
Abstract
Significance:
Hydrogen sulfide (H2S) is a multitasking potent regulator that facilitates plant growth, development, and responses to environmental stimuli.
Recent Advances:
The important beneficial effects of H2S in various aspects of plant physiology aroused the interest of this chemical for agriculture. Protein cysteine persulfidation has been recognized as the main reduction–oxidation (redox) regulatory mechanism of H2S signaling. An increasing number of studies, including large-scale proteomic analyses and functional characterizations, have revealed that H2S-mediated persulfidations directly regulate protein functions, altering downstream signaling in plants. To date, the importance of H2S-mediated persulfidation in several abscisic acid signaling-controlling key proteins has been assessed as well as their role in stomatal movements, largely contributing to the understanding of the plant H2S-regulatory mechanism.
Critical Issues:
The molecular mechanisms of the H2S sensing and transduction in plants remain elusive. The correlations of H2S-mediated persulfidation with other oxidative post-translational modifications of cysteines are still to be explored.
Future Directions:
Implementation of advanced detection approaches for the spatiotemporal monitoring of H2S levels in cells and the current proteomic profiling strategies for the identification and quantification of the cysteine site-specific persulfidation will provide insight into the H2S signaling in plants. Antioxid. Redox Signal. 39, 40–58.
Introduction
Sulfur is the 10th most abundant chemical element in the universe and is essential for all living organisms (Räisänen, 2005). It occupies a unique position in the reduction–oxidation (redox) biology due to its availability to reach many distinct oxidation states, ranging from −2 to +6 (Fig. 1). As the planet got oxidized, sulfate (+6) became the most abundant inorganic form of sulfur on earth. Hydrogen sulfide (H2S), the most reduced inorganic form of sulfur (−2) (Fig. 1), is a colorless, but flammable, gas, with a smell of rotten eggs that is naturally released from volcanic emissions or other geothermal activities and from decaying plant and animal proteins.
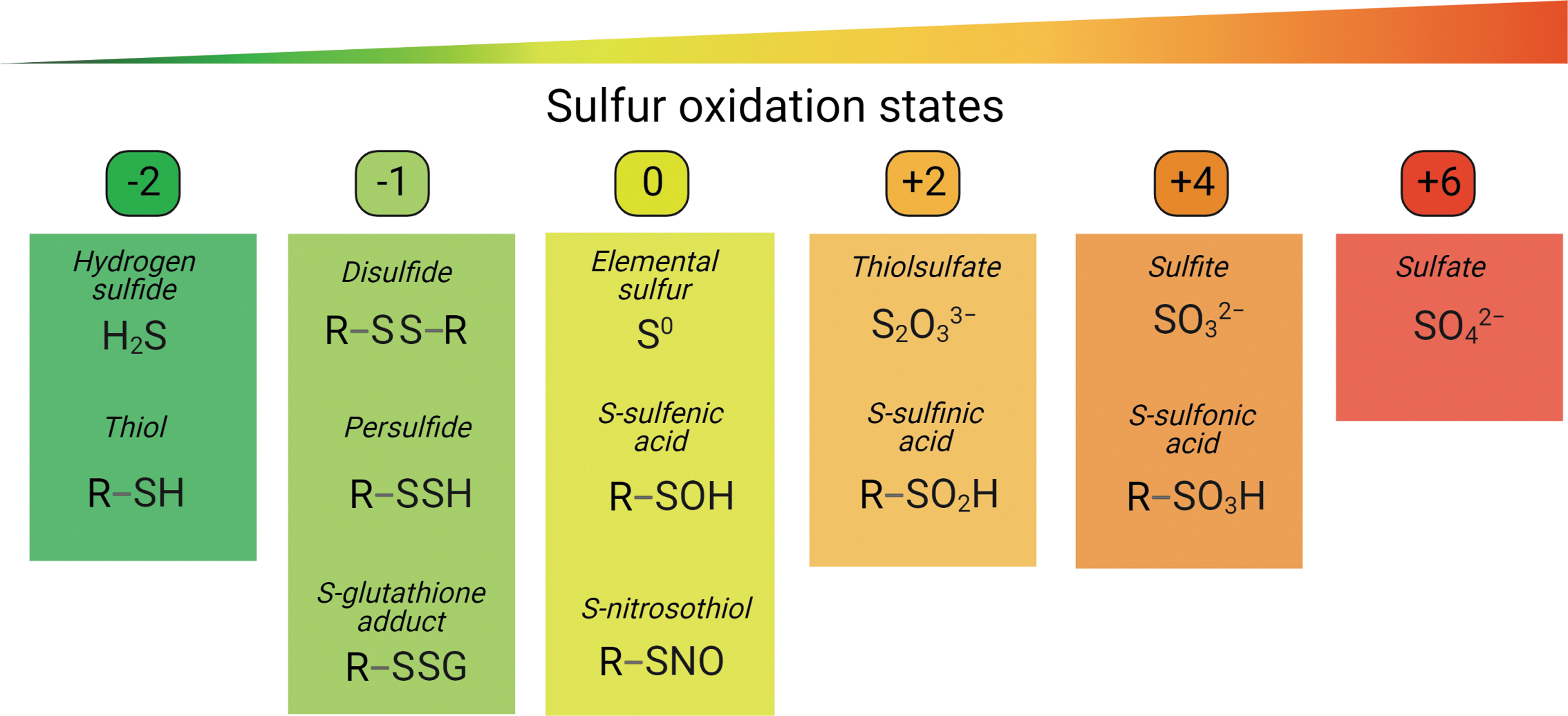
As H2S is a weak acid with the dissociation constants pKa1 of 6.9 (Pomeroy, 1941) and pKa2 of between 12 and 17 (Ellis and Golding, 1959; Meyer et al., 1983), it can be dissociated into hydrosulfide anion (HS¯) and sulfide anion (S2¯) in aqueous solutions. H2S usually stands for all species, including H2S, HS¯, and S2¯ (Paulsen and Carroll, 2013). Nevertheless, in solutions at an approximately physiological pH of 7.4, H2S releases negligible amounts of S2¯ and exists primarily as HS¯ (Hughes et al., 2009).
Given the basic chemical properties of H2S and HS¯ with the lowest oxidation state of −2, they both can only be oxidized. Whereas H2S is a gasotransmitter that can diffuse freely across cellular membranes, HS¯ needs specific ion channels to move between different subcellular organelles or cells (Kabil and Banerjee, 2010). Accordingly, the H2S and HS¯ might regulate cellular functions differently. H2S, HS¯, and S2¯, together with various chemically reactive forms of cysteine thiols (see in section H2S Signaling via Protein Persulfidation) and other sulfur-containing compounds that either reduce or oxidize biomolecules, can be classified as reactive sulfur species (RSS).
H2S has been implicated in the origin of life (Filipovic et al., 2018; Olson and Straub, 2016). Life began 3.8 billion years ago (bya) in an anoxic and ferrous ion (Fe2+)-rich ocean. Cyanobacteria, the first photosynthetic oxygen-generating organisms, are believed to have then evolved and contributed to the Great Oxygenation Event ∼2.5 bya (Demoulin et al., 2019; Fournier et al., 2021; Planavsky et al., 2014). Along with the slightly increased oxygen level, sulfur oxidized to sulfate, which was further reduced to sulfide by ubiquitous Fe2+ present in the ocean, greatly increasing the H2S level and, consequently, leading to an anoxic and sulfidic ocean (Cortese-Krott et al., 2017).
The first eukaryotes appeared and adapted under this condition using H2S as their major energy source (Olson and Straub, 2016). Green algae, one of the earliest photosynthetic eukaryotes of the Plantae kingdom that contained primary chloroplasts, derived from endosymbiosis with cyanobacteria, are assumed to have evolved as early as 1.0 bya (Tang et al., 2020). The combined activity of cyanobacteria and algae tremendously increased the level of ambient oxygen ∼0.6 bya, sequentially, land plants have evolved.
The antioxidant enzymes, such as superoxide dismutase (Cannio et al., 2000; Miller, 2012), catalase (Zamocky et al., 2008), glutathione (GSH) peroxidase (Margis et al., 2008), peroxiredoxins (Dietz, 2011; Knoops et al., 2007), thioredoxins (TRXs) (Balsera and Buchanan, 2019), and glutaredoxins (GRXs) (Alves et al., 2009), were already present in an anoxic and high H2S environment as early as 2.0 bya (Cortese-Krott et al., 2017). Therefore, these redox regulation systems most probably evolved primarily to use H2S as energy source or to deal with RSS, which was later amended to regulate reactive oxygen species (ROS) (Cortese-Krott et al., 2017; Olson and Straub, 2016).
Along with the evolution and increasing complexity of organisms, the ancient mechanisms of H2S metabolism and regulation had to be adapted. For example, the sulfate transport system in chloroplasts has been suggested to undergo several adaptions, spanning the evolution of green algae, liverworts, and flowering plants (Kopriva et al., 2015; Mendoza-Cózatl et al., 2005; Takahashi et al., 2012). In brief, H2S metabolism as well as regulatory mechanisms in living organisms has evolved as a result of environmental adaptation, while the fundamental principles might remain conserved (Yamasaki and Cohen, 2016).
Besides its implied vital role in evolution, H2S has been recognized as a potent signaling molecule in the regulation of critical cellular processes (Wang, 2014; Wang, 2002). Increasing evidence has shown that H2S is not only involved in plant growth, development, reproduction processes (Baudouin et al., 2016; Chen et al., 2011; Ma et al., 2021), and promotion of nodulation in the rhizobium–legume symbiosis (Zou et al., 2019), but also facilitates tolerance to various environmental stresses, such as drought (Jin et al., 2011; Shen et al., 2013), salinity (Christou et al., 2013; Li et al., 2014a), heavy metals (Fu et al., 2019; Zhang et al., 2020b), and extreme temperatures (Du et al., 2021; Li et al., 2012).
Furthermore, H2S has been reported to improve the quality maintenance during postharvest storage of fruit (Ge et al., 2017; Hu et al., 2012), vegetables (Li et al., 2014b), and flowers (Zhang et al., 2011). Given the important beneficial effects of H2S in multiple aspects of plant physiology, exogenous H2S seems to be a promising biotechnological strategy with a great agronomic interest.
Numerous studies have uncovered the positive physiological impact of H2S on plants (Corpas, 2019; Corpas and Palma, 2020; Zhang et al., 2021), but how plants sense and transduce H2S signals remains elusive. H2S can modify proteins through post-translational modification (PTM), a process named persulfidation. Persulfidation on critical proteins plays an important role in activating downstream signaling as demonstrated by both proteomic analyses and functional characterizations (Aroca et al., 2021a, Aroca et al., 2021b, Aroca et al., 2017b, Aroca et al., 2015; Fu et al., 2020a; Laureano-Marín et al., 2020; Zivanovic et al., 2019).
Recently, H2S-induced protein persulfidation has been found to regulate stomatal movements in the abscisic acid (ABA) signaling pathway (Chen et al., 2020; Shen et al., 2020; Zhou et al., 2021), contributing to the understanding of the molecular mechanism of H2S signaling in plants.
In this review, we provide a wide perspective of H2S in plants, including H2S biosynthesis, exogenous application, endogenous detection methods, and the mechanisms and identification strategies for protein persulfidation. We further give insights into the molecular mechanisms of persulfidation in the regulation of ABA-mediated stomatal closure by highlighting several recent functional studies.
H2S Biosynthesis in Plants
Plants generate H2S endogenously through several biosynthesis pathways in different subcellular organelles (Fig. 2). The major H2S source is associated with the photosynthetic sulfate assimilation pathway in chloroplasts. Plants take up sulfate from the environment through sulfate transporters (Takahashi et al., 2011), a protein class with high sulfate affinity that facilitates sulfate trafficking across membranes, into the chloroplasts, where H2S is mainly generated through sulfur metabolism.
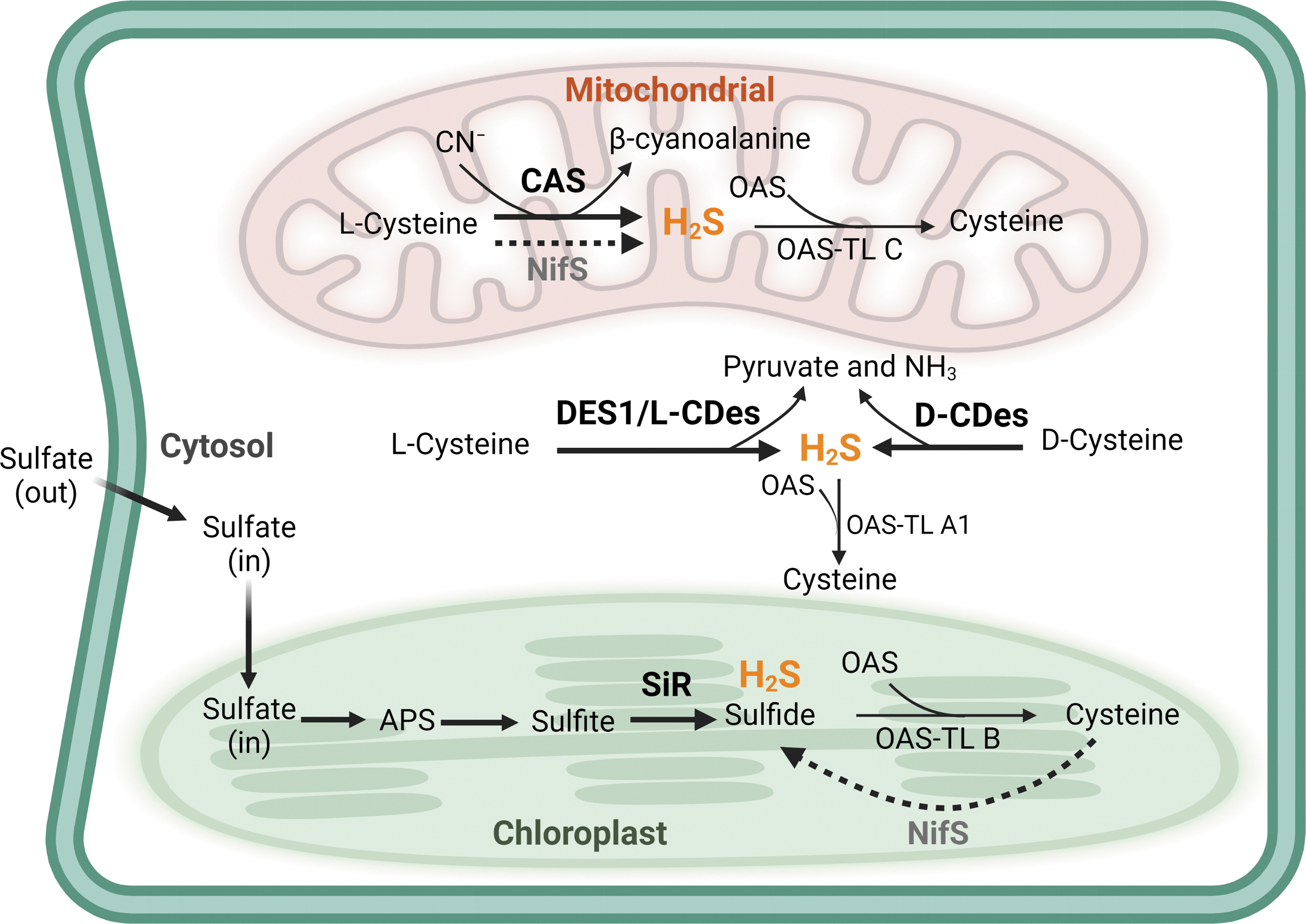
Sulfate is reduced by ATP sulfurylase to form the adenosine 5′-phosphosulfate (APS) intermediate that is further reduced to sulfite by the APS reductase. H2S is then produced from sulfite in the reaction catalyzed by sulfite reductase (Takahashi et al., 2011) (Fig. 2). H2S reacts with O-acetylserine (OAS), generating cysteine via catalyzation by OAS (thiol)lyase (OAS-TL) (Fig. 2). Based on an in vitro activity assay, OAS-TL has been suggested to catalyze the reverse reaction to break down cysteine into H2S and OAS (Burandt et al., 2001). However, negligible amounts of H2S were formed when compared with the cysteine production, indicating that the OAS-TL reaction is a net H2S-consuming reaction (Bloem et al., 2004).
Moreover, the production of endogenous H2S in planta by OAS-TL remains unclear. In plants, the main OAS-TLs responsible for cysteine synthesis are the cytosolic OAS-TL A1, the chloroplastic OAS-TL B, and the mitochondrial OAS-TL C (Fig. 2). Recently, the H2S level was found to be higher in an oas-tl a1 mutant than in wild-type Arabidopsis thaliana plants, confirming the major biological function of OAS-TL in cysteine biosynthesis rather than in H2S generation (Li et al., 2018).
Cysteine desulfhydrase (CDes) was the first studied H2S-producing enzyme (Harrington and Smith, 1980; Tishel and Mazelis, 1966). Because of its ubiquitous activity in various physiological processes of different plant species (Zhao et al., 2020), CDes is considered to be the most critical enzymatic source of H2S in plants. There are two types of CDes:
Moreover, the nitrogenase Fe–S cluster (NifS), localized both in chloroplasts and in mitochondria, is a putative H2S-producing enzyme due to its L-CDes-like activity (Pilon-Smits et al., 2002; Van Hoewyk et al., 2008). In addition, the mitochondrial β-cyanoalanine synthase (CAS) detoxifies cyanide that appeared in the cells to β-cyanoalanine in the presence of cysteine, along with the production of H2S (Hatzfeld et al., 2000) (Fig. 2).
In mammals, 3-mercaptopyruvate sulfurtransferase (MST) that belongs to the sulfurtransferase (STR) family is one of the most important H2S-producing enzymes, but information about MST in plants is scarce. Recently, two Arabidopsis MSTs, the mitochondrial STR1 and the cytosolic STR2, were characterized by means of an in vitro activity assay, revealing their H2S-producing capability in the presence of reducing systems, such as TRXs and GRXs (Moseler et al., 2021). Nevertheless, the in planta contribution to H2S biosynthesis from STR1 and STR2 awaits to be further investigated.
Exogenous H2S Application in Plants
Currently, investigation of the physiological roles of H2S is mainly based on studies applying exogenous H2S donors. To date, various H2S donors have been developed (Powell et al., 2018; Yang et al., 2022) and have been widely used in plant studies (Corpas, 2019; Corpas and Palma, 2020; Liu et al., 2021). Here, we provide a short discussion and an update on these donors.
Sulfide salts, such as sodium hydrosulfide (NaHS) and sodium sulfide (Na2S), are inorganic compounds that release H2S by hydrolysis. To date, NaHS is the most popular and widely applied H2S donor in various plant species (Corpas, 2019; Corpas and Palma, 2020; Liu et al., 2021). The use of NaHS as H2S donor has greatly improved our understanding of the biological function of H2S, but the NaHS shortcoming is that it does not mimic the biological effects of the physiological H2S generation.
Indeed, NaHS hydrolyzes immediately in aqueous solutions, and instantaneously releases large amounts of H2S, HS−, and S2− species, a process very different from the endogenously slow and continuous H2S enzymatic production. Therefore, chemicals with a slow H2S-releasing rate are required.
The morpholin-4-ium 4-methoxyphenyl(morpholino) phosphinodithioate (GYY4137) is such a slow releasing H2S compound that had initially been synthesized and evaluated with a vasodilatory and antihypertensive activity (Li et al., 2008). Due to the commercial availability and application feasibility, GYY4137 is the most widely used H2S donor, besides sulfide salts in the mammalian field (Powell et al., 2018). In plants, GYY4137 often applied in parallel with NaHS that has similar effects. For example, similarly to NaHS, GYY4137 application could induce stomatal closure in Arabidopsis (García-Mata and Lamattina, 2010; Honda et al., 2015) and Nicotiana tabacum (tobacco) (Papanatsiou et al., 2015), and could improve growth of Pisum sativum (pea), Lactuca sativa (lettuce), and Raphanus sativa (radish) (Carter et al., 2018).
Recent environmentally friendly slow H2S-releasing chemicals, dialkyldithiophosphates and disulfidedithiophosphates, have been shown to improve the growth of Zea mays (maize) plants, hinting at potential applicability in agriculture (Brown et al., 2021; Carter et al., 2019). Another novel H2S donor is a class of nitric oxide (NO)–hydrogen sulfide-releasing hybrid (NOSH) compounds that release NO and H2S simultaneously, which was designed for its extreme effectiveness in growth inhibition of human cancer cell lines (Kodela et al., 2012). NOSH and its aspirin hybrid (NOSH-aspirin) that also releases acetylsalicylic acid have been shown to improve drought tolerance of Medicago sativa (alfalfa) (Antoniou et al., 2020), suggesting that NOSH might be a promising plant priming agent against environmental stresses.
H2S has been reported to act as electron donor for respiration, and to contribute to ATP production in mitochondria of prokaryotes (Sakurai et al., 2010) and in a variety of species, such as California killifish (Fundulus parvipinnis) (Bagarinao and Vetter, 1990), marine mussel (Geukensia demissa) (Doeller et al., 2001; Doeller et al., 1999; Parrino et al., 2000), sandworm (Arenicola marina) (Vökel and Grieshaber, 1997), chicken (Gallus gallus) (Yong and Searcy, 2001), and human (Homo sapiens) (Goubern et al., 2007).
A mitochondria-specific H2S donor (10-oxo-10-(4-(3-thioxo-3H-1,2-dithiol-5yl)phenoxy)decyl), triphenylphosphonium bromide (AP39) (Le Trionnaire et al., 2014), can be used as a useful tool to specifically investigate the biological function of H2S in mitochondria. AP39 was initially used in murine microvascular endothelial cells, had an antioxidant and cytoprotective impact under oxidative stress conditions (Szczesny et al., 2014), and later improved the mitochondrial function in the nematode Caenorhabditis elegans (Fox et al., 2021). H2S is certainly tightly linked to the mitochondrial electron transport chain (ETC) activity in the above-mentioned species. However, the knowledge of this aspect in plants remains modest.
Recently, AP39 was applied in Arabidopsis to investigate stomata movement (Pantaleno et al., 2023). In addition to the induction of stomatal closure, AP39 could modulate mitochondrial ETC activity and redox homeostasis of guard cells, providing the first piece of evidence that H2S modulates mitochondrial energetics in plants.
In vivo Detection of H2S in Plants
The important physiological functions of H2S have attracted attention in plant research, compelling the development of detection techniques in living cells, tissues, and different organisms, but the direct detection of endogenous H2S in plants remains a challenge. Traditional detection methods for H2S, such as colorimetric assays (Siegel, 1965), gas chromatography (Hannestad et al., 1989), high-performance liquid chromatography (Shen et al., 2011), polarographic H2S sensor (Doeller et al., 2005), and ion-selective electrode (Li et al., 2000), typically require sample destruction and are limited to in vitro detection.
Fluorescent probes are emerging as tools for the noninvasive study of reactive species in situ in different biological systems, because of their high cell permeability and specificity. Different fluorescent approaches, including chemical and genetically encoded probes, have been used for H2S detection (Chen et al.,2013b, Chen et al., 2012; Lin et al., 2015; Liu et al., 2011a; Youssef et al., 2019). Despite the considerably fewer reports in plants than in the mammalian field, fluorescent probes are considered effective and noninvasive tools for the real-time detection and imaging of H2S in plants.
As the methods for H2S detection have been extensively reviewed (Filipovic et al., 2018; Kong et al., 2022; Lin et al., 2015; Luo et al., 2022; Zeng et al., 2021; Zhao et al., 2020), we will discuss the fluorescent probes recently used to discover endogenous H2S in different plant species and organisms. In general, the various strategies can be categorized into three groups based on the reaction types, namely azide/nitro/nitroso reduction, copper sulfide precipitation, and nucleophilic reaction-based methods (Luo et al., 2022). The fluorescent probes used in plant studies are mainly based on azide reduction and nucleophilic reaction (Fig. 3).

7-Azido-4-methylcoumarin
The azide reduction-based chemical probe, 7-azido-4-methylcoumarin (AzMC), is commercially available. The strong electron-withdrawing ability of the azide group of AzMC quenches its fluorescence, whereas H2S reduces the azide to amine, thus turning on the fluorescence (Kong et al., 2022). Initially, AzMC has been used in photoaffinity labeling of the substrate-binding site of the human phenol sulfotransferase (Chen et al., 1999), and later for H2S detection in living cells and cardiac tissues (Chen et al., 2013a). In plants, AzMC was first applied to measure the H2S level in tomato (Solanum lycopersicum) guard cells in response to ethylene (ET) signal transduction upon osmotic stress (Jia et al., 2018).
Recently, this probe has been utilized to determine H2S levels in Arabidopsis guard cells from wild-type and des1 mutant plants, which are deficient in cytosolic H2S generation due to the lack of DES1, in response to ABA-induced—stomatal closure (Shen et al., 2020; Zhang et al., 2020a). The ABA-induced H2S accumulation in guard cells of wild-type plants was abolished in the des1 mutant plants, whereas the H2S donor NaHS could clearly induce H2S in plants of both genotypes (Fig. 3), indicating that DES1 is responsible for the sensitivity of ABA-induced stomatal closure (Zhang et al., 2020a).
Silicon nanodots-4-azido-N-alanine-1,8-naphthalimide
Another azide reduction-based chemical probe is the silicon nanodots-4-azido-N-alanine-1,8-naphthalimide (SiND-ANPA-N3
Washington state probe-1
The Washington state probe-1 (WSP-1) is a nucleophilic reaction-based chemical probe (Liu et al., 2011a) that contains a pyridyl disulfide moiety and a fluorophore group. H2S reacts with the pyridyl disulfide and generates a persulfide (–SSH) intermediate that undergoes a spontaneous intramolecular cyclization to release the fluorophore. By using WSP-1 in tomato roots, an increased level of H2S was detected upon an exogenous NO donor treatment, which was inhibited by applying the NO scavenger cPTIO (Li et al., 2014c) (Fig. 3). Until now, WSP-1 was utilized for endogenous H2S detection in the roots of turnip (Brassica rapa), and the H2S level decreased upon selenium treatment (Chen et al., 2014) (Fig. 3).
Sulfane sulfur probe 4
The nucleophilic reaction-based sulfane sulfur probe, sulfane sulfur probe 4 (SSP4), has been developed based on the original design of the SSP1 and SSP2 probes (Chen et al., 2013b). Sulfane sulfur reacts with the nucleophilic thiols in the nonfluorescent SSP4 to form a persulfide intermediate that reacts with electrophilic ester groups, leading to spontaneous intramolecular cyclization and release of the fluorophore.
Recently, SSP4 has been used for endogenous H2S detection during the root nodule symbiosis in the legume Lotus japonicas, and an increased level of H2S has been observed during nodulation (Fukudome et al., 2020) (Fig. 3). SSP4 is commercially available, and has a relatively high selectivity and sensitivity to sulfane sulfurs. However, sulfane sulfur probes are not specific for H2S, because they also react with persulfides and polysulfides.
Sulfane sulfur near-infrared probe
Another sulfane sulfur probe that shares a thiophenol moiety and an ester linker with the SSP4 probe is attached to a near-infrared (NIR) fluorophore, designated sulfane sulfur NIR probe (SSNIP). The SSNIP probe has been tested in Arabidopsis roots (Fig. 3) during different growth stages (Jiang et al., 2019).
2-(2-Hydroxyphenyl) benzothiazole-based H2S probe
The 2-(2-hydroxyphenyl) benzothiazole-based H2S probe, HBTP-H2S, is a recent NIR fluorescent probe and contains a dinitrophenyl (DNP) ether that undergoes thiolysis under nucleophilic attacks by H2S, releasing the fluorophore. This NIR fluorescent probe has been applied for in situ bioimaging of endogenous H2S in rice (Oryza sativa) roots, and revealed an increased level of H2S under aluminum ion (Al3+) and flooding stresses (Wang et al., 2021) (Fig. 3).
Genetically encoded H2S sensors
Since 2012, reaction-based genetically encoded fluorescent H2S sensors have been studied, and a probe based on p-azidophenylalanine (pAzF) was originally developed and tested by expressing the cpGFP-Tyr66pAzF in Hela cells (Chen et al., 2012). This pAzF-based genetic probe has been optimized by modification with pAzF of the chromophore of a circularly permutated, superfolder green fluorescent protein (cpsGFP) to derive the cpsGFP-pAzF, which subsequently served as a Förster resonance energy transfer acceptor to an enhanced blue fluorescent protein EBFP2 (Youssef et al., 2019).
Thus far, the H2S genetic probe has not been utilized in plant research, whereas other genetic sensors, such as HyPer or the roGFP series for hydrogen peroxide (H2O2) sensing, and SoNar or Peredox for NADH/NAD+ sensing (Müller-Schüssele et al., 2021), have been extensively used for monitoring the redox states in living plant cells. The development or application of genetic probes for real-time H2S monitoring in plant research would be greatly beneficial for understanding the H2S-regulatory mechanisms in plants.
H2S Signaling via Protein Persulfidation
Protein persulfidation, also called S-sulfhydration, as a type of oxidative PTM of cysteines, has been increasingly recognized as the main redox mechanism directly regulating diverse biological processes in H2S signaling, such as protein function, structure, and subcellular localization. Protein cysteine thiols (–SH) are very susceptible to H2O2 and can undergo different oxidative PTMs. Initially, thiol reacts with H2O2 to form sulfenic acid (–SOH) (Allison, 1976) that is intrinsically unstable and an intermediary en route to other PTMs.
In the presence of H2O2 excess, –SOH forms the relatively more stable sulfinic acid (–SO2H) and sulfonic acid (–SO3H) (Cremlyn, 1996), which are generally considered to cause protein overoxidation (Fig. 4). The –SO3H formation is irreversible, whereas –SO2H can be reduced via an ATP-dependent reaction by sulfiredoxin of certain proteins (Akter et al., 2018; Biteau et al., 2003).

Alternatively, –SOH can react with proximal –SHs from proteins and with GSH, forming intra/intermolecular disulfide (–SS–) (Nagy and Winterbourn, 2010; Turell et al., 2021) and S-glutathione adduct (–SSG) (Turell et al., 2008), respectively (Fig. 4). Disulfides and GSH adducts can be enzymatically reduced to –SH by thiol reductases, so-called redoxins, such as TRXs and GRXs (Huang et al., 2018; Willems et al., 2021). Besides H2O2, reactive nitrogen species, which mainly refer to NO, trigger the formation of S-nitrosothiols (–SNO) (Hess et al., 2005) that can also be reduced by redoxins.
H2S reacts with oxidized, not reduced, thiols, –SOH, and disulfides specifically, to form persulfides (–SSH) (Cuevasanta et al., 2015). The kinetics of the H2S reactions with low-molecular weight albumin disulfides and –SOH have been determined. The rate constant of H2S with –SOH for the formation of persulfides is 270 M −1s−1, significantly higher than that of disulfides (0.6 M −1s−1), implying that the formation of protein persulfides might mainly occur through reaction of H2S with –SOH. However, extremely high reaction rates of protein disulfides with H2S have been detected in some special cases, such as human sulfide quinone oxidoreductase (Cuevasanta et al., 2017).
Furthermore, protein disulfides are relatively more stable than the labile –SOH in the environment; hence, the generation of protein persulfides via the H2S reaction with disulfides cannot be excluded. Persulfides might be formed by reaction of H2S with –SNO or –SSG (Francoleon et al., 2011; Iciek et al., 2015; Mishanina et al., 2015), but this reaction mainly remains hypothetical and needs further investigation.
Protein persulfides can be oxidized by H2O2 to form perthiosulfenic acid (–SSOH), perthiosulfinic acid (–SSO2H), and perthiosulfonic acid (–SSO3H). In contrast to the hardly reversible –SO2H and irreversible –SO3H formed upon H2O2 overoxidation, the corresponding –SSH and its oxidative derivatives can be reduced by redoxins (Dóka et al., 2020; Filipovic, 2015; Ju et al., 2016; Wedmann et al., 2016) (Fig. 4).
Protein Persulfidation Detection in Plants
In the past decade, a variety of detection methods for efficient persulfidation labeling and identification have been developed to investigate the H2S signaling executed via persulfidation. Protein persulfidation can be directly detected by means of mass spectrometry (MS), because of the mass increase of 31.972 Da by the addition of one sulfur atom, but is difficult to distinguish from other modification, such as –SO2H due to the addition of two oxygen atoms (mass increase of 31.99 Da). Thus, specific labeling with chemical probes is required for persulfidation detection.
Initially, a modified biotin switch method had been applied in human cells, in which methyl methanethiosulfonate (MMTS) was believed to specifically block –SH, whereafter –SSH was targeted and enriched with N-[6-(biotinamido) hexyl]-3′-(2′-pyridyldithio)propionamide (biotin-HPDP) (Mustafa et al., 2009). In plants, this method was first utilized in Arabidopsis leaf extracts, and 106 persulfided proteins were identified (Aroca et al., 2015). However, because the reactivity toward MMTS of SSH was higher than that of thiols (Pan and Carroll, 2013), this method was questioned and should be used with caution.
The most challenging aspect of persulfidation detection is the discriminative labeling from thiols, because of their similar reactivity to commonly used reagents through alkylation, such as maleimide, N-ethylmaleimide (NEM), and iodoacetamide (IAM) (Pan and Carroll, 2013). Due to their greater nucleophilicity, persulfides react even much faster than thiols to thiol-labeling reagents (Cuevasanta et al., 2015). Nevertheless, because alkylation of thiols yields thioethers, whereas persulfides generate disulfides; many thiol label-based detection approaches have been exploited based on this characteristic.
At first, a fluorescent probe, designated red maleimide, had been used to study the persulfidation of the p65 subunit of mice NF-κB (Sen et al., 2012). Both −SH and −SSH are labeled with red maleimide, followed by dithiothreitol (DTT) reduction, so that only the labeled −SSH, namely the R−S−S−maleimide-red adducts, are reduced (Fig. 5A). The protein samples are subsequently separated by gel electrophoresis and detected by in-gel fluorescence. The loss of fluorescence can be calculated for evaluation of the persulfidation level. This method revealed the persulfidation of recombinant human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) protein at Cys150, which enhances its catalytic activity (Gao et al., 2015).

Later, another thiol label-based probe, a maleimide compound containing a peptide arm, designated maleimide peptide (MalP, 1.95 kDa), was used to study the persulfidation of the iron–sulfur cluster machinery in mammalian proteins (Parent et al., 2015). As with the red maleimide fluorescent probe, both −SH and −SSH are labeled with MalP, and only the labeled −SSHs are further reduced by the subsequent DTT incubation and release the succinimide-peptide moiety (the product of the maleimide reaction with a sulfhydryl). As a result, the mobility shift in the protein migration can be detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (Fig. 5A).
The two aforementioned approaches offered many advantages, such as relative application simplicity and radiometric read-outs (−SSH vs. −SH), giving extra quantification information. Nevertheless, these methods are restricted to biochemical characterizations and are rather not applicable to large-scale MS-based proteomic analyses.
A MS-coupled thiol labeling-based proteomic approach, termed biotin thiol assay (BTA), had initially been utilized for mapping protein persulfidation in mammalian cells by means of maleimide-biotin (Cuevasanta et al., 2015), maleimide-PEG2-biotin (Gao et al., 2015), or iodoacetyl-PEG2-biotin (Dóka et al., 2019; Dóka et al., 2016). In this method, −SH and −SSH are first labeled with biotin-tagged alkylating reagents that are sequentially enriched on an avidin column, whereafter the labeled −SSHs are selectively eluted by reduction via DTT or tris(2-carboxyethyl)phosphine (Fig. 5B).
By labeling the eluted persulfide-derived thiols by isotope-labeled (D5, heavy) or normal (H5, light) maleimide, a quantitative analysis under different conditions could be achieved (Gao et al., 2015). Recently, a BTA combined with the iodoacetyl isobaric tandem mass tag system allowed the quantitative cysteine site-specific identification of persulfidation (Gao et al., 2020).
In addition, a “tag-switch” method was developed for persulfidation detection (Zhang et al., 2014). Here, methylsulfonyl benzothiazole (MSBT) was used to block both −SH and −SSH, whereafter solely MSBT disulfide adducts (–SS–MSBT) could react with cyanoacetate biotin (CN-biotin), hence designated “tag switch” (Fig. 5C). This method was further improved by attaching cyanoacetate with the fluorescent BODIPY moiety (CN-BOT) or the Cy3 dye (CN-Cy3), so that persulfidation could be visualized in situ by fluorescence confocal microscopy or in-gel fluorescent detection from the cell lysates (Kouroussis et al., 2019; Wedmann et al., 2016).
In plants, the “tag-switch” method was applied in wild-type and des1 mutant Arabidopsis plants (Aroca et al., 2017a). In total, 2015 and 2130 persulfidated proteins were identified in the wild-type and des1 plants, respectively, suggesting that a large fraction of the Arabidopsis proteome undergoes persulfidation even under nonstressed conditions (Aroca et al., 2017a). Recently, the application of the “tag-switch” method on Arabidopsis root tissue identified 5214 –SSH proteins (Jurado-Flores et al., 2021).
A variant of this tag-switch method for the –SSH identification is called the “dimedone-switch” assay (Zivanovic et al., 2019) (Fig. 5C). In this assay, –SSH, –SH, and –SOH first react with 4-chloro-7-nitrobenzofurazan (NBF-Cl), whereafter a dimedone-based probe, such as a biotin-conjugated analog dimedone-based biotin-conjugated analog (DCP-Bio1) (Poole et al., 2007) or azide-conjugated analog DCP-N3, selectively switches with –SS–NBF disulfides. The biotin-tagged persulfides are subsequently enriched, and identified by MS and by in-gel fluorescence or confocal microscopy when DCP-Bio1 and DCP-N3 and Cy5-alkyne click mix are used, respectively (Zivanovic et al., 2019). This dimedone-switch method recently applied in Arabidopsis revealed persulfidation of Cys103 of the autophagy-related protein 18a, thereby activating its phospholipid-binding activity in a reversible manner and, hence, regulating autophagy under endoplasmic reticulum stress (Aroca et al., 2021a).
Currently, a direct persulfidation detection method was developed for proteomic analysis by labeling –SSH at pH 5.0 by means of an alkyne-linked IAM, N-propynyliodoacetamide (IPM) (Fu et al., 2020a) (Fig. 5D). Given the lower pKa of persulfide (4.3) than that of thiol (8.29), persulfides maintain a relatively high reactivity at pH 5.0 when compared with protonated thiols. Hence, efficient labeling of –SSH can be achieved by labeling proteome extracts with IPM at a low pH, resulting in disulfide adducts (SS-IPM), in addition to thioether adducts (S-IPM) due to the unavoidable –SH labeling.
IPM-labeled peptides are further biotinylated by reaction with UV cleavable biotin-azide (Az-UV-biotin) reagents through a click reaction, the addition of biotin facilitating peptide enrichment. Two types of probe-modified peptides, including the disulfide forms derived from –SSH and the thioether forms derived from –SH, can be analyzed by MS (Fu et al., 2020a).
H2S-Mediated Persulfidation in ABA-Regulated Stomatal Movement
Stomata are pores on the epidermis of plant leaves surrounded by a pair of guard cells. Stomatal movement regulates gas and water exchange between the plants and the environment, and is important for plant growth, development, and response to environmental stimuli (Kim et al., 2010). That H2S induced stomatal closure and participated in ABA signaling was first evidenced by application of NaHS in epidermal strips of Vicia faba (broad bean), A. thaliana, and Impatiens walleriana (impatiens) (García-Mata and Lamattina, 2010). In contrast, H2S was reported to cause stomatal opening in Arabidopsis (Lisjak et al., 2010) and pepper (Capsicum annuum) (Lisjak et al., 2011).
The reason of the opposite effects of H2S on stomatal movement remains inconclusive. However, an increasing number of studies subsequently revealed that H2S is a key regulator of stomatal closure triggered by different environmental stresses, such as drought (Jin et al., 2017), cold (Du et al., 2019), and mediated by phytohormones, such as ABA that accumulates under drought stress (Jin et al., 2013), ET (Hou et al., 2016; Liu et al., 2011b), salicylic acid (SA) (He et al., 2020), and jasmonic acid (JA) (Deng et al., 2020) (Fig. 6).

To date, the H2S signaling function has been best characterized by the ABA-regulated stomatal movement. ABA has been generally recognized as eliciting the DES1 expression in guard cells, which increases the endogenous level of H2S, because DES1-mediated H2S production is required for downstream NO signaling (Scuffi et al., 2014) and respiratory burst oxidase homolog (RBOH)-dependent H2O2 signaling (Scuffi et al., 2018) to activate stomatal closure (Fig. 6).
Besides DES1 and RBOH (Shen et al., 2020), abscisic acid insensitive 4 (ABI4) (Zhou et al., 2020) and SNF1-related protein kinase2.6 (SnRK2.6), also known as Open stomata 1 (OST1) (Chen et al., 2020), have been found as key proteins involved in H2S signaling in ABA-regulated stomatal movement (see below). In addition, by means of pharmacological and genetic approaches, phospholipase D and mitogen-activated protein kinase 4 were shown to participate in H2S-mediated guard cell signaling (Scuffi et al., 2018) and to be an important downstream signal of H2S in stomatal movement in response to drought stress, respectively (Du et al., 2019).
Persulfidation on several key proteins in plants has been characterized, such as the critical antioxidant enzyme ascorbate peroxidase 1 (Aroca et al., 2015), the moonlighting cytosolic GAPDH protein (Aroca et al., 2017b), the autophagy-related protein 4 (Laureano-Marín et al., 2020) and 18 (Aroca et al., 2021a). For a recent review, see Aroca et al. (2021b). Here, we focus on the most recent proteins, that is, DES1, respiratory burst oxidase homolog D (RBOHD), OST1, and ABI4, which are involved in H2S signaling in ABA-regulated stomatal closure (Fig. 7).

The ABA level in guard cells under normal conditions remains low but increases upon stress stimuli. When ABA is absent, the protein phosphatase type 2C (PP2C) binds to the SnRK2.6 kinase domain and inhibits the kinase activity by dephosphorylation. In the presence of ABA, it binds to the ABA receptor pyrabactin resistance/PYR-like/regulatory component of ABA receptor (PYR/PYL/RCAR) to form a complex that binds to PP2C and inhibits the catalytic activity of PP2C (Fig. 7), thereby activating SnRK2.6 because of its dissociation from PP2C and autophosphorylation (Soon et al., 2012). The activated SnRK2.6 then transmits the ABA signal by phosphorylating downstream factors, such as RBOH, ultimately inducing rapid ROS production and cellular calcium cation (Ca2+) influx and activating specific ion channels that trigger stomatal closure (Fig. 7).
ABA induces the DES1 expression transcriptionally through an unclear mechanism, thereby inducing the H2S production in guard cells (Scuffi et al., 2014). The ABA-induced H2S production results in the persulfidation of SnRK2.6 at Cys131 and Cys137, which are adjacent to the catalytic loop of the kinase and the pivotal phosphorylation site Ser175 (Chen et al., 2020) (Fig. 7).
Persulfidation of SnRK2.6 promotes its activity and interacts with the transcription factor ABA response element-binding factor 2 (ABF2) that enhances the cytosolic Ca2+ signaling. Persulfidation of SnRK2.6 at Cys131 and Cys137 alters its protein structure, hence bringing the Ser175 residue closer to the phosphate-acceptor Asp140 and improving its activity (Chen et al., 2021). In contrast, NO negatively regulates ABA signaling in guard cells by inhibiting SnRK2.6 through formation of -SNO at Cys137 (Wang et al., 2015).
SnRK2 phosphorylates the downstream transcription factor related to ABI3/VP1-like 1 (RAV1) to inhibit the expression of ABI4 (Feng et al., 2014). ABI4 has been considered as a versatile activator or repressor of its downstream target genes (Wind et al., 2013). The DES1-mediated H2S production has been reported to induce the persulfidation of ABI4 at Cys250, which is essential for the ABI4 function in the regulation of plant responses to ABA (Zhou et al., 2021). The persulfidation of ABI4 at Cys250 enhances its transactivation activity on mitogen-activated protein kinase kinase kinase 18 (MAPKKK18), thereby activating a mitogen-activated protein kinase (MAPK) cascade (Fig. 7). Furthermore, transactivation of DES1 could be mediated by ABI4 persulfidation, hinting at the existence of a regulatory loop.
In addition, we recently demonstrated that H2S-mediated persulfidation is involved in ABA signaling in guard cells by directly regulating the activity of both ROS and H2S-producing enzymes (Shen et al., 2020). Upon ABA treatment, DES1-mediated H2S triggers its autopersulfidation at Cys44 and Cys205, leading to a burst of H2S in guard cells. The accumulation of H2S further induces the persulfidation of RBOHD Cys825 and Cys890, enhancing the RBOHD activity and leading to ROS overproduction (Fig. 7).
NO was demonstrated to negatively regulate Arabidopsis RBOHD by forming -SNO at Cys 890 during the hypersensitive response in plant immunity (Yun et al., 2011). Nevertheless, NO-mediated modifications on RBOHD still need to be investigated regarding the impact on the ABA signaling in guard cells. Here, the interplays between H2S, ROS, and NO involved in the regulatory ABA signaling mechanism in guard cells occur most probably through redox PTMs of key proteins.
Summary and Perspectives
In the past decades, numerous studies revealed the multitasking capacity of H2S that is involved in many physiological and pathological processes in mammals (Dilek et al., 2020; Kimura et al., 2012; Murphy et al., 2019; Shatalin et al., 2011) and growth, development, and response to environmental stimuli in plants (Baudouin et al., 2016; Chen et al., 2011; Jin et al., 2017; Jin et al., 2013; Li et al., 2014a; Zou et al., 2019). Nevertheless, our current knowledge on the molecular mechanisms by which H2S executes its signaling function remains limited.
The emerging studies focusing on H2S-mediated persulfidations in plants, especially the recently reported cases showing the key function of persulfidation in ABA-regulated stomatal movements, have greatly contributed to the understanding of the H2S regulatory mechanism in plants. Although several aspects regarding H2S signaling in plants still await to be assessed, we believe that they will be solved in the near future by the application of advanced techniques, such as quantitative proteomics, real-time imagining, and structural biology.
Intra/interdisulfide formation and S-glutathionylation are known to be the major redox mechanisms that protect –SH from overoxidation, with GSH being the most important low molecular weight antioxidant in the cells. Recently, the reaction rate of SOH with H2S has been reported to be 2700 M −1s−1 (Cuevasanta et al., 2015), which is much faster than that of –SH (21.6 M −1s−1) or that of GSH (2.9 M −1s−1) for the formation of disulfides and -SSG, respectively (Turell et al., 2008). In addition, increased persulfidation has been observed as a response to H2O2 stress in mammalian cells (Wedmann et al., 2016), indicating that the persulfidation chemistry involves the preferential addition of H2S to a –SOH protein.
Furthermore, H2S might play an important role as an antioxidant through persulfidation in protecting thiols from irreversible overoxidations, such as –SO3H. To compare the antioxidant role of H2S-mediated persulfidation with that of other PTMs, such as GSH-mediated S-glutathionylation, several features need to be taken into account, including the cellular concentration of H2S in the microenvironment.
The concentrations of H2O2 and GSH in plant cells have been well documented (Cheeseman, 2006; Hasanuzzaman et al., 2017; Smirnoff and Arnaud, 2019), and measurements at a spatiotemporal resolution of H2O2 and GSH have been improved by means of genetically encoded sensors (Niemeyer et al., 2021; Nietzel et al., 2019; Ugalde et al., 2022; Ugalde et al., 2021a; Ugalde et al., 2021b).
Unfortunately, the information on the cellular H2S content is scarce. Although the available H2S fluorescent probes provide useful tools to detect H2S production in situ, the noninvasive application of chemical fluorescent probes in planta for real-time H2S measurements remains a challenge because plant tissues are rather rigid. In our opinion, the implementation of genetic sensors for spatiotemporal monitoring of H2S levels in cells would greatly advance our understanding of H2S signaling in plants.
Besides GSH, ascorbate, another abundant antioxidant in plant cells, occurs in all subcellular compartments with particularly high levels in the chloroplasts (Smirnoff and Wheeler, 2000). Ascorbate–GSH cycle has been recognized as one of the major redox regulation pathways to detoxify H2O2 in plant cell (Foyer and Noctor, 2011). Although ascorbate was thought to selectively reduce –SNO formation and had been used extensively for the –SNO protein identification in proteomic studies (Willems et al., 2021), it has been found to also reduce –SOH of several proteins, including 1-Cys peroxiredoxin (Monteiro et al., 2007) and the thiol-specific antioxidant enzyme 2 (Anschau et al., 2020) from Saccharomyces cerevisiae and papain from Mus musculus (Zito et al., 2012). Therefore, concern about the selectivity of ascorbate toward –SNO in terms of proteomic analysis is increasing.
More importantly, these findings hint at an alternative pathway of thiol redox regulation via ascorbate. As H2S is known to react with –SOH to form –SSH, ascorbate might presumably affect persulfidation of certain proteins indirectly via reduction of –SOH. Nevertheless, the direct correlation between ascorbate and H2S-mediated persulfidation remains elusive.
In addition to ascorbate itself, ascorbate peroxidase (APX), the enzyme catalyzing the conversion of H2O2 into H2O in the presence of ascorbate, undergoes several types of thiol-based PTMs. The enzymatic activity of cytosolic APX1 in Arabidopsis is enhanced by NO-triggered –SNO formation (Begara-Morales et al., 2014; Yang et al., 2015) or H2S-induced –SSH formation (Aroca et al., 2015) at Cys32. Large-scale proteomic studies revealed that tAPX and sAPX from chloroplast were also sensitive to thiol modifications (Huang et al., 2019; Wei et al., 2020). However, which are the exact types of thiol-based PTMs and how they regulate the biological function of chloroplastic APXs still await to be explored by functional analysis.
To better understand the function of H2S-mediated persulfidation, it is crucial to find out the occupancy of –SSH and its correlation with other thiol-based PTMs. Such a correlation was evidenced by a comparative analysis based on proteomics data between –SSH and –SOH or –SH events in mammalian studies (Fu et al., 2020a; Zivanovic et al., 2019) and between –SSH, –SOH, and –SNO in Arabidopsis (Aroca et al., 2021b; Zhang et al., 2021; Zhou et al., 2020).
Likewise many thiol-based PTMs, –SSH sites have been mapped at the whole-proteome level in mammals (Fu et al., 2020a), but remain uncharted territory in plants. Adoption of advanced proteomic profiling strategies for identification and quantification of the complete repertoire of persulfidation-undergoing Cys sites in plants will prove a promising future endeavor.
Footnotes
Acknowledgment
The authors thank Martine De Cock for her kind support on the article preparation.
Authors' Contributions
J.H. conceived and wrote the article, Y.X. revised and submitted the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Jiangsu Natural Science Foundation for Distinguished Young Scholars (No. BK20220084 to Y.X.), the Fundamental Research Funds for the Central Universities (No. XUEKEN2022002 to Y.X.), and the Research Foundation-Flanders FWO Senior Postdoctoral fellowship (No. 1227020N to J.H.).