
Abstract
Aims:
The relationship between the gut microbiota and cardiovascular system has been increasingly clarified. Fecal microbiota transplantation (FMT), used to improve gut microbiota, has been applied clinically for disease treatment and has great potential in combating doxorubicin (DOX)-induced cardiotoxicity. However, the application of FMT in the cardiovascular field and its molecular mechanisms are poorly understood.
Results:
During DOX-induced stress, FMT alters the gut microbiota and serum metabolites, leading to a reduction in cardiac injury. Correlation analysis indicated a close association between serum metabolite indole-3-propionic acid (IPA) and cardiac function. FMT and IPA achieve this by facilitating the translocation of Nfe2l2 (Nrf2) from the cytoplasm to the nucleus, thereby activating the expression of antioxidant molecules, reducing reactive oxygen species production, and inhibiting excessive mitochondrial fission. Consequently, mitochondrial function is preserved, leading to the mitigation of cardiac injury under DOX-induced stress.
Innovation:
FMT has the ability to modify the composition of the gut microbiota, providing not only protection to the intestinal mucosa but also influencing the generation of serum metabolites and regulating the Nrf2 gene to modulate the balance of cardiac mitochondrial fission and fusion. This study comprehensively demonstrates the efficacy of FMT in countering DOX-induced myocardial damage and elucidates the pathways linking the microbiota and the heart.
Conclusion:
FMT alters the gut microbiota and serum metabolites of recipient mice, promoting nuclear translocation of Nrf2 and subsequent activation of downstream antioxidant molecule expression, while inhibiting excessive mitochondrial fission to preserve cardiac integrity. Correlation analysis highlights IPA as a key contributor among differentially regulated metabolites.
Introduction
Doxorubicin (DOX), a chemotherapy drug developed in the 1960s to combat cancer, is still widely used as a first-line treatment for tumors. Unfortunately, the significant cardiac toxicity of DOX limits its long-term use (Sangweni et al., 2022). A retrospective study showed that the incidence of DOX-related cardiomyopathy sharply increases with cumulative dose, with rates of 5.0%, 26.0%, and 48.0% at cumulative doses of 400, 550, and 700 mg/m2, respectively (Swain et al., 2003). Therefore, chemotherapy-related cardiomyopathy deserves our increased attention.
The heart, an energy-dependent organ rich in mitochondria, is significantly impacted by DOX toxicity, which is closely associated with mitochondrial damage (Songbo et al., 2019). DOX not only causes damage to mitochondrial DNA and interrupts the mitochondrial oxidative electron transport chain (Ashley and Poulton, 2009; Lebrecht et al., 2005) but also leads to an imbalance in the mitochondrial dynamics associated with fission and fusion, resulting in cell apoptosis (Catanzaro et al., 2019). The abnormality in mitochondrial fission and fusion plays an important role in DOX-induced cardiac injury, and inhibiting excessive mitochondrial fission or promoting mitochondrial fusion can alleviate the effect of DOX on the heart (Maneechote et al., 2022).
Innovation
In cardiovascular diseases, we have been overly focused on the heart itself, neglecting the interconnectedness between the heart and other organs throughout the body. In this study, we shifted our perspective from the heart to the gut microbiota and for the first time discovered that maintaining gut microbiota homeostasis through fecal microbiota transplantation (FMT) under doxorubicin stress regulates the balance of cardiac mitochondrial fission and fusion, thus protecting the heart. Furthermore, we demonstrated that these effects are mediated by the redox-related gene Nrf2. This undoubtedly provides evidence for the potential use of FMT as an adjunct therapy for cardiac diseases (Fig. 1).

Nfe2l2 (Nrf2) is a transcription factor with a crucial role in the cellular response to oxidative stress (Ma, 2013). A deficiency of Nrf2 exacerbates DOX-induced cardiac injury and dysfunction, indicating that Nrf2 is an important protective factor against DOX-induced stress (Li et al., 2014). Many drug molecules can exert protective effects by activating Nrf2 (Cheng et al., 2022; Singh et al., 2015). Upregulation of Nrf2 can enhance cellular resistance to oxidative stress and alleviate excessive mitochondrial fission (Dai et al., 2022). We previously demonstrated that yellow wine polyphenol (YWPC) could alleviate DOX-induced cardiac injury by activating Nrf2 and combating cardiac fibrosis (Lin et al., 2019), but how YWPC does this after entering the digestive system still requires elucidation.
Previously, research focused on the direct protective effects of antioxidant drugs on the heart, neglecting that the body is a unified and coordinated whole. With the deepening understanding in recent years, the status of the gut microbiota in the body has gradually been emphasized (Chen et al., 2021). Cardiac toxicity induced by DOX is closely related to the composition and function of gut microbiota (Huang et al., 2022). Our previous research has also shown that DOX decreases the abundance of gut microbiota and changes the proportion of bacterial groups such as Bacteroidetes, leading to fluctuations in the metabolite levels of gut microbiota, with significant changes observed in serum tryptophan metabolites and unsaturated fatty acid levels (Lin et al., 2021). Moreover, we found that YWPC, a powerful protector of the heart, had a greatly weakened effect when the gut microbiota was depleted by antibiotics (Lin et al., 2021).
Considering the low bioavailability of polyphenols and their ability to reach the colon intact (Mithul Aravind et al., 2021), we hypothesize that YWPC protects the heart via remodeling the disturbed gut microbiota induced by DOX, but this requires verification. These issues have aroused great interest, and we are eager to know whether maintaining gut microbiota stability during a DOX attack can reduce DOX-induced heart damage. Fecal microbiota transplantation (FMT) is undoubtedly the best choice to achieve this; it maintains gut microbiota stability by transplanting donor gut microbiota into the recipient's gut, changing the composition of the recipient's gut microbiota. FMT not only makes the recipient's microbiota similar to that of the donor but also allows the transplanted microbiota to exist in the recipient for up to 3 months (Kootte et al., 2017; Li et al., 2016). Moreover, through FMT technology, interventions such as gavage of YWPC can only occur in the donor mice, and the effects of gut microbiota on the recipient mice can be observed separately, excluding the direct interference effect between these interventions and the heart.
The diversity of metabolites produced by the gut microbiota is extensive, and one of the representative substances is the tryptophan metabolite indole-3-propionic acid (IPA). IPA is currently recognized as a molecule with anti-inflammatory and antioxidative properties, exhibiting favorable protective effects in various contexts, including the kidney (Karbownik et al., 2006), nervous system (Owumi et al., 2022), gastrointestinal tract (Li et al., 2021), and cardiovascular system (Xue et al., 2022). As a gut-derived metabolite, IPA enhances the expression of tight junction proteins in intestinal epithelial cells and amplifies the secretion of mucin by goblet cells, thereby reinforcing the barrier function of the intestines (Li et al., 2021). Within the cardiovascular system, IPA curbs atherosclerosis through the promotion of reverse cholesterol transport (Xue et al., 2022). However, whether IPA also plays a role in the context of FMT intervention in DOX-induced cardiac toxicity, as well as the mechanisms underlying the actions of IPA, are aspects that require elucidation in the present study.
Therefore, this study aims to investigate whether FMT can mitigate DOX-induced cardiac injury and elucidate the target mechanisms through which FMT exerts its cardiac protective effects. Building upon this foundation, an attempt will be made to identify key contributors among numerous gut-derived metabolites and utilize them to dissect more intricate underlying mechanisms.
Results
FMT can effectively improve DOX-induced cardiac injury
To clarify the effect of FMT on DOX-induced cardiomyopathy, we utilized DOX to induce heart damage in mice and administered FMT as a treatment. Given the demonstrated cardioprotective effect of YWPC against DOX, we used YWPC as a positive control (Fig. 2A). Echocardiography revealed that both FMT and YWPC significantly improved the left ventricular ejection fractions (LVEF) and left ventricular fractional shortening (LVFS), indicating an improved cardiac contractile function (Fig. 2A, B). Masson's staining showed that DOX increased collagen deposition in the heart, exacerbating cardiac fibrosis, whereas hematoxylin–eosin (HE) staining revealed that DOX increased cardiomyocyte vacuolization and induced cardiomyocyte disarray.
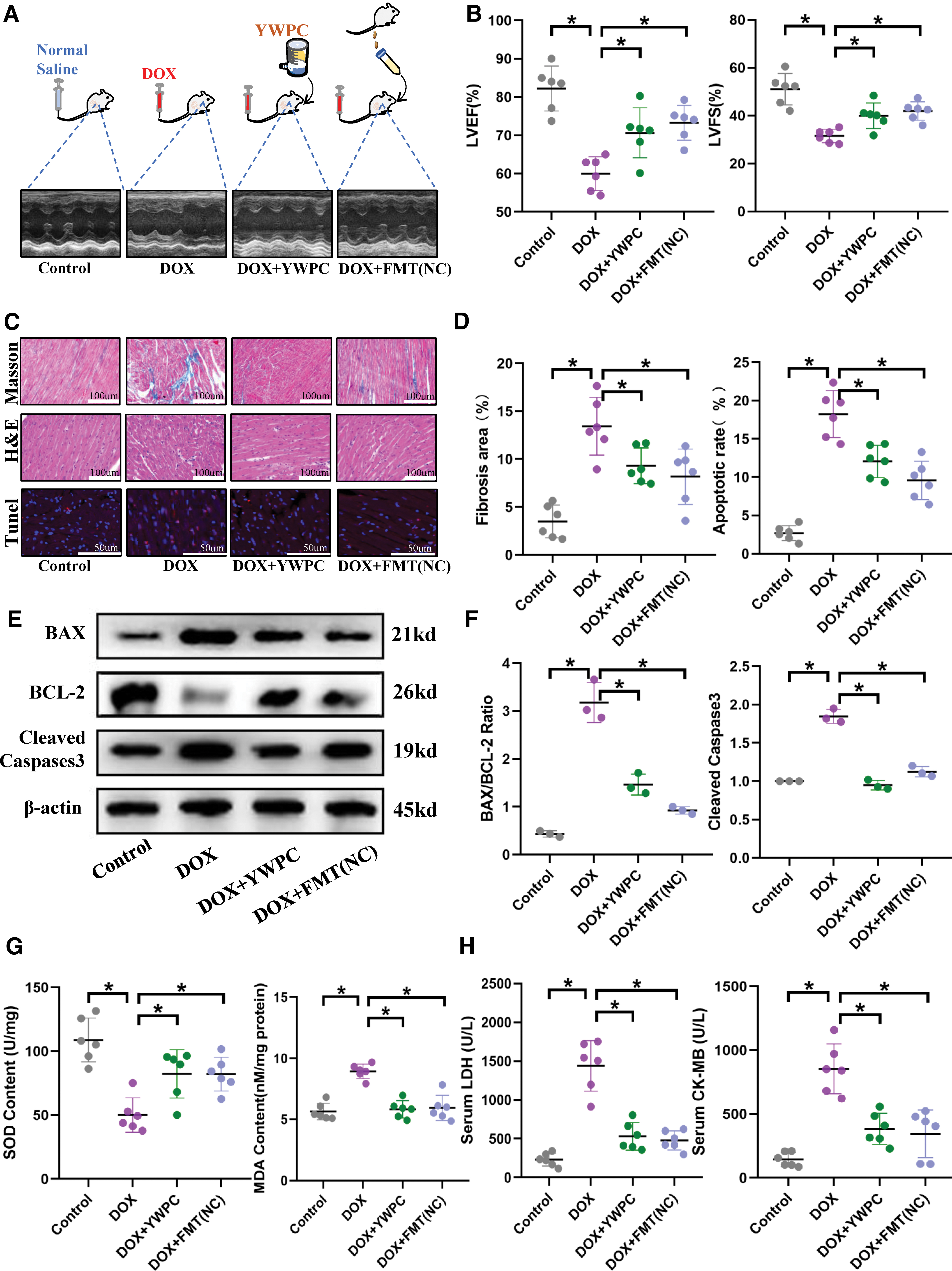
However, both YWPC and FMT significantly reversed these injuries and preserved the integrity of the myocardial structure (Fig. 2C, D). Moreover, YWPC and FMT reduced cardiomyocyte apoptosis (Fig. 2C–F) and attenuated cardiomyocyte damage (Fig. 2H). Furthermore, considering the substantial oxygen consumption of myocardial cells and the well-established property of DOX-induced impairment of mitochondrial respiratory chain, we assessed cardiac oxidative stress markers. As expected, compared with the control mice, DOX treatment significantly upregulated cardiac malondialdehyde (MDA) levels and impaired superoxide dismutase (SOD) content. However, the administration of FMT and YWPC attenuated this effect (Fig. 1G). Overall, the application of either YWPC or FMT alone provided protective effects against DOX-induced cardiac injury.
Gut microbiota plays an important role in protecting the heart
The gut microbiota plays an important role in protecting the heart. To clarify whether maintaining normal microbiota provides effective cardiac protection against DOX attack, we used an antibiotic cocktail to clear the gut microbiota of recipient mice. The Chao index analysis showed that antibiotics (ABX) effectively cleared the vast majority of gut microbiota; even after 4 weeks, if no new microbiota was implanted, the abundance of the original microbiota in the gut remained at a low level (Supplementary Fig. S2). We then transplanted either phosphate-buffered saline (PBS) or normal mouse microbiota into the receptor mice.
Echocardiography showed that the presence or absence of normal microbiota significantly impacts cardiac function under DOX stress. Compared with microbiota depletion, maintaining a normal gut microbiota significantly mitigated the damaging effect of DOX on cardiac ejection function (Fig. 3A, B). Histological examination also showed that under the maintenance of a normal microbiota, cardiac myocyte vacuolization, disorganization, and collagen deposition were significantly reduced (Fig. 3C, D), and myocardial apoptosis was significantly inhibited (Fig. 3C–F). In addition, transplantation of the gut microbiota from healthy mice alleviated oxidative stress levels in the heart (Fig. 3G), ultimately reducing cardiac damage (Fig. 3H).
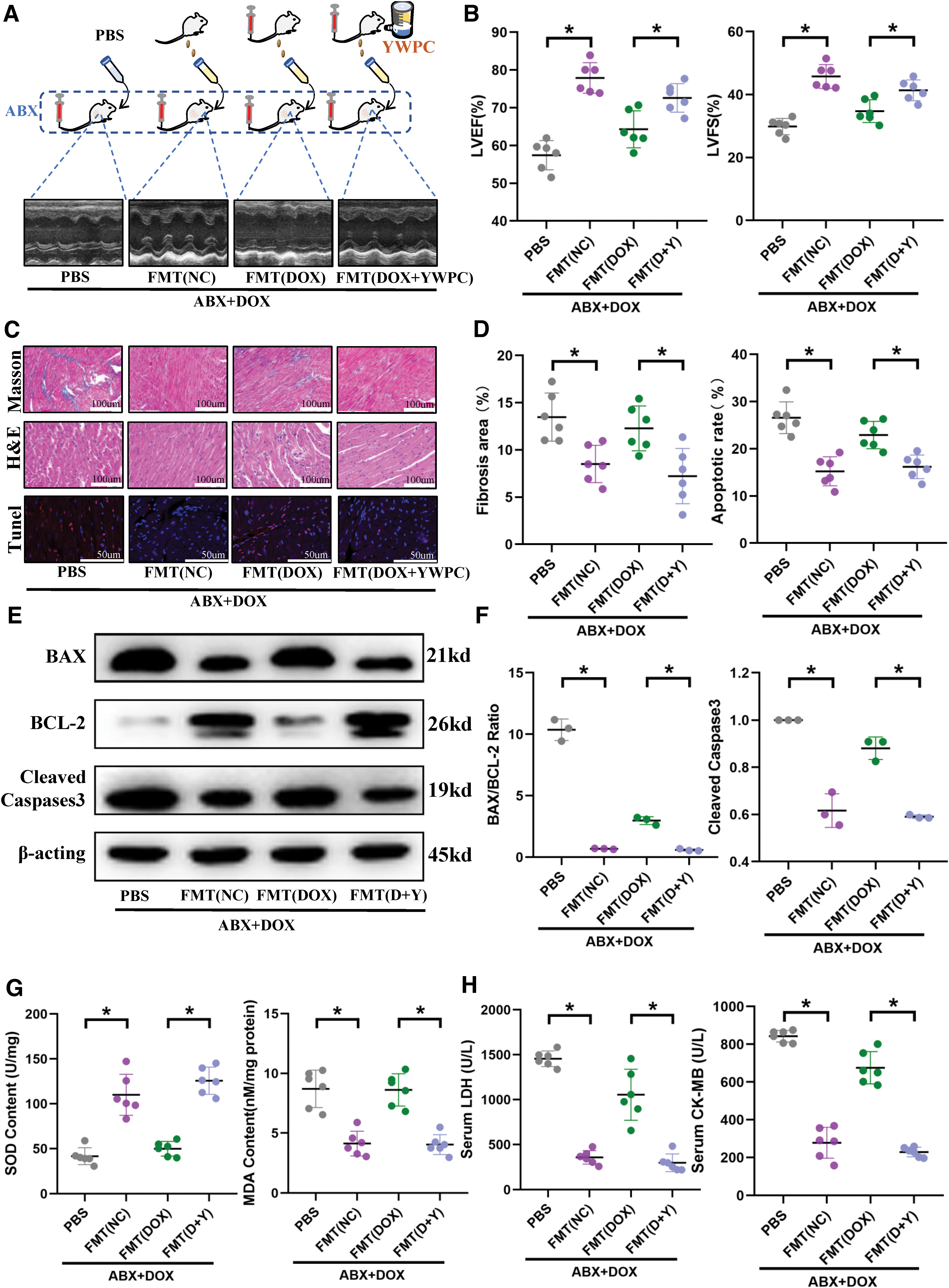
In our previous studies, we have demonstrated that YWPC can reconstitute the disrupted gut microbiota caused by DOX, significantly altering the composition and components of the microbiota, and using this as a basis to protect the heart (Lin et al., 2021). Therefore, in this study, we also transplanted the DOX-disrupted microbiota and the YWPC-remodeled microbiota separately into recipient mice (Fig. 3A) to observe the impact of transplanting different types of microbiota on cardiac function. Echocardiography showed that compared with mice transplanted with microbiota disrupted by DOX alone, mice transplanted with microbiota reshaped by YWPC had a better cardiac ejection fraction (Fig. 3A, B).
Histological examination also showed that transplanting YWPC-reshaped microbiota provided better protection to the cardiac fiber structure, delayed fibrosis, and cardiac remodeling (Fig. 3C, D), reduced the level of myocardial cell apoptosis (Fig. 3C–F), and reduced the DOX-induced damage to cardiac myocytes (Fig. 3H). Similarly, compared with the DOX-disrupted microbiota, the YWPC-remodeled microbiota can mitigate the oxidative stress caused by DOX (Fig. 3G). Therefore, the composition of the microbiota significantly influences the effectiveness of FMT.
In conclusion, we have reached the following findings: FMT significantly influences cardiac function under DOX-induced stress, and its effectiveness is strongly correlated with the composition of the transplanted microbiota. Microbiota derived from healthy mice or reconstituted by the antioxidant YWPC exhibits cardioprotective effects. Therefore, the stability of the microbiota plays a crucial role in heart protection.
FMT significantly alters the gut microbiota structure and protects gut barrier
We then performed 16S rDNA sequencing on the fecal samples of the recipient mice of the FMT to identify differences in the gut microbiota composition among the different groups of mice (Supplementary Table S1). Initially, we conducted principal component analysis (PCA) on fecal microbiota, as shown in Figure 4A, the microbial structure of the ABX+DOX+PBS group differs significantly from the other three groups to such an extent that it requires a large measurement scale, which diminishes the differences between the other three groups (PCA Analysis 1). Therefore, we conducted another PCA (PCA Analysis 2) on the other three groups. The results showed that the gut microbiota structure of the ABX+DOX+FMT (DOX) group was slightly different from that of the ABX+DOX+FMT (NC) group, indicating many differences in the composition of their microbiota.
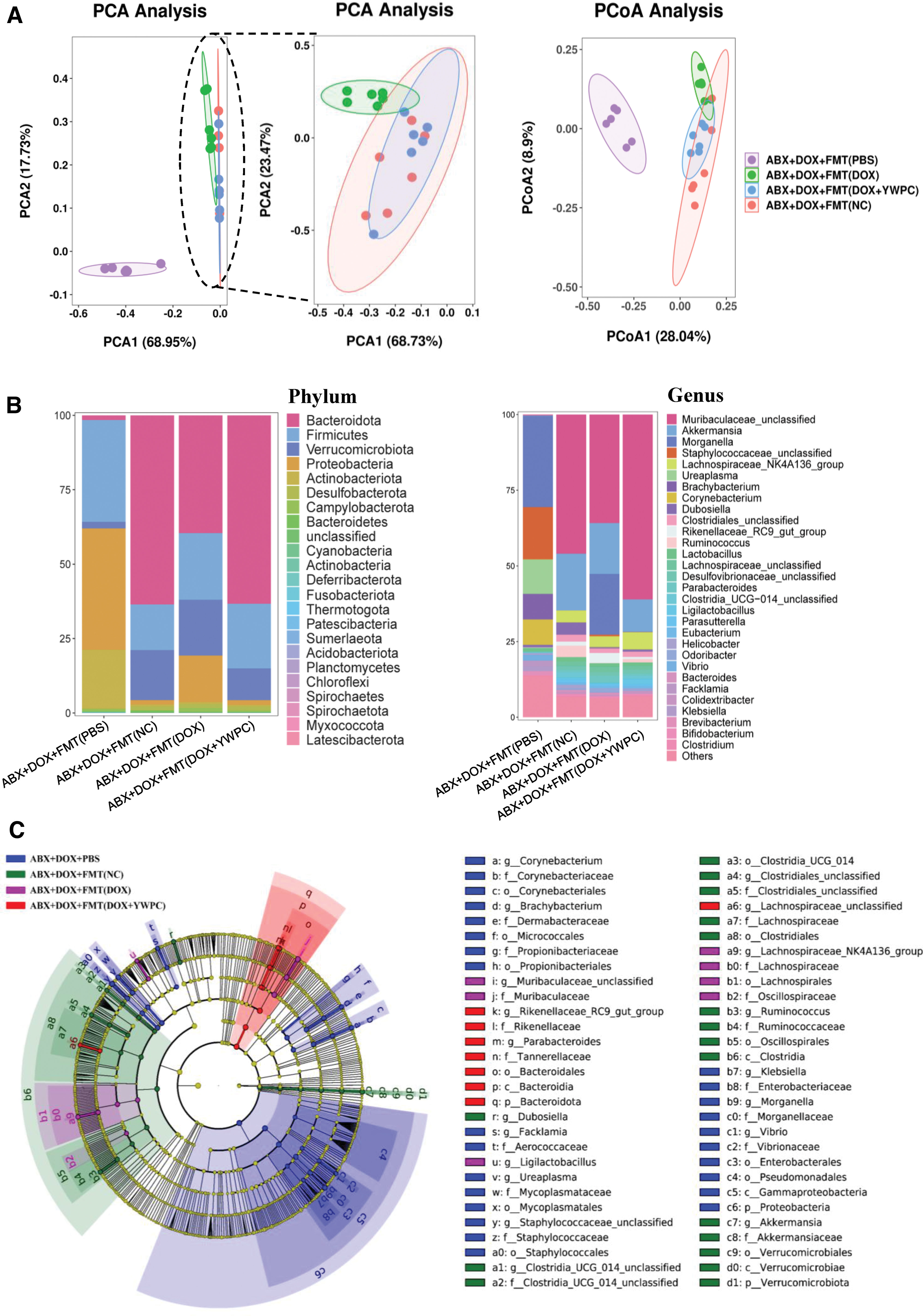
However, surprisingly, the distribution of the ABX+DOX+FMT (DOX+YWPC) group on the coordinate axis was highly consistent with that of the recipient mice transplanted with normal microbiota, suggesting that the reshaping effect of YWPC counteracted the destructive effect of DOX and preserved the normal composition microbiota as much as possible. Similarly, principal coordinates analysis also obtained consistent results (Fig. 4A). We further analyzed the specific components of the microbiota of each group of mice, as shown in Figure 4B. After antibiotic treatment, the remaining stubborn microbiota at the phylum level, Firmicutes and Proteobacteria, accounted for a large proportion in the gut, and the Bacteroidota was almost completely eliminated. Compared with the ABX+DOX+FMT (NC) group, the ABX+DOX+FMT (DOX) group had a lower proportion of Bacteroidota and a higher proportion of Proteobacteria.
However, the microbiota reshaped by YWPC increased the proportion of Bacteroidota and decreased the proportion of Proteobacteria, which was more similar to that of the ABX+DOX+FMT (NC) group. Compared with the ABX+DOX+FMT (NC) group, the ABX+DOX+FMT (DOX) group mainly had a lower proportion of Muribaculaceae_unclassified and a higher proportion of Morganella at the genus level, but these changes were reversed after YWPC reshaping. Then, we used linear discriminant analysis (LDA) to identify specific bacterial groups in the different groups, namely biomarker microbiota. The phylogenetic tree showed that there were specific bacterial groups in all four groups, among which the ABX+DOX+FMT (DOX) group was the most different from the other groups because most of its microbiota abundance had been cleared to very low levels (Fig. 4C). Furthermore, we displayed the representative biomarker bacterial groups according to the LDA score and performed receiver operating characteristic curve (ROC) analysis to explore their ability to distinguish different groups as signature microbiota.
The results showed that p_Proteobacteria, c_Gammaproteobacteria, o_Bacteroidales, and p_Bacteroidota were the most representative biomarker bacterial groups when comparing ABX+DOX+FMT (NC) with ABX+DOX+PBS (Supplementary Fig. S3). When comparing ABX+DOX+FMT (DOX) with ABX+DOX+FMT (DOX+YWPC), the bacterial groups s_Ligilactobacillus_unclassified, g_Ligilactobacillus, s__Rikenellaceae_RC9_gut_group_unclassified, and g__Rikenellaceae_RC9_gut_group were highly representative biomarkers (Supplementary Fig. S4). The ROC analysis of the top three representative bacterial groups for each group yielded very high area under curve values, almost perfectly distinguishing between the different groups (Supplementary Figs. S5 and S6). In conclusion, FMT from different sources can significantly alter the composition of gut bacterial groups, each containing representative bacterial groups.
Moreover, the changes in the gut microbiota structure also led to alterations in the microscopic intestinal tissue structure. Although no significant differences were observed in the colon length, the HE and alcian blue periodic acid schiff (AB-PAS) staining revealed that the group transplanted with healthy mouse microbiota or YWPC-reshaped microbiota significantly preserved the colonic mucosa thickness and number of goblet cells compared with the non-transplant group or group transplanted with microbiota disrupted only by DOX (Supplementary Fig. S7A, B). We also examined the changes in the tight junction protein zonula occludens-1 (ZO-1) in the intestine and found that the transplantation of healthy mouse microbiota or YWPC-reshaped microbiota also upregulated ZO-1 (Supplementary Fig. S7C, D), indicating that intestinal barrier function was protected.
Serum metabolites are important link between the gut microbiota and the heart
Since the main connection between the gut microbiota and the heart is through the bloodstream, we analyzed serum metabolites. PCA showed that there were significant differences in metabolites between the ABX+DOX+PBS group compared with the ABX+DOX+FMT (NC) group and between the ABX+DOX+FMT (DOX) group compared with the ABX+DOX+FMT (DOX+YWPC) group (Fig. 5A, B). The volcano plot showed 655 upregulated and 540 downregulated metabolites in the ABX+DOX+FMT (NC) group compared with the ABX+DOX+PBS group (Fig. 5C), which were mainly enriched in organismal systems-related pathways (Supplementary Fig. S8A). Meanwhile, there were 208 upregulated and 294 downregulated metabolites in the ABX+DOX+FMT (DOX+YWPC) group compared with the ABX+DOX+FMT (DOX) group (Fig. 5D), which were mainly enriched in purine metabolism and other metabolic pathways (Supplementary Fig. S8B).
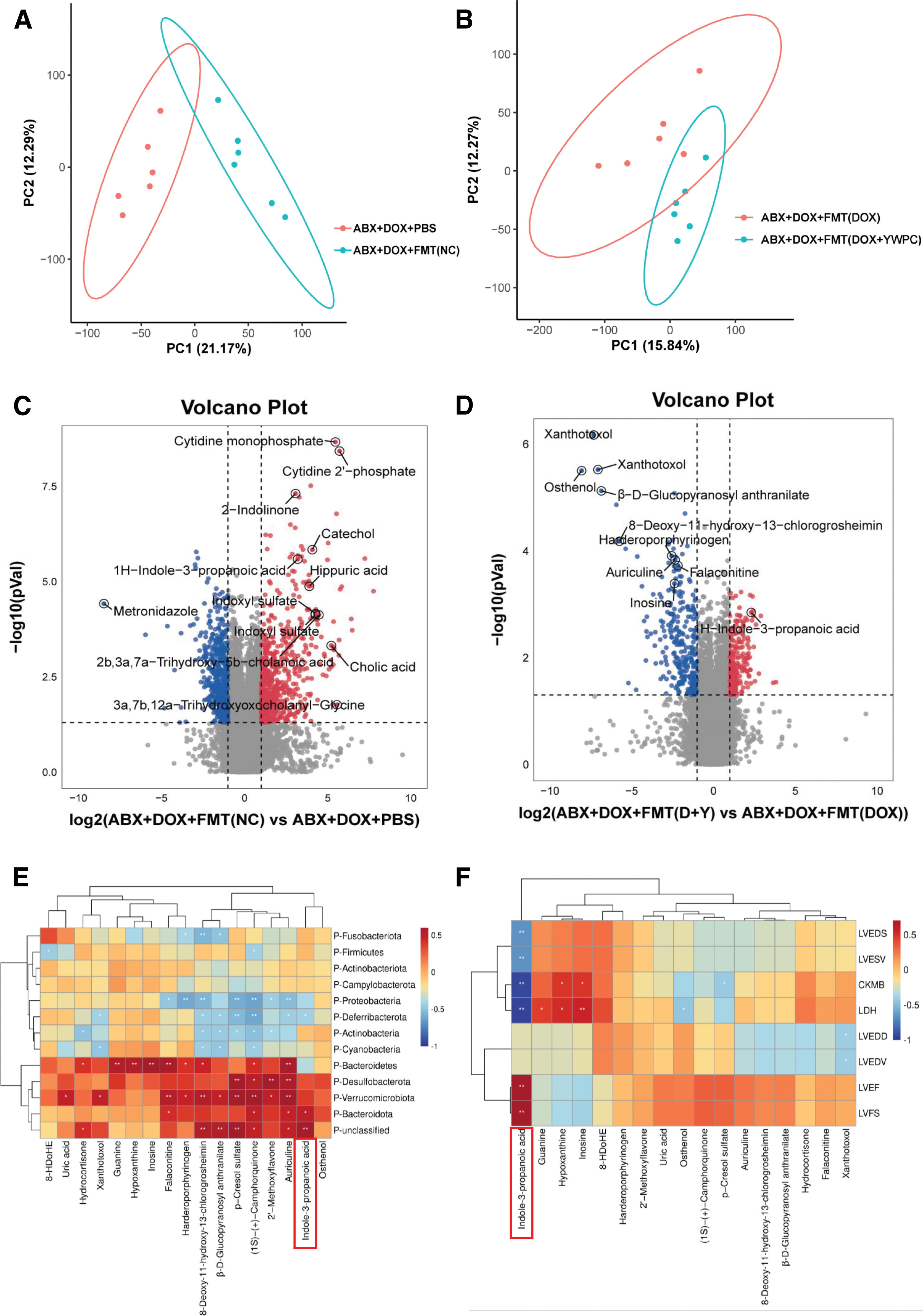
Based on the Spearman correlation analysis, we identified a strong correlation between the gut microbiota and serum metabolites and further revealed potential connections between various serum metabolite components and cardiac function, as well as important metabolites that may play a key role (Fig. 5E, F). The results showed that at least 5 bacterial groups were associated with 17 metabolites. At the phylum level, Bacteroidota, Verrucomicrobiota, and Desulfobacterota showed a positive correlation trend with many metabolites, whereas Proteobacteria, Deferribacterota, and Actinobacteria showed a negative correlation trend with many metabolites.
This suggests a significant relationship between fluctuations in serum metabolites and changes in gut microbiota (Fig. 5E). Furthermore, we explored the interaction between metabolites and cardiac function. Inosine and hypoxanthine were positively correlated with the cardiac injury markers creatine kinase-MB (CK-MB) and lactate dehydrogenase (LDH), whereas IPA exhibited significant cardioprotective effects: the levels of LVEF, LVFS, and IPA showed a significant positive correlation, whereas LDH and CK-MB were significantly negatively correlated with IPA levels (Fig. 5F). These results strongly suggest a close relationship between gut microbiota, metabolites, and cardiac function.
FMT modulates mitochondrial fission–fusion dynamics and impacts mitochondrial function
To further elucidate the target organelle responsible for the cardioprotective effects of FMT and considering the high oxygen consumption of the heart along with DOXs known mitochondrial damage, we assessed changes in mitochondrial respiratory chain function and mitochondrial fission–fusion dynamics under DOX-induced stress and investigated the effects of FMT. We observed that DOX stress led to excessive mitochondrial fission, characterized by reduced mitochondrial size and an overall increase in quantity. However, FMT restored mitochondrial size (Fig. 6A, B). Further assessment of proteins related to mitochondrial fission and fusion revealed that FMT lowered the expression of p-DRP-S616 while promoting the expression of mitofusin 2 (MFN2), resulting in a shift of the mitochondrial fission–fusion balance toward fusion (Fig. 6D, E). Activity tests performed on mitochondrial respiratory chain complexes I–IV demonstrated that DOX disrupted the functions of complexes I and III. However, FMT significantly mitigated these damaging effects (Fig. 6C).

Nrf2 mediates the gut microbiota effect on the heart
Based on our previous research, we investigated whether the important redox gene Nrf2 mediates the effects of the gut microbiota on cardiac function. To begin, to eliminate the impact of Nrf2 knockout on baseline cardiac function, we performed cardiac ultrasound on age-matched wild-type mice and Nrf2−/− mice without any intervention. The results revealed that under physiological conditions, the sole absence of Nrf2 does not influence cardiac function (Supplementary Fig. S10A, B). Previously, from the preceding section, we showed that transplanting normal mouse gut microbiota, which maintained a normal gut microbiota in the recipient mice, effectively protected against DOX-induced cardiac injury.
However, when we performed the same operation on the Nrf2-deficient mice, we found that transplanting normal microbiota did not seem to improve the cardiac ejection function (Fig. 7A, B). Pathological examination of the hearts of Nrf2−/− mice revealed that microbiota transplantation did not alleviate DOX-induced collagen fiber deposition, vacuolization, and disorganization of myocardial cells (Fig. 7C, D). There was also no significant improvement in the myocardial cell apoptosis level (Fig. 7C–F) or cardiac injury markers (Fig. 7H). Similarly, we reevaluated the cardiac oxidative stress, and unsurprisingly, we found that in the absence of Nrf2, even with the maintenance of normal gut microbiota in mice, oxidative stress remains unmitigated (Fig. 7G). This strongly suggests that the cardioprotective effect of FMT requires mediation by the Nrf2.
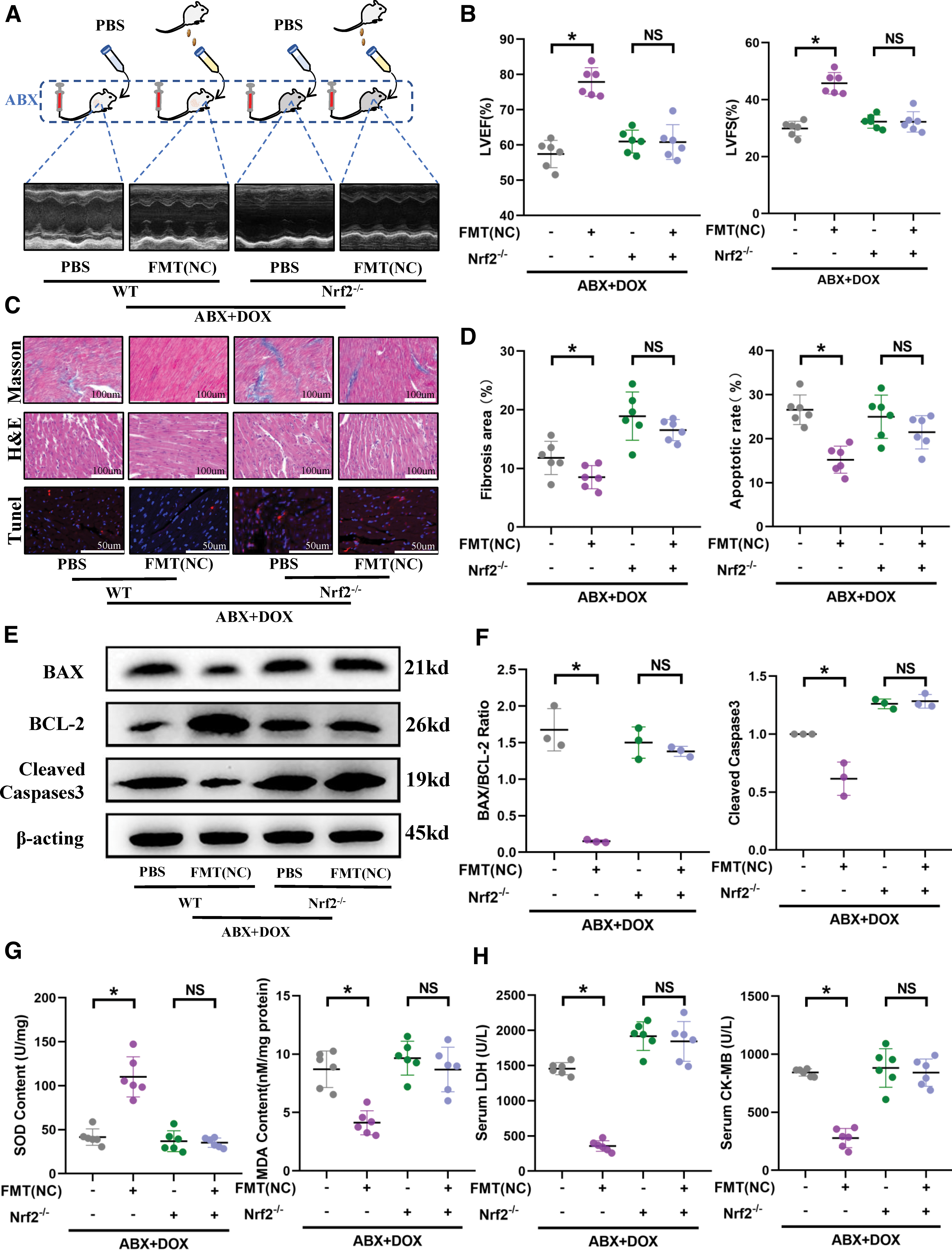
In wild-type mice, compared with the transplantation of microbiota disrupted by DOX, the transplantation of microbiota initially disrupted by DOX but subsequently reshaped by YWPC significantly improved cardiac function. In this context, we transplanted these two distinct compositions of microbiota into Nrf2−/− mice once again. As anticipated, we found that under Nrf2 deficiency, the transplantation of YWPC-reshaped microbiota failed to exert cardioprotection (Supplementary Fig. S9A–H). This once again verifies our hypothesis that the cardioprotective effect of FMT requires mediation by the Nrf2.
FMT-mediated regulation of mitochondrial dynamics and function requires Nrf2 involvement
We further explored whether FMTs regulation of mitochondria is also accomplished through the Nrf2. Initially, we assessed the morphology and function of mitochondria in untreated wild-type mice and Nrf2−/− mice. The results indicated that under physiological conditions, the absence of the Nrf2 gene does not affect mitochondrial dynamics (Supplementary Fig. S10C, D) and mitochondrial respiratory chain complex activity (Supplementary Fig. S10E). Moreover, we found that under DOX-induced stress, mitochondria in Nrf2−/− mice exhibited excessive fission, as expected. However, surprisingly, the beneficial effects of FMT on mitochondrial dynamics and respiratory chain complexes disappeared upon Nrf2 knockout (Fig. 8A–D). This suggests that Nrf2 mediation is required for FMTs protective effects on mitochondria.

FMT facilitates nuclear translocation of Nrf2, leading to the activation of downstream antioxidant molecules
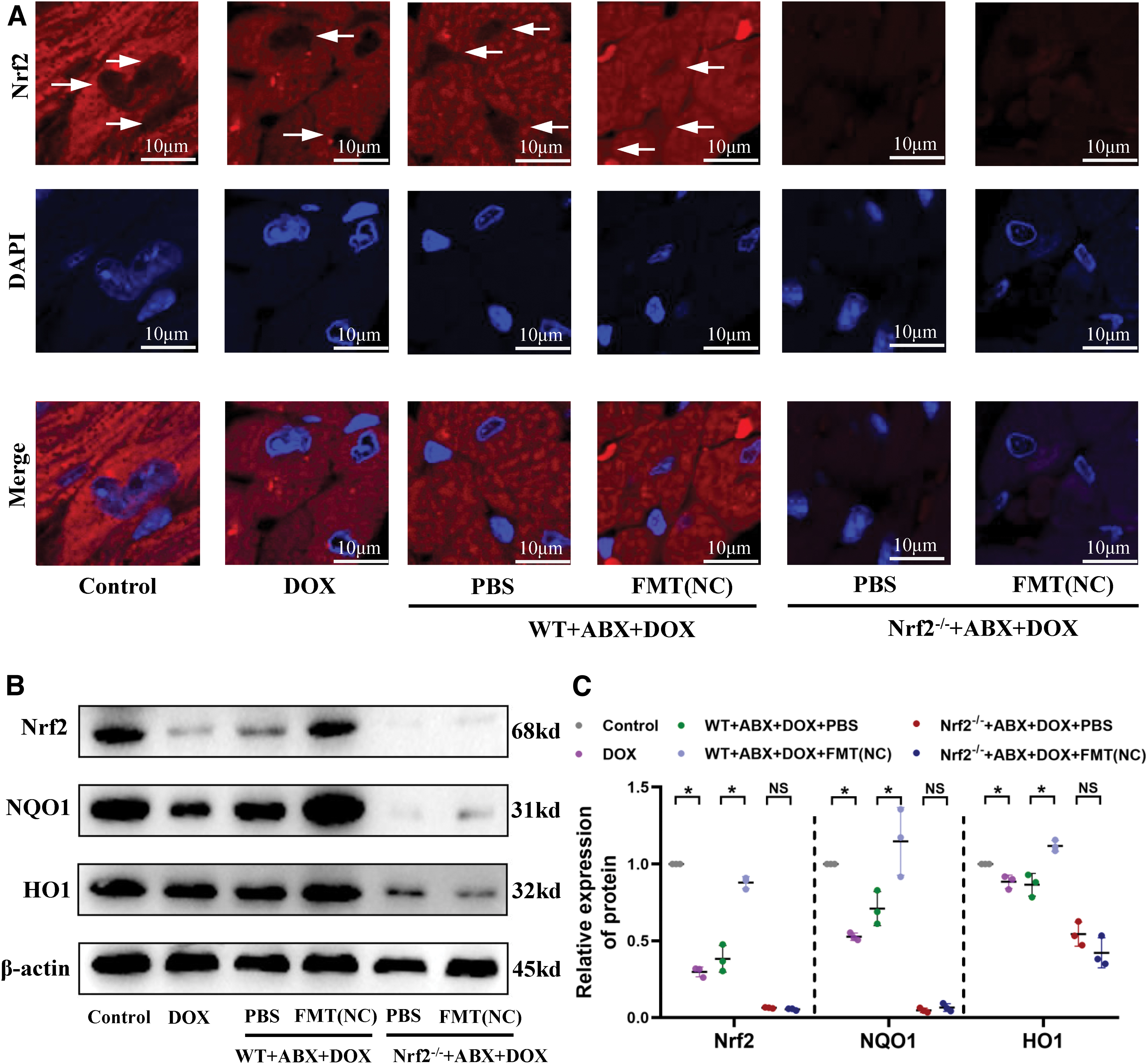
Thus far, we have elucidated that FMT can alter the gut microbiota of recipient mice, influencing serum metabolites, mitochondrial dynamics, and respiratory chain complex function, thereby exerting cardioprotective effects under DOX-induced stress, which require Nrf2 mediation. We further investigated the impact of FMT on Nrf2 synthesis, distribution, and downstream activation. As depicted in Figure 9A, in the control group mice, Nrf2 is primarily located in the cytoplasm. Under DOX stress, insufficient Nrf2 translocates into the nucleus to initiate antioxidant responses. However, FMT significantly promotes Nrf2 translocation into the nucleus (Fig. 9A). Western blot analysis of Nrf2 and its downstream proteins revealed that FMT also upregulates Nrf2 protein levels and stimulates the expression of downstream antioxidant molecules NAD(P)H quinone oxidoreductase-1 (NQO1) and heme oxygenase-1 (HO1) (Fig. 9B, C), thereby enhancing cardiac antioxidant capacity. Consequently, the cardioprotective effects of FMT may indeed rely on the facilitation of Nrf2 expression and nuclear translocation.
Differential metabolite IPA regulates cardiac mitochondrial fusion and fission through Nrf2
In Figure 6F, we analyzed the possible relationship between metabolites and cardiac function. We found a significant correlation between serum metabolite levels and cardiac-related indicators; IPA showed the strongest association, as its expression level was strongly correlated with the LVEF and LVFS. Furthermore, IPA was significantly elevated in groups transplanted with either the normal microbiota or the microbiota remodeled by YWPC (Supplementary Fig. S11A, B). Therefore, we selected IPA as the representative metabolite and administered it to both DOX-treated mice and primary cardiomyocytes to confirm that IPA was the primary effector of gut microbiota metabolites. The results showed that the oral administration of IPA effectively inhibited DOX-induced reductions in the LVEF and LVFS; however, this effect appears to be mediated through the Nrf2 gene, as this cardioprotective effect of IPA vanished upon Nrf2 gene knockout (Fig. 10A, B). Similarly, we also observed that Nrf2 knockout abolished IPAs alleviating effects on myocardial apoptosis and oxidative stress (Supplementary Fig. S12A–C).
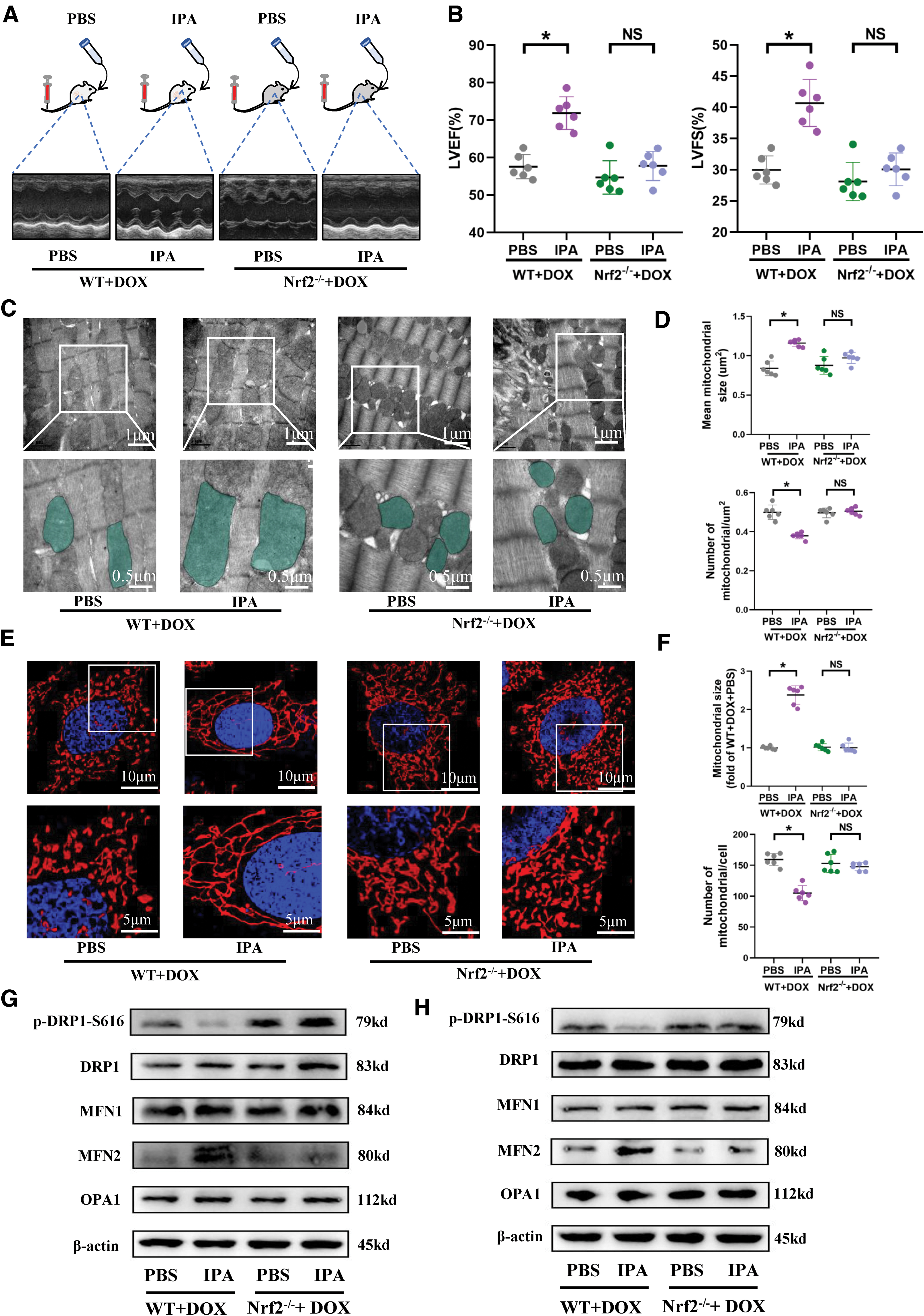
To further elucidate the target organelle of IPA effects, in conjunction with changes in mitochondrial morphology and function following FMT, we reevaluated mitochondrial morphology and function in this context. Transmission electron microscopy revealed that in wild-type mice, IPA intervention reversed DOX-induced excessive mitochondrial fission, increasing mitochondrial size and reducing the quantity of mitochondria per unit area (Fig. 10C, D). Analysis of proteins associated with mitochondrial fission–fusion dynamics indicated that IPA reduced cardiac tissue p-DRP1-S616 and increased MFN2 expression (Fig. 10G and Supplementary Fig. S12E), restoring mitochondrial respiratory chain complexes I and III activity (Supplementary Fig. S12D). However, Nrf2 knockout abolished these effects of IPA (Fig. 10C, D, G and Supplementary Fig. S12D, E). In primary cultured cardiomyocytes, normal mitochondria exhibit elongated rod shapes, and under physiological conditions, the sole absence of the Nrf2 gene does not affect mitochondrial morphology (Supplementary Fig. S13B).
DOX significantly promoted mitochondrial fission, leading to short rod-like forms, whereas incubation with IPA restored mitochondria to their elongated rod-like morphology (Fig. 10E, F). Protein analysis related to mitochondrial fission–fusion dynamics showed that IPA lowered p-DRP-S616 protein expression and promoted MFN2 expression, tilting the balance of mitochondrial fission–fusion dynamics toward fusion (Fig. 10H and Supplementary Fig. S14B). However, in the absence of the Nrf2 gene, IPA inhibition of cardiomyocyte mitochondrial fission became exceedingly weak (Fig. 10E), and p-DRP1 and MFN2 expression showed no significant differences (Fig. 10H and Supplementary Fig. S14B). These findings suggest that the gut microbiota-derived metabolite IPA may exert crucial cardioprotective effects through the regulation of mitochondrial fission–fusion dynamics, with Nrf2 mediation of IPA regulation of cardiac fission–fusion.
IPA facilitates Nrf2 nuclear translocation and activates antioxidant molecules, reducing mitochondrial-derived reactive oxygen species generation and alleviating oxidative stress
We investigated the mechanism through which IPA exerts its antioxidative effects. Under DOX-induced stress, the redox-sensitive fluorophores targeting mitochondrial-derived reactive oxygen species (MitoROS) (red fluorescence) and cellular ROS (green fluorescence) displayed intriguing colocalization patterns (Supplementary Fig. S14A), suggesting that intracellular ROS mainly originate from mitochondria. IPA intervention significantly reduced the generation of MitoROS and cellular ROS. However, this effect of IPA was profoundly weakened when the Nrf2 gene was absent (Supplementary Fig. S14A), implying that IPA inhibition of MitoROS and cellular ROS generation depends on Nrf2.
Further investigations in vivo and in vitro delved into how IPA combats ROS generation through Nrf2. Fluorescent staining of Nrf2 in myocardial tissues revealed that under DOX stress, there was not sufficient Nrf2 translocating into the nucleus, but IPA intervention significantly promoted Nrf2 nuclear translocation. In Nrf2−/− mouse myocardial tissues, Nrf2 was extensively absent (Supplementary Fig. S15A). Western blot analysis of Nrf2 and its downstream proteins revealed that IPA not only increased the total protein level of Nrf2 but also upregulated the expression of downstream antioxidant molecules NQO1 and HO1 (Supplementary Fig. S15B).
A similar Nrf2 nuclear translocation phenomenon was observed in cultured primary cardiomyocytes under IPA intervention (Supplementary Fig. S16A). Furthermore, through nuclear–cytoplasmic fractionation, we found a substantial amount of cytoplasmic Nrf2 translocated to the nucleus after IPA intervention (Supplementary Fig. S16B, C). Collectively, these data suggest that IPA alleviates oxidative stress in myocardial cells by promoting Nrf2 nuclear translocation, stimulating downstream antioxidant molecules NQO1 and HO1, and consequently reducing mitochondrial MitoROS and cellular ROS, thus protecting against oxidative stress.
Discussion
Increasing evidence suggests a close relationship between the gut microbiota and various diseases, including digestive and metabolic disorders (Capparelli et al., 2023; Gerard, 2016) and various cardiovascular diseases (Bhat et al., 2022), where a disrupted microbiota can have adverse consequences. Our study has revealed the therapeutic potential of FMT in DOX-induced myocardial injury and its underlying mechanisms. We found that FMT can alter the structure of the gut microbiota, thereby changing the composition of serum metabolites. Through upregulating known anti-inflammatory and antioxidant molecules such as IPA, promoting Nrf2 translocation to the nucleus, and activating downstream antioxidant molecules, IPA prevents mitochondrial-derived ROS production and inhibits excessive mitochondrial fission, thus achieving myocardial cell protection under DOX-induced stress.
FMT is an effective therapeutic strategy for various diseases, and the transplanted microbiota from donors can coexist with the existing microbiota of the recipients for up to 3 months (Li et al., 2016). FMT effectively treats digestive system diseases and difficult Clostridium difficile infections (Juul et al., 2018). As our previous study has already demonstrated the strong anti-DOX-induced cardiomyopathy effect of YWPC, and the role of YWPC is mediated through gut microbiota (Lin et al., 2021), we used YWPC as a therapeutic reference to evaluate the therapeutic efficacy of FMT. As shown in the Results section, FMT has a strong cardiac protective effect, similar to YWPC. DOX toxicity mainly manifests in cardiomyocyte apoptosis and loss, collagen fiber deposition, and cardiac fibrosis exacerbation, causing a decline in cardiac ejection function (Zhang et al., 2022).
However, we found that FMT can significantly inhibit these effects, and the effect is equivalent to that of YWPC, indicating that FMT is a feasible means to improve DOX-induced cardiomyopathy. At a more microscopic level, DOX also accumulates in mitochondria, causing damage to the mitochondrial electron transport chain function and promoting mitochondrial swelling, which leads to a disorder in energy metabolism in cardiomyocytes. YWPC is capable of resisting these destructive effects of DOX (Lin et al., 2021). Surprisingly, the cardiac mitochondria of the mice receiving FMT were unaffected by the above damage, which confirms the effectiveness of FMT and further suggests that the cardiac protective mechanism of FMT may be similar to that of YWPC.
To further reveal the cardioprotective mechanisms of FMT, separate studies on the transplanted microbiota are needed. We eliminated most of the original microbiota in the receptor mice using antibiotics and transplanted the microbiota from standardized mouse models. As expected, the microbiota in FMT had a distribution similar to that of the microbiota from YWPC remodeling. The gut microbiota after FMT had a larger proportion of Bacteroidota and a lower proportion of Proteobacteria. These two types of bacteria can serve as biomarkers to distinguish whether the FMT intervention was performed. Bacteroidota dominates the gut microbiota of healthy individuals and participates in human metabolic processes, while obese patients have a lower proportion of Bacteroidota (Ley et al., 2006). Proteobacteria account for only a small fraction of gastrointestinal bacteria but usually comprise a higher proportion in patients with gastrointestinal diseases (Pflughoeft and Versalovic, 2012).
At the genus level, Muribaculaceae, a beneficial bacterium of the phylum Bacteroidetes, effectively inhibits the growth of pathogenic bacteria in the gut by competing for specific nutrients and has a protective effect on bodily health (Pereira et al., 2020). Our study showed that the proportion of Muribaculaceae was higher in the receptor mice transplanted with microbiota from healthy mice or YWPC remodeling. The disrupted gut microbiota activates inflammation and disrupts the intestinal mucosal barrier, leading to the translocation of bacteria and harmful substances into the blood and even the “waterfall effect” release of inflammatory mediators (Gumus et al., 2013). Our results suggest that whether transplanting microbiota from healthy mice or YWPC remodeling, significant changes in the structure of the gut microbiota can be made, stabilizing the intestinal environment by restoring the levels of some beneficial bacteria, with a great protective effect on the intestinal barrier.
The pathways linking the gut microbiota and the host include metabolites, immune regulation, and neural regulation (Cryan et al., 2019); the metabolites produced by the gut microbiota are critical mediators of the host–microbiota interaction (Krautkramer et al., 2021). Studies on the metabolic pathways have indicated that trimethylamine/trimethylamine N-oxide, short-chain fatty acids, and secondary bile acids production influence the development and progression of cardiovascular diseases, including heart failure (Tang et al., 2019). The Spearman correlation analysis showed that at least five phylum-level gut microbial categories were correlated with metabolite levels.
In the analysis of the correlation between metabolites and cardiac function, we were surprised to find that IPA level was strongly positively correlated with cardiac ejection fraction and negatively correlated with the extent of cardiac injury, implying that IPA is the primary effector of cardioprotection. This is expected since IPA, a derivative of tryptophan metabolism, was enriched in the serum of mice with diabetic cardiomyopathy treated with YWPC in our previous research (Yang et al., 2022), and a considerable body of research has demonstrated that IPA derived from the gut microbiota has potent anti-inflammatory and antioxidant properties (Xiao et al., 2020; Zhuang et al., 2023). Furthermore, clinical studies have reported an inverse relationship between the occurrence of atherosclerotic events and IPA concentration (Ho et al., 2022).
As energy-intensive cells, myocardial cells rely heavily on the homeostasis of mitochondria. Mitochondrial fusion and fission are important mechanisms for maintaining mitochondrial function and cell survival (Chan, 2020). Mitochondrial fusion optimizes mitochondrial activity by allowing the exchange of contents between them, and forming a network of fused mitochondria facilitates energy transfer to the cellular core (Skulachev, 2001). Optic atrophy 1 (OPA1), mitofusin 1 (MFN1), and MFN2 are molecules that mediate the fusion process. Mitochondrial fission is also important for maintaining the proper distribution and quantity of mitochondria in daughter cells and is beneficial in situations where large amounts of energy are required (Adebayo et al., 2021).
However, excessive mitochondrial fission is often observed under stress conditions (Hernandez-Resendiz et al., 2020), and DOX-induced stress causes excessive mitochondrial fission (Ding et al., 2022). Excessive fission can lead to changes in mitochondrial morphology and skeleton, affecting their function and stability and ultimately triggering mitochondrial-mediated apoptotic pathways that lead to cell death (Westermann, 2010). There is a link between excessive mitochondrial fission, diabetic cardiomyopathy, and myocardial ischemia–reperfusion injury, with DRP1 being the main protein that promotes mitochondrial fission (Ding et al., 2018a; Forte et al., 2021). To elucidate the mechanism underlying myocardial cell protection by FMT, we investigated its impact on mitochondrial dynamics and function.
Our findings demonstrate that FMT effectively suppresses excessive mitochondrial fission under DOX-induced stress. Moreover, it downregulates p-DRP expression while upregulating MFN2 content, thus restoring a balanced state of mitochondrial fission and fusion. Furthermore, we observed that mitochondria are a major source of intracellular ROS, and the key metabolite IPA, which plays a predominant role, reduces ROS production from mitochondria, and curbs excessive mitochondrial fission. Consequently, we propose that the gut microbiota, through its metabolites, counteracts ROS generation from mitochondria and restrains excessive mitochondrial fission, thereby safeguarding the heart under DOX-induced stress.
Research in this field has mainly focused on the direct impact of FMT on cardiovascular outcomes, with little exploration of the underlying molecular mechanisms. Our previous research demonstrated that YWPC can protect the heart by activating Nrf2 (Lin et al., 2019). As an important redox-sensitive gene, Nrf2 exerts antioxidant effects through various pathways, including promoting glutathione synthesis, activating glutathione S-transferase, peroxiredoxin systems, and classical antioxidant enzymes such as SOD-1, catalase, and glutathione peroxidase, and inducing NQO1 and HO1 expression; it plays a protective role in many cardiovascular and neurological diseases (Chen and Maltagliati, 2018; George et al., 2022). Our findings indicate that the effect of FMT appears to be greatly diminished in the absence of the Nrf2.
In-depth analysis of this crucial phenomenon revealed that both FMT and the predominant metabolite IPA exert their antioxidative effects by promoting Nrf2 translocation to the nucleus and activating the downstream expression of antioxidant molecules NQO1 and HO1. Therefore, we believe that FMT, through its metabolite IPA, facilitates Nrf2 nuclear translocation and the activation of antioxidant molecule expression, mitigating ROS production from mitochondria. Consequently, it significantly alleviates cardiac oxidative stress, disturbances in mitochondrial dynamics and function, thereby safeguarding cardiac functionality.
The limitations and advantages of this study are worth discussing. First, there are certain differences between the gut microbiota of mice and humans, which means that these research findings need to be further supported by more relevant clinical trials. However, it is worth acknowledging that studying mice allows for better control of standard clean diets and uniform specific pathogen-free (SPF)-level living environments, which significantly reduces the variation in microbiota caused by factors such as diet and environment between individuals. Second, germ-free mice are the best recipients for FMT research, but our method of using antibiotics to clear gut microbiota has also been widely used in various related studies of gut microbiota, and the use of germ-free mice for validation is one of our next research plans. In addition, to obtain a healthy microbiota, the control group, one of the microbiota donors, did not undergo any tail vein injection sham procedure.
However, the tail vein injection performed on the other donor groups, which could potentially cause microbiota disturbance due to the stress response induced by the procedure itself, might influence the cardiac function of recipient mice; we plan to conduct relevant experiments in the future to validate this possibility. It is worth noting that the intervention measures disrupting the microbiota, such as DOX-induced disturbance and YWPC-mediated remodeling of the disrupted microbiota, occur in the donor mice, the microbiota acts on the heart, and this process occurs in the recipient mice, which isolates the direct intervention effect of YWPC on the heart, making the microbiota the only variable. Using this advantage, we have demonstrated that the process from microbiota to the heart relies on the Nrf2 gene. Finally, this study validated the differential metabolites exerting the main effect through IPA; however, the microbiota that exerts the main effect still needs to be further isolated and individually validated from numerous bacterial species, which will deepen our understanding of the gut ecosystem.
Conclusion
FMT alters the gut microbiota and serum metabolites of recipient mice, promoting nuclear translocation of Nrf2 and subsequent activation of downstream antioxidant molecule expression, while inhibiting excessive mitochondrial fission to preserve cardiac integrity. Correlation analysis highlights IPA as a key contributor among differentially regulated metabolites.
Materials and Methods
Intervention of donor mice and fecal microbial preparation
All experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals and approved by the Experimental Research Ethics Committee of Shaoxing People's Hospital. Healthy 5-week-old male C57BL/6J mice were purchased from Shanghai SLAC Laboratory Animal Co., Ltd. Five-week-old Nrf2-knockout mice (Nrf2−/−) (Supplementary Fig. S1A) were purchased from Jiangsu Jicui Yaokang Co., Ltd., and housed in an SPF animal laboratory. They were provided with SPF-grade irradiated sterilized feed (Xietong Biotechnology, Nanjing, Jiangsu) and were allowed to acclimatize for 1 week before grouping.
Donor mice used to provide the microbiota were divided into three groups of six mice each: the normal control (NC), DOX, and DOX+YWPC groups. The NC group received no treatment. The DOX group received weekly tail vein injections of DOX (4 mg/kg) (MedChemExpress, NJ) for four consecutive weeks (Lin et al., 2021). The DOX+YWPC group received oral administration of YWPC (30 mg/kg/day in water) for 8 weeks and tail vein injections of DOX (the same dosage and frequency as the DOX group) for the last 4 weeks (Lin et al., 2021). To eliminate the effect of age on the microbiota, fresh feces were collected from all groups of mice at 15 weeks of age and stored at −80°C; the collection process lasted for 1 week. For the FMT, 500 mg of feces was resuspended in 5 mL of PBS buffer, and the supernatant was collected after centrifugation. Male recipient mice at 8 weeks of age received one oral gavage (200 μL) of fecal microbiota each day for three consecutive days in the first week of colonization. Then, the fecal microbiota was introduced every other day for the next 3 weeks to enhance colonization (Supplementary Fig. S1B) (Liu et al., 2020).
Intervention of recipient mice
To validate the effectiveness of YWPC and FMT in alleviating DOX-induced cardiomyopathy, we established four groups (n = 6): the control group received an equal volume of normal saline instead of DOX via tail vein injection; the DOX group received weekly tail vein injections of DOX (4 mg/kg) for four consecutive weeks; the DOX+YWPC group received daily oral administration of YWPC (30 mg/kg/day in water) for 8 weeks, followed by tail vein injections of DOX (as above) for the last 4 weeks; the DOX+FMT (NC) group received tail vein injections of DOX (as above) and oral gavage of fecal microbiota from the NC mice for 4 weeks (as above) (Fig. 2A).
To validate the protective effect of maintaining the normal microbiota and reshaping the DOX-disturbed microbiota using YWPC, we established four groups (n = 6). Initially, all mice received drinking water containing multiple antibiotics (0.5 g/L vancomycin, 1 g/L neomycin, 1 g/L metronidazole, and 1 g/L ampicillin) for 14 consecutive days to deplete the original gut microbiota (Zarrinpar et al., 2018), followed by tail vein injections of DOX (as above) for the next 4 weeks. During the 4 weeks of DOX injections, the ABX+DOX+PBS group received gavage of PBS solution (same dose and frequency as FMT), the ABX+DOX+FMT (NC) group received transplanted fecal microbiota from the NC donors, the ABX+DOX+FMT (DOX) group received transplanted fecal microbiota from the DOX donors, and the ABX+DOX+FMT (DOX+YWPC) group received transplanted fecal microbiota from the DOX+YWPC donors (Supplementary Fig. S1B) (transplantation method as described above).
To verify whether Nrf2 mediates the microbiotic effect on DOX-induced dysbiosis and subsequent cardioprotection, we established four groups (n = 6). Initially, we cleared the intestinal microbiota of all WT/Nrf2−/− mice with an antibiotic cocktail (as above) and administered DOX via tail vein injection (as above). During the 4 weeks of DOX injection, the WT+ABX+DOX+PBS group and the Nrf2−/−+ABX+DOX+PBS group were gavaged with PBS solution (same dose and frequency as FMT), whereas the WT+ABX+DOX+FMT (NC) group and the Nrf2−/−+ABX+DOX+FMT (NC) group were transplanted with fecal microbiota from NC mice (transplantation method as described above) (Fig. 6A).
To investigate whether the YWPC remodeling of DOX-induced dysbiotic microbiota exerts cardioprotective effects mediated by Nrf2, we established four groups (n = 6). Similarly, we first depleted the gut microbiota of WT/Nrf2−/− recipient mice using an antibiotic cocktail therapy (as described above) and then injected them with DOX via the tail vein (as above). During the 4 weeks of DOX injection, both the WT+ABX+DOX+DOX and Nrf2−/−+ABX+DOX+DOX groups were orally gavaged with DOX donor mouse fecal microbiota (at the same dose and frequency as above). In addition, the WT+ABX+DOX+FMT (DOX+YWPC) group and the Nrf2−/−+ABX+DOX+FMT (DOX+YWPC) group received DOX+YWPC mouse FMT (as described above) (Fig. 7A).
To verify whether IPA is the primary metabolite responsible for the observed effects, we established four groups (n = 6) of all WT/Nrf2−/− mice injected with DOX via the tail vein (as above). During the 4 weeks of DOX injections, the WT+DOX+IPA group and the Nrf2−/−+DOX+IPA group were given drinking water containing 0.1 mg/mL IPA (Alexeev et al., 2018), whereas the WT+DOX+PBS group and the Nrf2−/−+DOX+PBS group were given drinking water with an equal weight of PBS powder added (Fig. 8A).
Measurement of cardiac injury markers, oxidative stress, and antioxidant enzyme activity
Following anesthesia of the mice with isoflurane, blood was promptly collected from the inferior vena cava. After blood coagulation, centrifuge to collect the separated serum. The levels of the cardiac injury markers LDH and CK-MB were determined using commercial assay kits according to the manufacturer's instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Part of fresh cardiac tissue was weighed, and tissue homogenates were prepared on ice. Measurements were conducted using a lipid peroxidation MDA assay kit and a SOD assay kit (Nanjing Jiancheng Bioengineering Institute). The procedures followed the instructions provided by the kit manufacturers.
Echocardiography
After anesthetizing the mice with isoflurane and shaving the chest fur in the thoracic area, the LVEF and LVFS were measured using the VisualSonics Vevo 2100 Imaging System (Toronto, Canada).
Histological examination
After euthanizing the mice with isoflurane, the hearts and colons were quickly removed and fixed in 10% formalin. Following tissue gradient dehydration and paraffin embedding, 5-μm-thick sections were prepared. The heart and colonic morphology were examined using the HE staining kit (Beyotime, Haimen, China), and the myocardial collagen deposition was evaluated using a Masson's trichrome staining kit (Maixin, Fuzhou, China). The number of colonic goblet cells was determined using the AB-PAS Staining Kit (Solarbio, Beijing, China). Images of the stained slides were captured using a Leica DM3000 microscope (Leica, Wetzlar, Germany), and image quantification was performed using the Image-Pro Plus software (NIH, Bethesda, MD).
TUNEL staining
Tissue sections were deparaffinized in xylene and rehydrated in a series of graded alcohols. The sections were then incubated with proteinase K at 37°C for 20 min, followed by incubation with the TUNEL reaction mixture (Roche, Basel, Switzerland) at 37°C for 1 h. The nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). Apoptotic cells were quantified by counting the number of TUNEL-positive cells using a Nikon Eclipse Ti-U fluorescence microscope (Tokyo, Japan).
Transmission electron microscopy
The apex of the heart was cut into small fragments with a long axis of ∼1 mm. The fragments were fixed with 2.5% glutaraldehyde and 1% osmium tetroxide, sequentially washed, dehydrated with acetone, permeated with epoxy propane, embedded in epoxy resin, and sectioned into ultrathin slices. The mitochondrial images were obtained using a transmission electron microscope (Titan G2 60-300; FEI, Hillsboro, OR) equipped with a corrected scanning corrector. The morphometric analysis of mitochondria using the Image-Pro Plus software (NIH) includes at least 300 mitochondria from at least 8 pictures per heart (Ding et al., 2020).
Cell culture and treatment
Primary neonatal cardiomyocytes were isolated from 1- to 2-day-old WT C57BL/6J mice or Nrf2−/− mice. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Sigma, St Louis, MO) supplemented with 10% fetal bovine serum (FBS) (Gibco, Grand Island, NY) and 1% streptomycin–penicillin (Beyotime) at 37°C in a 5% CO2 atmosphere. Cells were treated with 5 μmol DOX to establish the cell toxicity model (Lin et al., 2019) and co-incubated with 0.1 mmol IPA (Aladdin, Shanghai, China) (Du et al., 2021) or PBS in the culture medium. After 24 h, the culture medium was replaced, and the above treatment reagents were removed.
Immunofluorescence staining of Nrf2 in vivo and in vitro
After deparaffinization with xylene and subsequent rehydration through a graded ethanol series, the sections were blocked using a 10% bovine serum albumin (BSA) solution at 37°C for a duration of 30 min. Next, the sections were incubated with the Nrf2 antibody (#AF0639; Affinity, Jiangsu, China) at 4°C overnight, followed by a 1-h incubation with Alexa Fluor 594-conjugated secondary antibody (ProteinTech, Rosemont, IL). Finally, nuclear staining was achieved utilizing DAPI.
As for the primary cardiomyocytes cultured in vitro, cells were fixed with paraformaldehyde after which they were permeabilized with 0.1% Triton. Subsequently, the cells were blocked with 10% BSA at 37°C for 30 min. Following this, they were incubated with the Nrf2 antibody (#AF0639; Affinity) overnight at 4°C, followed by a 1-h incubation with Alexa Fluor 594-conjugated goat anti-rabbit secondary antibody (ProteinTech). Finally, nuclear staining was performed using DAPI. Images were acquired using a confocal microscope (Leica Stellaris, Wetzlar, Germany).
Cellular ROS and MitoROS detection methods
Cellular ROS levels were measured using an ROS assay kit (S0033S; Beyotime), and MitoROS levels were measured using MitoROS staining solution (AAT Bioquest, CA) according to the manufacturer's instructions. After removing the cell culture medium, an appropriate volume of diluted DCFH-DA and MitoROS staining was added to the cells. The cells were incubated at 37°C in a cell culture incubator for 20 min. After incubation, the cells were washed three times with serum-free cell culture medium to eliminate any residual fluorescent dyes that had not entered the cells. Images were acquired using a confocal microscope (Leica Stellaris).
Extraction of nuclear and cytoplasmic proteins
Nuclear and cytoplasmic proteins were extracted according to the manufacturer's instructions (P0028; Beyotime). Initially, cells were subjected to hypotonic conditions using Cell Cytoplasmic Protein Extraction Reagents A and B to induce cellular swelling, followed by disruption of the cell membrane to release cytoplasmic proteins. Nuclear pellets were obtained through centrifugation, with subsequent extraction of nuclear proteins using a high-salt nuclear protein extraction reagent.
Western blot
The total protein was extracted from heart tissue or primary neonatal cardiomyocytes cultured in DMEM (Sigma) with 10% FBS (Gibco) and 1% streptomycin–penicillin (Beyotime) at 37°C in a 5% CO2 atmosphere, using RIPA lysis buffer (Beyotime) containing Cocktail II (MedChemExpress) as a phosphatase inhibitor. Equal amounts of protein were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferred onto polyvinylidene fluoride membranes, blocked with 5% skim milk for 1 h, and incubated with specific primary antibodies overnight, followed by secondary antibody labeling.
Bands were visualized using enhanced chemiluminescence (Beyotime). Band quantification was performed using Image-Pro Plus (NIH). The specific antibodies used in this study were as follows: anti-BAX (ab32503; Abcam, Cambridge, England), anti-BCL2 (ab182858; Abcam), anti-cleaved-caspases 3 (ab214430; Abcam), anti-p-DRP1 (ser616) (#AF8470; Affinity), anti-DRP1 (ab184247; Abcam), anti-MFN1 (ab126575; Abcam), anti-MFN2 (ab124773; Abcam), anti-OPA1 (ab119685; Abcam), anti-ZO-1 (ab276131; Abcam), anti-Nrf2 (#AF0639; Affinity), anti-NQO1 (ab80588; Abcam), anti-HO1 (ab305290; Abcam), anti-Lamin B1 (ab16048; Abcam), and anti-β-actin (#4970; CST, Danvers, MA) (Supplementary Material).
Mitochondrial morphology imaging
Primary neonatal cardiomyocytes were labeled with 200 nM of MitoTracker to specifically label live mitochondria, and Hoechst dye was used to label cell nuclei. Images of the mitochondria were captured using a confocal laser scanning microscope (Leica Stellaris). For mitochondrial Z-axis volume reconstruction, the ImageJ 3D Object Counter plugin (NIH) was employed to analyze and quantify the number and size of mitochondrion (Ding et al., 2018b).
Statistical analysis
All data are expressed as mean ± standard error of the mean. Electronic laboratory notebook was not used. Statistical analysis was performed using SPSS 22.0 software (SPSS, Inc., Chicago, IL), and graphs were generated using Prism 8 software (GraphPad, San Diego). The Shapiro–Wilk test was employed to assess the normality of the variable distribution. t-Tests were used for statistical analysis between two groups. One-way or two-way ANOVA was used for comparisons among more than two groups, followed by Tukey's post hoc analysis. The correlations between cardiac function, metabolites, and bacterial taxonomic units were assessed using the Spearman correlation analysis in the R language (utilizing the heat map package). A p value <0.05 was considered statistically significant.
Footnotes
Acknowledgment
We deeply appreciate MD. Liping Meng for pointed advice and discussion for writing this article.
Ethics Approval and Consent to Participate
All animal protocols were approved by the Animal Care and Use Committee of Shaoxing People's Hospital.
Statement
Parts of the Figure 1 were drawn by using resources from Servier Medical Art (http://smart.servier.com). Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (![]() ).
).
Authors' Contributions
J.Z.: Experimental operation (lead) and writing—original draft (lead). J.H.: Visualization/data presentation (lead). Z.Z., J.Y., T.L., and B.Z.: Experimental operation (supporting). H.L.: Data analysis (lead). J.C.: Experimental design (lead). H.G.: Conceptualization (lead).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (No. 82174204 and No. 81873120) and the Science and Technology Plan Special Project of Shaoxing city (No. 2020B33002).
Supplementary Material
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
Supplementary Figure S13
Supplementary Figure S14
Supplementary Figure S15
Supplementary Figure S16
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.