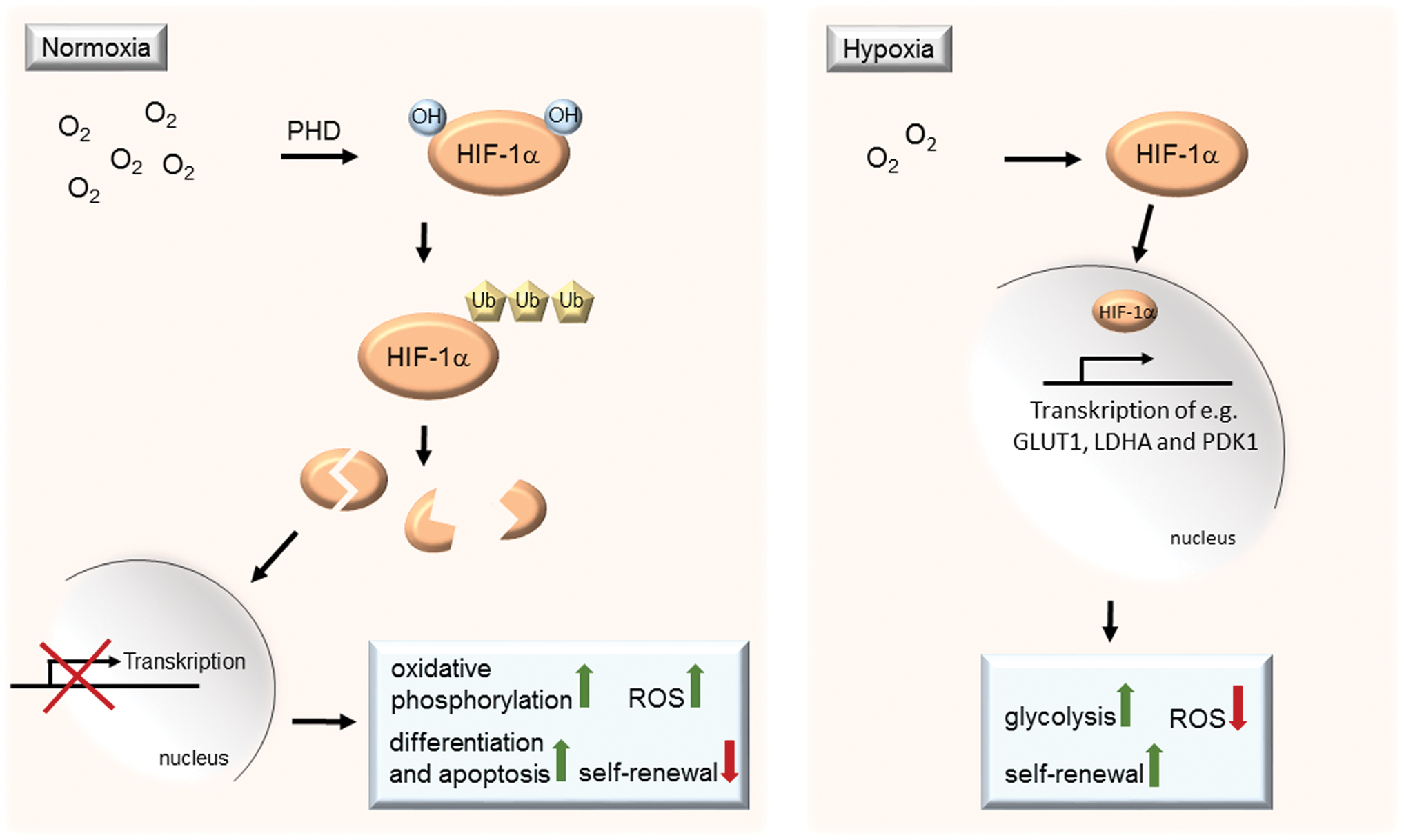
In hypoxia, reduced protein modification allows HIF-1α to escape from its degradation (47). Stabilized HIF-1α acts as a transcription factor changing gene expression of a large number of metabolic enzymes. HIF-1α mediates the expression of transporters and enzymes, including GLUT1, lactate dehydrogenase α, and PDK1, which contributes to the uptake of glucose and the reduction of pyruvate to lactate. To a large part, metabolic regulation of HIF-1α occurs via the transcription factor MEIS1 (123). The glycolytic metabolism also prevents generation of ROS. HSCs are sensitive to ROS, and ROS induce differentiation or apoptosis. Interestingly, HIFs can also promote the expression of stemness factors, such as nanog homeobox (NANOG), SOX2, and Oct4 in stem cells including hESCs (129). Block of HIF-1α signaling by genetic ablation of HIF-1α induces oxidative phosphorylation and an increase in ROS, resulting in the subsequent loss of quiescence and the self-renewal properties of HSCs (158). Conversely, increasing HIF-1α activation enhances glycolysis, inhibits oxidative phosphorylation, and improves proliferation and maintenance of ESCs, while it is repressing differentiation (83).
Cellular sources of ROS
The enzymatic mechanism of the mitochondrial respiratory chain ensures the full reduction of molecular oxygen, avoiding the generation of toxic ROS. The complete reduction by the transfer of four electrons turns molecular oxygen into water (H2O). Incomplete reduction by the transfer of less than four electrons converts oxygen (O2) into ROS. With the transfer of one electron at a time, oxygen is converted into superoxide anions (O2 •−), hydrogen peroxide (H2O2), and hydroxyl radicals (HO−) plus hydroxyl anion, respectively (Fig. 5). The main source of ROS in the cell is the mitochondrial respiratory chain. Despite the efficient electron transduction within the respiratory chain complexes, 0.2% of oxygen is converted to ROS. Complexes I and III most prominently contribute to ROS generation (28). ROS generated at complex I are emitted exclusively into the mitochondrial matrix, whereas ROS generated at complex III emit into the matrix and into the IMS, and are particularly relevant for cellular damage and cell signaling processes in the cytosol.
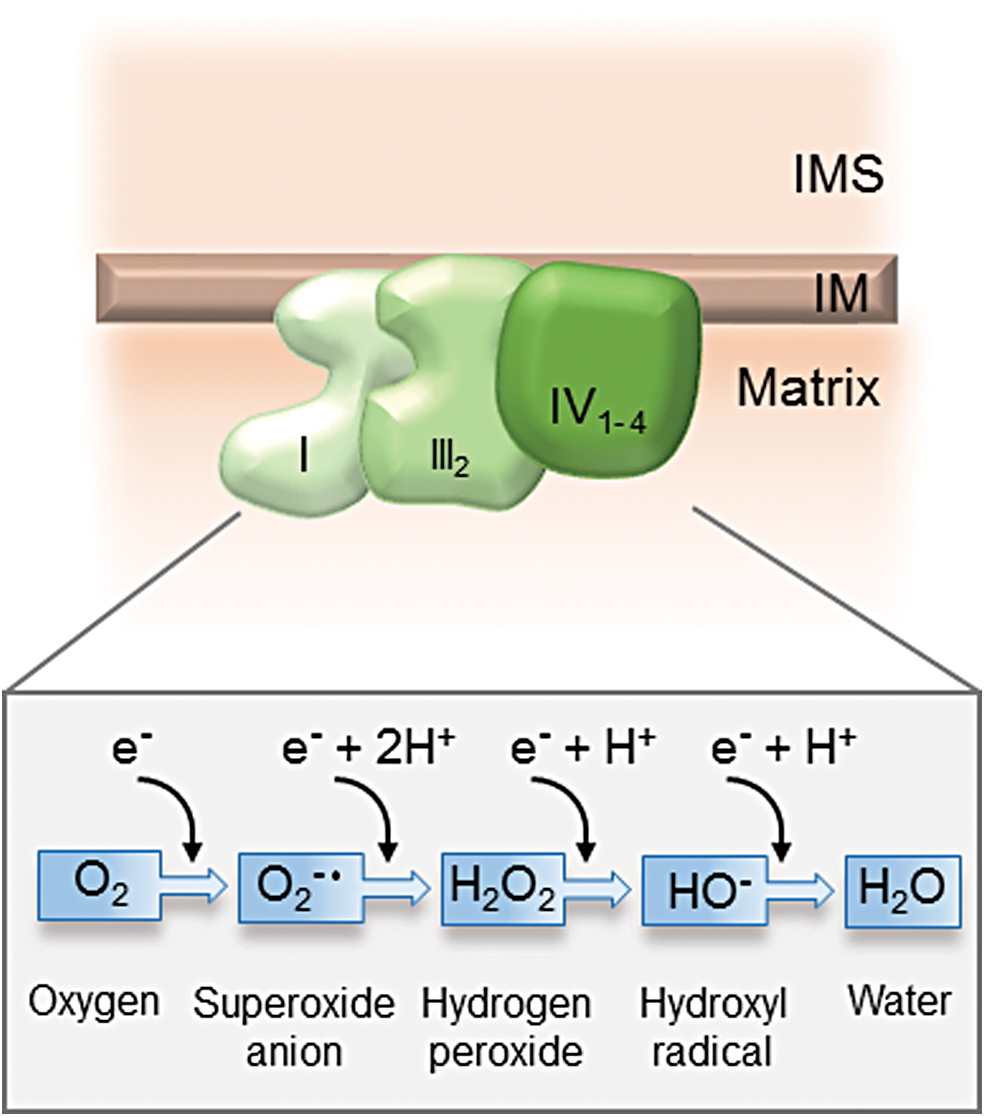
Another source of O2 − in undifferentiated human MSCs is the succinate dehydrogenase (complex II) (18). This enzymatic complex catalyzes the oxidation of succinate to fumarate resulting in the conversion of flavin adenine dinucleotide (FAD) to its reduced form (FADH2), which promotes superoxide production at the FAD-binding site of the complex.
ROS are highly reactive and can cause damage to cellular components, including proteins, lipids, and nucleotides. Under normal physiological conditions, the generation of ROS is tightly regulated by a ROS scavenging system. ROS scavengers are antioxidant enzymes that can neutralize ROS by directly reacting with and accepting electrons from ROS. These include superoxide dismutase (SOD), catalase, peroxiredoxins, thioredoxin (TRX), glutathione peroxidase (GPX), and glutathione reductase (GR) (114). The SOD catalyzes a rapid reduction of superoxide to H2O2, which is further reduced to H2O by catalase or GPX. Oxidized proteins are reduced by glutathione through the glutaredoxin and TRX systems. TRX is a small protein that is regenerated in an NADPH-dependent manner by the TRX reductase. Glutaredoxin transfers electrons onto GSH, which is regenerated in an NADPH-dependent manner by the GR.
Another major source of ROS is the membrane-bound protein nicotinamide adenine dinucleotide phosphate oxidase (NADPH oxidase, NOX, Fig. 6). NOX is a membrane-bound enzyme complex that uses cytosolic NADPH to generate primarily superoxide (O2 •−), which is converted into H2O2 by the SOD (125). Mammals express seven isoforms of NOX enzymes (NOX1, NOX2, NOX3, NOX4 and NOX5, DUOX1 and DUOX2) located predominantly at the plasma membrane but also at ER (NOX2, NOX4, and NOX5), the mitochondrial membrane (NOX4), and the nuclear membrane (NOX4 and NOX5) (125). NOX isoforms in the vascular system include NOX1, NOX2, and NOX4 in MSC-like stem cells (72), which can also be found in endothelial cells (124), vascular SMCs (109), and adventitial fibroblasts (50).
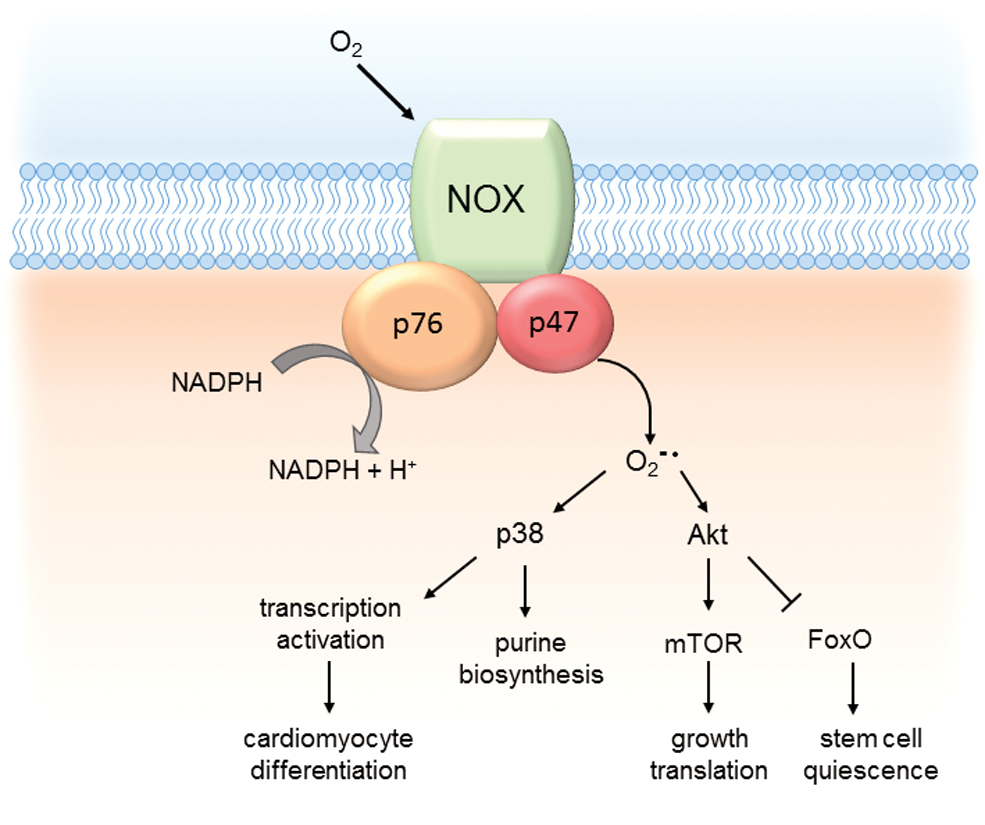
NOX are integrated into the membrane with six transmembrane domains, which also contain conserved binding sites for FAD and NADPH and a heme-binding domain. These cofactors are involved in the transfer of an electron from cytosolic NADPH to molecular oxygen producing superoxide anions as a primary ROS product (9). The NOX proteins form hetero-oligomeric complexes with the transmembrane protein p22phox and recruit various cytosolic proteins to regulate the individual activity. ROS generated by NOX play a particular role in stem cell differentiation and immune response. Although initially considered as harmful byproducts of metabolism, accumulating evidence indicate that some ROS, in particular H2O2, also play a role in cell signaling processes.
ROS and stem cell differentiation
Pericytes are embedded in the basement membrane in proximity to endothelial cells forming the blood capillaries. As a response to the ischemic microenvironment, pericytes exhibit predominantly glycolytic metabolism (16). Mitochondrial respiration has a low contribution to ATP synthesis, indicated by a limited sensitivity of these cells to oligomycin (66, 136). The low levels of oxidative metabolism also limit the generation of ROS. High expression levels of SOD and low expression of NOX maintain ROS levels low. Interference with this protective upregulation of ROS defense mechanism may have fatal consequences for pericyte function. Silencing of SOD3 interferes with the ability of adventitial pericytes to restore blood flow upon transplantation (54).
The high susceptibility of pericytes to ROS may also provide the pathological mechanism for loss of retinal vascular cell function under conditions of high glucose. Under physiological glucose levels, pericytes are spared from ROS generation, compared with other cell types, such as endothelial cells. Under high glucose however, pericytes become more dependent on mitochondrial respiration for energy conversion. Associated with this switch is an increased generation of ROS, providing a mechanism for the stress sensitivity of pericytes under high glucose conditions (42).
The switch to oxidative metabolism during differentiation of stem cells into mature cells also establishes a more oxidative environment associated with increasing levels of ROS (102). A single electrical field pulse induced cardiomyocyte differentiation of ESCs by enforcing ROS accumulation (117). Lowering ROS levels, using radical scavengers, inhibited stem cell differentiation in many studies, indicating that ROS are a prerequisite for stem cell differentiation (117). In neuronal stem cells, the increase of mitochondrial ROS by administration of specific inhibitors for complex I and complex III alleviated differentiation of neuronal stem cells. Knockdown of NOX1 and NOX4 in MSCs attenuated IL-17 dependent proliferation of hMSCs (52). Studies on the reverse pathway, the generation of iPSCs from differentiated cells, showed that limiting ROS increased the dedifferentiation process. Similarly, the efficiency of reprogramming increased by maintaining cells at low oxygen levels, which suppresses mitochondrial ROS emission, shifting metabolism toward glycolysis (35). These data indicate that ROS play an important role in shifting metabolism during differentiation.
The forkhead box (FOX) proteins are a large family of transcription factors of which the FOXO subfamily plays an important role in regulating stem cell homeostasis, self-renewal, and pluripotency (135). FOXO proteins are regulated by post-translational modifications such as phosphorylation, acetylation, and ubiquitination (140). Post-translational modifications can have a different impact on FOXO activity, since Akt-mediated phosphorylation inactivates FOXO family members, whereas c-Jun-N terminal kinase (JNK) signaling results in activation (34). Sirt1-mediated deacetylation of FOXO1 regulates subnuclear localization and fine-tunes the selection of transcriptional programs (37). FOXO plays a role in inducing cell cycle inhibitors, such as p27, p21, Cyclin G2, Cyclin B, and Polo like kinase and repressing cell cycle activators such as Cyclin D (86). Knockout of FOXO in HSCs causes a decrease in HSC numbers due to stem cell exhaustion (135). FOXOs function is to keep cells in quiescence and prevent exit into cell cycle, and therefore it prevents depletion of the stem cell pool. At the same time, FOXO induces antioxidant genes, such as SOD2 and CAT, indicating that to a large part, maintaining stem cell self-renewal is due to FOXOO's ability to limit oxidative stress. Accordingly, FOXOO-deficient HSCs show increased levels of ROS (135). The hematopoietic defect in the FOXO-deficient mice can be reverted with the antioxidant N-acetylcysteine (NAC), which rescues the cell cycle and apoptosis defects and restores the HSC pool (135). The HSC defects in FOXO-deficient mice are possibly caused by ROS-induced activation of the p53 tumor suppressor or the activation of p38 mitogen-activated protein kinase (MAPK) (92, 151). Transition of HSCs to myeloid progenitors is associated with a strong increase in ROS and results in terminally differentiated myeloid lineage cells with a robust emission of ROS upon their activation (135). The activity of FOXO inducing ROS defense mechanisms decreases during terminal differentiation of myeloid cells (135). Akt signaling, activated by exogenous cytokines, inactivates FOXO by direct phosphorylation. Akt-mediated FOXO phosphorylation induces its translocation into the cytoplasm, where it is ubiquitinated and targeted to the proteasome for degradation (12). In this model, Akt signaling is the initiating trigger eliminating FOXO activity to maintain stem cell quiescence. Therefore, it becomes increasingly clear that ROS act as signaling molecules directing the differentiation of HSCs to myeloid progenitor cells.
The role of ROS as second messengers in stem cell differentiation has also been observed during the differentiation of ESCs into cardiomyocytes. Administration of H2O2 induced differentiation and increased the expression of NOX1, NOX2, and NOX4. Specific NOX inhibitors prevented differentiation into cardiomyocytes (74). These studies indicate that NOX-mediated ROS signaling triggers redox-regulated molecular switches, inducing the differentiation process (Fig. 6). One of the best studied molecular mechanisms for ROS-mediated signaling is the regulation of protein phosphatases. Substrate phosphorylation by kinases is often balanced by opposing actions of protein phosphatase reactions, including protein tyrosine phosphatases (PTPs). These enzymes are sensitive to the redox state due to critical cysteine residues. The oxidation of these cysteine residues inhibits phosphatase activity and promotes kinase signaling.
The p38 MAPK pathway is a well-known target of redox regulation. H2O2 administration to glia cells reduces protein phosphatase activity, and increases phosphorylation and activation of the p38 MAPK protein kinase (115). In HSC, ROS-induced p38 MAPK activated the gene expression of enzymes involved in purine biosynthesis, which is an important mechanism for stress-activated hematopoiesis (61). Upon hematopoietic stress induced by serial bone marrow transplantation, p38 MAPK becomes activated and induces genes of the purine biosynthesis pathway, required for proliferation and bone marrow repopulation. NOX4-derived ROS were a critical component of the differentiation of mouse ESCs into cardiomyocytes. They activate p38 MAPK, which results in nuclear translocation of the cardiac transcription factor myocyte enhancer factor 2C (MEF2C) (74). MEF2C, Nkx2.5, and GATA4 are central transcription factors expressed in cardiac cells, and they play an important role in transcriptional regulation of cardiac specific genes (29). Phosphorylation by p38 MAPK induces their transcriptional activity and promotes differentiation to cardiomyocytes.
A role of ROS in regulating cell signaling pathways has been demonstrated also for the Akt-mTOR (mechanistic target of rapamycin) pathway during stem cell differentiation. Early studies using rapamycin-mediated mTOR inhibition revealed a role for mTOR in the differentiation of adipocytes (154a). Later work demonstrated important roles of mTOR in promoting the differentiation of ASCs, driving the growth and proliferation of stem and progenitor cells (91, 40a, 94a). Akt is important for the precise regulation of mTOR signaling in stem cell self-renewal and differentiation. It is activated by phosphatidyl-inositol-(1,3,4)-phosphate (PIP3) produced by PI3K. The phosphatase and tensin homolog (PTEN) is an important negative regulator of Akt by dephosphorylating PIP3 to PIP2 (137). PTEN belongs to the family of PTPs and is also redox regulated by the oxidation of cysteine residues (133). Oxidative stress inhibits PTEN, and thereby fosters Akt-mediated activation of mTOR signaling.
APE-1 is a redox-sensitive transcriptional modulator, which was originally discovered as an endonuclease in the base excision repair pathway of DNA lesions. Independent of this, it also mediates a redox switch in transcription factors, which can enhance, repress, or direct stem cell differentiation toward specific cell types. Oxidative stress promotes the nuclear translocation of APE-1, regulating the binding activity of several transcription factors such as NF-κB, p53, and HIF-1α (113). APE-1 reduces the oxidized cysteine residues within the DNA-binding domain of transcription factors. Maintaining these cysteine residues in a reduced state is critical for transcription factor activity. This activity is modulated by phosphorylation of APE-1 by casein kinase or protein kinase C, which switches APE-1s function from DNA repair to redox regulation of transcription factors (38, 150). APE-1 nuclear export to the cytosol depends on nitrosylation of cysteine residues, which triggers conformational changes, exposes nuclear export signals, and facilitates nuclear export of APE-1.
Recently, it was observed that APE-1 is targeted to mitochondria in a redox-specific manner (75). The mitochondrial intermembrane space import and assembly (MIA) pathway is a protein import machinery that uses a redox switch to drive cysteine-rich proteins into the IMS (17). The central component of this pathway, Mia40, interacts with APE-1 by forming a disulfide bridge between the cysteine residues on APE-1 and cysteine residues on MIA40 (8). Interestingly, MIA-driven import is strictly dependent on the redox state of the cell, with balanced glutaredoxin levels, required for oxidative folding in the reducing environment of the IMS (69). The function of APE-1 in the mitochondria is not resolved; however, overexpression of APE-1 alleviates mitochondrial dysfunction upon ischemia/reperfusion in cardiomyocytes.
Stem cells in the endothelia of blood vessels exist in three distinct layers: the tunica intima, an inner lining of endothelial cells, which contains the endothelial progenitor cells (EPCs), the tunica media, a thick middle layer composed of SMCs and stem cells, and the tunica adventitia (32). The tunica adventitia forms an outer layer of connective tissue, containing a heterogeneous population of cells, including fibroblasts, resident inflammatory cells, microvascular endothelial cells, adrenergic nerves, pericytes, and stem and progenitor cells. These stem cells deliver all cell types that are needed for new vessel formation such as endothelial and SMCs as well as pericytes (32, 68). They are involved in neovascularization due to tumor (31, 148), stress, or insult (32, 88). ROS are critically involved in the activation of stem cells. Knockout of the NADPH oxidase impairs EPC activation and resulted in reduced tissue repair after ischemia. In endothelial stem cells, proliferation is associated with higher levels of superoxide and hydrogen peroxide than in quiescent cells. NOX2 and NOX4 play a role in activating several signal transduction pathways, including receptor tyrosine kinases and activation of p38 MAPK, ERK, and Akt, thereby enhancing endothelial cell proliferation and survival. ROS also play a role in endothelial cell migration. Angiogenic factors such as VEGF or angiopoietin-1 utilize ROS for signal transduction inducing cell migration (65). Upon VEGF binding to its receptor VEGFR, the GTPase Rac1 is recruited (152). Activated Rac1 recruits NOX subunits to initiate ROS production (55). Migration is initiated in quiescent endothelial cells by breaking up their intercellular connections. The major adhesion component is vascular-endothelial- (VE-) cadherin, which forms a dimer and binds directly to β-catenin. ROS-mediated phosphorylation of VE-cadherin and β-catenins leads to disassembly of VE-cadherin-catenin complex and EC junctional breakdown (142).
ROS and cellular senescence
Mitochondrial ROS are primarily generated from electron leakage of the mitochondrial respiratory chain complexes I and III, which damage all components of the cell, including DNA, lipids, and proteins. Dysfunction of the mitochondrial respiratory chain and inefficient oxidative phosphorylation may lead to more electron leakage and further increases in ROS generation, resulting in a detrimental cycle that causes eventual, irreversible damage to cells and contributes to aging. One of the first observations that suggested a role for ROS in cell aging was that the replicative life span of in vitro-cultured cells was significantly extended by culturing the cells in a low-oxygen environment, which might be due to less ROS generation (101).
ROS signaling in vascular stem cells is an important regulator of stem cell differentiation. On the contrary, ROS limits the self-renewal potential of stem cells. This might be regarded as beneficial as limitation of proliferation might be a protective defense from cancer. Limiting the self-renewal ability, however, is associated with senescence and aging. Aging is associated with reduced cardiomyocyte turnover as a consequence of the reduced cardiac growth reserve (116, 145). Senescent and dysfunctional cardiac resident stem and progenitor cells accumulate as a consequence of cardiac pathology (96). The intracellular redox state is one of the driving factors of aging. In cardiomyocytes, an increased oxidative burden is associated with oxidative modification of regulatory molecules and contractile proteins affecting cardiomyocyte contraction. Performing analyses on mouse aortic tissue of different ages it was recently shown that aortae of aged mice (9 months) exhibit enhanced expression of NOX2 and 4-HNE, particularly in the aortic intima and adventitia that serve as a vascular stem cell niche (67). Interestingly, these alterations were suppressed in the aortae of mice that were deficient for the cell adhesion molecule CEACAM1 (carcinoembryonic antigen-related cell adhesion molecule-1) (67).
Age also affects the stem cell microenvironment. Increased levels of DNA damage and reduction in telomere length are eventually causing a senescent phenotype in stem cells (20). The regenerative potential of stem cells has also been found to be compromised or even lost with aging as a consequence of senescence (134). The embryonic, hematopoietic, and mesenchymal compartment depends on a hypoxic environment to keep stem cells in a quiescent state. Physiological oxygen tension is required for stem cell commitment, but causes senescence and exhaustion. A critical balance between hypoxic and normoxic conditions is required for tissue homeostasis and for the preservation of the stem cell pool during the life span (138, 139). It has been shown that a decrease in ROS levels improves stem cell maintenance, suggesting that antioxidant enzymes could play a significant role in the preservation of the regenerative function of stem cells (57, 98).
A critical role of oxygen tension in the induction of a senescence phenotype has been observed in MSCs. MSCs express higher levels of glycolytic enzymes and lower levels of respiratory chain components, suggesting that MSCs rely more on glycolysis. When cultured in vitro, however, enzymes of oxidative phosphorylation and O2 consumption increase. Under these conditions, MSCs show a three- to fourfold increase in senescence, suggesting that hypoxia-induced glycolysis limits MSC proliferation to prevent oxidative stress-induced senescence and preserve MSC long-term self-renewal (103).
EPCs are derived from the bone marrow and circulate in the bloodstream, home to sites of new blood vessel formation (particularly in ischemic tissues), and contribute to new blood vessel formation. Because acute ischemia and in particular, reperfusion are characterized by enhanced ROS production in affected microenvironments, it was suggested that EPCs may be particularly resistant to ROS-induced damage. Indeed, the upregulation of manganese SOD kept levels of ROS very low (26, 51). Interestingly, the protection against ROS seems to deteriorate with age. The long-lived EPCs are repeatedly exposed to oxidative stress over the lifetime of an organism. Oxidative damage may accumulate over time, which likely contributes to reduced cellular EPC function (15). Increased ROS production during MI was associated with reduced EPC levels (131). Increased production of ROS is also strongly associated with diabetes (99). Hyperglycemia may contribute to accumulating oxidative damage, diminishing EPC function as well. EPCs derived from diabetic patients were reduced in number, showed premature senescence phenotype, and had a reduced ability of vessel formation (56). These dysfunctions may contribute to the increased risk of developing vascular dysfunction in diabetic patients.
The molecular mechanism of how ROS are limiting the self-renewal capacity in stem cells was demonstrated in HSCs in mice deficient in the protein ataxia telangiectasia mutated (ATM). ATM is a serine threonine protein kinase and is a critical enzyme in the regulation of cellular stress response to DNA damage. ATM deficiency is associated with ataxia telangiectasia, an autosomal disorder characterized by neuronal degeneration, immunodeficiencies, predisposition to lymphomas, and extreme sensitivity to ionizing radiation (87). ATM also plays an important role in reducing ROS levels. Consequently, increased ROS levels are detected in ATM-deficient mice. ATM-deficient mice develop early-onset bone marrow failure, indicating a defect in the maintenance and repopulation ability of HSCs.
Correlating with increased levels of ROS, an activation of the p38 MAPK pathway and increased expression of p16 and p19 in the HSC compartment have been observed (73). p16INK4a and p19ARF are tumor suppressors generated from alternative open reading frames of the INK4a locus. Upregulation of p16INK4a, but not p19ARF, mediates the HSC defects in ATM knockout mice. Treatment with the antioxidant NAC or a p38 MAPK-specific inhibitor rescues these mice from bone marrow failure, indicating that ROS-mediated activation of p38 MAPK induces a block in cell cycle progression. The gene POLG encodes for the mitochondrial DNA polymerase G. Similar to APC deletion, the deletion of POLG in mice results in a significant upregulation of ROS. In POLG-deficient mice, dysfunctions in neuronal stem cells and in HSCs were also rescued by supplementation with NAC (3). NAC also restored the ROS-mediated deficiency of POLG-mutant cells to be reprogrammed into iPSCs (48), suggesting the effects of ROS in regulating the quiescence of stem cells.
A direct link of mitochondrial ROS production and regulation of aging is provided by p66shc. The protein p66shc is a splice variant of p52shc/p46shc, which are inductors of the Ras signaling pathway. The splice variant p66Shc is a redox-regulated protein implicated in producing mitochondrial ROS and inducing apoptosis. Accordingly, p66Shc−/− mice show prolonged life span, reduced production of ROS, and increased resistance to oxidative stress (95). These data show that H2O2 generated by p66Shc significantly contributes to the intracellular ROS burden. p66Shc primarily localizes to the mitochondrial IMS, where it directly oxidizes cytochrome c, receiving electrons from Cyt c and transferring them onto molecular oxygen, thereby producing H2O2 (43). p66shc is regulated via reversible phosphorylation (106). Phosphorylation is an essential regulator of p66Shc function, and phosphorylation of this site increases with age in several organs, leading to higher ROS production rates and accumulation of oxidative damage, as demonstrated in aged mice (71). In summary, p66shc is part of a signal transduction pathway that regulates stress apoptotic responses and life span in mammals (111).
Induction of proliferation in stem cells is associated with a significant increase in protein synthesis. Transport and folding of proteins into the ER is a redox-sensitive process. Oxidative folding is mediated by the protein disulfide isomerase (PDI) catalyzing the formation of disulfide bond in its substrate proteins. Disulfide bond formation requires the transfer of one electron from PDI to the Ero1 (endoplasmic reticulum oxidoreductin-1) and molecular oxygen (46). Therefore, oxidative folding may result in an increased generation of ROS. The cellular glutathione pool is required in this process to maintain ER oxidoreductases in a reduced state and to buffer ROS produced by the oxidative folding process. The balance between reduced and oxidized glutathione may be challenging under conditions of increased protein synthesis that saturates the folding capacity of the ER and eventually leads to the accumulation of unfolded proteins in the ER lumen. ER stress is sensed by the unfolded protein response. The transcription factor CCAAT-enhancer-binding protein homologous protein (CHOP) is upregulated by ER stress in HSCs. CHOP plays an important role in the induction of apoptosis. A particular role for the ASK1 (apoptosis signal regulating kinase 1), an oxidative stress-responsive kinase, that leads to the activation of p38 MAPK and JNK was found in stem cells (146). Although ER stress and the activation of stress response pathways have been suggested to be contributors to senescence and aging, clear evidence for this hypothesis is still missing (25, 85, 122).
Conclusion and Future Perspective for Therapy
Understanding the molecular mechanisms, how metabolism and ROS regulate stem cell renewal and differentiation is a key for advancing the utilization of stem cells in therapeutic approaches. Stem cells and progenitor cells including EPCs are susceptible to excessive oxidative stress. A promising therapeutic approach would be to decrease oxidative stress and to improve EPC function (120, 154). Antioxidative agents, such as resveratrol and salvianolic acid, have been found to enhance activity of EPCs (46a, 77). The regenerative potential of stem cells has attracted much attention in replacing damaged tissue supporting wound healing and tissue repair. Bone marrow mononuclear cells (BMNCs) were considered for cardiac cell therapy, as they are relatively easy to obtain. BMNCs were shown to promote angiogenesis and neovascularization in models of skin wound healing, MI and hindlimb ischemia (6, 40).
However, injected cells largely failed to differentiate, so that clear evidence for their potential to successfully undergo differentiation and tissue replacement is still lacking (82). MSCs are a subset of bone marrow-derived stem cells. Successful differentiation of MSC into a large range of cell types including adipocytes, chondrocytes, and osteoblasts has been shown in vitro (27). Therefore, MSCs have been tested to promote regenerating human heart tissue. Considering the low numbers of MSCs after transplantation, the improvement in myocardial function was quite substantial. But also here, evidence for a differentiation of MSC into cardiomyocytes is limited. Instead, the improvements of cardiac function after stem cell treatment have been largely attributed to the secretion of paracrine factors, which may play a tissue-protective role (49). The generation of cardiomyocytes using induced pluripotent stem cells provides a reliable source of human cardiomyocytes for therapeutic applications. Studies have shown successful integration of these cells into the heart muscle and electrical coupling and a beneficial effect on cardiac function in several heart disease models (21, 79, 132, 162). Clinical trials are ongoing and show promising outcomes regarding safety and support the therapeutic concept (81, 90).
Footnotes
Authors' Contributions
J.D. and C.M. contributed to conceptualization; J.D. and I.K. contributed to collection and analysis of bibliography; J.D., C.M., and I.K contributed to writing original draft. All authors read and approved the final article.
Funding Information
The work in the laboratory of the authors is supported by the Deutsche Forschungsgemeinschaft (DFG; Jan Dudek: DU1839/2-1; CM: Ma2528/7-1; SFB 894; TRR-219) and the Bundesministerium für Bildung und Forschung (BMBF; DZHI, 01EO1504; CF.3, RC.2), Corona Foundation (Christoph Maack), and Barth Syndrome Foundation (Jan Dudek and Christoph Maack).