
Abstract
Significance:
Among the 200 or so cell types that comprise mammals, spermatozoa have an ambiguous relationship with the reactive oxygen species (ROS) inherent in the consumption of oxygen that supports aerobic metabolism.
Recent Advances:
In this review, we shall see that spermatozoa need the action of ROS to reach their structural and functional maturity, but that due to intrinsic unique characteristics, they are, perhaps more than any other cell type, susceptible to oxidative damage. Recent studies have improved our knowledge of how oxidative damage affects sperm structures and functions. The focus of this review will be on how genetic and epigenetic oxidative alterations to spermatozoa can have dramatic unintended consequences in terms of both the support and the suppression of sperm function.
Critical Issues:
Oxidative stress can have dramatic consequences not only for the spermatozoon itself, but also, and above all, on its primary objective, which is to carry out fertilization and to ensure, in part, that the embryonic development program should lead to a healthy progeny.
Future Directions:
Sperm oxidative DNA damage largely affects the integrity of the paternal genetic material to such an extent that the oocyte may have difficulties in correcting it. Diagnostic and therapeutic actions should be considered more systematically, especially in men with difficulties to conceive. Research is underway to determine whether the epigenetic information carried by spermatozoa is also subject to changes mediated by pro-oxidative situations. Antioxid. Redox Signal. 37, 481–500.
Spermatozoa Ambiguous Relationship with Reactive Oxygen Species
The impact of reactive oxygen species (ROS) on sperm structure and function has been the subject of long-standing debate in basic science and clinical practice. A simple PubMed search with the keywords “ROS” and “spermatozoa” will bring up more than 2000 publications, starting as early as 1946 with the report in Nature that mature spermatozoa were one of the first cell types to produce large amounts of ROS, in particular, hydrogen peroxide (H2O2) (166). This was somewhat to be expected, as this cell type is characterized by a high metabolic rate to support its motility but a limited capacity to counteract the damaging ROS that are inevitably generated as a by-product of aerobic metabolism.
The particular intrinsic susceptibility of spermatozoa to ROS attacks
Due to their extreme cytodifferentiation, spermatozoa have haploid and highly compacted chromatin, which renders them transcriptionally silent and prevents them from mounting genomic antioxidant responses. In addition, the resorption of most of the cytoplasm at the onset of spermiation by the Sertoli cells renders spermatozoa poorly endowed with cytosolic antioxidants, whether these are enzymatic (e.g., superoxide dismutase [SOD], catalase [CAT], glutathione peroxidases [GPxs], glutathione-S-transferases [GSTs], peroxiredoxins [PRDXs], and glutaredoxins [GRXs]) or small molecular mass ROS scavengers (e.g., glutathione [GSH], thioredoxin [TRX]) as well as vitamins (i.e., vitamin E [alpha-tocopherol] and vitamin C [ascorbic acid]) (158). The poor ability to defend against ROS is, therefore, a characteristic of mature spermatozoa, which partly explains their high susceptibility to oxidative attack (7, 43).
In addition to its quasi-“silent” metabolic nature, the unique cytology of the sperm cell also explains its sensitivity to oxidative attack, especially when it comes to oxidative DNA/nuclear damage, a point that will be elaborated on later. In the absence of a significant cytoplasmic compartment, the sperm nucleus, despite its great compactness compared with any somatic cell, is still vulnerable to ROS attack.
A second critical aspect of sperm physiology that accounts for their susceptibility to oxidative damage is the particular composition of their plasma membrane, which has a high content of polyunsaturated fatty acids (PUFAs) (100) that are highly susceptible to ROS-mediated lipid peroxidation (4, 7, 42). When initiated, lipid peroxidation of the sperm plasma membrane enters a vicious cycle in which the generation of toxic aldehydes such as 4-hydroxynonenal (4-HNE) and malondialdehyde (MDA) exacerbate ROS generation by the mitochondria, among other deleterious effects on sperm structure and function (121).
The plasma membrane of mammalian spermatozoa has evolved to generate the membrane fluidity needed to support both motility and sperm/egg membrane fusion during fertilization. One way to achieve these goals was to reduce the amount of cholesterol in the sperm plasma membrane and, at the same time, increase the degree of unsaturated fatty acid. This has led to spermatozoa having a high phospholipid-cholesterol ratio (PL/Chol) and a high PUFA content, making them more susceptible to lipid peroxidation.
Spermatozoa need oxidation processes to mature and function
Mammalian spermatozoa become structurally and functionally capable of fertilization as they migrate through the epididymis (80). Among the various modifications to which spermatozoa are subjected as they transit the epididymis, the oxidation of thiols to disulfides is of particular interest. Elegant reports have shown that disulfides in mammalian sperm proteins are closely involved in two important aspects of sperm physiology, namely maturation of the flagellar structure and chromatin condensation (154).
This discovery was made possible by the use of a fluorescent labeling agent, monobromobimane (mBBr), which specifically alkylates thiol groups (151), allowing the thiol-disulfide status of spermatozoa to be determined with or without dithiothreitol (DTT) reducing treatment. Using flow cytometry (FCM), fluorescence microscopy, gel electrophoresis, and spectrofluorometric quantification, it was shown that the thiol content of spermatozoa decreases sharply as they descend into the epididymis. It has also been shown that disulfide bridge formation involves several distinct protein fractions located in the sperm head, mitochondrial sheath, and tail and that the extent of thiol oxidation to disulfides in these proteins is variable (150, 154).
Originally described in rats, this epididymal oxidation of thiols has been shown to be very similar in other animal models (21), including humans (143). These observations led to the proposal that mBBr labeling of sperm estimated by FCM could be a valuable test for evaluating sperm quality (151). However, when cohorts of infertile men were examined, it was found that, while discriminating between oligozoospermic, teratozoospermic, and normozoospermic samples, the mBBr signal neither correlated with World health Organization (WHO) basic semen parameters nor correlated with fertilization capacity. Further, it bore no relationship to whether the sperm population had been fractionated or not (143, 149). In addition, considerable heterogeneity in the proportion of thiols to disulfides has been reported between ejaculates (149).
Looking more closely at the status of sperm thiol protein, it has been shown more recently that there is a relationship between sperm thiol levels and phosphotyrosine phosphatase (PTP) activity (152). PTP, which is known to be inhibited by oxidation, counteracts the action of tyrosine kinases involved in protein tyrosine phosphorylation, a process closely associated with the acquisition of sperm motility and the events that control sperm fertilizing ability (i.e., capacitation, acrosomal response, and hyperactivation) (11, 98). During epididymal maturation, tyrosine phosphorylation of sperm tail proteins increases significantly. Induced oxidation of sperm thiols with diamide was shown to increase phosphorylation of sperm tail proteins, whereas disulfide reduction with DTT had a negative effect (149).
These observations suggest that PTP activity is somehow correlated with sperm thiol status and that tyrosine phosphorylation of sperm proteins during epididymal maturation is enhanced by thiol oxidation, which is itself associated with lower PTP activity (149). This is an important and largely unconsidered aspect linking the thiol/disulfide status of sperm proteins to protein tyrosine phosphorylation processes that are critical for gamete function. Overall, sperm capacitation can be partly seen as a redox-regulated process (8, 131).
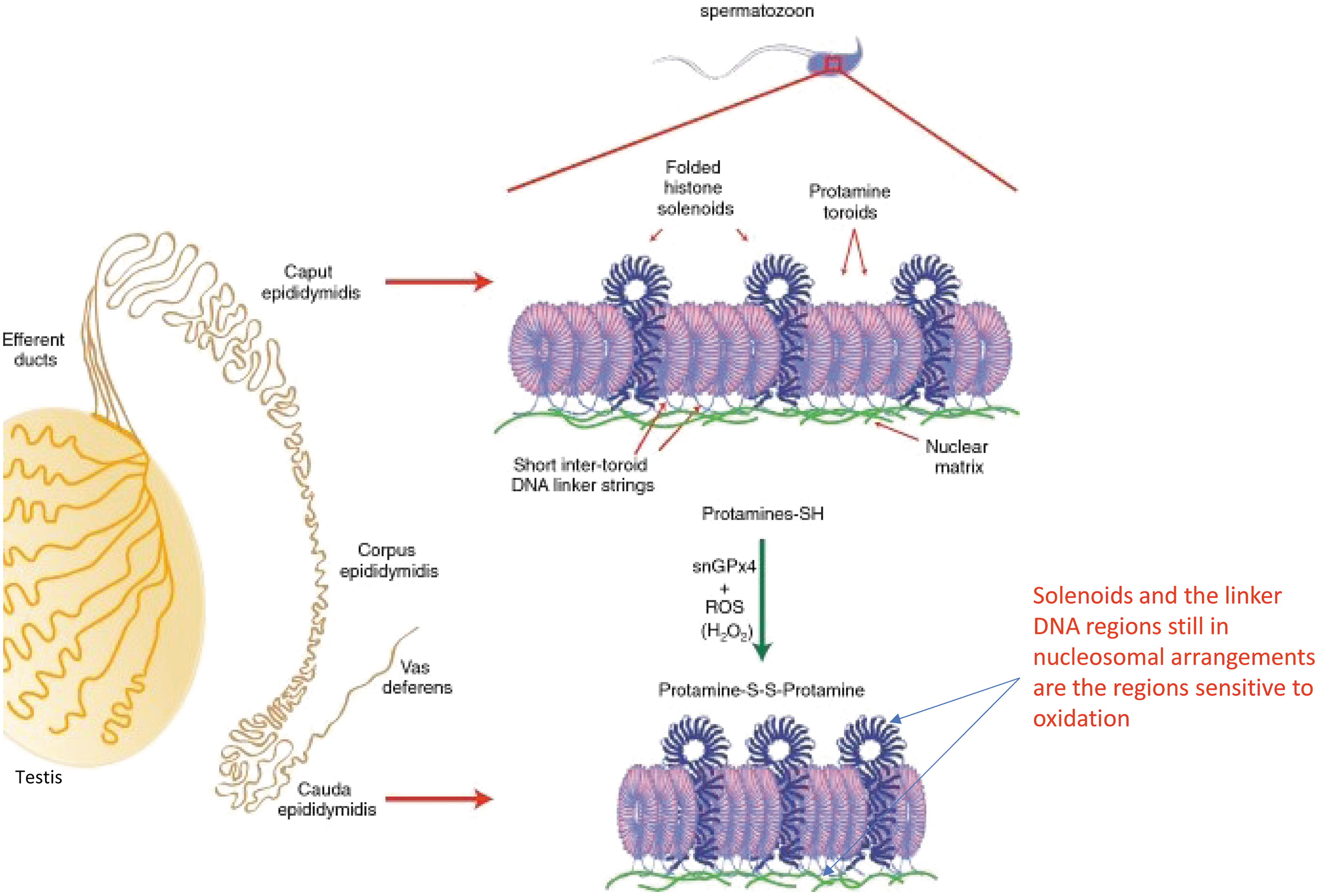
The nucleus is another compartment of spermatozoa where the thiol/disulfide status is particularly important. At the end of spermatogenesis, during spermiogenesis, testicular histones are transiently replaced by transition proteins, which are themselves later replaced by histone H1-related sperm nuclear basic proteins, called protamines (12, 140). The small size and positive charge of protamines, as well as their supramolecular organization with DNA into a toroidal structure, allows for great compaction of the sperm chromatin into the smallest cell type found in mammals.
It is assumed that this serves hydrodynamic purposes as well as protection of paternal DNA. In Eutherian mammals, arginine-rich protamines are also thiol-containing cysteine proteins involved in the establishment of abundant disulfide bridges during epididymal descent, creating intra-protamine and inter-protamine disulfide bonds that have been shown to have distinct roles in maintaining sperm chromatin integrity (75). Using transgenic mouse models, it has been demonstrated that protamine-thiol oxidation during epididymal transit contributes to further condensation and stabilization of the sperm nucleus to achieve a state of optimal compactness.
This has been shown to be orchestrated by the activity of epididymis-secreted antioxidant enzymes (glutathione peroxidase 5 [GPx5], in mice, but PRDXs in humans), which through their activities regulate luminal H2O2 concentration (21, 130). At an optimal concentration, H2O2 serves as a substrate for a sperm nuclear glutathione peroxidase 4 (snGPx4) that functions as a disulfide isomerase (27).
In mice, it has been shown that inactivation of GPx5 activity has dual consequences for the sperm nucleus. Transiently, for spermatozoa in the proximal epididymis, it causes higher disulfide bridging activity due to increased H2O2 availability, resulting in spermatozoa with smaller nuclei compared with spermatozoa from control mice (21). However, when examining sperm cells from the cauda epididymidis of GPx5 knockout (KO) animals, it was shown that the sperm nuclei, though properly compacted, were more sensitive to a reducing challenge, suggesting an inherent instability (21).
In addition, caudal epididymal spermatozoa from KO animals exhibited high levels of DNA base oxidation (21, 91, 127). The generation of double knockout (DKO) animals in which both GPx5 and snGPx4 were inactivated confirmed the role played by H2O2 and sperm nuclear disulfide isomerase activity, as this DKO resulted in a worsening of the sperm phenotype, including greater sensitivity to nuclear decondensation and higher levels of DNA base oxidation (21, 128). With both genetic backgrounds, no other structural or functional alterations in the spermatozoa were found to be significant, apart from the fact that the sperm nucleus tended to decondense and carry many oxidized DNA bases; wild-type (WT) and KO animals had equivalent sperm counts, motility parameters, membrane lipid peroxidation levels, viability, and DNA fragmentation scores (128).
However, despite what appeared to be a mild sperm phenotype, fertilization of oocytes with oxidized DNA sperm was associated with increased miscarriage, aberrant embryonic development, as well as increased perinatal mortality (21). This has led to the conclusion that oxidative stress (OS) within the sperm nucleus, which includes, but is not limited to, DNA base oxidation, may be an important factor in paternally mediated reproductive failure.
Additional KO mouse models have also emphasized the importance of OS in male reproductive function. For example, the importance of SOD in the preservation of normal sperm function has been emphasized by the Sod1-KO mouse (128). In this case, insemination of WT oocytes in vitro with Sod1-KO sperm suppressed fertilization rates compared with WT spermatozoa via mechanisms that could be partially reversed by a reductant (2-mercaptoethanol) (168). Sperm motility was also compromised in these Sod1-KO mice, whereas levels of lipid peroxidation were found to increase (168). Incubation of such spermatozoa in vitro led to a time-dependent decrease in adenosine tri phosphate levels and a concomitant decline in the levels of tyrosine phosphorylation seen in these cells, suggesting the impairment of sperm capacitation.
The lack of SOD-1 has also been shown to accentuate the impact of aging on sperm quality, increasing levels of lipid peroxidation and oxidative DNA damage in these cells as a function of age (124). Interestingly, the deletion of catalase does not have the same impact, suggesting that this is not a major protective enzyme as far as the male germ line is concerned and/or that compensatory pathways are upregulated in such animals, that effectively neutralize the OS created by catalase deficiency (153). The PRDX system seems to be particularly important in this regard (153), although even the upregulation of this system cannot prevent the OS created in the epididymis in the GPx5/snGPx4 DKO mouse (128).
Interestingly, in the reverse situation where both the testis-specific and somatic forms of peroxiredoxin 4 (Prdx4t/Prdx4) are deleted, fertility is normal, at least, in part because GPx4 levels show a compensatory increase under these conditions (128). The impression given by these KO models is that the control of OS is so important for male reproduction that multiple defensive pathways have developed to protect the germ line. Moreover, if one element of this defective system should fail to function, others will compensate to ensure that a high level of protection is provided over a broad range of circumstances.
These different detrimental and beneficial actions of ROS on sperm structure and function are summarized in Figure 1.

Damage Caused by ROS to Genetic and Epigenetic Information of Spermatozoa
Localization of nuclear oxidative damage in spermatozoa
Because sperm DNA oxidation appears to be an important player in reproductive failure, a more detailed study was conducted to determine where and to what extent paternal DNA might be damaged. First performed on mouse sperm nuclei, immunocytochemistry revealed that oxidation of the sperm nucleus occurred at preferential locations characterized by high persistent histone content (27). In mice, these locations correspond to the basal region of the sperm head (neck) as well as the dorsal peripheral nuclear zone (22, 127) (Figs. 2 and 3).
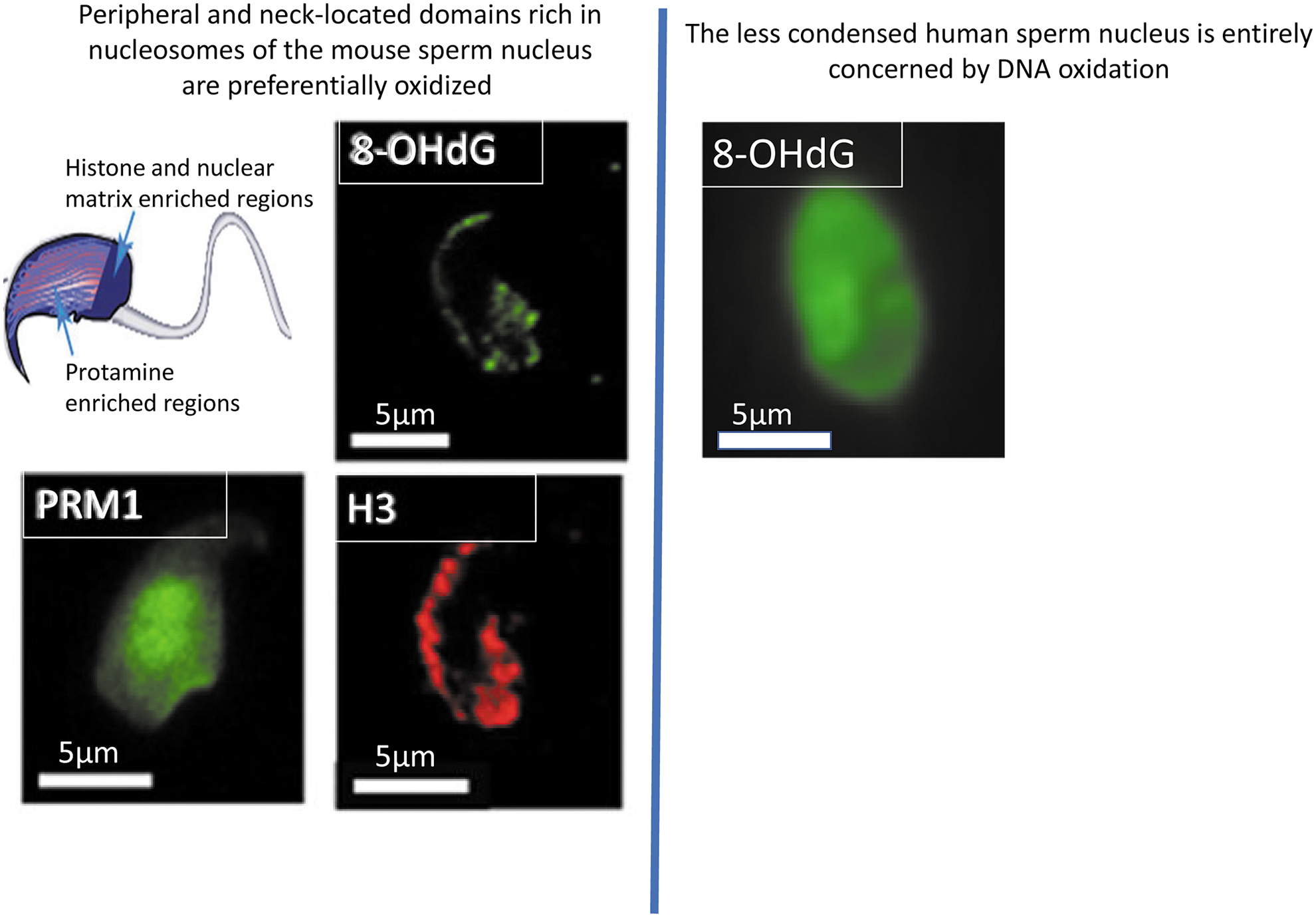
This made some sense, as these regions retain a nucleosomal organization with DNA wrapped around histone octamers as in somatic cells, generating lower compaction with only 146 bp of DNA per octamer (169, 183). When the oxidized chromosomal regions of paternal DNA were further characterized (91), we observed that DNA wrapped in nucleosomes containing the histone variant histone 2A variant Z type (H2AZ) was preferentially the most susceptible to oxidative damage. This also made sense because H2AZ-containing nucleosomes are less condensed than canonical histone-containing nucleosomes, which facilitates DNA repair (101).
In protamine-associated sperm nuclear regions, toroidal organization condenses 50–100 kb of DNA to a near-crystalline state (13, 28, 179), limiting access to stressors, including ROS (127). In the human sperm, nucleus histones are about 10 times more represented than in the mouse and, as a result, oxidative DNA damage is not regionalized within the human sperm nucleus but involves the entire sperm head (170).
Sensitivity of sperm chromosomes to oxidative damage
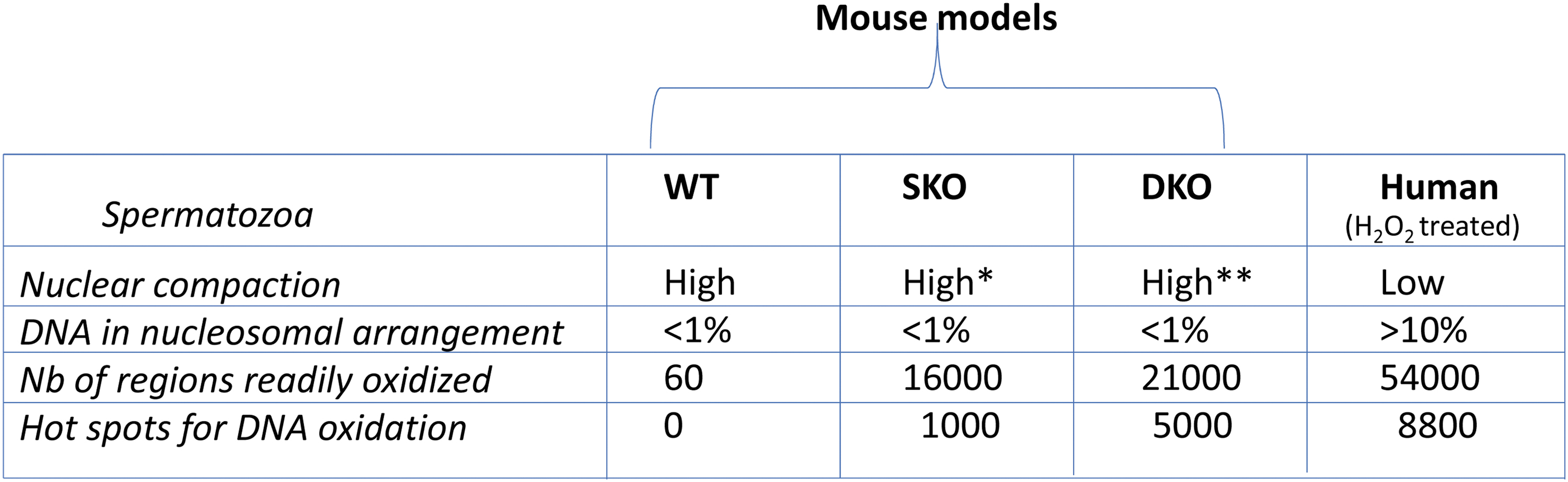
Using a modified chromatin immunoprecipitation protocol coupled with next-generation sequencing, each base oxidation-sensitive chromosomal region of spermatozoa was identified. This was first conducted in the mouse KO models described earlier (91) and then in human sperm cells (180). In mice, these experiments revealed that, despite the mild epididymal OS created in KO animals, which showed no obvious phenotype apart from the higher level of sperm DNA oxidation, the number of oxidized regions on the sperm nucleus increased to 16,000 regions in gpx5 KO animals (single KO [SKO] mouse) and to more than 21,000 regions in sngpx4;gpx5 KO animals (DKO mouse) whereas it was less than 60 regions in the control WT mouse (91) (Fig. 4).
Interestingly, the distribution of these oxidized regions on mouse sperm chromosomes was found to obey a certain level of rhythmicity as they were found approximately every 50 kb (91). This information, coupled with the fact that oxidation of sperm DNA preferentially targets histone-rich domains, highlighted the small DNA linker domains (∼1 kb long and known for their nucleosomal organization) connecting each protamine torus that envelop 50 kb of DNA, as the sperm DNA sites most susceptible to oxidative damage (91) (Fig. 3).
Interestingly, these regions correspond to DNA domains attached to the sperm nuclear matrix that participates in the highly ordered organization of sperm chromatin in the nucleus (127, 174), which also explained the sensitivity of the nuclear peripheral regions to oxidation.
Not all chromosomal regions identified in mouse sperm were oxidized to the same extent, as a subset of ∼1000 regions in SKO animals and a subset of nearly 5000 regions in DKO animals were found to be highly oxidized and were defined as hotspots for DNA oxidation (91). The 5-fold increase in the number of DNA oxidation hotspots in DKO sperm nucleus compared with SKOs illustrates the importance of optimal nuclear condensation for protecting paternal DNA from oxidative damage (128). Regarding the distribution of these oxidized regions on mouse sperm chromosomes, it is interesting to note that the smaller the chromosomes, the more sensitive they were to oxidative damage, exhibiting a higher number of oxidized regions (91).
This was not related to the fact that smaller chromosomes contain more persistent histones than longer ones, as histones were shown to be evenly distributed among all autosomes (91). Only sex chromosomes in the mouse sperm nucleus appeared to contain more nucleosomes, which may partially explain their greater sensitivity to oxidative damage than autosomes (91).
Because histone/nucleosome density could not explain why some chromosomes were more sensitive to oxidative damage than others, it was hypothesized that chromosome location within the sperm nucleus may be important. Fluorescent in situ hybridization experiments coupled with immunocytochemistry revealed that in the mouse sperm nucleus, small chromosomes were located closer to the neck region and were also more peripheral than long chromosomes (127). These locations corresponded well with the sperm nuclear domains that were affected by oxidative damage (21, 127). Thus, it is the position of chromosomes in the mouse sperm nucleus that determines their sensitivity to oxidation (91).
This was confirmed by a further study using confocal immunocytochemistry coupled with three-dimensional image reconstruction that identified specific chromosomal territories in the mouse sperm nucleus (185). These investigations were carried out in parallel on the different genetic backgrounds (WT, SKO, DKO) and revealed that oxidative damage affects the integrity of paternal DNA without altering the topology of chromosomes within the nucleus (185). Alterations of chromosome territories could have been possible, because inter-chromosome protamine disulfide bridges are suspected to maintain chromatin organization in the sperm nucleus (66, 176, 185).
These investigations also revealed that SKO sperm nuclei showing higher oxidative damage were more compacted than WT sperm nuclei, while still being more sensitive to reducing agent-mediated decondensation. This confirmed that epididymal maturation of the sperm nucleus uses a very delicate balance in which ROS, on the one hand, ensure optimal compaction of the sperm nucleus and, on the other hand, promote its decondensation, most likely through the DNA breaks they induce (185).
In human spermatozoa, the observations and conclusions were very similar and consistent with the intrinsically lower level of compaction in the human sperm nucleus, which is supported by the fact that 10%–15% of human sperm DNA is still associated with histones. As many as 54,000 regions sensitive to DNA oxidation have been identified on human sperm chromosomes, including ∼8800 regions labeled as “highly significantly” oxidized (180) (Fig. 4). As in mice, oxidized chromosomal regions were found at regular intervals of ∼50 kb, again designating the DNA binding domains connecting two adjacent protamine toroids as the target for oxidative damage.
In contrast to observations in mice, the most oxidation-sensitive regions of human sperm nucleus were domains lacking nuclear-organizing proteins, either histones and/or protamines (180). Further, in contrast to the situation in the murine sperm nucleus, the number of oxidized regions on human sperm chromosomes followed a linear relationship with chromosome size. The longer the chromosome, the more oxidized regions it contained (180). This could have been expected from the immunocytochemical data showing in humans that the entire sperm nucleus is involved in oxidation in contrast to the mouse sperm nucleus where DNA oxidation is limited to a few specific areas (170, 180).
It is noteworthy that three chromosomes escape this linear relationship, namely, the two sex chromosomes that were found to be particularly well protected from oxidative damage in human spermatozoa; on the contrary, autosome 15 was found to be very sensitive. Similar to the mouse sperm chromosomes, the position of these chromosomes in the sperm nucleus partly explains these observations since chr15 is located at the periphery of the nucleus whereas the allosomes occupy a more central position (118, 184) (Fig. 5).

Ten chromosomal regions highly susceptible to DNA oxidation have been identified in the human sperm nucleus and it was remarkable to find that these regions were associated with sites where mutations are known to lead to the development of syndromes in the offspring, which have been shown to be associated with paternal age (30, 46, 109) and increased paternal DNA oxidative damage (25). These include Prader-Willi syndrome and Angelman syndrome, autism spectrum disorders, abnormal developmental situations and multiple congenital anomalies in offspring, as well as a predisposition to autism, bipolar disorder, and schizophrenia (6, 14, 17, 18, 77, 78, 85, 90, 117, 119, 145, 171). It was also very interesting to find that when comparing a cohort of fertile men with an infertile cohort, most of these 10 sperm DNA oxidation susceptibility regions were more oxidized in the infertile cohort (180).
This corroborated a recent follow-up of men in couples consulting for difficulty conceiving, which showed that 7 out of 10 had moderate to high levels of sperm DNA oxidation (170), further suggesting that sperm DNA oxidation is a very common occurrence in the clinic. It is also consistent with studies demonstrating a significant relationship between sperm DNA damage (measured with the sperm chromatin structure assay [SCSA®]) and pregnancy outcome in an IVF program (16, 54). Correlations have also been observed between levels of sperm DNA oxidation and defective sperm or/and seminal parameters that have long been known to be associated with OSs (170).
This was particularly the case for leukocytospermic and asthenozoospermic samples, as well as sperm samples from obese male patients for whom OS was found to be the main factor contributing to their infertility (170).
Genetic/epigenetic risks as consequences of oxidative damage to the sperm nucleus
Genetic risks: promotion and transmission of de novo mutations
Any aerobic cell is at risk of oxidative DNA damage depending on the intensity of OS it faces. At moderate levels of OS, base adducts, such as 7,8-dihydro-8-oxo-2′deoxyguanosine (8-oxodG), are formed. Guanine is the most sensitive base, due to its low redox potential compared with other bases, adenosine is also very sensitive to base oxidation, and thymine and cytosine are less susceptible to oxidative attack (87). When OS increases, base oxidation can be associated with the generation of abasic sites (AP) that are often likely to generate DNA breaks (15).
When OS increases further, it may cause covalent bridging events in nuclear proteins that could affect chromatin organization and plasticity (71). Finally, in situations of acute OS, DNA strand breaks can be recorded, either single-strand breaks (SSBs) or double-strand breaks (DSBs) leading to DNA fragmentation (102, 144). Spermatozoa exhibit all these forms of DNA damage, including fragmentation, which has been clearly linked with reproductive failures in both natural conception and assisted reproduction (113, 159, 164, 181).
However, sperm DNA fragmentation may also have other origins, including unresolved meiotic breaks, apoptotic germ cells incompletely discharged, and sperm DNA breaks resulting from mechanical shear after protamination during spermiogenesis (5). Therefore, monitoring sperm DNA fragmentation by any of the techniques available to date (but not yet routinely used in the clinic to assess sperm nuclear integrity!), including deoxynucleotidyl transferase dUTP nick end labeling (TUNEL), SCSA, Comet, and sperm chromatin dispersion, cannot give an accurate and complete indication of sperm DNA oxidation status.
Somatic cells repair their DNA throughout the cell cycle by using corrective enzyme systems, one of which is the base excision repair (BER) pathway. BER is important for removing chemically damaged bases, whether oxidized, alkylated, or deaminated, that could otherwise cause mismatch mutations or lead to DNA breaks during replication. BER uses DNA glycosylases such as 8-oxo-guanine DNA glycosylase (OGG1) (10, 89) in the case of oxidized base removal. Glycosylases recognize damaged bases and generate abasic sites (AP).
These are then cleaved by an AP-endonuclease (apurinic/apyrimidinic endonuclease 1 [APE1]), resulting in single-strand breaks that can be processed by polymerases to replace missing nucleotides (45, 65, 105). Due to their peculiar and extreme cytodifferentiation and metabolism, sperm cells do not have a functional BER system. They have been shown to possess only the first step consisting of OGG1-mediated processing of oxidized bases, but they do not contain the second enzymatic activity (APE1) that prepares AP sites for the insertion of a new base (160, 161). It is only during the early postfertilization stages, before paternal DNA replication that precedes the first segmentation division, that the BER machinery of the oocyte is involved in repairing AP sites in the paternal pronuclei.
This shared process of DNA repair, half of which is performed by the sperm and half by the oocyte, highlights the importance of both sperm and egg quality for reproductive success (107). Unfortunately, this otherwise laudable example of male–female collaboration is marred by the oocyte being relatively deficient in OGG1. So, if a spermatozoon with highly oxidized DNA fertilizes an oocyte, the 8-oxodG residues may remain unresolved and persist into the round of DNA replication that precedes the first mitotic division.
Further if, for any reason, the BER machinery of the oocyte is not optimal (whether due to the age of the mother or due to any other extrinsic factor, such as the hormonal induction of oocyte maturation) this can further compromise the ability of the oocyte to effectively repair the DNA damage brought into the egg by the fertilizing spermatozoon. In situations such as advanced parental age or infertility, it would be possible to imagine a degree of collusion between the male and female germ lines that significantly compromises the developmental potential of the embryo.
Under these circumstances, the clinical impact of excessive oxidative DNA damage in spermatozoa will become accentuated by the impaired capacity of the oocyte for DNA repair. As a result of such factors, patients may be directed straight to invasive assisted reproductive technologies (ART) such as ICSI, without adequate assessment and, as a result, experience potentially preventable failed IVF attempts (36, 62, 134).
What, then, will be the impact of incomplete or defective repair of oxidized DNA bases in the germ line? There are several consequences of such a situation that are not mutually exclusive. First, an excess of AP sites in the paternal pronucleus will favor SSBs and increase the time required for repair, since the efficiency with which BER processes SSBs is four to five times lower than with AP sites (122). Further, as we have found that sperm DNA base oxidation occurs preferentially in clusters at specific chromosomal locations (91, 180), this opens up the possibility of having, within these clusters, AP sites in close proximity to 8-oxodG lesions. It has been reported that damage in bistranded clusters associating both AP sites and 8-oxodG lesions is more likely to terminate in DSBs, resulting in increased mutagenic potential compared with isolated lesions (106).
It has also been shown elsewhere that in a cluster containing an AP site or SSB, DNA repair activity is greatly reduced when 8-oxodG residues are in close proximity. It is hypothesized that 8-oxodG lesions in the vicinity of SSB and AP sites introduce conformational destabilization of DNA that prevents proper interaction of the repair machinery (96, 97). Therefore, one consequence of the presence of clusters of 8-oxodG residues on sperm chromosomes next to AP sites and SSBs will be to delay oocyte-driven DNA repair of the paternal nucleus and potentially increase the occurrence of mutagenic DSBs that are more difficult to repair (74, 163).
The second consequence of having a paternal chromosomal complement with an excess of 8-oxodG residues will be the risk that some of these oxidized residues completely escape the shared sperm/oocyte repair process. If this happens, before the first cleavage division it may lead to de novo mutations (DNM). One mechansism by which this occurs involves Hoogsteen base pairing whereby 8-oxodG is mistakingly seen as a thymine (106) leading to the aberrant incorporation of an adenine (74). These events underlie the so-called G:C to T:A “transversion” mutations (163) that are believed to be primarily paternally inherited and to increase with paternal age (133, 182).
The increase in the occurrence of transversion mutations in the zygote is of concern because these mutations, which are known to have a greater impact on gene regulation than transitions (60), will be passed on to every cell in the offspring (115). Since point mutations (such as transversions and transitions) are highly pathogenic (31) and largely arise in the paternal line, it is of interest to know the level of sperm DNA oxidation, since this may define the susceptibility of the embryo to de novo point mutations. This is particularly relevant in the context of assisted reproductive procedures where it is further suspected that the techniques themselves may contribute to increased DNM (92).
Epigenetic risks: OS and alteration of epigenetic information in spermatozoa
In addition to base oxidations and its associated genetic consequences, OS is also likely to affect the epigenetic information carried by spermatozoa, thus disrupting the messages that these epigenetic marks carry.
In addition to the classical epigenetic marks represented by methylation of cytosine residues and various post-translational modifications involving nuclear histones (methylation, acetylation, and many others), spermatozoa have been shown to contain a complex and highly dynamic pool of small and long noncoding RNAs (ncRNAs) that are suspected to modulate the developmental program of the embryo after fertilization (79).
To our knowledge, oxidation of sperm nuclear proteins has not been studied outside of its structural impact on sperm nuclear protamines and its potential consequences on the size of the sperm head as previously mentioned (22). Although this may impact sperm motility and the kinetics of sperm nuclear decondensation after fertilization, it is unlikely to affect the early developmental program of the embryo, as protamines will be rapidly cleared from the zygote. Persistent histones in spermatozoa, however, may be better candidates for introducing altered epigenetic marks associated with the so-called “histone code” in the embryo if they are affected by OS.
A recent analysis of DNA methylation and histone H3 lysine 4 tri-methylation (H3K4me3) in human spermatozoa revealed considerable overlap, suggesting a potential interplay between these marks that might be mutually exclusive in sperm. Indeed, comparisons between H3K4me3 marked regions in sperm and the embryonic transcriptome suggest a possible paternal influence on embryonic gene expression (99). The etiology of dynamic changes in histone methylation will have to be resolved in future studies; however, there are already data to indicate that the induction of OS in spermatogonial cell lines results in changes in histone methylation status that could potentially impact embryonic development (120).
Alteration of nuclear cytosine methylation of the sperm nucleus is a powerful route by which OS could alter one of the important epigenetic marks associated with paternal DNA. There are several levels to consider. First, under acute OS during spermatogenesis, when paternal cytosine methylation occurs (132), the presence of 8-oxodG residues in CpG doublets has been reported to interfere with the methylation process of the adjacent cytosine residue (173, 179). It is likely that this may lead to changes in the pattern of paternally imprinted regions.
Second, as recent research has established that methylation of post-testicular sperm DNA is unlikely (48), the way in which OS may affect sperm DNA 5 methylcytosine (5meC) residues during post-testicular maturation is, therefore, limited to the oxidation of 5meC, leading to 5 hydroxymethylcytosine (5hmeC) residues. This is particularly interesting and may potentially have important consequences, as 5hmeC is increasingly considered to be a particularly important epigenetic mark for male germ cells in itself (50).
In support of this, we recently observed that in the context of the SKO mouse strain with epididymal sperm nucleus oxidation, the levels of 5meC and 5hmeC on sperm DNA were altered (M. Gentil and E. Hug, unpublished data). The increase in 5hmeC representation in sperm DNA could initiate demethylation of paternal DNA regions, as 5hmeC generation is the first step in the ten eleven translocase (TET)-dependent oxidative process that replaces meC with cytosine after fertilization (93, 111).
In this process, there is massive demethylation of paternal DNA (173) but specific male pronucleus regions must escape this demethylation, particularly DNA regions that are important for transposon silencing and DNA regions of paternally imprinted genes (64). It is conceivable that if there is an excess of 5hmeC on the paternal oxidized nucleus, the regions involved could be erroneously committed to demethylation. Hypomethylation of the sperm genome in a 5,10-methylenetetrahydrofolate reductase deficiency model has been shown to generate a sub-fertility phenotype that extends across at least two generations (86).
Thus, oxidative damage of the sperm nucleus could alter the dynamics of epigenome reprogramming and possibly affect the developing embryo and the fertility of future generations (142). This issue will, undoubtedly, need to be investigated in more depth in the future.
Let us now turn to the spermatozoon's cargo of ncRNAs and their susceptibility to OS. Here again, several issues can be considered. First of all, as is the case for DNA, RNA can be oxidized, leading, for example, to the formation of 8-hydroxygunaosine residues (8-OHG or 8-oxoG). It has been reported that RNA is actually more vulnerable to oxidative damage than DNA (95). The oxidation of messenger RNA has been fairly well studied and has been shown to disrupt translation and protein synthesis (95). It is suggested to be an important contributor to cell dysfunction, cell death, and pathogenesis (55).
To our knowledge, the consequences of sperm ncRNA oxidation have never been studied. Because ncRNAs are not translated, oxidative damage to ncRNAs should not have the same impact as oxidation of mRNAs. In fact, there is compelling evidence suggesting that ncRNAs function as intermediaries in OS responses, as different classes of ncRNAs are up- or downregulated in response to OS (49) and, in turn, regulate the antioxidant genetic response. This is exactly what we found in the epididymal compartment of the SKO mouse strain (24).
In short, comparative RNAseq analyses of the proximal epididymis of WT and SKO mice showed clear changes in epididymal ncRNA profiles that could be correlated with the tissue antioxidant gene response (24). As it is now well accepted that the epididymis supplies transiting spermatozoa with many ncRNAs, it is likely that in response to OS, the ncRNA load of spermatozoa also changes. In fact, we have preliminary data showing that this is the case and that some microRNA (miR) and piwiRNA (PiR) ncRNAs accumulate differently in SKO spermatozoa compared with WT spermatozoa (C. Chu and A. Moazamian, unpublished data).
We hypothesize that this specific sperm ncRNA signature may serve as a signal to stimulate/regulate the oocyte DNA repair machinery. Whether these changes in sperm ncRNA content in response to post-testicular OS will have an impact on the developing embryo and beyond is not yet known. However, it is a reasonable assumption, given that changes in sperm ncRNA in response to specific environmental stresses, including unbalanced paternal nutrition or behavioral stress, are responsible for the transgenerational transmission of phenotypic change (20, 23, 24, 47, 51, 56, 88, 155, 156).
Figure 6 provides a summary of the major genetic and epigenetic problems that could be faced with oxidatively stressed spermatozoa.

The Most Common Exposures of Sperm to OS
During the long journey from its genesis in the testicular germinal epithelium to the site of fertilization in the female genital tract, the spermatozoon may be subjected to oxidative attack. This is particularly significant during post-testicular life, when spermatozoa are heavily dependent on antioxidants in their extracellular environment. As soon as this redox balance is challenged, by either intrinsic or extrinsic causes (i.e., individual genetic factors and/or environmental stressors), spermatozoa are at risk of OS.
Apart from situations such as varicocele and acute orchitis, which are known to generate high testicular OS (2, 63), spermatozoa are more susceptible to oxidative damage in the epididymal compartment than in the testis. This hypothesis is supported by the fact that the seminiferous tubule is a sealed compartment that limits its access to systemic stressors as well as to innate immune cells, which are classical producers of ROS in inflammatory situations. It is also supported by the fact that apoptotic processes during spermatogenesis remove ROS-damaged sperm before they leave the gonad (157), thus preventing them from entering the post-testicular maturation stage.
In addition, repair processes, particularly DNA repair processes, exist in the early stages of spermatogenesis, before spermiogenesis, which can correct some of the DNA damage caused by ROS to the male germ line (52, 59).
In the epididymis, the situation is quite different. Recent research has definitely shown that, unlike the seminiferous tubule, the luminal compartment of the epididymis is not well isolated from systemic influences (172). First of all, the epididymal tubules are densely irrigated by fenestrated blood capillaries (1). Second, the epididymal epithelium is a classical permeable epithelium with conventional tight junctions (TJs) located at the apical-lateral site, very different from the basally located and specialized TJs that seal the Sertoli cells (32, 33, 67).
This means that any environmental stressors that are systemically expressed, such as the low-grade inflammation associated with obesity or metabolic syndrome (38, 39), will have no difficulty reaching the epididymal lumen and attacking the spermatozoa. Also, a wide array of viruses and bacteria are likely to reach the epididymis (e.g., Zika, human immunodeficiency virus [HIV], Herpes-simplex, possibly SARSCOV2 and Chlamydia), inducing a pro-inflammatory state and creating OS (19, 165, 186). The cauda epididymidis is particularly well equipped to cope with such ascending infections, eliciting a powerful inflammatory response when challenged with live bacteria or other inflammatory stimuli (139).
Autoantigenic responses to sperm antigens also tend to be focused on the cauda epididymidis, acutely inducing inflammatory mediators and cytokines/chemokines that could induce OS. The immunological responsiveness of the cauda epididymidis may reflect its role in aggressively combating ascending infections through a robust inflammatory response, whereas the caput provides a more tolerogenic environment to protect the auto-antigenic spermatozoa released from the testis (177). Third, the fact that the blood-epididymis barrier (BEB) is very different from the blood-testis barrier (BTB) also explains the ease with which immune cells reach the epididymal luminal compartment and spermatozoa, which explains the very common clinical conditions of epididymitis (112) and leukocytospermia (68, 178), which compromise male reproductive success, in part by ROS-induced sperm damage.
Exposures to environmental toxicants, whether from food or air quality, as well as exposures to physical environmental stressors such as radiation and heat are thus classical creators of OS that spermatozoa could potentially face (40, 70, 72, 73). The high diversity and abundance of environmental factors that are capable of generating systemic or/and local OS in the vicinity of spermatozoa may explain why sperm DNA oxidation (an early marker of oxidative damage to sperm cells) was found to affect two-third of men from couples with difficulties conceiving regardless of the origin of the couple's infertility (170).
In comparison, when similar male patients were evaluated for sperm DNA fragmentation, only one in seven had what is considered a pathological DNA fragmentation index (DFI >30) (54). These observations highlight the relative importance of sperm DNA oxidation in a clinical context.
In addition to the earlier mentioned environmental factors that induce OS damage in spermatozoa, the contribution of ART must also be considered (40, 70, 72, 73). The preparation of spermatozoa for in vitro fertilization (IVF) or intracytoplasmic sperm injection (ICSI), through either the use of density gradient centrifugation or any sperm selection protocol/device/medium available to date, are all likely to place spermatozoa into a state of OS (3, 57, 104, 136, 137). Sperm cryopreservation, an established technique in assisted reproduction, is also known to be associated with oxidative damage to male gametes (136).
As another very trivial example, natural conception never exposes sperm to light, which is suspected of generating photooxidative stress (26). The ART protocols do expose sperm to light and in some cases, such as when using high-magnification selection methods (motile sperme organelle morphology examination/intracytoplasmic morphologically selected sperm injection [MSOME/IMSI] techniques) to choose the most morphologically perfect cell for ICSI (9), exposure to the microscope's light beam could result in DNA damage.
If this was the case, it may lead some clinics to offer their patients a therapeutic advantage by selecting the highest quality sperm, when, in fact, they may have actually damaged sperm DNA by this approach to sperm selection and compromised the chances of success. The validity of such speculations will have to be examined in future studies.
Importance of Spermatozoa Oxidative Damage Detection in the Clinic
Although it has long been known that OS alters sperm structure and function, this diagnostic criterion is still not routinely assessed to determine the reproductive competence of the male partner in infertile couples. For more than 40 years, reproductive endocrinologists, gynecologists, and other IVF specialists have mostly relied solely on the basic, fundamental WHO recommendations for the evaluation and processing of human semen samples (29, 175).
These recommendations refer to sperm count and morphology, two parameters for which there is nowadays a consensus that they are poorly correlated with reproductive success (37, 94, 103). Sperm motility is occasionally discussed, but with the advent of ICSI, which now accounts for more than 60% of ART attempts worldwide (61), sperm motility tends to be considered a secondary parameter. Although over the past decade the major European and American societies covering the field of clinical infertility (i.e., European Society for Human Reproduction and Embryo [ESHRE] and American Society for Reproductive Medicine [ASRM]) have begun to raise awareness of the importance of sperm nuclear integrity as a key determinant of reproductive success and offspring health, this has not yet translated into clear recommendations for routine andrological assessment.
This is not due to a lack of testing techniques or knowledge, as this information has been widespread for decades. Rather, it is due to the fact that several tests are available to assess different aspects of sperm nuclear integrity such as condensation, fragmentation, and, more recently, oxidation, each with “pros” and “cons,” creating great confusion and preventing their inclusion in the infertility clinicians' arsenal. It is only very recently that excessive sperm DNA fragmentation has been recognized as a critical parameter determining the success of ART (141).
However, very few clinics have made monitoring sperm DNA fragmentation a mandatory step in their male infertility workup. As for sperm DNA oxidation, which should be dissociated from sperm DNA fragmentation, we have seen earlier how common it is and how detrimental it could be to reproductive success and offspring health; however, no clinic in the world has chosen to assess it routinely. Reliable and reproducible protocols are available, but they are still difficult to implement in the clinic due to practicality and cost. This is unfortunate because if they were realized, they could provide clinicians with sound arguments for the appropriate use of therapeutic options that could potentially avoid the overuse of ART.
Antioxidant therapy
Because oxidative problems have long been suspected of altering the structure and function of male gametes, antioxidant supplementation (138) has logically emerged as a means to potentially correct sperm defects. The benefits of oral antioxidant supplementation for the male and/or female partner of an infertile couple are the subject of much controversy. This is mainly due to the fact that to date, no carefully considered, well-designed clinical trials have been conducted. On the contrary, there is a multitude of poorly designed clinical trials due to many factors, including: (i) aberrant patient selection criteria; (ii) small patient cohorts; (iii) arbitrarily and inadequately chosen doses and duration of supplementation; (iv) the great heterogeneity of single molecules or complex formulations that have been used without clear justification for their composition and use; and (v) the evaluation criteria that have been monitored, such as sperm parameters and/or the benefit in terms of reproductive success. Moreover, the impact that antioxidants (AO) have on the levels of OS experienced by the spermatozoa has not been assessed. The literature on this particular subject is, therefore, extensive but very confusing (53).
Several studies examining the effects of AO treatments on sperm quality, embryo quality, and live birth rate after ICSI have obtained conflicting results (82, 148, 162). In a randomized controlled trial of 200 infertility patients, Joseph et al. (162) could find no impact of antioxidant treatment (vitamin C, vitamin E, and zinc) on the incidence of clinical pregnancy, live birth rates, or semen quality. Similarly, in a double-blind, randomized, placebo-controlled trial of antioxidant therapy (vitamin E, vitamin C, selenium, L-carnitine, zinc, folic acid, and lycopene) in 174 male infertility patients, Steiner et al. (161), failed to find a significant change in DNA fragmentation, semen quality, or live birth rates as a consequence of treatment.
Schisterman et al. (147) conducted a large randomized controlled study of antioxidant (zinc and folic acid) treatment for male infertility patients and also failed to find any significant impact on semen quality, DNA integrity, or live birth rate. Matorras et al. (110) studied the impact of vitamin E therapy in a double-blind, placebo-controlled, randomized study of 101 patients but did find a significant increase in life birth rate per transfer as a result of antioxidant therapy, although no impact on clinical pregnancy rate or semen quality was discernable (148). In contrast, Scaruffi et al. (146) conducted a multicenter, longitudinal, uncontrolled prospective study of antioxidant (myo-inositol, alpha-lipoic acid, folic acid, coenzyme Q10, zinc, selenium, and vitamins B2, B6, and B12) therapy in men who had exhibited low fertilization rates in a previous ICSI cycle.
In this instance, antioxidant treatment improved semen quality, decreased sperm DNA damage, and improved both embryo quality and live birth rate (147). Similarly, Nazari et al. (123) looked at the use of antioxidant therapy (vitamin E and zinc) to treat the male partners of couples suffering from recurrent early pregnancy loss. Treatment of 60 patients with this antioxidant formulation revealed a statistically significant improvement in semen quality, including a reduction in levels of DNA damage. Success has also been achieved by Nikitaras et al. (125), who used the co-enzyme Q analogue, Idebenone, to treat the OS experienced by aged human and mouse spermatozoa.
In vitro, this compound reduced ROS generation whereas, in vivo, the administration of Idebenone to aging mice was found to improve fertilization success, enhance blastocyst formation, and increase implantation rates after embryo transfer. The commercial antioxidant preparation, “Menevit,” has also recently been found to generate superior live birth rates after IVF in a retrospective cohort study (167).
After reading the earlier paragraphs and understanding the dual impact of oxidation on spermatozoa, which is both necessary for optimal sperm function and harmful in many ways when in excess, it is clear that indiscriminate AO supplementation is highly likely to be ineffective. Indeed, there are data to indicate that the over-aggressive use of antioxidants in patients who are not suffering from OS may have adverse impacts on spermatozoa by, for example, stimulating an increase in sperm nuclear decondensation (41, 69, 116).
Only when a properly designed clinical trial is conducted will we be able to verify the beneficial impact of this treatment option. This means that the initial selection of patients must be done by examining the level of oxidative damage in the spermatozoa. Since the earliest mark of oxidative damage is DNA base oxidation, this means that assessment of sperm DNA oxidation should be the pathway to select candidates who could benefit from AO supplementation. Further, the level of sperm DNA oxidation should be taken into account, as it will determine the dose and duration of AO supplementation to be considered.
Other key diagnostic tests might be used to confirm a state of OS, for instance measuring ROS by chemiluminescence or FCM or possibly employing other OS markers such as the lipid peroxidation products, 4-HNE and MDA, or measuring the oxido-reductive potential in semen. Finally, one must be very careful in selecting an effective AO, because very few formulations have been well designed.
ROS and Spermatozoa: Evolutionary Perspectives
One of the main purposes of this review has been to highlight the dual effect of oxidative processes on male gametes, as it is clear that sperm cells both require an oxidizing environment to mature and function while, simultaneously, being particularly sensitive to oxidative and reductive stress (135). One may wonder how the evolution of aerobic metazoans could have placed mammalian spermatozoa in such a schizophrenic and potentially dangerous situation. In these final paragraphs, we would like to look at things a little differently, escaping the traditional pessimistic approach that OS is only a risk to spermatozoa.
This line of thinking was stimulated on reading that in a transcriptionally active cell, the 8-oxodG residues located at certain G-rich gene promoters might be epigenetic marks in their own right, which are capable of stimulating transcription of the genes involved (44). Although this concept does not apply to the sperm cell, which is transcriptionally inactive, it does open up the possibility that DNA oxidation might be a powerful signaling event as well as a biomarker of pathology (see Fig. 7 for a summary of the points developed later).
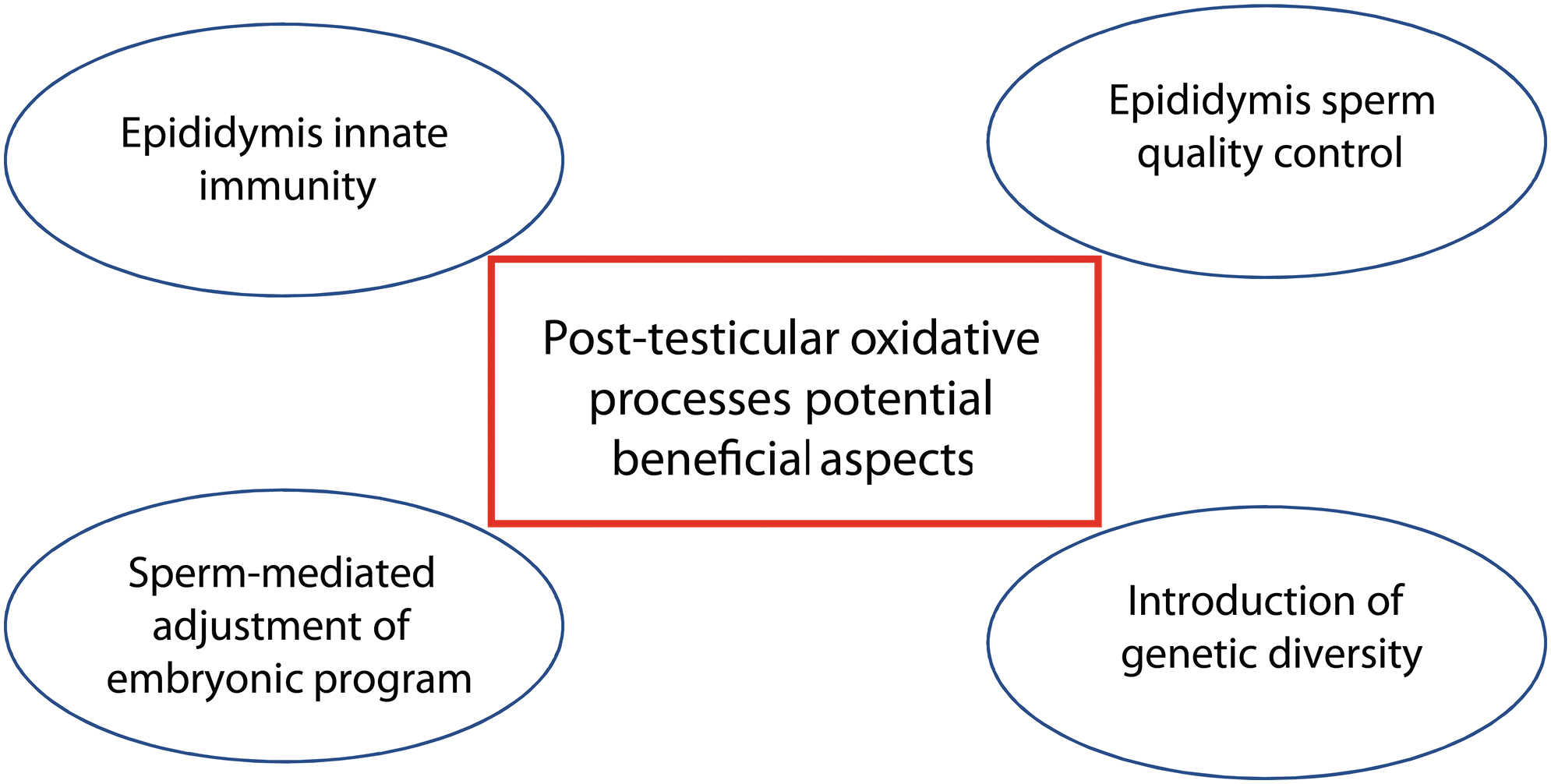
Post-testicular oxidative process and innate immunity
Our current understanding of how immune responses are regulated in the mammalian epididymis may explain why OS is a positive contributor to male reproductive function. Differentiating late in life, spermatozoa and their germline-specific antigens are foreign to the host's immune system whose self-repertoire has been fully established about 2 years after birth (158). Although mammalian evolution has chosen to strictly restrict the access of immune cells and antibodies to the germinal testicular epithelium by developing a tight BTB, it has not done so outside the gonad. This is logical, because being connected to the environment through the excretory ducts, there is a high risk of pathogens ascending to the immune-privileged gonad if effective control is not achieved outside of it.
Therefore, a strong immune checkpoint in the epididymis is necessary to detect any ascending pathogens that may eventually migrate to the testis. Our current view of the organization of the immune biology of the mammalian epididymis supports this as it appears that the BEB in the caput epididymidis is very limited, with each tubule surrounded by a dense network of antigen-presenting cells (34, 35) as well as a very dense network of fenestrated blood capillaries and lymph vessels (1). Taken together, these arrangements support the idea that the epididymal tubule has the capacity to sample its luminal content.
If this occurs, then sperm antigens will also be exposed to the host immune system and various mechanisms are then called upon to establish what is called peripheral tolerance (58). In this context, in mature males, sperm antigens continuously challenge the host immune system and create a controlled, low-level but constitutive inflammatory situation in the epididymis, which classically involves activated ROS-producing leukocytes. In such a scenario, the exposure of spermatozoa to ROS in the epididymal compartment is an inevitable consequence of innate immunity and the necessary establishment of peripheral tolerance by this tissue to cope with the dual immune challenge of orchestrating close surveillance of pathogens while avoiding autoimmune responses.
In support of this, it is observed that macrophages, which are very good producers of H2O2 when activated, are suspected to readily enter the epididymal caput and to be involved in the formation of granulomas in severe inflammatory situations (126). In addition, in support of this hypothesis, it has also been observed that the epididymal caput is the site of constitutive and strong expression of indoleamine 2,3 dioxygenase (IDO), an important player in immune suppression and the establishment of peripheral tolerance (116, 167).
The IDO triggers the kynurenic pathway, whose metabolites have various properties, including anti-inflammatory, anti-apoptotic, and antioxidant (83). Interestingly, when inactivated in a transgenic mouse model, the absence of IDO expression in the epididymis leads to increased local epididymal inflammation and various alterations in sperm structures and functions, providing a link between innate immunity/peripheral tolerance issues and post-testicular sperm quality control (84, 146).
Post-testicular oxidative processes as part of a sperm quality control system
H2O2 is unique among general toxins in that it rapidly kills all cell types by producing highly reactive hydroxyl radicals (OH˙) through the Fenton reaction (76, 108). The cell-killing and DNA-damaging effects of H2O2 have been shown to be mediated primarily by the reaction of H2O2 with chromatin-bound Fe2+ (81, 114). Since sperm cytology allows rapid access to chromatin and given that the epididymal lumen is most likely (as discussed earlier) an H2O2-rich environment, we can assume that spermatozoa harboring a weakly condensed nucleus will readily undergo oxidative DNA alterations that could lead to cell death.
Protamination defects during spermiogenesis as well as other phenomena such as unrepaired meiotic breaks, unremoved apoptotic germ cells, and nuclear decondensation phenotypes due to mechanical shear during nuclear protein replacement are examples of situations that will lead to spermatozoa with poorly condensed chromatin in the epididymis that will be more prone to H2O2-mediated attacks. In this context, the oxidative environment of the epididymis could be considered as part of a true sperm quality control system sending chromatin-defective spermatozoa to a similar apoptotic fate with accumulation of SSB and DSB.
Post-testicular oxidative attack on spermatozoa, as well as sperm sensitivity to oxidative damage, could then be viewed as a well-orchestrated evolutionary mechanism for limiting the reproductive capacity of males under high OS, regardless of its origin. Although clearly speculative, this concept could theoretically be seen as an adaptation to an aerobic way of life ensuring the genetic and reproductive integrity of any given species.
Post-testicular oxidative processes as a means to introduce genetic diversity
We have already mentioned earlier that base oxidation is a highly mutagenic process that could promote the transmission of de novo transversion mutations in the offspring if not repaired by the oocyte. Whether this is only a negative problem, invariably associated with adverse consequences for the embryo and offspring, is perhaps too limited a view. From a more optimistic perspective, one could envision the tremendous selection pressure exerted by oxygen, since the colonization of the atmosphere by aerobic plants and animals, is also at play to contribute to organismal adaptation and genetic diversity.
The wide-open connection between the luminal compartment of the epididymis and the bloodstream may be a means by which the power of oxygen and its by-products can generate random mutagenesis that adds to the genetic shuffling already achieved by meiotic recombination during spermatogenesis (129). The high number of oxidized bases generated in post-testicular sperm can be seen as a means of introducing random mutations if the oocyte machinery leaves them unattended or if false repair is performed.
Both these outcomes can be expected for some of these oxidized bases, as there is no repair system that is not overwhelmed at some point or is error-free. Some of these DNM could have detrimental effects whereas others could potentially have beneficial effects. This could partly support the concept that the testis and spermatozoa are engines of evolution enhancing the creation of spontaneous mutations of the male germ line.
Post-testicular oxidative processes and adjustment of embryonic development by spermatozoa
Dynamic changes in the antioxidant responses mounted by the epididymal epithelium clearly modulate the qualitative and quantitative presence of sncRNAs that will be transmitted to the transiting spermatozoa. Via such means, OS in the epididymis can be considered as a means of adjusting embryonic development so that the offspring are optimally adapted to the environmental influences that affected the father. To this end, the loading of sperm with OS-modified sncRNAs could modulate the earliest stages of embryonic development that depend heavily on mRNAs and proteins accumulated in the oocytes, to rapidly correct the consequences of environmental exposures, whether due to paternal lifestyle choices or because of environmental constraints.
A major consequence of OS on sperm sncRNA might, therefore, be to permit an adaptive response during early embryonic development. Our preliminary investigations exploring the changes in sncRNA profiles recorded in WT versus SKO spermatozoa (C. Chu and A. Moazamian, unpublished data) reinforce this hypothesis. In response to OS, particularly miRs and piRs have been found to predominate whose targets are mRNAs controlling very early events in embryogenesis, in particular DNA repair processes and the balance between proliferation and differentiation. Although speculative, this hypothesis casts the epididymis in a new light.
It is not just a conduit for spermatozoa from the testes to the ejaculate, but also a means by which spermatozoa can be selected and modified to optimize the chances of conception and viable embryonic development.
Conclusion
The ROS and OS have long been known to have complex interactions with mammalian sperm cells. More than any other cell, spermatozoa illustrate how ROS metabolites and their consequences can have not only detrimental impacts on these cells but also beneficial physiological effects to optimally refine sperm structure and function. As highlighted throughout this review, the ROS and OS are often portrayed as destructive because of their impact on the integrity of the paternal genome as well as the long-term health and well-being of the offspring.
However, in some respects, these factors could also be considered as beneficial to innate immunity, sperm quality control, adaptation of the fertilized oocyte/zygote to paternal-inherited environmental stresses, or as a basic mechanism by which aerobic life can contribute to genetic diversity. Fundamentally, the complex relationships that ROS and OS have with spermatozoa, mammalian reproduction, and evolution are, in our opinion, good examples of the delicate redox balance that has to be struck in committing to an aerobic pattern of metabolism.
Because both OS and reductive stress are harmful, a delicate balance has to be struck between oxidative (ROS) and reductive (antioxidants) stress to maintain a fully functional reproductive system. The cellular mechanisms that underpin the maintenance of a balanced redox environment within the male reproductive tract have implications for our ability to both understand the etiology of male infertility and develop suitable strategies for its amelioration.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this work.