
Abstract
Significance:
Acquired sensorineural hearing loss is a major public health problem worldwide. The leading causes of sensorineural hearing loss are noise, aging, and ototoxic medications, with the key underlying pathology being damage to the cochlea. The review focuses on the phenomenon of preconditioning, in which the susceptibility to cochlear injury is reduced by exposing the ear to a stressful stimulus.
Recent Advances:
Cochlear conditioning has focused on the use of mono-modal conditioning, specifically conditioning the cochlea with moderate noise exposures before a traumatic exposure that causes permanent hearing loss. Recently, cross-modal conditioning has been explored more thoroughly, to prevent not only noise-induced hearing loss, but also age-related and drug-induced hearing losses.
Critical Issues:
Noise exposures that cause only temporary threshold shifts (TTSs) can cause long-term synaptopathy, injury to the synapses between the inner hair cells and spiral ganglion cells. This discovery has the potential to significantly alter the field of cochlear preconditioning with noise. Further, cochlear preconditioning can be the gateway to the development of clinically deployable therapeutics. Therefore, understanding the underlying mechanisms of conditioning is crucial for optimizing clinical protection against sensorineural hearing loss.
Future Directions:
Before the discovery of synaptopathy, noise exposures that caused only TTSs were believed to be either harmless or potentially beneficial. Any considerations of preconditioning with noise must consider the potential for injury to the synapses. Further, the discovery of different methods to precondition the cochlea against injury will yield new avenues for protection against hearing loss in the vulnerable populations. Antioxid. Redox Signal. 36, 1215–1228.
Introduction
Hearing loss affects millions of children and adults in the United States and worldwide. The Johns Hopkins Bloomberg School of Public Health estimates that 14.3% of Americans aged 12 years or older have hearing loss (23).
That percentage climbs to 33% of Americans between the ages of 65 and 74, and ∼50% at age 75 and older (60). There are numerous potential causes of hearing loss, including ear diseases that affect the outer or middle ear, age-related hearing loss (ARHL; also known as presbycusis), congenital syndromic and nonsyndromic hearing losses, and acquired sensorineural hearing losses based on experiences. Sensorineural hearing losses are those affecting the cochlea, auditory nerve, and/or the central auditory nervous system. Two of the leading causes of acquired sensorineural hearing losses are exposure to high-level noise or ototoxic drugs.
Both noise and ototoxic drugs are known to damage the cochlea, the sensory receptor organ of the auditory system. The main function of the cochlea is transduction of mechanical energy from the movement of the tympanic membrane and middle ear bones into the electro-chemical signaling of the nervous system. Two sensory receptor cell types are housed within the organ of Corti, as well as a set of specialized support and structural cells that maintain an exquisitely complex mechanical architecture and delicate metabolic homeostasis. Either mechanical or metabolic disruption of the organ can result in irreversible damage or cell death that underlies functional hearing loss.
Noise and ototoxic drugs can cause mechanical and metabolic injuries to the cochlea. Mechanical injuries are largely defined by the magnitude of injury to the organ of Corti, which is itself a product of the intensity and duration of noise exposure or the dose and timing of an ototoxic drug exposure. Short-duration, catastrophic injuries to the organ of Corti can result from blast or impulse noise exposures. Likewise, very high levels of ototoxic drugs given in single bolus doses can cause extensive cochlear damage to sensory cells. More commonly, noise- and drug-induced hearing losses are cumulative and progressive, occurring over a period of time with repeated exposures. In these instances, the metabolic homeostasis of the cochlea is likely disrupted in a significant way (66).
The cochlea is inherently susceptible to injury from metabolic changes. However, that susceptibility is variable across individuals. For example, susceptibility to noise-induced hearing loss (NIHL) can have a broad range across individuals, even when accounting for age (39). Similarly, prevalence data on ototoxicity vary widely, and they reflect differences in dosing, clinical delivery protocols, and individual inherent susceptibility. Prevalence of ototoxicity from drugs such as cisplatin (CDDP), a powerful chemotherapy compound used against several cancers, has been reported at 40% (88), 55% (94), and 72% (71).
Susceptibility to cochlear injury from noise or ototoxicity has been modified in numerous animal models through a variety of interventions, including exogenous pharmaceutical or nutraceutical treatments, genetic modifications, and manipulation of endogenous factors. The cochlea's endogenous defenses can be strengthened by using a cochlear preconditioning or toughening paradigm.
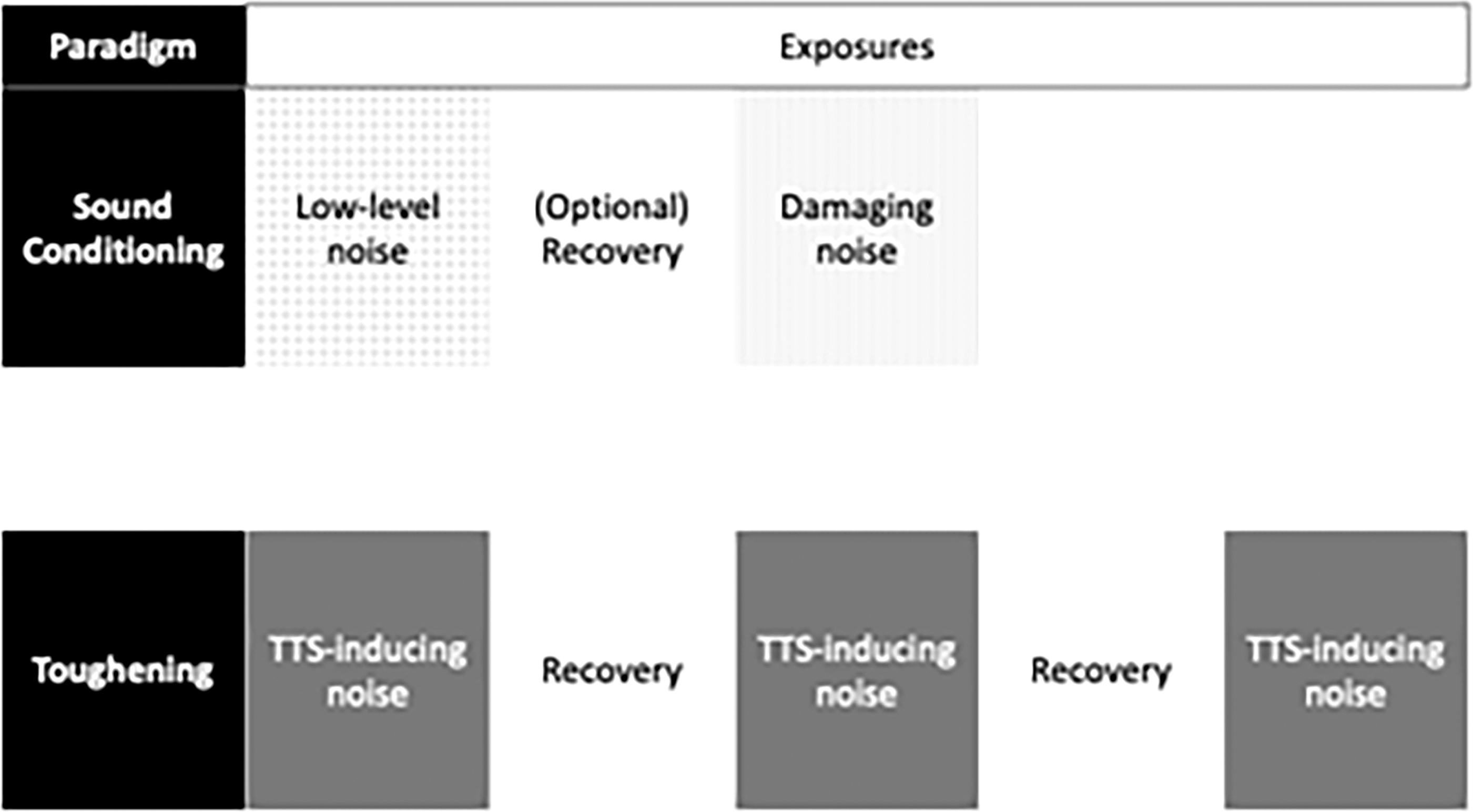
Cochlear preconditioning involves exposing a subject to a stressor that is intended to render the body more resistant to future cochlear injury. Specifically, sound conditioning involves exposing the subject to a noise that stresses the cochlea without permanently damaging it. The underlying principle is that exposure to stressful levels of sound will stimulate the ear's defenses against permanent injury, thus decreasing its endogenous susceptibility to injury and subsequent hearing loss. Cochlear preconditioning can be mono- or cross-modal. As will be discussed next using noise as a stressor for cochlear preconditioning has been researched the most thoroughly.
Definition and Discovery
Though the terms “sound conditioning” and “toughening” are used interchangeably, Niu and Canlon offer two separate definitions. Both phenomena have been demonstrated in a variety of animal species, as discussed next. The two terms describe a process in which the susceptibility of NIHL is reduced, but sound conditioning is the phenomenon where a paradigm of low-level, nondamaging noise is presented continuously or intermittently to reduce susceptibility to a subsequent acoustic trauma. The term “noise conditioning” is often used instead of “sound conditioning,” but these terms are synonymous. The toughening paradigm uses an intermittent noise that causes a temporary threshold shift (TTS). The degree of TTS is reduced after repeated exposures, sometimes resulting in no measurable postnoise TTS (61). TTS is an elevation of the hearing sensitivity threshold that completely recovers over time.
TTS is contrasted with permanent threshold shift (PTS) or compound threshold shift (CTS). PTS is an irreversible elevation of threshold, with a corresponding and permanent cochlear injury. CTS is a combination of TTS and PTS. For noises that cause PTS, hearing loss often begins as CTS. In the acute time window during and shortly after sound exposure, threshold elevation reflects both TTS and PTS components, but after the cessation of the noise, the TTS component will recover over time, leaving only PTS (57). Cochlear preconditioning or toughening can reduce susceptibility to TTS or PTS resulting from noise or ototoxic compounds (chemicals/drugs that injure the inner ear, specifically the cochlea).
There is a robust body of literature on sound conditioning and toughening. However, concerns have arisen over the past several years regarding the use of conditioning with sound that produces TTS. This is because TTS injury from noise has been linked to previously undetected cochlear damage. Although TTS does not appear to produce loss of sensory cells, TTS has been associated with damage to synaptic terminals on inner hair cells (IHCs), the primary conduits of acoustic information to the central nervous system. This synaptopathic damage occurs between IHCs and the dendrites of afferent spiral ganglion neurons (45, 46, 51). The topic of synaptopathic loss will be discussed in more detail later in the review of the current status of cochlear preconditioning research.
Resistance to the loss of cochlear hair cells or supporting cells after the presentation of noise was first described by Miller et al. in experiments using a cat animal model. When cats were exposed to intermittent noise for 16 days, threshold shifts at the end of the exposure were lower than those at the beginning, suggesting an ear-toughening effect as a function of repeated exposure (58). This was followed by a publication that demonstrated that the toughening phenomenon was also present in chinchillas, specifically reducing TTS (92). The same group showed evidence that toughening after TTS occurred in humans as well (91).
Later research described the histological effects of toughening in the chinchilla cochlea. The location of damaged cell lesions (including IHCs and outer hair cells [OHCs] and select populations of supporting cells) was the same in conditioned and unconditioned chinchillas, though the conditioned chinchillas had smaller lesions than their unconditioned counterparts (6).
Later that year, the same group published evidence that the toughening paradigm does not produce asymptotic threshold shifts (ATSs) in chinchillas (14). ATS is the phenomenon in which noise induces a threshold shift over time, and repeated exposure to that same noise will not increase the overall threshold shift. Instead, the threshold shift asymptotes or plateaus at a specific magnitude related to the sound pressure level of the noise (89). The toughening in the chinchillas prevented the ATS from developing, and instead led to recovery of as much as 30 dB of threshold shift using an interrupted noise exposure, as compared with a continuous noise exposure (14).
Preliminary evidence for the sound conditioning paradigm (as opposed to toughening) was presented in the early 1980s. Pigmented guinea pigs experienced significant TTS after exposure to a 10 kHz tone for 1 min at 97 dB sound pressure level (SPL), then again at 103 dB SPL for 1 min. This group showed reduced TTS compared with a group that received only the second, 103-dB SPL exposure (68). Despite these results, the paradigm of a prolonged conditioning period was not developed until years later.
Canlon et al. exposed Hartley guinea pigs to a 1 kHz tone at 81 dB SPL for 24 days and allowed a short recovery period, after which their auditory brainstem response (ABR) thresholds were measured, and then exposed the animals to the same noise at 105 dB SPL for 72 h. The group that received conditioning showed lower ABR thresholds immediately after the traumatic exposure and at 8 weeks postnoise, than the control group that underwent no prenoise exposure conditioning (Fig. 1). At the time, the authors hypothesized that elevated metabolic activity in the OHCs increased neural or synaptic plasticity, or that the middle ear muscles (MEMs) could play a role in the conditioning phenomenon (11). During the time since that paper was published, the majority of the research in the field has been related to conditioning, rather than toughening.

Characteristics of Toughening and Conditioning with Sound
Sound toughening has been shown in a variety of species, most notably humans (59, 91). Other species include Wistar rats (1), cats (58), and chinchillas (14, 92). Sound conditioning has also been demonstrated across a number of species, including chinchillas (10, 79, 80), guinea pigs (11, 12), gerbils (73, 93), Wistar rats (66), rabbits (21), and CBA/CaJ mice (103). Despite the use of the term “toughening” in the study describing the effect in Wistar rats, by the definition presented earlier, the study concerns the conditioning phenomenon (66).
The key distinction between sound conditioning and other forms of cochlear preconditioning is the use of a moderate-intensity sound as the conditioning stimulus. Sound conditioning has been studied most often by using paradigms with a conditioning period either immediately or shortly preceding an exposure to traumatic noise (Fig. 2). One of the earliest sound-conditioning experiments is representative of this phenomenon.
This experiment used a 0.5 kHz octave band noise at 95 dB for 6 h/day for 10 days as the conditioning stimulus. The chinchillas undergoing conditioning were given a 5-day rest period before noise exposure whereas unconditioned chinchillas were exposed to the 0.5 kHz noise for 6 h at 106 dB SPL. Both the conditioned and unconditioned chinchillas showed threshold shift and cochlear damage, but the conditioned group showed lower CTS and PTS than the unconditioned group (10). Interestingly, when the conditioning and damaging noises were presented at a higher frequency centered at 4 kHz, the conditioning effect was not present with a 5-day rest interval between the conditioning and the damaging noise. However, a strong conditioning effect was seen when the rest interval was only 18 h, with lower threshold shift and OHC loss (81).
The durability of a sound-conditioning effect is unclear. There is evidence, however, that protection against noise-induced PTS may last at least 60 days after the conditioning period, though the strength of the effect decreases as time postconditioning increases. To date, the longevity of this effect has only been tested in chinchillas presented with an octave band noise, centered at 0.5 kHz. Conditioning levels of 90 or 95 dB SPL (6 h/day for 10 days), preceding a traumatic noise of 106 dB SPL for 48 h, both produced and decreased PTS relative to un-conditioned animals, despite a lack of significant differences in OHC loss (Fig. 3) (55).

Not all published reports on sound conditioning have shown efficacy. Contrary to the results presented by McFadden et al., a recent experiment in albino guinea pigs showed that a conditioning noise exposure of 85 dB SPL 8 h/day for 3 days does not offer any protection against a traumatic exposure of 106 dB SPL for 2 h with respect to thresholds. The conditioned group, however, showed significantly fewer temporary and permanent synaptic losses after the acoustic trauma than the unconditioned group (18). Clearly, more research is needed to fully understand the relationship among sound conditioning paradigms, changes in hearing sensitivity, and physiological protection. Still, these results are encouraging, given the recent attention given to synaptopathic losses without functional evidence of threshold elevation (45).
Cross-Conditioning with Sound
Studies on sound conditioning have focused on protection against acoustic trauma, but there is also evidence that noise cross-conditioning, or sound conditioning that protects against other types of hearing loss, exists. First, noise conditioning appears to protect against the effects of ARHL. In two strains of mice (C57BL/6J and DBA/2J), exposure to 70 dB SPL of noise for 12 h/day was associated with preservation of physiological (ABR thresholds) and behavioral (startle amplitudes) measures of hearing. The C57BL/6J mice were exposed daily for 14 months, and DBA/2J mice were exposed daily for 9 months. The latter strain showed little difference in ABR thresholds at high frequencies (16 and 24 kHz), but they did show improvement at lower frequencies (95).
A similar effect was observed in Fischer 344/NHsd rats. The conditioned group was exposed to 80 dB SPL noise for 12 h/day, 5 days/week, for 13 weeks. The exposures began after the onset of ARHL. Animals that received sound conditioning showed lower ABR thresholds and less OHC damage compared with the unconditioned group. In this case, the effect was strongest at high frequencies (15 kHz and higher). Interestingly, these data show that the conditioning noise exposures were able to slow the progression of ARHL, even though the loss was already in progress at the onset of the experiment (84).
Cross-conditioning with noise has also been shown to have a protective effect against drug ototoxicity. In these cases, animals were exposed to low- to mid-level noise before the administration of ototoxic drugs. Both platinum-based anticancer drugs (e.g., CDDP) and aminoglycoside antibiotics (e.g., kanamycin and gentamicin) are commonly used medical treatments. One of the main side effects of these drugs is ototoxicity that leads to and induces PTS (47, 74, 85). For example, sound conditioning has been used successfully to protect against the ototoxic effects of gentamicin. A conditioning noise of 1.4–5.6 kHz at 81 dB SPL for 3 weeks was presented to Mongolian gerbils. After acute delivery of gentamicin onto the round window of the cochlea, the conditioned group had significantly less OHC and IHC loss than the unconditioned group.
Interestingly, gentamicin is known to be vestibulotoxic in addition to being ototoxic, and the conditioning paradigm provided no protection against such vestibulotoxicity (82). It should be noted that, although this experiment featured a small sample size of two gerbils per condition, the otoprotective finding is supported elsewhere in the literature. In Sprague-Dawley rats, white noise centered at 8 kHz was presented at 85 dB SPL for 15 min before the administration of CDDP. Even this short duration of sound conditioning was enough to mitigate the ototoxic effects of CDDP. The study also tested two otoprotectant compounds, L-N acetylcysteine and D-methionine, each of which provided more protection against CDDP ototoxicity. However, the best results were achieved when the otoprotective drugs were used in combination with sound conditioning. The data show that sound conditioning with noise may augment the protective effects of antioxidant compounds against drug ototoxicity (86).
Similar sound conditioning against CDDP ototoxicity was found in CBA/CaJ mice. Roy et al. found that this protection was primarily in the mid-frequencies, with the greatest protection at 16 kHz. The authors speculated that the protection was related to the 8–16 kHz noise used in preconditioning (72). Theneshkumar et al. also found otoprotective effects, with the greatest protection observed at 8 kHz where the preconditioning noise was centered (86).
Finally, noise can also condition against ROS-induced damage from the administration of paraquat, a pesticide that generates ototoxic superoxides in the inner ear and can induce hearing loss and hair cell death (28). In experiments using a chinchilla animal model, subjects were conditioned with a 0.5 kHz octave band noise at 90 dB SPL for 6 h. The exposure was then followed by the administration of one of two doses of paraquat to the round window. Sound conditioning before administration was associated with reduced PTS and IHC loss in animals exposed to 5 mM paraquat (Fig. 4) and reduced IHC loss in animals exposed to 10 mM paraquat (28). Taken together, these experiments show the ability of noise to cross-condition against ototoxicity from several compounds known to induce oxidative stress in the inner ear.

Cochlear Preconditioning with Ototoxic Drugs
Although there is a robust literature surrounding sound conditioning, fewer studies have evaluated the effects of cochlear preconditioning using other challenges to the inner ear, such as ototoxic drug exposure. Ototoxic drugs have been evaluated both mono-modally as protectants against future ototoxic exposure to the same drug and cross-modally as a protectant against future noise exposure. Of the aminoglycoside antibiotics, mono-modal conditioning has been evaluated for both amikacin (15) and gentamicin (54) ototoxicity, and cross-modal conditioning has been evaluated with gentamicin (54) and kanamycin (19, 64) as the conditioning stimuli.
Ohlemiller and colleagues found kanamycin to be an effective protectant against PTS from noise exposure in the CBA/J mouse (19), but not in the C57Bl/6J mouse (64). The preconditioning paradigm used 300 mg/kg of kanamycin 24 to 48 h before noise exposure, and this preconditioning offered almost complete protection from noise-induced PTS while providing significant but incomplete protection from OHC loss. Based on findings from OHC loss and threshold shift across doses, the authors speculated that the smallest dose with the greatest efficacy in protecting against threshold shift would be 3+ doses of 300 mg/kg spaced no more than 48 h apart before exposure to damaging noise.
Although the aforementioned research group saw robust protection of kanamycin against noise in the CBA/J mice studied, there was almost no protection from PTS or OHC loss in the C57Bl/6J mice. Similarly, Roy et al. saw little protection from OHC loss or ABR threshold shifts with the use of 750 mg/kg of kanamycin twice daily for 17 days as the preconditioning stimulus before exposure to 2 h of 90.1 dB SPL noise. The only protection observed in this group was at 32 kHz and in the corresponding region for the OHCs for these CBA/CaJ mice (72). It is possible that that the effects of preconditioning do not generalize from the CBA/J mouse to the CBA/CaJ mouse, but it is of note that both the C57Bl/6J in Ohlemiller et al.’ 2011 study (64) and the CBA/CaJ mice in this study received twice daily doses of kanamycin and yielded negative results, whereas the CBA/J mice were dosed less frequently and exhibited a protective effect. Thus, the difference may be attributable to the dosing schedule.
These are the only published studies of cross-modal conditioning using kanamycin in the mouse. As such, additional research is needed to determine the generalizability of these findings to other species and to identify whether the dosing paradigms that were most successful in the CBA/J mouse would yield different results in other inbred mouse strains.
Mono-modal evaluation of both amikacin and gentamicin has demonstrated a protective effect of low-dose pre-exposure to the drug before a high (simulated therapeutic) dose of the same drug in albino guinea pigs. Maudonnet et al. reported that 30 days of exposure to 10 mg/kg/day of gentamicin protected both OHCs and distortion product otoacoustic emissions (DPOAEs) from the damaging effects of 10 days of exposure to 160 mg/kg gentamicin/day (54).
Similarly, De Oliveira et al. found that 30 days of exposure to 20 mg/kg/day of amikacin provided almost complete protection of the most basal OHCs when exposed to 400 mg/kg/day of the same drug (15). It is worth noting that in the De Oliveira study, no direct auditory function measures were collected, and animals were exposed to amikacin until the point that the acoustic startle reflex was absent. This may explain the degree of damage to OHCs in the more apical turns of the cochlea, despite the high-frequency protection seen in this study.
It is worth noting that although both amikacin and gentamicin are known ototoxins, neither has been found to be as harmful to the cochlea as kanamycin, and so additional study is required to determine both how these results would generalize to other species and how these results might generalize across drugs, even within the same class. This is particularly of note, as these findings differ significantly from those seen in the next section for another well-known ototoxin: CDDP.
Of the available literature, only one study evaluated the effects of mono-modal conditioning for CDDP. Harrison and colleagues (2015) evaluated the effects of alternating doses of either 2 or 3 mg/kg of CDDP every other week for a total of 10 mg/kg over a period of 8 weeks. The Fischer 344/NHsd rats in the study were then exposed to a 12 mg/kg dose of CDDP, a dose sufficient to create significant ABR threshold shifts and OHC loss (30). The OHC counts indicated that the low conditioning doses alone caused a mild OHC loss. The major finding was that the group exposed to both the preconditioning and damaging CDDP doses saw significantly higher threshold shift and OHC loss than the group exposed to the high dose alone (Fig. 5) (29). Thus, the “conditioning” stimulus potentiated the hearing loss rather than limiting it.

Subsequent literature has indicated that ears exposed to CDDP have an elevated risk for injury even long after the exposure (16), and it has been postulated that CDDP is retained in the cochlea indefinitely (9). When pairing these findings with data showing that CDDP is known to be increasingly damaging with an increasing cumulative dose (50), it is unsurprising that preconditioning of the cochlea with additional exposure to CDDP was found to be ineffective. This does not mean, however, that preconditioning cannot be used as a means of protection against CDDP ototoxicity as was demonstrated using noise, to precondition the cochlea before CDDP exposure.
The studies covered in this review provide promising explorations of noise preconditioning for ototoxicity across two rodent species. Additional research is warranted, especially as these studies focused on threshold shift and OHC counts rather than ribbon synapse counts (see Current State of the Literature) that may be more vulnerable to change from these insults. However, given the variability described in other sections of this article, this confirmation of results across species is a promising beginning to the evaluation of this effect.
Potential Mechanisms
The exact mechanisms behind sound conditioning are still unknown, however decades of experimentation have ruled out many contenders. Later, we present a short history of the proposed mechanisms for sound conditioning, culminating in a review of the mechanisms currently believed to be the most likely candidates.
Canlon et al. were some of the first to predict underlying mechanisms for sound conditioning. Among these predictions was the function of the MEMs, and particularly the stapedius muscle (11). This was in contrast to the earliest report of sound conditioning, which hypothesized that the MEMs were not the underlying cause of the conditioning phenomenon (68). Two groups presented research years later that dispute this prediction as well. Mongolian gerbils (73) and chinchillas (33) underwent surgery to section their MEMs. In both cases, there were no significant differences between the ABR thresholds in normal ears and ears with sectioned MEMs in animals that underwent sound conditioning. Therefore, both groups concluded that the MEMs did not contribute to sound conditioning.
The auditory olivo-cochlear efferent system has also been hypothesized as an influence or underlying cause of the sound conditioning phenomenon (104). However, there is considerable evidence that the efferent system is, in fact, not responsible for sound conditioning. Yamasoba and Dolan reported that strychnine de-efferentation of pigmented guinea pigs raised DPOAE thresholds, but that the protective effect of conditioning showed similar threshold shifts in normal and de-efferented ears (99).
In addition, Kujawa and Liberman tested this relationship with de-efferentation surgery in albino guinea pigs and found that the groups that underwent sham surgery were more resistant than the de-efferented animals to severe NIHL as a result of traumatic noise exposure, regardless of whether or not the animals experienced the conditioning noise exposure (44). The effect does not occur in ears contralateral to the presentation of noise, which further suggests that the auditory efferent system is not directly involved in protecting against NIHL after sound conditioning. It, instead, suggests that the effect is local to the cochlea, rather than a systemic effect (100).
Despite evidence ruling out certain mechanisms, there are many yet to fully explore, and it is not known which of the following is primarily responsible for the protection associated with sound conditioning, including F-actin modulation; apoptosis suppression; stress responses, including heat shock proteins (HSPs) and glucocorticoids (GCs); and protection against reactive oxygen species (ROS) and reactive nitrogen species (RNS).
In addition to protecting against NIHL from continuous noise exposures of long duration, sound conditioning can also protect chinchillas against ABR threshold shift and hair cell loss from exposure to repeated impulse noise (34), which is known to induce mechanical damage to the cochlea (78). One indirect measure of cochlear integrity is quantification of actin, a structural protein present in OHCs and some support cells that plays a role in the motility of the OHCs and of the organ of Corti as a whole. F-actin, the filamentous form of actin, can be labeled, quantified, and used to estimate the relative motility of the organ of Corti. Chinchillas were conditioned with a 0.5 kHz octave band noise at 90 dB SPL for 6 h/day for either 10 days, followed by a traumatic noise at the same frequency and 105 dB SPL for 4 h. This exposure successfully created a sound-conditioning effect, and F-actin staining showed that the conditioning group showed a decrease in F-actin density in the stereocilia and cuticular plates of the OHCs, as well as the Deiters' cells.
Though more research is required to determine the effect to which this plays a role in sound conditioning, it was hypothesized that the decrease in F-actin after sound conditioning represents a lessening of mechanical rigidity in the organ of Corti. This may protect the organ of Corti from some of the mechanical trauma incurred during a traumatic noise exposure (37).
In addition to the mechanical damage discussed earlier, hair cells in the cochlea can undergo metabolic damage as a result of noise exposure. This damage leads to apoptosis, a type of programmed cell death that can occur in cochlear cells (32). To test the effect of apoptosis suppression in the sound-conditioning phenomenon, the activity of the apoptosis-suppressor gene, bcl-2, and the level of cytochrome c, a protein involved in the apoptotic pathway, were tested during a typical sound conditioning experiment.
Fifty-six pigmented guinea pigs were conditioned with a 1 kHz tone at 81 dB SPL for 24 days and then exposed to the same tone at a traumatic level of 105 dB SPL for 72 h. The study showed that cytochrome c is released into the cytosol of the OHCs after an acoustic trauma and that sound conditioning protected against this release. Further, the expression of bcl-2 increased after sound conditioning. Taken together, the findings suggest that bcl-2 and apoptosis suppression play a role in sound conditioning (62).
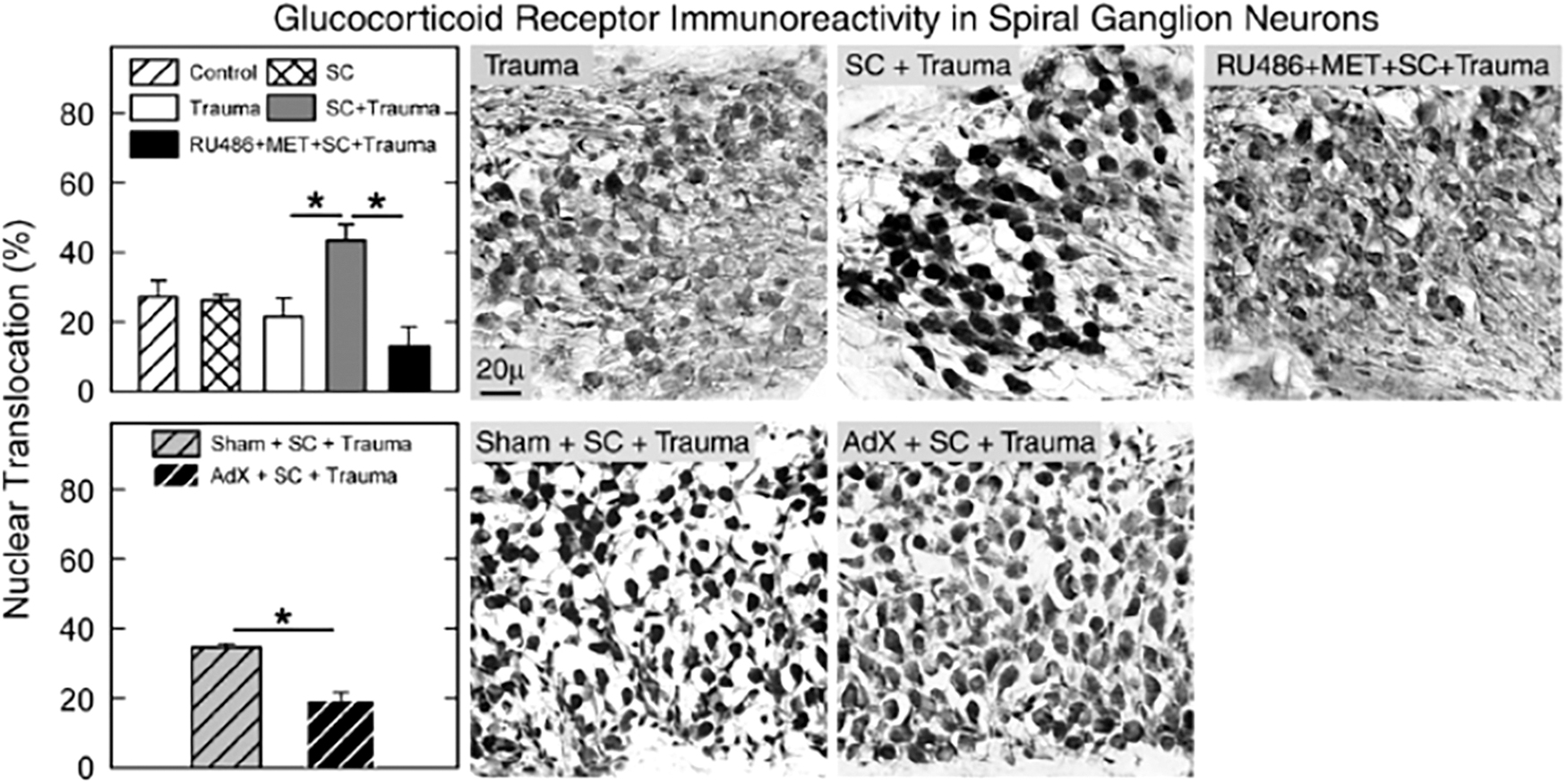
The effects of stress on the auditory system and how it affects sound conditioning involve both GCs and HSPs. The GCs are steroid hormones released as a result of stress-induced activation of the hypothalamic-pituitary-adrenal axis. Corticosterone is the predominant GC in rodents, and levels of corticosterone are increased after noise exposures (56). For an in-depth review of GCs and the auditory system, see Meltser and Canlon (56). The CBA mice were assigned to a variety of experimental groups to investigate the role of corticosterone on sound conditioning. For all tests, the conditioning noise was an 8–16 kHz stimulus at 89 dB SPL for 15 min and the acoustic trauma was the same noise at 100 dB SPL for 2 h.
The study had four important findings. First, the noise paradigm used successfully produced a sound-conditioning effect. Second, the protection of sound conditioning disappeared when mice were injected with a GC synthesis inhibitor (metyrapone) and a GC receptor antagonist (RU486). Third, the protection of sound conditioning also disappeared when mice underwent adrenalectomy surgery. Finally, the administration of a corticosterone supplement restored the protective effect of sound conditioning (Fig. 6). Taken together, these data show that without corticosterone (either endogenous or exogenous), sound conditioning does not protect against traumatic NIHL in CBA mice (83).
Further evidence for the importance of GCs in conditioning comes from a stress-conditioning study in which CBA/CaJ mice were restrained (12 h/day for 2 days) to cause a stress response before being exposed to an acoustic trauma (8–16 kHz at 100 dB SPL for 2 h). Noise was not used for conditioning. The mice that were exposed to the acoustic trauma 2 h after the end of the restraint showed significantly increased corticosterone levels and had significantly less PTS (measured by compound action potentials [CAPs] and DPOAEs). This protective effect was reduced to insignificant levels by 24 h after restraint (90).
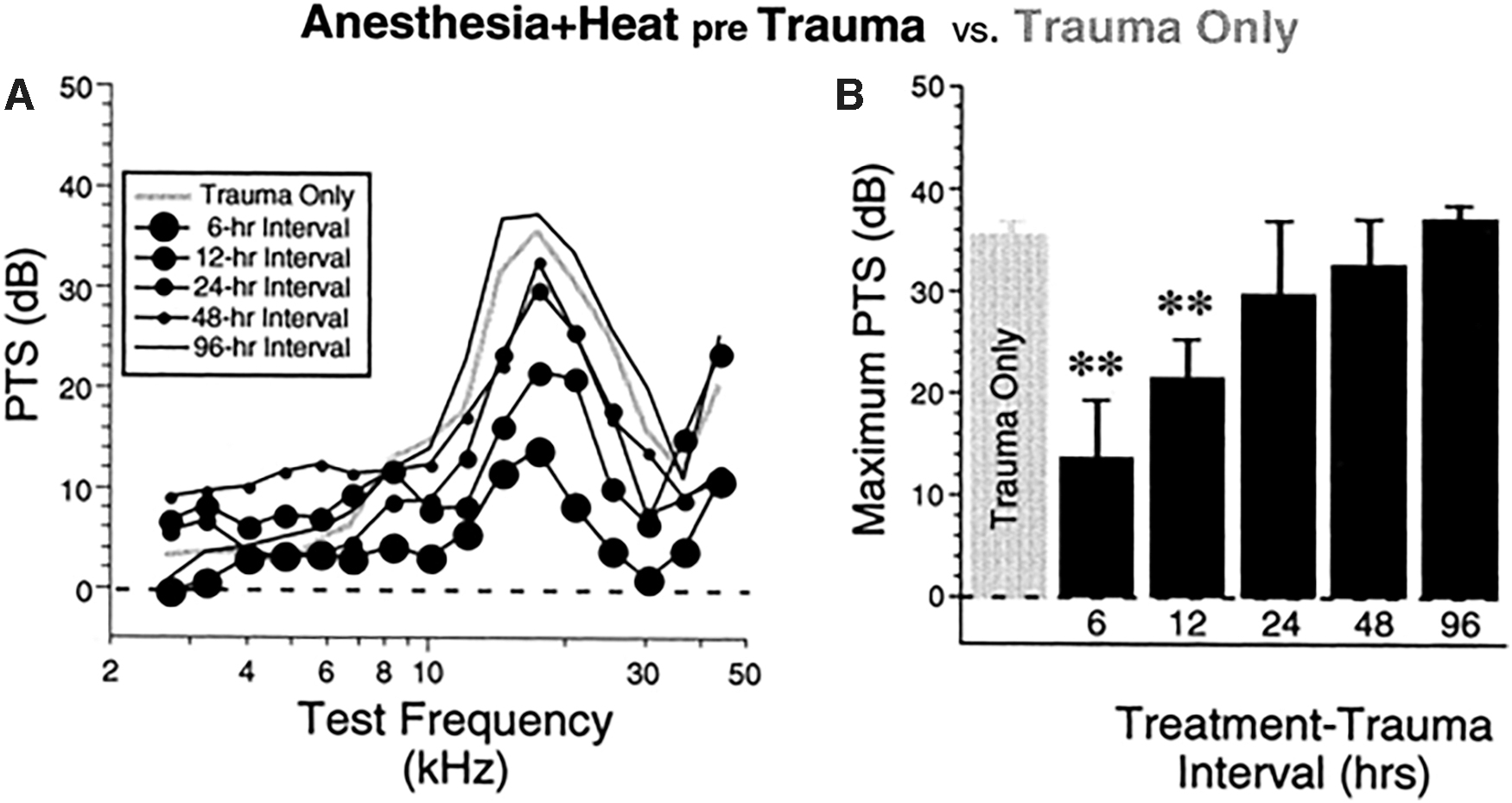
GCs are not the only stress-related molecules that are studied in relation to conditioning. CBA/CaJ mice were heat-stressed before exposure to a damaging noise. Again, groups were determined by the length of the interval between the stressor and the damaging noise exposure. Mice that were heat-stressed 6 or 12 h before the noise showed significantly less PTS (measured by CAPs and DPOAEs), though the protective effect did not extend to conditioning delivered 24 h or more before the traumatic noise (Fig. 7). The specific HSP studied in this article, HSP70, was upregulated at least 30 min after heat stress, but levels returned to baseline by 6 h after the stressor (102). Despite the relationship between HSP upregulation and conditioning against NIHL, there is no evidence that there is a direct connection, and the authors hypothesized that this could be indicative of other mechanisms, such as F-actin and GCs.
HSP70 was also upregulated in guinea pigs that had undergone sound conditioning and high-level noise exposure. This change was also associated with changes in F-actin and calmodulin (106). HSP70 upregulation has also been specifically detected in spiral ganglion neurons of sound conditioned cochleae (105). This is particularly interesting, because so many of the reports on the conditioning phenomenon have focused on hair cells. Other HSPs, such as HSP27, are associated with increases in F-actin in Chinese hamsters (48). Heat stress is also associated with an increase in corticosterone and cortisol (another GC) levels in Kunming mice (24).
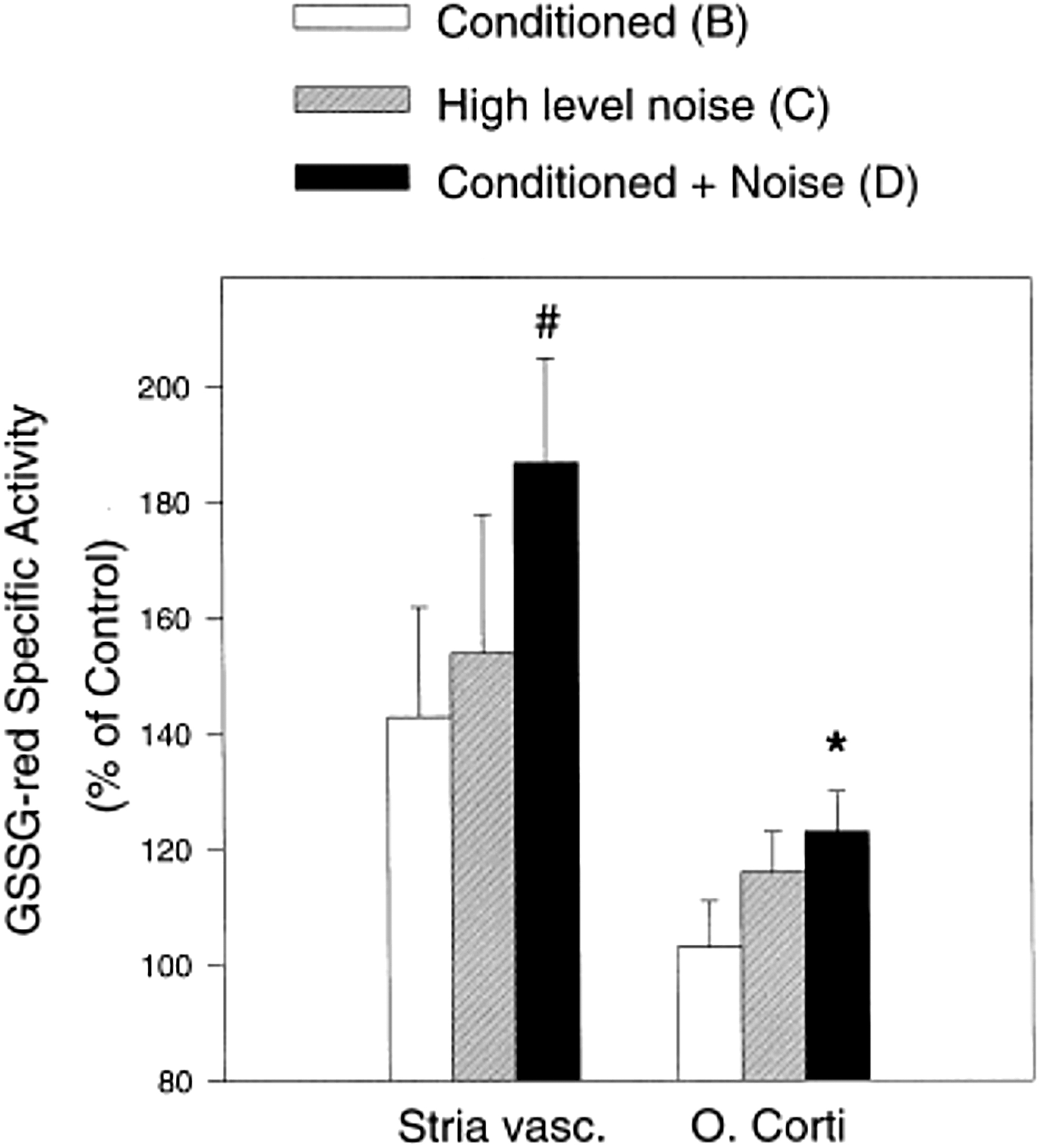
The final potential mechanism behind sound conditioning is antioxidant upregulation. It is well documented that noise exposures induce the formation of ROS throughout the cochlea, including in the OHCs, and potentially to levels where oxidative stress occurs (7, 32, 97, 98). There is a strong body of evidence indicating that noise causes ROS increases in the cochlea (63, 65, 97, 98). Further, several studies have demonstrated that enhancement of antioxidants can reduce NIHL in animal models (4, 27, 36, 38, 41, 42, 43, 65, 67, 75, 101). Changes in antioxidant enzymes have also been implicated in the sound conditioning effect. Chinchillas were conditioned with a 0.5 kHz octave band noise at 90 dB SPL for 6 h/day for 10 days and exposed to a damaging noise, consisting of the same octave band noise at 105 dB SPL for 4 h. Levels of three antioxidant enzymes, glutathione reductase, γ-glutamyl cysteine synthetase, and catalase, were measured in the stria vascularis and organ of Corti.
The animals were placed into one of four conditions: (i) no conditioning or damaging noise; (ii) sound conditioning alone; (iii) damaging noise without prior sound conditioning; or (iv) sound conditioning plus damaging noise 5 days later. The antioxidant enzymes in the cochlear lateral wall tissues and the organ of Corti were increased by all three noise conditions, relative to the untreated controls. Specifically, the highest antioxidant enzyme levels were found after the combined sound conditioning plus damaging noise (Fig. 8) (40). The implication was that the increased antioxidant enzymes reflected the cochlear tissues responding to the heightened ROS formation in the cochlea induced by the noise exposures. The preconditioning with sound prepared the cochlea to respond with a more robust antioxidant defense to minimize oxidative stress and reduce susceptibility to injury.
Superoxide dismutase (SOD) 1 and 2 are also enhanced in the cochlea after sound conditioning. This increase was linked to increases in HSP70, Bmi1, and FoxO1 in cochlear tissue from animals that experienced sound conditioning followed by high-level noise. In animals that underwent the high-level noise only, levels of the molecules decreased (105). This evidence, combined with the ability of noise to cross-condition against paraquat (28), indicates that sound conditioning can upregulate cochlear antioxidant capacity. Since ARHL (31) and ototoxic drugs (aminoglycosides, CDDP, etc.) (70, 96) have also been linked to oxidative stress as a mechanism underlying cochlear damage, antioxidant changes may also be a key mechanism in the cross-conditioning discussed earlier.
ROS upregulation in the cochlea also links to changes in calcium homeostasis. Increased free intracellular Ca2+ has been detected in animals exposed to high-level noise (22) and has been linked to ROS formation through NOX3 (69). Further, TRPV1, which is also increased by high-level noise (2), creates a loop with NOX3 that increases Ca2+ entry into cochlear cells, and ultimately leads to cell death through apoptosis (17). Increased intracellular Ca2+ can be attenuated by prior sound conditioning (106). Therefore, prevention of calcium dysregulation in the hair cells may be a possible contributing factor for sound conditioning as well.
In addition to ROS, noise exposures can increase levels of RNS, such as nitric oxide (NO), in the cochlea (76, 77). Further, NO may have protective effects based on its redox state (52) or concentration (13). Therefore, there is evidence that NO may play a role in toughening. Wistar rats were exposed to a 118 dB SPL white noise for 1 h, and the exposure was repeated four times. After repeated noise exposures, the degree of TTS incurred was reduced (compared with the TTS that occurred after the first exposure) and thresholds returned to levels that were not significantly different compared with baseline thresholds. Further, after noise exposures, neuronal NO synthetase, an enzyme that synthesizes NO, is upregulated in the spiral ganglion. These data together suggest that NO may play a role in toughening (1).
Despite the research done to date, there is still no conclusive single mechanism responsible for sound conditioning and toughening. Several other potential mechanisms have been postulated, and the current review is certainly not exhaustive. Future research should continue to explore potential mechanisms to explain sound conditioning, while paying particular attention to the two most promising candidates: stress responses and antioxidants.
Current State of the Literature
Thus far, this review has discussed the progression of the literature on cochlear conditioning after initial reports of the effect in cats in 1963. In animal models, preemptive noise exposures have been shown to have promising protective effects against NIHL and ARHL, though these effects appear to be present primarily at cellular and synaptic levels in the cochlea, and do not always protect hearing as assessed audiometrically. Similarly, priming the cochlea for a high-dose ototoxic drug by successive low doses of the same drug achieves similar conditioning effects in terms of cochlear survival, as shown with gentamicin and amikacin, but not kanamycin or CDDP.
Further, cross-modal conditioning has been observed by preventing ototoxic damage via conditioning noise exposure and vice versa. The mechanisms behind these phenomena are still under investigation. It is possible that milder cochlear stress (i) decreases F-actin to lessen mechanical trauma; (ii) suppresses apoptosis to reduce metabolic damage; and (iii) upregulates antioxidants to minimize oxidative stress. It is more than likely that these mechanisms occur in combination to reduce overall cochlear injury. Additional research in this area must continue to investigate the effects of sound conditioning in detail, as well as the cochlear processes driving the effects. It is also critical to interpret future sound conditioning results in the context of synaptopathy.
Cochlear synaptopathy is defined as noise-induced damage to IHC afferent nerve fiber synapses that produces TTS but not PTS and goes undetected in typical clinical assays of hearing loss. For this reason, synaptopathy has also been referred to as “subclinical hearing loss,” “threshold-independent hearing loss,” or “hidden hearing loss.” If moderate-level noise exposure induces cochlear synaptopathy, the boundary between synaptopathy and sound conditioning must be clearly defined before sound conditioning research can proceed beyond the animal model.
Importantly, the two phenomena may not be independent. In particular, ear-toughening experiments may enhance hair cell survival, but the preserved hair cells may have irreversible synaptic damage. In the case of IHCs, toughening may enhance OHC survival, but IHC synapses may be, and likely are, damaged. The lack of threshold changes alone or the absence of hair cell loss may fail to reveal synaptic damage. Even in cases of synaptic damage, the boundary at which synaptic damage begins to affect auditory function is not well understood. Repeated experiments have shown that synaptopathic noises produce no long-term effects on OHCs, as assessed by DPOAEs (20, 45). However, there is evidence from animal models that these exposures produce permanent reductions in ABR Wave-I amplitudes at higher sound presentation levels, data consistent with loss of afferent low-spontaneous rate fiber function (5, 53).
Histological studies have shown loss of synaptic connections in mice and guinea pig IHCs, with no measurable damage to OHCs (35, 51). Data from these studies have fueled speculation that noise exposures that produce only TTS may produce hearing deficits that are not detected by threshold measures. The potential for suprathreshold or threshold independent hearing deficits have led to a number of human studies (8, 25, 26, 87). The human studies motivated by the animal experiments have yielded mixed results. Individuals with noise exposure histories but no evidence of hearing loss have not reliably been shown to have suprathreshold hearing deficits, such as poorer hearing in noise (49).
To date, one animal study (53) has shown elevated thresholds in competing background noise, but not in quiet, after a synaptopathic noise exposure. These data support the hypothesis of hearing deficits after sound exposures that do not appear to produce PTS. Animal models will likely play a major role in future conditioning studies and the relationship among conditioning noise exposure, synaptopathy, and effects on threshold and suprathreshold measures.
There is a clear need to determine the effect of conditioning or toughening on hearing loss outcomes beyond simply PTS. Suprathreshold measures of the auditory system will be critical evaluators of potential synaptic injuries induced by the conditioning stimuli themselves, or that go unprotected by conditioning paradigms that are effective in reducing PTS. Indeed, future conditioning studies will likely reveal a complex interaction of level duration and pattern of cochlear damage that will enhance our understanding of the complexities of the inner ear and its response to environmental and hazardous challenges.
Footnotes
Authors' Contributions
R.T.H.: Main writer for content on mono-modal sound conditioning and co-writer of “Potential Mechanisms.” J.R.D.: Main writer for content on cross-modal conditioning. M.T.: Co-writer for “Current State of the Literature” and references coordinator. E.C.B.: Co-writer for “Potential Mechanisms” and editing. E.L.: Co-writer for “Current State of the Literature” and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors declare no external funding sources for this review.