
Abstract
Significance:
Circadian-controlled cellular growth, differentiation, and metabolism are mainly achieved by a classical transcriptional–translational feedback loop (TTFL), as revealed by investigations in animals, plants, and fungi.
Recent Advances:
Recently, reactive oxygen species (ROS) have been reported as part of a cellular network synchronizing nontranscriptional oscillators with established TTFL components, adding complexity to regulatory mechanisms of circadian rhythm. Both circadian rhythm and ROS homeostasis have a great impact on plant immunity as well as fungal pathogenicity, therefore interconnections of these two factors are implicit in plant–fungus interactions.
Critical Issues:
In this review, we aim to summarize the recent advances in circadian-controlled ROS homeostasis, or ROS-modulated circadian clock, in plant–fungus pathosystems, particularly using the rice (Oryza sativa) blast fungus (Magnaporthe oryzae) pathosystem as an example. Understanding of such bidirectional interaction between the circadian timekeeping machinery and ROS homeostasis/signaling would provide a theoretical basis for developing disease control strategies for important plants/crops.
Future Directions:
Questions remain unanswered about the detailed mechanisms underlying circadian regulation of redox homeostasis in M. oryzae, and the consequent fungal differentiation and death in a time-of-day manner. We believe that the rice–M. oryzae pathobiosystem would provide an excellent platform for investigating such issues in circadian-ROS interconnections in a plant–fungus interaction context. Antioxid. Redox Signal. 37, 726–738.
Introduction
As is known, Earth's rotation around its own axis and the Sun results in robust rhythmic changes in its environment, so life on Earth has evolved in accordance with an ∼24-h cycle, termed circadian rhythm (95, 119). The molecular basis of the circadian timing system, also called circadian clock, comprises a few core genetic elements driving output pathways and rhythmic physiology, forming a transcriptional–translational feedback loop (TTFL) (83, 91).
The 2017 Nobel Prize in Physiology or Medicine was awarded to researchers for their contribution to the study of molecular mechanisms controlling the circadian rhythm using fruit fly as a model organism (53). However, studies on molecular mechanisms of circadian rhythm were carried out almost at the same time using a fungus, Neurospora crassa. Circadian clock mutants of N. crassa were first reported in the 1970s (35) and then the clock gene frequency (FRQ) was identified in the 1980s (39, 81). The TTFL-controlled circadian rhythm and rhythmic biological processes, including fungal growth and differentiation, were established based on oscillating mRNA and protein levels of the FRQ gene (2).
In the 1990s, the light sensor complex, white collar complex (WCC), comprising white collar-1 (WC-1) and white collar-2 (WC-2), was identified and characterized (24). The FRQ-centered TTFL model was then modified to accommodate light entrainment of the circadian rhythm based on light-induced transcription of the FRQ gene mediated by WCC. Such circadian timekeeping machinery was progressively improved, and recent studies added more layers of regulation to the central oscillators, FRQ and WCC, including post-transcriptional regulation (43), epigenetic and post-translational modifications (7, 8, 12, 50, 77, 92, 93, 99, 123), and ubiquitin/proteasome-mediated protein degradation (51).
Overall, investigation using N. crassa as a preeminent model has led to elucidation of circadian clock regulatory mechanisms that are common to all eukaryotes. At present, different elements driving the TTFL-based clockwork have been found in fungi (N. crassa as a representative: FRQ and WCC) (78), plants (TOC1 and CCA1) (83, 91), and animals (BMAL1 and PERIOD) (13, 62). Although using different elements in different organisms, basic rhythmic oscillation is commonly driven by a positive and negative feedback loop at transcriptional and translational levels, as illustrated in Figure 1.

Moreover, the circadian timekeeping machinery could also employ non-TTFL mechanisms; examples include phosphorylation cycles of the KaiABC system in cyanobacteria (55) and redox cycles of the antioxidant proteins, peroxiredoxins (PRXs), in human red blood cells (85) as well as in N. crassa (33). A schematic summary of the TTFL-based and non-TTFL circadian systems is presented in Figure 1. The circadian clock controls growth, proliferation, differentiation, metabolism, movements, and many other dark/light-associated biological behaviors in various organisms (19, 52, 60, 79, 89, 104) and is also involved in plant immunity (79, 117, 127, 128) and microbial pathogenicity (19, 30, 68).
Redox (oxidation and reduction reactions) involves the reactive oxygen species (ROS), including the superoxide anion (O2 •−), hydrogen peroxide (H2O2), and hydroxyl (OH•) radicals (11, 88). When reaching elevated intracellular levels, ROS are usually associated with damage to cellular lipids, proteins, and DNA. However, accumulated evidence supports that ROS could also serve a physiological function as signaling molecules, named redox signaling (37, 103).
Redox signaling has been reported to regulate cell proliferation and survival, differentiation, iron homeostasis, and programmed cell death (56, 94, 107). Particularly, it has long been known that ROS burst is part of the plant hypersensitive response (HR) against microbial pathogen invasion (1, 38, 118).
Given that both circadian rhythm and ROS homeostasis/signaling closely regulate growth, development, and stress response in both plants and pathogenic fungi, they (unsurprisingly) also affect plant–fungus interactions. In this review, we aim to summarize the recent advances in plant–fungus interactions, under circadian and redox regulation, respectively, as well as in the interconnection between them. Particularly, we use the rice blast fungus pathosystem as an example to demonstrate a potential circadian redox rhythm-based plant–fungus interaction, which may also be applicable to other plant–fungus pathosystems.
Circadian Rhythm and ROS Homeostasis in N. crassa
Recent studies using the model fungal organism, N. crassa, have revealed complex interconnections between circadian rhythm and ROS homeostasis/signaling. Fungi generate ROS as metabolism by-products, using nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) and mitochondrial respiration (14, 34, 87, 96). The antioxidant systems include major ROS-detoxifying enzymes, for example, superoxide dismutases (SODs), catalases (CATs), carotenoids, and PRXs (46).
Glutathione (GSH) is a simple sulfur compound comprising three amino acids, glutamate, cysteine, and glycine, serving a redox-homeostatic buffering function in fungi, including N. crassa (3, 46). In N. crassa, intracellular ROS levels oscillate rhythmically, in accordance with N. crassa morphological changes, at different stages of cellular differentiation (49, 109, 110). NOX, the active ROS generator, shows no obvious circadian rhythmicity at either the transcriptional level or in enzymatic activities, while the ROS-detoxifying enzymes, CATs, do (125).
Transcription of CAT genes and biosynthesis of another antioxidant, carotenoid, are both induced by light and under TTFL control, executed by the FRQ-WC-1-WC-2 oscillator (FWO) (70, 100, 122, 124). Circadian oxidation cycles of PRXs also contribute to ROS oscillation in N. crassa (33). Therefore, ROS oscillation in N. crassa is controlled by both TTFL and non-TTFL mechanisms.
On the other hand, ROS positively regulate WCC activity, either by transcriptional activation of WC1 via the Ras signaling pathway (6, 9) or by stabilizing the WCC via activation of protein phosphatase 2A (47). The ROS-activated WCC in turn enhances transcription of FRQ and other clock-controlled genes (125). A simplified model for circadian-ROS interconnection is shown in Figure 2.

However, N. crassa is not a pathogenic fungus, therefore we could not uncover the mechanism by which such circadian redox rhythm affects plant–fungus interaction and eventually determines disease occurrence and/or severity. In the following sections, we, respectively, discuss about regulation of plant immunity and fungal pathogenicity by the interconnected circadian redox signaling pathways.
Circadian Redox Rhythm of Plant Immunity
Both circadian oscillation and redox homeostasis play important roles in plant growth and development/differentiation. For example, photosynthesis, stomatal opening, and leaf movement occur during the daytime (15, 48, 65, 120), while cell cycle progression and hypocotyl elongation occur during the nighttime (84). On the other hand, redox homeostasis and signaling contribute to plant cell cycle progression (64), root growth (63), flowering (126, 132), senescence, and cell death (27).
H2O2 production is a common result of multiple phytohormone actions (4), therefore we could anticipate the role of redox homeostasis/signaling in plant immunity. In this section, we focus on ROS-based plant defense response against microbial pathogens, potentially under circadian control.
Plants use ROS as a defense response
It has long been known that ROS is an important part of plant immune responses, including pathogen-associated molecular pattern-triggered immunity and effector-triggered immunity (42, 57, 112). Plant peroxidases and NOX, also known as respiratory burst oxidase homologs (RBOHs), are two major types of ROS generators (28, 58, 67). In Arabidopsis thaliana, two RBOH genes, AtrbohD and AtrbohF, are the main generators of intracellular ROS upon infection by pathogens, including bacteria, fungi, and parasitic nematodes (90, 106, 111).
Such Rboh-generating ROS could either trigger (90, 111) or prevent (106) HR cell death, depending on the nature of pathogen–plant interactions, and under regulation of the well-established receptor-like cytoplasmic kinase, Botrytis-induced kinase 1 (BIK1), a component of the flagellin-sensing 2 (FLS2) immune receptor complex (59, 116), through protein phosphorylation (71). In addition, the plasma membrane-localized RBOHs and cell wall peroxidases are also considered as positive regulators of cell death, thus conferring plant defense against various invading pathogens (10, 21).
Two class III peroxidases, peroxidase 33 (AtPrx33) and peroxidase 34 (AtPrx34), are responsible for ROS production crucial for A. thaliana immunity against fungal and bacterial pathogens (10). Generally, plants produce elevated levels of ROS as a barrier to stop/restrict microbial, including fungal pathogen, invasion.
ROS burst is also critical for rice (Oryza sativa) resistance against the blast fungus, M. oryzae. Recently, a single-nucleotide (A to G) change in the promoter of the broad-spectrum resistance Digu 1 (bsr-d1) gene was identified in the rice line, Digu, contributing to inhibition of ROS turnover during M. oryzae infection and thus conferring broad-spectrum resistance to blast disease (72). During incompatible rice–M. oryzae interactions, OsRBOHB (86) of rice is strongly induced and produces ROS specifically distributed at the sites surrounding M. oryzae invasive hyphae and thus restricts their spreading (17).
A recent study showed that such rice ROS generated by nicotinamide adenine dinucleotide phosphate-malic enzyme 2 (OsNADP-ME2) and OsRbohB, besides restricting in planta growth of blast fungus, also contributes to HR cell death (25). Such HR cell death is iron and ROS dependent and thus was defined as ferroptotic cell death (25) under regulation of the mitogen-activated protein kinase (MAPK) signaling cascade comprising mitogen-activated protein kinase kinase 2 (OsMEK2), mitogen-activated protein kinase 1 (OsMPK1), and OsWRKY90 (WRKY is the amino acid sequence within a conserved heptapeptide motif, WRKYGQK, at the N-terminus of the transcriptional factor, therefore it is used to name the families of transcriptional regulators) (26).
Overall, ROS burst, followed by HR cell death, is an important component of plant immunity, including in rice, against the blast fungus.
Circadian regulation of ROS-based plant defense response
Accumulating evidence supports reciprocal control of the circadian clock and ROS homeostasis in plants, both of which could regulate plant immunity. A daily rhythmic accumulation of ROS in plants has been reported, possibly resulting from the rhythmic expression of genes encoding ROS-generating or -neutralizing enzymes (23, 66). On the other hand, ROS functions as an input signal to regulate the transcriptional output of the plant clock (66).
Several phytohormone signaling pathways involved in plant immunity have been shown to be controlled by the circadian clock, including jasmonates (JAs) and salicylic acid (SA) (41). Particularly, SA mediates systemic acquired resistance, part of which is ROS production (18). Inhibition of catalase by SA results in elevated intracellular ROS levels, which suppress the master immune regulator, nonexpressor of pathogenesis-related gene 1 (NPR1), by keeping it in the oligomeric form (82).
Following such early ROS burst, a transient increase in cell reduction potential by accumulation of antioxidants such as GSH triggers conversion of the NPR1 oligomer to a monomeric form, which in turn enters the nucleus and activates transcription of pathogenesis-related genes involved in plant resistance (82), as well as core circadian clock genes, conferring a circadian redox rhythm of plant immunity response (130, 134).
It has also been reported that plant auxin/indole-3-acetic acid (IAA) singling and response are controlled by the circadian clock (22, 121). Auxin/IAA could be utilized as an ROS-detoxifying strategy, likely via transcriptional activation of genes involved in ROS detoxification, DNA repair, and iron sequestration, as recently reported in auxin-producing bacteria (113). The circadian redox regulation of plant immunity is briefly summarized as the scheme in Figure 3.

Such phenomenon of circadian-gated immune responses may be prevalent in plants, including rice. A dark phase immediately after M. oryzae–rice contact is required for development of disease symptoms as the infected rice seedlings constantly exposed to light fail to develop disease symptoms (61). This phenomenon is termed light-dependent disease suppression (61). We infer that it is partially due to stronger resistance of rice against M. oryzae during the daytime.
Supporting this hypothesis, light-induced ROS accumulation in rice during blast fungus infection has been recently reported (76) and provides new evidence supporting circadian/light regulation of the blast disease resistance mechanism. Circadian-controlled auxin biosynthesis and signaling are also reported in rice (36).
Although auxin/IAA is not directly involved in plant immunity response, it is observed that an elevated auxin/IAA level caused reduced resistance of rice against blast fungus (129). Therefore, circadian rhythmic auxin/IAA fluctuation also potentially contributes to higher resistance to blast in the daytime and elevated susceptibility in the night.
In summary, we propose that plants boost their immunity systems in a phase when pathogen infection is anticipated, while allowing growth in the opposite phase of the day. Such circadian-controlled immunity response–growth balance helps plants minimize fitness costs.
Circadian and Redox Regulation of Fungal Pathogenicity: Rice Blast Fungus as an Example
As the host plants use ROS as a defense mechanism against microbial, including fungal, pathogens, the pathogens evolve multiple strategies to suppress or detoxify such oxidative stress during their invasion. On the other hand, recent studies revealed that ROS not only causes cellular damage but also acts as a signaling molecule carrying out physiological functions, including pathogenic development, in pathogenic fungi. In principle, pathogenic fungi utilize ROS-detoxifying enzymes and/or the antioxidant system to counteract the oxidative stress imposed by the host plant, while producing endogenous (low level) ROS as a physiological signal to regulate pathogenic development (101).
In the following sections, we will give examples of M. oryzae counteracting the oxidative stress during infection of host rice, as well as the mechanisms underlying redox signaling during M. oryzae pathogenic development. Particularly, we focus on the circadian control of M. oryzae pathogenicity, potentially via redox homeostasis/signaling.
Antagonizing ROS stress during host infection
Fungi utilize ROS-detoxifying enzymes or produce antioxidants to antagonize ROS stress and thus facilitate infection of the host. In this section, we list a few examples from the rice blast fungus. M. oryzae secretes ROS-scavenging enzymes, peroxidase and laccase, to prevent accumulation of host-derived ROS and the consequent host defense response (20). Expression levels of peroxidases and laccases were under regulation of a novel pathogenicity gene, defense suppressor 1 (DES1) (20), and the basic leucine zipper transcription factors, activator protein 1 (MoAp1) (44) and activating transcription factor 1 (MoAtf1) (45).
M. oryzae produces the antioxidant, GSH, via the gamma-glutamyl cycle, which is under regulation of sorting nexin-41 (MoSnx41)-mediated endosomal protein sorting, and acts to suppress the host-derived ROS stress and ensure its invasive growth (29). M. oryzae could also manipulate the rice ROS system by secreting fungal effector proteins. For example, it secretes the effector, avirulence Pii (AVR-Pii; Pii is the name of the corresponding resistant protein in rice), to interfere with the rice ROS production system, OsNADP-ME2 (as discussed in the Plants use ROS as a defense response section), and thus disrupt ROS burst and host immunity (108). M. oryzae produces the phytohormone, auxin/IAA (32, 54)
Redox signaling during pathogenic differentiation
Besides dealing with the ROS stress derived from the plant host defense response, pathogenic fungi could also actively utilize ROS as a physiological molecule to regulate their pathogenic development (102, 133). In M. oryzae, two NOXs (5), Nox1 and Nox2, are responsible for ROS production critical for fungal pathogenicity (34). Nox1 is required for ROS accumulation during two critical steps of M. oryzae pathogenic development: appressorium (a specialized infection cell) formation and penetration peg elongation within host tissues (34, 97).
Nox2 is necessary for ROS accumulation to form functional appressorium and develop the penetration peg (16, 34, 97). Our recent study demonstrated that M. oryzae uses Nox2 to generate lipid peroxides (oxidized lipid of the membrane), which triggers a programmed death of the conidia (asexual spores) essential for appressorium maturation (105). Suppression of ROS is required to facilitate fungal invasive growth, but compromises appressorium formation and functioning (74).
Circadian/phototropic regulation of M. oryzae pathogenicity
We can see that M. oryzae needs to precisely control intra- and extracellular redox oscillation during pathogenic development, namely to increase the ROS level during conidial and appressorium development and decrease the ROS level during appressorium-mediated host invasion. Such spatiotemporal control of redox oscillation is indicative of underlying circadian/phototropic regulation. Indeed, M. oryzae possesses a conserved fungal core circadian timekeeping machinery, FWO.
The rhythmic conidial banding documented in N. crassa can also be observed in M. oryzae grown on solid medium under dark/light cycles (Fig. 4). The FRQ ortholog in M. oryzae, MoFRQ, displays rhythmic oscillation at the transcriptional level (30), suggesting the presence of a functional circadian clock. Interestingly, such banding could also be observed in the mofrqΔ mutant (our unpublished result), but is lost in the deletion mutant of the MoWC1 gene (69) or MoWC2 gene (our unpublished result) (Fig. 4). This suggests that the FWO in M. oryzae may function differently than in N. crassa. The circadian/phototropic control of fungal development may also exist in M. oryzae.

We mentioned that rice develops higher resistance to blast fungus in the daytime and shows elevated susceptibility in the night. Correspondingly, M. oryzae has evolved to adapt to such circadian oscillation in host resistance. Forming of M. oryzae asexual spores (conidia), termed conidiation, is light induced (69). The blue light receptor, MoWc-1, is required for release of conidia, which is suppressed by light, but not for light-induced formation of conidia (61, 69).
MoWc-1 is also required for sensing darkness immediately after inoculation of the conidia on rice, thus mediating light-dependent disease suppression (69). This light-dependent disease suppression phenomenon may reflect the outcome of coevolution of rice and M. oryzae. Furthermore, MoWc-1 is required for light-induced MoFRQ transcription (61). Therefore, the core FWO circadian timekeeping machinery is conserved in M. oryzae and plays important functions in regulating pathogenic development.
We reported a circadian-controlled gene, Twilight (TWL), in M. oryzae (30) whose encoding product undergoes phosphorylation and acetylation modifications under dark/light cycles and shuttles between the nucleus and cytosol. Twl regulates the phototropic expression of a transcriptional factor, Tfb, and thus triggers light-induced conidiation (30).
We also identified a histone acetylase-encoding gene, general control nonrepressed 5 (GCN5), downstream of Twl (30). Gcn5 negatively regulates M. oryzae autophagy by acetylation of the autophagy protein, Atg7, in the dark, but loses interaction with Atg7 under light, thus conferring phototropic induction of autophagy (131). Autophagy promotes M. oryzae conidiation (31) in response to light exposure.
On the other hand, programmed cell death of developing conidia, which is also autophagy dependent, is crucial for M. oryzae appressorium functioning (115). Such conidial death is iron and ROS (including lipid peroxide and products of oxidative lipid damage) dependent, termed ferroptosis (105). We found that Gcn5 regulates autophagy-dependent degradation of pyruvate kinase (PK) at the stage of conidial development to ensure successful host infection (75). PK was reported to regulate iron homeostasis (80, 98, 114) and the cellular ROS level (73).
Therefore, we infer that such Twl-Gcn5-autophagy-PK axis is indicative of circadian-controlled oscillation of the intracellular ROS level and/or cell death, coordinating M. oryzae pathogenic development. Furthermore, our recent study reveals that M. oryzae-derived IAA promotes such ferroptosis-based conidial death (32), but the detailed mechanism is still unclear. IAA goes through oxidation under light (40), therefore this fungus-derived IAA may also function in controlling oscillation of fungal ROS homeostasis and/or pathogenesis on a circadian basis.
We summarize the M. oryzae ROS homeostasis controlled by rhythmic autophagy and IAA activities in Figure 5.

Summary and Outlook
ROS is an important regulator of interactions between rice and M. oryzae and is potentially kept under control by circadian rhythm. In this review, we summarized the recent advances in circadian-controlled plant immunity and fungal pathogenic development, both on a redox oscillation basis. We infer that M. oryzae has evolved an ability to regulate its pathogenic development, including conidial formation, conidial release and development, conidial death (for appressorium formation and maturation), and appressorium-mediated invasive growth, according to the light/dark conditions.
Such ability can help M. oryzae adapt to the Earth dark/light cycles and the consequent periodical change of environmental factors, including host plant physiological processes. A schematic presentation of M. oryzae pathogenic development under the dark/light cycle is proposed in Figure 6.
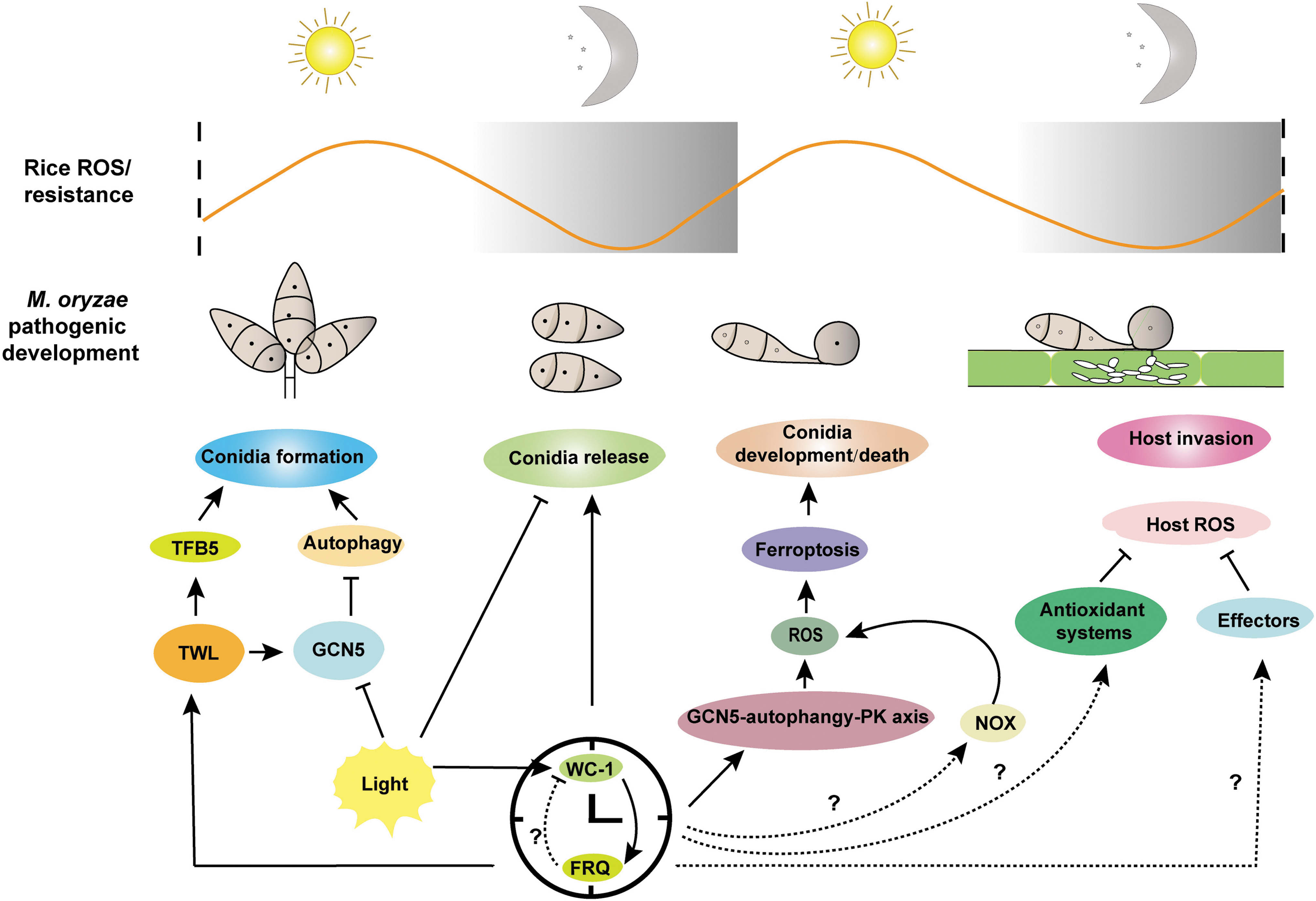
However, we notice that important questions remain unanswered regarding the circadian/redox regulation of M. oryzae pathogenic development: (i) Is there an FWO-based regulation on M. oryzae ROS oscillation, which increases ROS levels in the daytime for conidial formation and/or developmental conidial death (daytime) and decreases fungus- or plant-sourced ROS during the invasion stage (nighttime)? (ii) Does ferroptosis-based conidial death oscillate in dark/light cycles and is it dependent on FWO? (iii) Is fungal IAA production controlled by the FWO clock (and how)? (iv) What is the role (if any) of fungal ROS (including lipid peroxides)/IAA in shaping the FWO clock?
The rice–M. oryzae pathosystem is an excellent platform for investigation of these detailed mechanisms underlying circadian-ROS interconnections during plant–fungus interactions. Understanding the circadian regulation of redox homeostasis and the consequent fungal differentiation and death would also provide a theoretical basis for developing disease control strategies for important plants/crops.
Footnotes
Author Contribution Statement
M.L. and L.D. contributed equally to the concept and design of the review. Y.Z.D. had final responsibility for the decision to submit for publication. Y.Z.D. completed the literature search, drafted the review, and, with M.L. and L.D., finalized the review. L.D created the figures, with Y.Z.D., and conducted the literature search associated with the figures. All authors have reviewed and approved the manuscript before submission.
The review has been submitted solely to Antioxidants & Redox Signaling and is not published, in press, or submitted elsewhere.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work is supported by grants from the National Natural Science Foundation of China (Grant No. 32022070) and the Key Realm R&D Program of Guangdong Province (2020B0202090001) to Y.Z.D. and the China Postdoctoral Science Foundation (2020M682723) to M.L.