
Abstract
Significance:
Reactive oxygen species (ROS), reactive nitrogen species (RNS), and reactive sulfur species (RSS) act as signaling molecules, regulating gene expression, enzyme activity, and physiological responses. However, excessive amounts of these molecular species can lead to deleterious effects, causing cellular damage and death. This dual nature of ROS, RNS, and RSS presents an intriguing conundrum that calls for a new paradigm.
Recent Advances:
Recent advancements in the study of photosynthesis have offered significant insights at the molecular level and with high temporal resolution into how the photosystem II oxygen-evolving complex manages to prevent harmful ROS production during the water-splitting process. These findings suggest that a dynamic spatiotemporal arrangement of redox reactions, coupled with strict regulation of proton transfer, is crucial for minimizing unnecessary ROS formation.
Critical Issues:
To better understand the multifaceted nature of these reactive molecular species in biology, it is worth considering a more holistic view that combines ecological and evolutionary perspectives on ROS, RNS, and RSS. By integrating spatiotemporal perspectives into global, cellular, and biochemical events, we discuss local pH or proton availability as a critical determinant associated with the generation and action of ROS, RNS, and RSS in biological systems.
Future Directions:
The concept of localized proton availability will not only help explain the multifaceted nature of these ubiquitous simple molecules in diverse systems but also provide a basis for new therapeutic strategies to manage and manipulate these reactive species in neural disorders, pathogenic diseases, and antiaging efforts.
Introduction
Reactive molecular species derived from molecular oxygen (O2), nitric oxide (NO), and hydrogen sulfide (H2S)—often referred to as reactive oxygen species (ROS) (Mittler, 2017; Sies and Jones, 2020), reactive nitrogen species (RNS) (Fang, 2004; Patel et al., 1999), and reactive sulfur species (RSS) (Giles et al., 2017; Gruhlke and Slusarenko, 2012)—play pivotal roles in cellular function and homeostasis of all living organisms. Numerous studies have demonstrated that these reactive molecular species function as crucial signaling molecules, regulating gene expression (Cerny et al., 2018; Griendling et al., 2000; Perillo et al., 2020), enzyme activity (D'Autreaux and Toledano, 2007; Sies and Jones, 2020), and physiological responses (Mittler et al., 2022; Sies et al., 2022). Simultaneously, it has been also shown that an excess of these species is harmful, causing cellular damage and death.
The dual nature of ROS, RNS, and RSS, which has been likened to a “double-edged sword” (Kalogeris et al., 2014; Martin and Barrett, 2002), “two sides of the same coin” (Lechner et al., 2005), or “Yin-Yang” (Lundberg et al., 2008; Sakihama et al., 2002a), has presented an intriguing conundrum in life science and potentially causes confusion among nonspecialists.
The chemical interplays among ROS, RNS, and RSS result in the generation of a variety of reaction products, such as peroxynitrite (ONOO−), which is produced by the reaction between NO and the ROS superoxide (O2 •−) (Yamasaki, 2000). Due to the close relationship between ROS and RNS in oxidative stress, they have sometimes been collectively referred to as reactive oxygen and nitrogen species (RONS) (Weidinger and Kozlov, 2015). Moreover, hybrid species of RNS and RSS, such as nitrosopersulfide (SSNO−) and dinitrososulfite (N-nitrosohydroxylamine-N-sulfonate [SULFI/NO]), have been reported as key molecules in the NO/H2S cross talk (Cortese-Krott et al., 2016; Cortese-Krott et al., 2015). The presence of complex cross talk among ROS, RNS, and RSS further compounds the difficulty in understanding the physiological actions of these molecules, especially in vivo (Cortese-Krott et al., 2017; Cortese-Krott et al., 2015).
Assuming that chemical reactions provide the foundation upon which biomolecules interact, we can consider that redox chemistry, associated with reactive species, constitutes a universal language that enables communication between different entities and layers of regulation (Santolini et al., 2019). Santolini et al. (2019) applied the concept of “Lingua Franca” to explain the multifaceted nature of redox reactions. “Lingua Franca” refers to a language adopted as a common means of communication among speakers whose native languages are different (Seidlhofer, 2004). This analogy underscores the complexity that may arise from the history of human language and redox processes associated with reactive species. Considering reactive species as a universal language among living organisms, each species could be seen as a “word” in intra- or intercellular communication. Similar to words in human language, the meanings of these species can shift based on their historical background, often carrying contradictory meanings in different contexts.
Contronyms, also known as auto-antonyms or Janus words, are words with multiple meanings, some of which are contradictory or opposite of each other (Murodova and Djumabayeva, 2017). For example, the word “cleave” can mean both “to split or separate” and “to adhere or cling together” (Jackson, 2018). The biological roles of ROS, RNS, and RSS may be viewed as such “contronymic” molecular species. By integrating ecological and evolutionary perspectives, we aim to create a framework that bridges seemingly independent findings observed in various biological systems. Our goal is to discuss the “context” factors in biology that determine the roles and meanings of contronymic molecular species, specifically ROS, RNS, and RSS, in different biological contexts.
Echoing Peter Mitchell's chemiosmotic theory from the 1960s, which has enhanced our understanding of the role of the ubiquitous proton (Rich, 2008), we propose that understanding the spatiotemporal differentiation of reactions, along with the regulation of local proton availability, is crucial for grasping the diverse actions of ROS, RNS, and RSS in cells. Adopting a holistic approach, which includes considering proton availability as discussed in this article, will not only help explain the multifaceted nature of these molecules in various systems but also pave the way for novel therapeutic strategies in treating cancer, neural disorders, pathogenic diseases, and in antiaging research.
Hans Selye's Stress Concept
Oxidative stress often leads the public and nonspecialists to perceive “stress” as inherently damaging. However, according to Selye's (1955) initial proposal, “stress” primarily refers to the body's response to demands or challenges (Iwama, 1998). He emphasized that stress is not solely a negative experience, but it can encompass any demand that requires the body to adapt or adjust (Selye, 1955).
Incorporating the concept of oxidative stress into stress theory should inherently encompass both positive and negative aspects (Forman and Zhang, 2021). Selye differentiated between pleasant stress, which he labeled “eustress,” and unpleasant stress, referred to as “distress” (Selye, 1955). Similarly, oxidative stress is sometimes categorized into “oxidative eustress” and “oxidative distress” to clarify their distinct physiological implications (Sies and Jones, 2020). Therefore, it is logical to designate ROS, RNS, and RSS as “stressors” that elicit a broad range of physiological responses, exerting both positive and negative effects on all living organisms.
Figure 1 illustrates the diverse origins of O2, NO, and H2S, which are primary sources of ROS, RNS, and RSS. These sources can be categorized as either extrinsic or intrinsic, both of which significantly influence the physiology and metabolism of all living organisms. While medical sciences often focus on intrinsic sources, which are regulated by multiple enzymes or proteins, it is crucial to recognize that plants, microbes, and even symbiotic bacteria colonizing the human gut—which metabolize environmental inorganic compounds—are also significantly influenced by extrinsic sources of O2, NO, and H2S. The levels of these extrinsic sources can fluctuate greatly in natural environments.
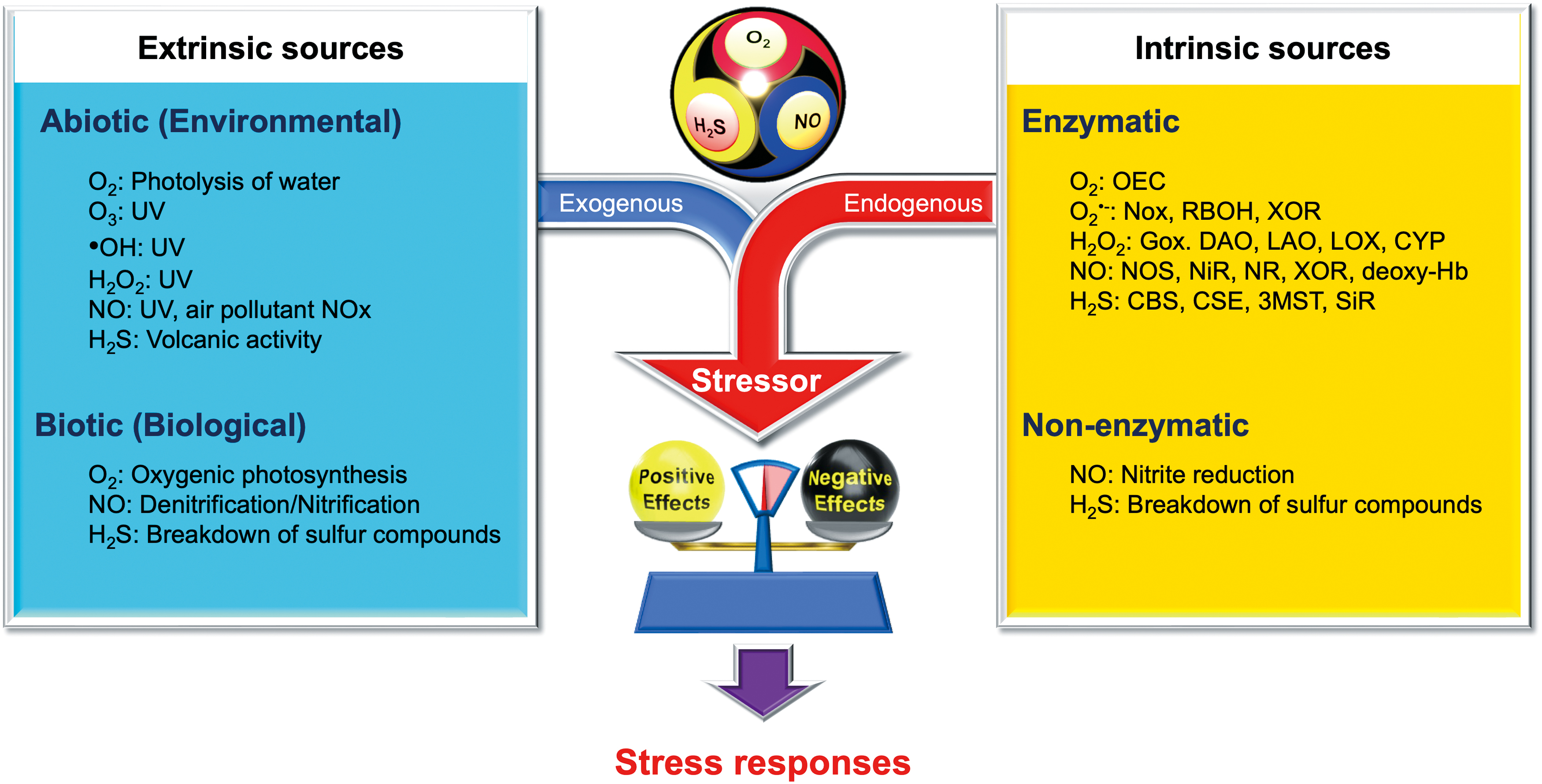
Environmental Perspectives
In nature, biogenic O2, NO, and H2S, and their derivatives, circulate within the biosphere, contributing to the global geochemical cycle (Fowler et al., 2013; Lasaga and Ohmoto, 2002; Maier, 2015). Fundamental roles in these biogeochemical cycles are played by plants, soil bacteria, and oceanic phytoplankton (Maier, 2015). To explore the biological implications of these “contronymic” molecules in ecological and evolutionary contexts, it is necessary to consider the environmental ROS, RNS and RSS as extrinsic sources. Figure 2 illustrates the biogeochemical cycles of nitrogen and sulfur, commonly referred to as the nitrogen cycle and the sulfur cycle, respectively (Madsen, 2011).
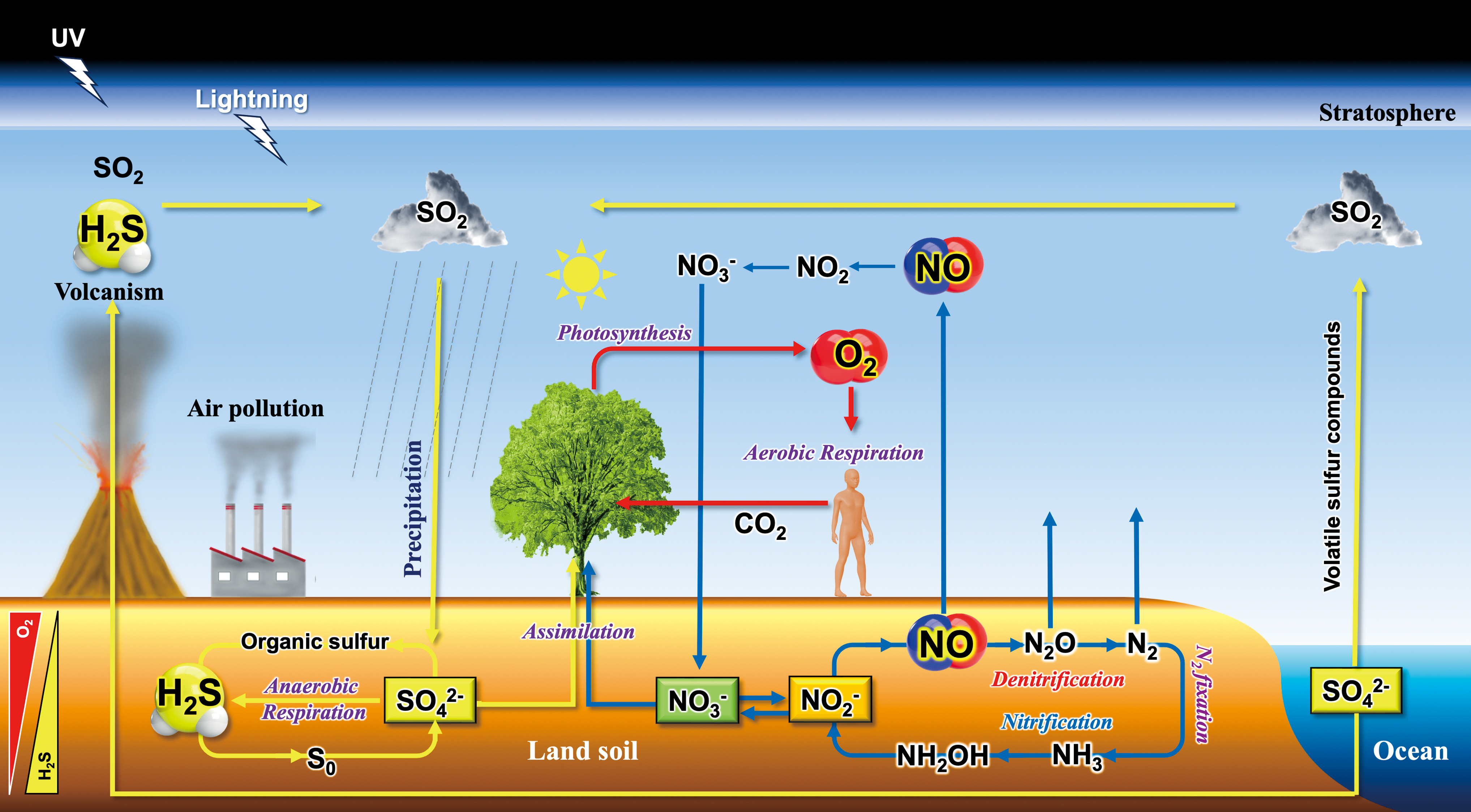
In addition to biogenic sources, the abiotic photochemical generation of reactive species is also significant, particularly in the context of biological evolution (Blake and Carver, 1977). Ozone (O3), a form of ROS, is primarily produced in the stratosphere through a process involving the interaction of O2 with ultraviolet-C (UV-C) radiation from the sun. This process, known as the ozone–oxygen cycle, is essential for protecting life on Earth from the harmful effects of UV-C radiation (Staehelin et al., 2001).
Ionization of atmospheric molecules results in the creation of plasma (da Silva et al., 2019). Lightning is a visible example of this process, with a plasma channel manifesting as a lightning bolt. This process facilitates the formation of reactive species, including ROS and RNS such as •OH, NO, and NO2 (Brune et al., 2021). Once formed in the atmosphere, these reactive species can be deposited onto the soil surface via rainwater or other forms of precipitation.
The hydroxyl radical (•OH) can be also generated by the reaction of O3 with water vapor (H2O) in the presence of UV-C radiation. Often referred to as the “detergent” of the atmosphere, •OH plays a crucial role in natural atmospheric chemistry (Li et al., 2008). It participates in the oxidation and removal of many atmospheric pollutants such as SO2, CO, NO, and NO2, greenhouse gases such as methane, and volatile compounds emitted from plants such as isoprene (Tambunan et al., 2006), thereby playing a significant role in maintaining the self-cleaning capacity of the atmosphere (Rohrer et al., 2014). This self-regulating mechanism bears similarities to those of living organisms, a concept that aligns with the Gaia hypothesis (Lovelock and Margulis, 1974). This hypothesis proposes that Earth's self-regulated physicochemical properties are maintained within a range favorable for life, forming a superorganism (Maier, 2015).
Biological evolution is closely linked with changes in Earth's atmospheric composition throughout history (Catling and Zahnle, 2020; Yamasaki and Cohen, 2016). In early cells, protein Fe-S clusters would function to transfer electrons from H2 to carbon dioxide (CO2), producing organic acids (Muchowska et al., 2020; Roldan et al., 2015). Abundant lightning activity in an anaerobic atmosphere rich in NH3 and CO2 would generate nitrogen oxides (NOx), including NO and nitrite, which could be readily reduced to NO. The binding of NO to Fe-S clusters would initially have inhibited autotrophic electron transfer. The consequent requirement for NO detoxification, along with positive selection pressure for the use of higher potential electron acceptors to maximize proton translocation across the membrane, would lead to gene duplication events. These would result in proteins designed to reduce NO and other NOx, ultimately leading to a complete denitrification pathway (Catling and Zahnle, 2020).
As volcanic activity diminished, natural selection favored the evolution of pathways for reductive assimilatory electron transfer to NOx compounds to generate ammonia for cellular biomass (Catling and Zahnle, 2020). Eventually, depletion of NOx, due to increased cellular utilization and concurrent decreases in production from lightning (Navarro-González et al., 2001), led to the evolution of N2 fixation and the eventual achievement of a balance between biotic production of N2, via denitrification and anammox, and its utilization via N2 fixation (Catling and Zahnle, 2020).
In observing the current biosphere, one might note that there is always a gradient from oxic (O2) environments to suboxic (NO) and then to anoxic (H2S) environments, progressing from the surface to deeper layers or interiors. This can be observed at scales ranging from microbial (e.g., biofilms) to global (e.g., soil or ocean) (Yamasaki and Cohen, 2016). It is important to recognize that the abiotic or environmental reactive oxygen, nitrogen, and sulfur species—collectively abbreviated henceforth as RONSS (Deng et al., 2017)—potentially influence all living organisms, in addition to those synthesized endogenously within organisms themselves. Assuming that RONSS function as stressors, one could speculate that organisms may detect environmental changes through fluctuations in ambient RONSS levels, subsequently regulating their metabolism and development.
Minimizing Environmental Impacts
The chemical behavior of NO should be differentiated between the aqueous phase and the gas phase. In an aqueous solution, NO rapidly reacts with O2 •− to form ONOO−, a highly harmful RNS closely linked to human diseases (Szabo et al., 2007). O2 •− spontaneously decays in acidic solutions to form another ROS, hydrogen peroxide (H2O2). This spontaneous decay of O2 •− is strongly dependent on pH, showing the highest second-order rate constants near pH 5 (Bielski and Allen, 1977). Under physiological conditions, the decay of O2 •− at acidic pH can be more than 100 times faster than at neutral pH (Bielski and Allen, 1977). As discussed later, this spontaneous decay of O2 •− does not occur in the absence of H+ or H2O, such as in aprotic environments. For example, less than 10% decomposition occurs in 1 day at room temperature in dimethyl sulfoxide (Bielski and Allen, 1977).
The presence of the enzyme superoxide dismutase (SOD) significantly accelerates the decay of O2
•− making it faster than its spontaneous decay (Fridovich, 1975): k = 4.5 × 105 M
−1s−1 and k = 0.62 × 109 M
−1s−1 for spontaneous reaction and Cu/Zn-SOD reaction, respectively (Han et al., 2003).
In the presence of SOD or the absence of O2 •−, NO is relatively stable in aqueous solutions, exhibiting half-lives of 500s or longer (Ignarro et al., 1993).
Similarly, the chemical fate of H2S differs between the aqueous phase and the gas phase. Once dissolved into an aqueous solution, H2S reaches equilibrium with its ionic form, the sulfide ion (HS−):
Above pH 7, HS− is the dominant species in sulfide solutions (pK a = 7.0) (Cortese-Krott et al., 2015) while the protonation of HS− predominantly results in H2S, which is not readily oxidized by O2 in the aqueous phase (Chen and Morris, 1972). Although physiological relevance is questioned, a trace amount of HS− could be further deprotonated to form S2− (pK a >14) (Nagy et al., 2014). In the absence of target molecules in waters, HS− may be slowly oxidized to elemental sulfur. The elemental sulfur can then combine with the remaining HS− to form polysulfides (SX 2−) (Chen and Morris, 1972).
Membrane permeability of RONSS
Noncharged small molecules, including gaseous O2, NO, and H2S, present in the aqueous phase, can diffuse across lipid bilayer membranes (Fig. 3). It is worth remembering that some of the RONSS derived from these gases are neutral radicals (e.g., •OH and •NO), some are ions (e.g., O2 •− and ONOO−), and others are nonradicals (e.g., H2S and H2O2) (Pryor, 1986). In principle, the ionic forms of RONSS, such as O2 •− (Mao and Poznansky, 1992; Takahashi and Asada, 1983), or other charged molecules cannot diffuse across the membranes without the aid of ion transporters (Fig. 3). Interestingly, H2O2 is naturally membrane-permeable, although facilitated diffusion via aquaporins, also known as peroxiporins, has recently been suggested (Cerny et al., 2018; Rodrigues et al., 2019; Sies, 2017; Tong et al., 2019).
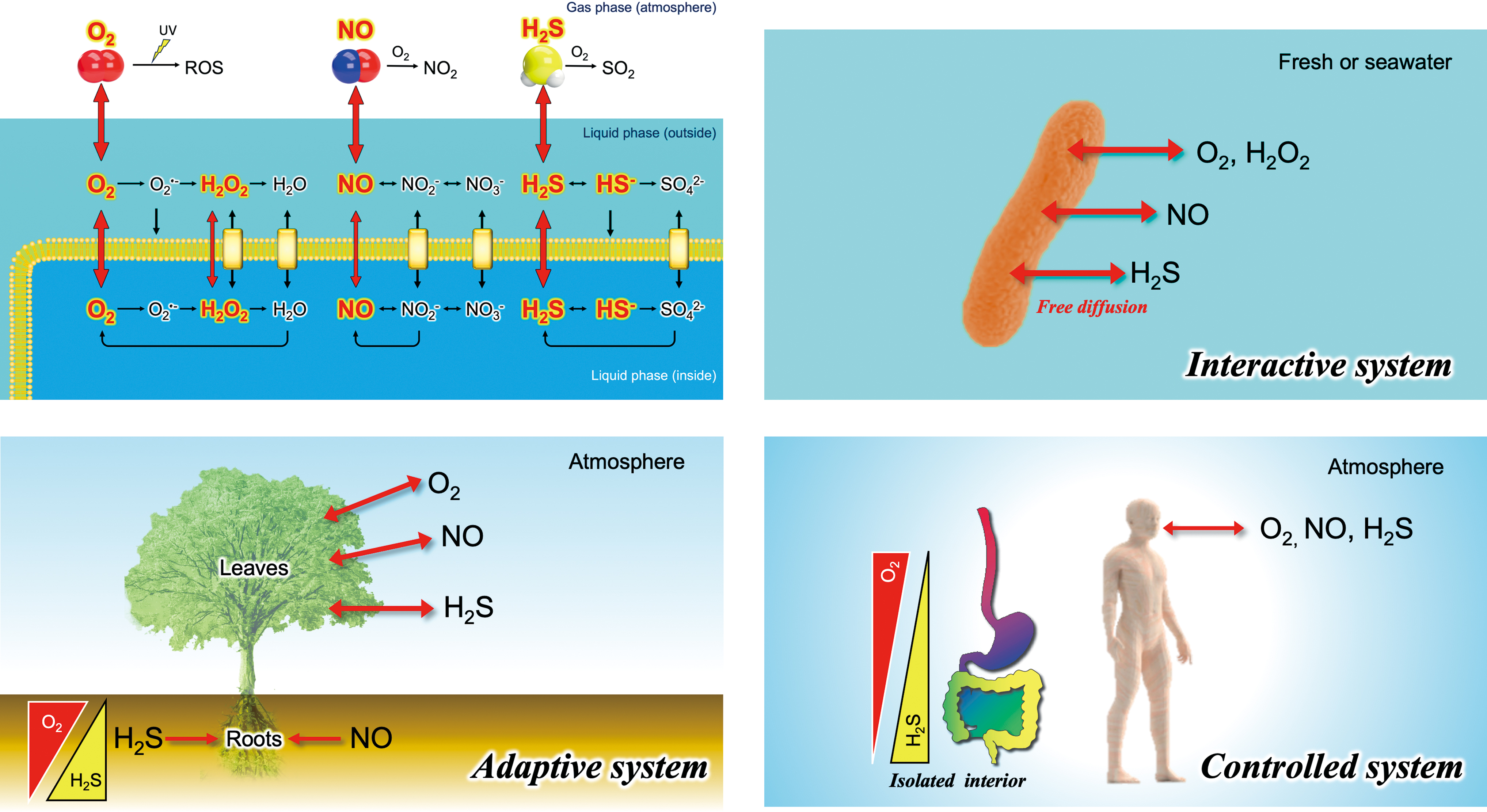
The ability of H2O2, NO, and H2S to pass freely across biomembranes positions them as promising candidates for universal signaling molecules in intra- or intercellular communication. In view of environmental and ecological perspectives, we propose the categorization of three biological systems, each adopting different strategies for coping with and productively utilizing RONSS (Fig. 3).
Interactive systems
Free-living bacteria, archaea, and microalgae residing in aqueous environments are directly influenced by environmental RONSS (Imlay, 2019). In laboratory experiments involving such aquatic organisms, oxidative damage can be observed when the concentration of RONSS, such as H2O2, becomes excessive (Imlay, 2019). However, in natural environments, unlike controlled laboratory experiments with a limited volume of medium, membrane-permeable RONSS—including H2O2, NO, and H2S—are likely to diffuse into the surrounding water due to dialysis effects. This diffusion does not significantly increase the concentration in the surrounding water due to its vast volume in natural environments. Therefore, an accumulation of membrane-permeable RONSS may not occur in an interactive system under natural conditions, assuming the volume of the surrounding water is virtually unlimited.
In contrast, microorganisms that colonize the bodies of animals and plants, establishing symbiotic, infectious, or parasitic relationships, may face circumstances distinct from those encountered by free-living organisms. These organisms could face challenges due to the dynamic fluctuations in RONSS levels induced by the host immune systems (Andres et al., 2022).
Controlled systems
Heme-containing proteins, such as hemoglobin and myoglobin, which are capable of binding O2, NO, and H2S, are abundant in the muscles and blood of vertebrate animals (Lancaster, 1994). Consequently, the impact of environmental RONSS (Fig. 1) can be minimized within these animals' bodies, including humans. From the RONSS perspective, vertebrate animals, equipped with heme protein armor, can be regarded as a controlled system offering control mechanisms that organisms have in place to manage their exposure and response to environmental RONSS (Fig. 3).
Insects absorb O2 through a network of tubes known as tracheae (Westneat et al., 2003), which are directly connected to the atmosphere via openings, valve-like spiracles, located on the abdomen and thorax's surface (Hetz and Bradley, 2005). They have sophisticated processes for adjusting the hydrostatic pressure in the haemocoel, actively ventilating the tracheal system (Hetz and Bradley, 2005). Most arthropods are believed to reduce gas exchange through the trachea during periods of low metabolic activity to avoid O2 toxicity (Hetz and Bradley, 2005). While a small amount of O2, NO, and H2S might be able to diffuse across the insect exoskeleton, the primary route by which gases enter and exit the insect's body is through the tracheal system. Therefore, arthropods can also be considered a controlled system (Mahdi et al., 2010; Trimmer et al., 2001).
The development of a controlled system was likely necessary for evolutionary adaptation to terrestrial environments to prevent water loss from the body. Indeed, aquatic invertebrate animals, such as jellyfish (Moroz et al., 2004), reef-building corals (Bouchard and Yamasaki, 2008), and cuttlefish (Palumbo, 2005), are susceptible to water eutrophication or water pollution (Yuen et al., 2009), where environmental levels of NO or H2S are elevated due to bacterial activity (Meyer-Reil and Köster, 2000). They appear to retain an interactive system that is sensitive to environmental RONSS fluctuation.
Adaptive systems
Terrestrial plant leaves control gas exchange and water loss through stomata, small pores surrounded by a pair of guard cells on their surfaces (Sakihama et al., 2003). Meanwhile, roots absorb water as well as other inorganic nutrient ions dissolved in the soil water (Alam, 1999). Terrestrial plants, balancing interactive and controlled system traits, are thus categorized as adaptive systems. Their ability to neutralize or isolate RONSS may be less than that of animals, except in nodules rich in hemoglobin found in legume roots (Hichri et al., 2015). Notably, terrestrial plants produce NO and H2S as by-products of nitrate and sulfate assimilation under stress conditions (Yamasaki, 2000; Yamasaki and Cohen, 2016). An adaptive system facilitates the diffusion of these gases into the atmosphere to prevent internal concentration buildup. Terrestrial plants likely sense abiotic and biotic stresses through RONSS fluctuations, influencing their growth and development (Yamasaki, 2005).
A controlled system provides a significant advantage in mitigating the influence of extrinsic or environmental RONSS fluctuations (Fig. 1), maintaining the lowest possible RONSS level under normal conditions compared with interactive or adaptive systems (Fig. 3). Although further clarification of this classification is needed, the concept aids in understanding the biological implications of RONSS across diverse organisms and environments (Ignarro et al., 1993). This may explain the loss of NO synthase (NOS) in terrestrial plant lineages, despite the prevalence of NO-based signaling systems (Jeandroz et al., 2016). An NOS-based signaling system might require low background levels of NO, provided by a controlled system.
Lessons from the Oxygen-Evolving Enzyme
Oxygen-evolving complex of photosystem II
In the context of biological evolution, it is widely accepted that the Great Oxygenation Event (GOE) 2.4 billion years ago was a pivotal step in the evolution of life on Earth (Hamilton, 2019; Sanchez-Baracaldo and Cardona, 2020). This event paved the way for the development of aerobic organisms that require O2 for respiration. A significant biological innovation during this period was the oxygen-evolving or oxygenic photosynthesis of ancestral cyanobacteria, which began using H2O, rather than H2S, as the electron donor for the light-driven electron transport system (Hamilton, 2019). This crucial change has played a significant role in shaping the current O2-rich atmosphere on Earth (Hamilton, 2019).
Water oxidation, or water-splitting, proceeds by coupling the photochemistry at the reaction center chlorophylls with the oxidation of water, which involves the four-electron and four-proton reaction at the oxygen-evolving complex (OEC) located deep within photosystem II (PSII). It is believed that PSII evolved from type II reaction centers of anoxygenic phototrophs (Cardona, 2019).
Oxygen evolution in photosynthesis follows the Kok cycle, also known as the S-state cycle (Kern et al., 2018), which outlines the sequential steps occurring within the OEC. These steps are essential for the splitting of water molecules into oxygen, protons, and electrons, as illustrated in Figure 4. This process includes five states, ranging from S0 to S4, and is cyclical in nature, as also depicted in Figure 4. In this context, the “S” in “S-state” specifically refers to “state,” indicating the different oxidation states of the manganese cluster within the OEC of PSII. (Note that “S” does not represent “sulfur”; therefore, “S4” in this context does not refer to polysulfide.)

The transition from S3 to S4 and back to S0 is a crucial step in the Kok cycle. It involves the formation of a reactive S4 intermediate, the release of O2 and two protons, and the binding of a water molecule. This final step is regulated by proteins surrounding the OEC, which play a functionally critical role. The Mn4CaO5 cluster within the OEC (Fig. 4) is widely recognized as harboring the molecular mechanism underpinning the Kok cycle (Kern et al., 2018).
O2 evolving mechanism
Bhowmick and coworkers (2023) revealed rapid structural changes at the active site of the OEC during the S3 to S4 to S0 transition. They used an advanced technique known as serial femtosecond crystallography with X-ray free electron lasers. This technique allows for the elucidation of time-dependent structural changes within the Mn4CaO5 cluster of the OEC and its protein environment (Bhowmick et al., 2023).
After light exposure in the S3 state, the oxidation of the redox-active D1-Y161 tyrosine (referred to as YZ) in the OEC occurs in less than 50 μs. YZ mediates electron transfer from the OEC to a special pair of chlorophyll molecules. The formation of the tyrosine radical initiates an early deprotonation event at 200–500 μs, involving a manganese-bound water ligand and two amino-acid residues (Asp61 and Glu65). The YZ radical then extracts an electron from the manganese cluster, transitioning the system to the transient S4 state. This process begins ∼500 μs after the initial light exposure (Kern et al., 2018).
In their 2023 study, Greife et al. (2023) used an approach known as time-resolved microsecond Fourier-transform infrared spectroscopy to investigate the kinetics of the S3→S4→S0 transition. They demonstrated that the formation of the YZ radical after the S3 state is succeeded by a deprotonation event at 340 μs (Greife et al., 2023). Their quantum-chemical calculations attribute this to either Asp61 or a pair of residues (Glu65 and Glu312), findings that align remarkably well with the conclusions drawn by Bhowmick and coworkers (2023) and Pantazis (2023).
Proton Transfer as a Key Factor
Recent breakthroughs in the study of photosynthesis have yielded significant insights at the molecular level, with high time resolution, into how the OEC manages the water-splitting process without harmful ROS production. The formation of a reactive manganese-bound oxygen radical (Mn–O•, the S4 transient reactive intermediate) involves a simultaneous transfer of one electron and three protons, identified as the rate-limiting step of the entire reaction (Bhowmick et al., 2023). The subsequent O–O bond formation and O2 release are relatively faster (Bhowmick et al., 2023). The O–O bond formation can proceed only after proton removal is complete (Greife et al., 2023), implying a molecular mechanism for avoiding the generation of highly reactive intermediate products (Pantazis, 2023).
A key factor appears to be the dynamic structural changes in the distance between redox components (Bhowmick et al., 2023), and regulation of proton transfer, presumably through hydrogen bond networks, as discussed later, between the outer aqueous phase and the active site of the OEC (Hussein et al., 2023). These recent findings suggest that a precise and dynamic spatiotemporal arrangement on an Å scale is crucial for the safe operation of the water oxidation in PSII. In other words, the mechanism for controlling the transfer of protons to a bulk aqueous phase, along with the strict regulation of the redox events, is important for mitigating potentially deleterious ROS formation.
As mentioned in the case of the spontaneous disproportionation reaction of O2 •−, many reactions associated with RONSS are highly pH-dependent (Andres et al., 2023; Nosaka and Nosaka, 2017). This suggests that pH (or proton availability) is a critical factor that alters the protonation status of target molecules, as well as ionic forms of RONSS themselves. It plays a significant role in regulating RONSS-associated reactions, as demonstrated in the water-splitting process (Bhowmick et al., 2023; Greife et al., 2023). Therefore, controlling the local pH or proton availability at a reaction site can be considered essential for managing the seemingly nonspecific RONSS reactions in living systems.
Mitchell's Chemiosmotic Theory
Proton gradient
When discussing the biochemical function of proton in biological systems, it is worthwhile to revisit the chemiosmotic theory (Mitchell, 1961), which established a primary role for ubiquitous protons (H+) in cellular energetics. This theory, proposed by the Nobel laureate Peter Mitchell in the 1960s, is a fundamental concept in the field of bioenergetics (Rich, 2008). It explains how ATP is synthesized during the processes of aerobic respiration (oxidative phosphorylation) and photosynthesis (photophosphorylation). Mitchell suggested that these processes involve the formation of an H+ gradient across a membrane. This gradient, known as the proton motive force (pmf), generates a difference in the electrochemical potential of H+ (ΔμH+) between two aqueous phases that drives ATP synthesis (Barber, 1982).
where Δp represents the total pmf, ΔμH+ represents electrochemical difference of H+ between two aqueous phases, Δψ represents the electrical potential difference across the membrane, ΔpH represents the difference in pH across the membrane, R is the gas constant, T is the absolute temperature, and F is Faraday's constant. Thus, the overall pmf (Δp) can be given as the sum of the two components: Δψ (membrane potential or electrical component) and ΔpH (pH difference or H+ concentration component) across the membrane. It should be noted that ionic RONSS molecules, such as O2 •−, contribute to the membrane potential (Δψ).
Chemical perspective of ΔpH formation
In terms of energetics, the Δψ and ΔpH components of pmf are mutually convertible, which can be expressed as in millivolt (Nicholls, 2005). However, from a chemical perspective, a fundamental difference exists between them. The ΔpH formation results in pH changes or pH shifts on either side (or both sides) of the aqueous phase, which strongly impacts the pH-dependent chemical reactions as well as the protonation status of molecules exposed to the bulk aqueous phase. Interestingly, the contribution of ΔpH to pmf varies significantly among mitochondria, bacteria, and chloroplasts. Under normal conditions in mitochondria and bacteria, the primary component of the pmf is Δψ (about 99%), and the contribution of ΔpH is small, typically less than 0.5 pH units (Dzbek and Korzeniewski, 2008). In contrast, ΔpH is a significant component of the pmf in chloroplasts and cyanobacteria (Takahashi and Yamasaki, 2002), which carry out oxygenic photosynthesis.
In chloroplasts, the ΔpH under illumination exceeds 3 pH units (Schuldiner et al., 1972). It should be noted that the relationship between pH and [H+] is logarithmic; therefore, a 3-unit change in pH represents a 1000-fold difference in proton concentration. In the darkness, the pH of both the stroma and lumen (outside and inside of thylakoid membranes, respectively) is ∼7. Under illumination that excites PSII and PSI to drive the photosynthetic electron transport system, the pH of the stroma may reach to 8, while the lumen pH decreases to 5. Synchronizing with this pH shift, in addition to the redox regulation of thiol enzymes by thioredoxins (Hisabori et al., 2013; Mills and Mitchell, 1982), many enzymatic activities are optimized to drive metabolisms, including CO2 fixation, known as the Calvin–Benson cycle (Buchanan, 1980).
The alkalinization in the stroma (along with the acidification of the lumen space) may be a simple mechanism for controlling a variety of redox metabolisms, including RONSS reactions (Fig. 4). This is extremely important in photosynthesis because the fluctuation of light energy in natural environments is beyond the speed of gene expression and protein translation. In contrast, in mitochondria and bacteria under normal conditions, pH does not significantly change either inside or outside the membranes during electron transport, which could contribute to pH homeostasis during ATP synthesis. If matrix pH increases, it stimulates ROS generation in mitochondria (Selivanov et al., 2008).
Therapeutic perspectives of uncouplers
Mitchell's chemiosmotic theory has contributed to explaining the cytotoxicity of 2,4-dinitrophenol (DNP). Historically, DNP and the related compounds were used as food coloring agents known as “Martinus Yellow” or “Victoria Yellow” (Parascandola, 1974). Although these compounds were also tested in clinical trials for weight loss in patients, they were banned for clinical use in 1938 due to safety concerns (Childress et al., 2018). Excessive intake of DNP can cause severe hyperthermia (elevated body temperature), which may result in organ failure and death (Grundlingh et al., 2011). The mechanism of this toxicity was not previously understood. However, Mitchell explained this effect by proposing that DNP increases the H+ permeability of membranes, thereby dissipating the Δμ H +, generated by the respiratory electron transport chain, as heat (Skulachev, 1998). This phenomenon, known as “uncoupling” or “proton leak” in oxidative phosphorylation, disrupts the coupling mechanism between ATP synthesis and electron transport activity.
According to Mitchell's chemiosmotic theory, proton carriers, often referred to as “uncouplers,” inhibit ATP synthesis by dissipating the pmf across energy-transducing membranes including the inner membrane of mitochondria, the thylakoid membrane of chloroplasts, and the cell membrane of bacteria.
Given that mitochondria are the major source of ROS in eukaryotic cells, the process of uncoupling has been brought into focus once again due to its close association with many diseases (Zhao et al., 2019). Chemical uncouplers, as well as endogenous uncoupling proteins (UCPs) (Cadenas, 2018), have garnered significant attention from medical researchers as potential therapeutics for treating obesity, diabetes, neurological disorders, cardiovascular diseases, aging, and metabolic diseases (Childress et al., 2018). The cardioprotective effects of red wine have been attributed to the uncoupling action of dietary polyphenols, such as galangin (Dorta et al., 2008; Modriansky and Gabrielova, 2009). Moreover, the pharmaceutical effects of chemical uncouplers, such as DNP, carbonyl cyanide m-chlorophenylhydrazone (CCCP), and nemorosone, have been evaluated in various models of cancer, both in vitro and in vivo (Pardo-Andreu et al., 2011; Shrestha et al., 2021). Some of these compounds are currently undergoing clinical trials (Shrestha et al., 2021).
Effects of Uncouplers on Ferroptosis
Ferroptosis induced by uncouplers
Recently, uncouplers have been found to induce ferroptosis that could lead to potential therapeutic approaches in treating cancer (Fernandez-Acosta et al., 2023). Ferroptosis, a nonapoptotic cell death, has attracted great attention due to its crucial roles in organ injury, degenerative disease, and vulnerability of therapy-resistant cancers (Mishima et al., 2022; Viswanathan et al., 2017). The term ferroptosis was coined in 2012 (Dixon et al., 2012) to describe a type of regulated cell death characterized by iron-dependent lipid peroxidation, which can be distinct from apoptosis, necrosis, and autophagy (Tang et al., 2021). Ferroptosis has been demonstrated to be induced by drugs, ionizing radiation, and cytokines, which suppress tumor growth (Chen et al., 2021).
It has been revealed that ferroptosis is prevented by glutathione peroxidase-4 (GPX4) (Bersuker et al., 2019), ferroptosis suppressor protein-1, which was previously known as apoptosis inducing factor mitochondrial 2 (Bersuker et al., 2019), coenzyme Q10 ubiquinone (Bersuker et al., 2019), or vitamin K (Mishima et al., 2022).
Lipid ROS accumulation is a hallmark of ferroptosis, but where and how it is generated during ferroptosis has not been defined. In particular, the role of mitochondria in ferroptosis remains highly controversial. Gao and coworkers (2019) provided evidence to show that mitochondria indeed play a crucial role in cysteine-deprivation-induced ferroptosis but not in that induced by inhibiting GPX4. Using BODIPY 581/591 C11, a fluorescent probe for membrane-localized ROS, they demonstrated that the uncoupler CCCP (10 μM) completely blocked the lipid ROS accumulation and ferroptosis (Gao et al., 2019). Based on the experimental evaluation, they proposed that ferroptosis is a natural tumor suppressive mechanism under diverse biological conditions (Gao et al., 2019).
Fernandez-Acosta and coworkers (2023) further explored the association between uncoupling and ferroptosis. They reported that nemorosone, a naturally occurring polycyclic polyprenylated acylphloroglucinol isolated from Clusia plants, which exhibits anticancer activity, induces ferroptosis (Fernandez-Acosta et al., 2023). Along with its anticancer activity, nemorosone is known to exhibit potent uncoupling activity comparable with the classical uncoupler, CCCP (Pardo-Andreu et al., 2011). It was demonstrated that both CCCP-induced cell death and lipid peroxidation could be inhibited by the ferroptosis inhibitor ferrostatin-1 and the iron chelator deferoxamine, as well as by the respiratory electron transport chain inhibitors rotenone and antimycin A.
The team suggested that the effects of CCCP demonstrated dual roles: the induction of ferroptosis at high concentrations (50 μM) presumably through the increase of the intracellular labile Fe2+ pool via heme oxygenase-1 upregulation, and protection from ferroptosis at lower concentrations (Fernandez-Acosta et al., 2023). It appears that a “mild uncoupling (Fernandez-Acosta et al., 2023)” induced by CCCP or nemorosone may exhibit a distinct effect on lipid peroxidation in mitochondria.
Effects of uncouplers on lipid peroxidation
Under normal conditions, the reduction of O2 to O2 •− by redox components in mitochondria is the primary cause of the lipid peroxidation during respiratory electron transport. This is followed by the subsequent production of H2O2, either through enzymatic SOD reactions or spontaneous disproportionation reactions. The reaction between O2 •− and H2O2, capable of producing •OH, is part of a complex set of reactions known as the Haber–Weiss reaction [Eq. (7)], which operates in conjunction with the Fenton reaction [Eq. (9)]. Since O2 is not an efficient reactant, heme-iron proteins may catalyze lipid peroxidation in vivo (Cheng and Li, 2007). Equation (8) illustrates a reaction from the iron-catalyzed Haber–Weiss cycle, often associated with the Fenton reaction in biological systems. The •OH generated initiates chain reactions that produce lipid peroxides (LOOH).
Lipophilic antioxidants, such as α-tocopherol (vitamin E) (Kajarabille and Latunde-Dada, 2019), can prevent lipid peroxidation and terminate the chain reaction. NO is also known to act as a chain-breaking antioxidant in lipid peroxidation (Wink et al., 2001). Thus, the O2
•−, H2O2, and •OH generated by the respiratory chain are primarily involved in the process of lipid peroxidation.
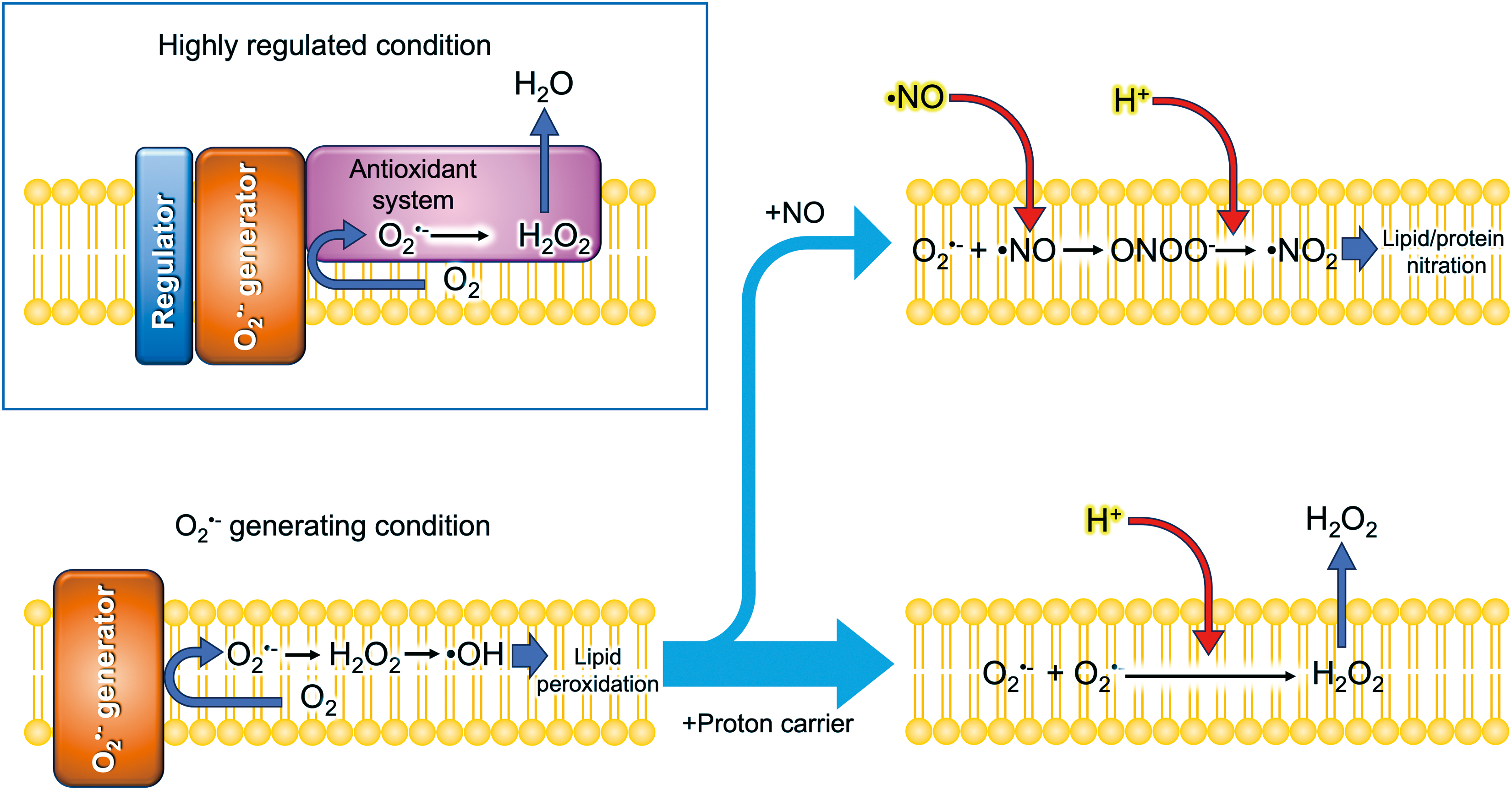
where LH is a lipid with allylic hydrogens, which are present in polyunsaturated fatty acids (Forman and Zhang, 2021). L• and LOO• represent the lipid radical and peroxyl radical, respectively. The dual mode of action of classical uncouplers in ferroptosis may provide important insights into the mechanism controlling lipid peroxidation-related reactions in the membranes. The half-life for the decay of several micromolar O2 •− is about a few seconds at neutral pH in an aqueous solution (Takahashi and Asada, 1988). Since O2 •− is, in principle, membrane impermeable (Fig. 3), the O2 •− produced in hydrophobic regions or the aprotic interior within the membranes remains much longer unless protons are supplied [Eq. (2)]. At low concentrations, classical uncouplers or protonophores (proton channels) may have a potential to deliver protons into the aprotic interior of the membranes where O2 •− has accumulated.
O2 •− generating site in membranes
To explore the reactions associated with O2 •− generation, the thylakoid membranes offer an obvious advantage as they proved the Mitchell's chemiosmotic theory in the 1960s (Jagendorf and Uribe, 1966). Unlike respiratory electron flow in the inner membrane of mitochondria, the electron flow is easily controlled by light exposure with a high time resolution. Even a single electron transfer can be elicited by using a flash of light, thereby enabling us to chase rapid sequential reactions associated with ROS formation (Kruk et al., 2003; Sonoike et al., 1995). In their pioneering study, Takahashi and Asada (1988) clarified that the classical uncoupler NH4Cl, as well as methylamine and gramicidin D, induces H2O2 production and O2 uptake in illuminated thylakoid membranes, providing evidence for O2 •− generation at the aprotic interior of the thylakoid membranes.
At low concentrations, beyond those needed for uncoupling, proton conductors (classical uncouplers or protonophores) are suggested to deliver protons to such aprotic or hydrophobic microenvironments where O2 •− is generated by redox components (Kozuleva et al., 2011). This could facilitate the spontaneous disproportionation of O2 •− to generate H2O2 [Eq. (2)], which diffuses into the aqueous bulk phase before reacting with O2 •− within the membrane (Fig. 5). Consequently, proton delivery into the O2 •− generating site could prevent lipid peroxidation, which is required for inducing ferroptosis. Despite the theoretical importance (Fig. 5), little is known what mechanism might transfer protons into ROS generated in membranes.
Localized H+ at the Liquid–Membrane Interface
The chemiosmotic theory provides a framework for bioenergetics. However, there has been a long-standing debate on whether delocalized (bulk aqueous) or localized (liquid–membrane interface) pmf contributes to ATP synthesis in energy transducing membranes (Ort et al., 1976; Tedeschi, 2005). Thus, the significance of localized pmf at a liquid–membrane interface remains a subject of debate even in the field of bioenergetics (Lee, 2021; Nicholls, 2005; Xiong et al., 2010).
Surface potential generated by macromolecules
In addition to the conventional membrane potential difference between two aqueous bulk phases (Δψ∞), surface charges of the membrane create the surface electrical potential between the membrane surface and nearby aqueous phase (Barber, 1982). Distribution of charged molecules such as ions follows the surface potential. As a result, pH theoretically becomes lower on the surface at the negatively charged density and higher at the positively charged density (Itoh and Nishimura, 1986).
The charge state of membrane proteins is primarily determined by the ionization of their side chains of amino acid residues: the side chains of glutamate (Glu) and aspartate (Asp) are negatively charged, while those of lysine (Lys) and arginine (Arg) carry a positive charge at neutral pH. Modifications on the side residues such as acetylation or succinylation of Lys alter the charge state (Trist et al., 2021). Consequently, these charged molecules create a charge on the local membrane surface, thereby forming the surface potential. In addition, the hydrophilic head groups of lipid molecules form partial charges on the surface of the lipid bilayer.
These charges, either negative or positive, generate the local surface potential of the membrane, which influences the distribution of H+ and presumably the ionic form of RONSS as well. Generally, proteins and lipids with positive charges are less abundant, leading to a typically negatively charged exterior membrane surface at neutral pH. The surface potential is involved not only in dynamic changes of ion distributions near the membranes (Itoh, 1978) but also in the regulation of ion permeability of membranes such as in mitochondrial permeability transition pore (PTP) (Bernardi, 1996). Interestingly, cationic polyamines (e.g., spermidine and spermine) and amphipathic polypeptides (e.g., antimicrobial peptides [AMPs]) have been reported to exhibit unique multifunctionalities in both animals (Bahar and Ren, 2013; Gao et al., 2021; Madeo et al., 2020; Minois et al., 2011) and plants (Gupta et al., 2016; Kunstler et al., 2020).
It is of a great interest to investigate the effects of these cationic molecules on modulation of the surface potential, which can alter the local pH or proton availability at the liquid–membrane interface.
Hydrogen Bonded Networks for Proton Transfer
H-bonded network
The original proposal of the chemiosmotic theory postulated that energy-transducing membranes are impermeable to protons (H+). This allows the formation of a pmf between two compartments. Consequently, the presence of protons (H+) is strictly limited within these membranes, functioning as a diffusion barrier.
Protons have been believed to transfer along the hydrogen bonds of water, a mechanism referred to as “Grotthuss wires/bridges” or the “Grotthuss mechanism” for hydrogen-bonded water wires (Fig. 6) (Marx, 2006). Techniques involving crystallography and spectroscopy, coupled with computational simulations, have unveiled that proteins can also form proton transfer channels composed of an array of ionizable polar residues and water molecules (Ishikita and Saito, 2014; Luecke et al., 1998). These polar groups form an “H-bonded network” that is analogous to the one observed in water (Ishikita and Saito, 2014). A mechanism involving H-bonded networks has been proposed for the proton transport activity of the AMP gramicidin (Pomes and Roux, 2002), which has been known as a classical uncoupler to diminish pmf in mitochondria, chloroplasts, and bacteria (Yamasaki et al., 1991).
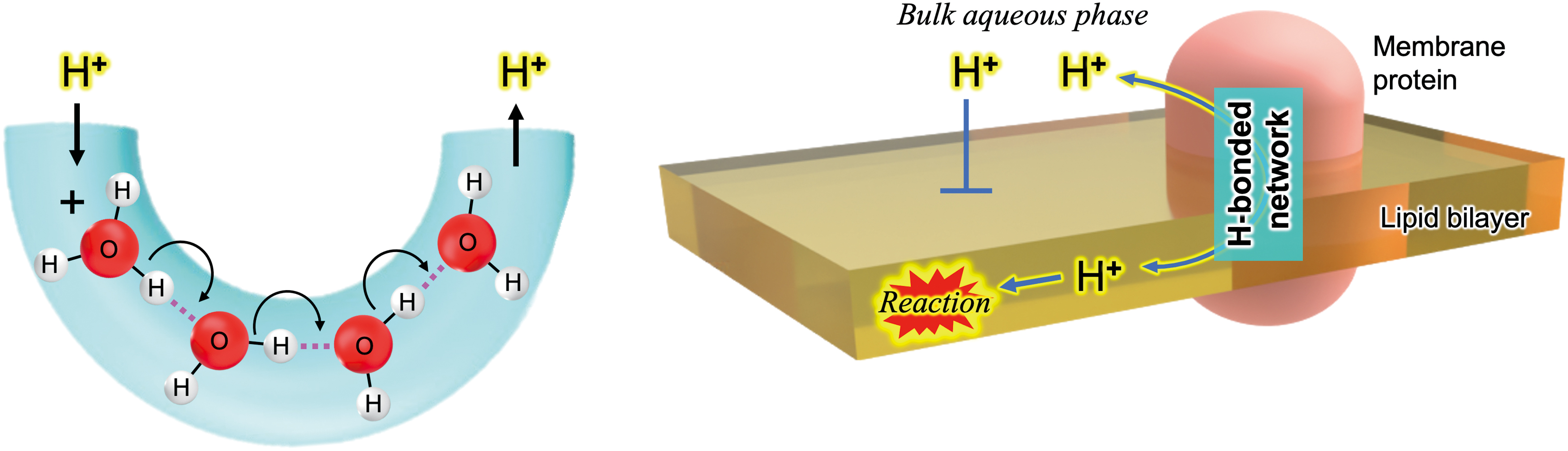
Proton exchange between hydrophobic and hydrophilic microenvironments
Intraprotein proton transfer through H-bonded networks has been extensively explored to reveal the energy conversion processes in biological systems. Examples include the OEC of PSII (Ishikita and Saito, 2014) and bacteriorhodopsin in Halobacterium salinarum, an archaeon (Luecke et al., 1998; Rammelsberg et al., 1998), both of which are involved in generating a pmf across membranes. Even in soluble enzymes, such as SOD, that require H+ for their reactions [Eq. (2)], it is suggested that H-bonded networks facilitate proton transfer to the enzyme's catalytic site (Trist et al., 2021).
While evidence is limited, it is thought that internal water molecules included in the membranes may exchange their protons with water molecules on the membrane surface, thereby constructing a proton transfer channel (Serowy et al., 2003). In membrane-bound proteins, the internal H-bonded network-based proton channels require strict spatial arrangements and precise distances between polar groups to form an H-bonded wire capable of exchanging protons between the membrane interior and the bulk aqueous phase. Thus, the reactions associated with ROS, such as the H2O2 generated from O2 •−, in the interior of lipid bilayers, can be regulated by proton uptake or release through the H-bonded networks (Fig. 6).
Subcellular Structures for H2O2 Transport?
Both mitochondria and chloroplasts, owing to their ancestral bacterial origins, may retain a primordial form of communication with the host nucleus of eukaryotic cells. Indeed, these organelles possess their own genomes, although highly reduced. The majority of proteins required by mitochondria and chloroplasts are encoded by nuclear DNA, synthesized by cytoplasmic ribosomes, and imported into the organelles. In this system, the cell nucleus serves as the “command center,” issuing “anterograde” signals to mitochondria and chloroplasts. Conversely, the state of these organelles can affect the expression of certain nuclear genes, indicating “retrograde” communication, where signals travel from the organelles back to the nucleus. Retrograde signaling is an evolving field linked with the evolution of eukaryotic cells, multicellularity, pathogen infections, and the chronic responses of mitochondria and chloroplasts.
As recently discussed by Speijer et al. (2020), ROS are considered to play significant roles in the evolution of these two organelles, which have an endosymbiotic origin. To date, numerous studies suggest that H2O2 functions as a retrograde signaling molecule in intracellular communication (Itoh, 2023).
Nanotubes
The nanoconduits or nanotubes exist ubiquitously across all domains of life, from archaea and bacteria to plants and mammals, suggesting they play crucial roles in evolution (Gozen and Dommersnes, 2020). The formation of nanotubes has been found to be associated with the infection mechanisms of bacteria or viruses in plants (Gozen and Dommersnes, 2020). It is plausible that nanotube formation and functionality in plants and animals could have been inherited from their bacterial ancestors during endosymbiotic evolution. There is growing interest on the function of these nanotubes as a conduit for H2O2 translocation.
Stromules
Chloroplasts are recognized as internal sensors of environmental stressors (Murata et al., 2007), influencing photosynthesis and transmitting information to the nucleus to coordinate plant growth, development, and stress responses (Seo et al., 2023). Stromules or stroma-filled tubules (Köhler and Hanson, 2000), the thin tubular extensions from plastids or chloroplasts, have been demonstrated to be induced by both biotic and abiotic stressors, suggesting their roles in stress-associated signal transduction from the plastid body. Caplan et al. (2015) provided evidence for the movement of H2O2 from the plastid to the nucleus. To monitor change of H2O2 level in the cells, they applied a fluorescent sensor protein called HyPer (Belousov et al., 2006) in tobacco leaves activated by p50 effector from Tobacco Mosaic Virus. The stroma-targeted HyPer signal increased at the interface between plastids/stromules and the nucleus.
In addition, when H2O2 bursts were induced in perinuclear plastids by laser at the region of interest, the nuclear-targeted HyPer signal also increased. H2O2 is therefore suggested a strong candidate for the role of retrograde signaling molecules in chloroplasts (Foyer and Hanke, 2022). However, there is currently no evidence for the fusion of plastid envelope membranes with the nucleus (Hanson and Hines, 2018).
Matrixules and peroxules
Mitochondria have been found to transiently form tubular membrane extensions, termed “matrixules” (Logan et al., 2004; Scott et al., 2007) or mitochondrial “nanotunnels” in both plant and animal cells (Lavorato et al., 2020; Vincent et al., 2017). These mitochondrial extensions in mammalian cells have been shown to be induced by various stress conditions, such as dysregulation of calcium homeostasis (Lavorato et al., 2017), exposure to manganese (Morcillo et al., 2021), and inhibition of respiratory complex III (Yao et al., 2020). The function of these stress-induced extensions remains unclear, but increased mitochondrial interconnectivity through “nanotunneling” might provide a protective effect (Carmichael et al., 2023).
Peroxisomes can also intermittently extend thin membrane tubules, similar to stromules, which is designated as “peroxules” (Scott et al., 2007). Although the dynamic nature of these organelle extensions was initially extensively studied in plant cells (Mathur et al., 2012), similar structures have subsequently been reported in mammalian cells (Carmichael et al., 2023), suggesting that the formation of thin tubules from mitochondria and peroxisomes may be a common feature shared among multicellular eukaryotes. Similar to stromules, the formation of peroxules in plant cells has been observed to be induced by exogenous H2O2 (Sinclair et al., 2009).
Given the ability of H2O2 to readily diffuse across lipid bilayers (Fig. 3), it seems unlikely that nanotubes would act as conduits for H2O2 transfer from organelles to the nucleus or other organelles. Consequently, a critical area for future research is to ascertain whether these nanotube structures are involved in the translocation of H2O2, other signaling molecules, metabolites, or macromolecules, such as proteins, between subcellular compartments. There may exist unidentified mechanisms that efficiently convey the H2O2 signal to the nucleus (Foyer et al., 2020; Mittler et al., 2022; Sies and Jones, 2020).
Recently, Imachi et al. (2020) successfully cultured one of the closest living Archean relatives of eukaryotic cells, an Asgard archaeon. Before their discovery, it had been presumed that this eukaryotic ancestor would be a large cell capable of potentially engulfing other cells for food. However, their finding has suggested that the ancestor was likely small, similar in size to this Asgard archaeon (∼550 nm in diameter), and utilized membrane “nanotube” extensions to exchange nutrients with other cells (Imachi et al., 2020). This ancestor may have enveloped the proto-mitochondrial bacterium as a means of protection against O2 toxicity. Beyond their functions in eukaryotic cells, the occurrence of nanotubes in an early life remains an important area for further investigation.
Localization of Redox Events
Localized versus delocalized reactions
It is now established that H2O2 serves both as an intracellular signaling molecule in mammals, plants, and bacteria, and as a cause of oxidative damage. This dual nature of H2O2 is often attributed to its concentration or “level” (Mittler, 2017; Sies and Jones, 2020). However, it is critical to recognize that concentration is a relative measure that fluctuates depending on the volume of a cellular compartment. For instance, decreasing the volume of vesicles can increase the concentration. It is known that organelles such as mitochondria (Massari and Azzone, 1972) and chloroplasts (Caplan et al., 2015; Dilley and Vernon, 1965) can dynamically alter their volume, such as demonstrated by observed swelling. Unlike in vitro model experiments that involve adding a known amount of chemicals to bulk aqueous solutions, evaluating the physiological effects of ROS based on concentration requires careful consideration.
Moreover, an increase in the H2O2 concentration of a bulk aqueous solution may lead to unfavorable and uncontrollable reactions, such as the Fenton reaction to produce •OH [Eq. (9)]. In this context, we propose that delocalized or bulk phase ROS generations could potentially cause harm (oxidative distress), as their rapid reactions with other molecules, such as NO [Eq. (1)], are challenging to control. In contrast, when ROS generation is localized and coupled with other signaling systems, they are not harmful due to their isolation from the bulk aqueous phase. This sequestration enables them to play a role in signaling without impacting regular metabolism (oxidative eustress). A finely tuned assembly of functional units is likely necessary for this localization mechanism. With respect to this localization mechanism, lipid microdomains are of considerable interest.
Membrane rafts: specialized microdomains orchestrating cellular signaling
Sterols are common in eukaryotic cells but rare in prokaryotes, including cyanobacteria, suggesting that sterols might have evolved adaptively to cope with an oxygenated environment after the GOE (Bloch, 1991). The plasma membrane in eukaryotic cells contains microdomains enriched with sterols (such as cholesterol), forming membrane/lipid rafts (MLRs) (Bieberich, 2018; Bloch, 1991; Lefebvre et al., 2007; Miserocchi, 2023). These regions may exist as caveolae (“little caves”), which can be morphologically observed as flask-like invaginations, or in a less observable planar form (Head et al., 2014; Navarro et al., 2014; Parton, 2018).
MLRs include O2 •− generators such as Nox (Anagnostopoulou et al., 2020) and RBOH (Noirot et al., 2014), NO generators such as endothelial NOS (Anagnostopoulou et al., 2020; GarciaCardena et al., 1996; Shaul et al., 1996) and neuronal NOS (Keshet et al., 2000; Marques-da-Silva and Gutierrez-Merino, 2014), signaling receptors (Del Pozo et al., 2021), and ion channels (O'Connell et al., 2004) that communicate extracellular stimuli to the intracellular milieu (Head et al., 2014; Roy and Patra, 2023). An increasing number of reports suggest that these microdomains play crucial roles in the physiology of animals (Head et al., 2014) and plants (Lefebvre et al., 2007; Luthje and Martinez-Cortes, 2018) as well as in mammalian caveolae (Anagnostopoulou et al., 2020; Del Pozo et al., 2021; Miserocchi, 2023; Navarro et al., 2014; Parton, 2018). These microdomains can be viewed as mechanisms to localize reactions with forming a functional unit.
On the contrary, the H2O2 or NO bursts observed in the bulk aqueous phase, which can be viewed as delocalized production observed in neutrophils during pathogen infections in mammals (Andres et al., 2022) and in the oxidative bursts in plants during pathogen infections (Noirot et al., 2014), are suicidal, even though they are part of the stress response against pathogens.
Spatial dynamics of RONSS: from beneficial to harmful roles
Small hydrophilic or water-soluble antioxidants, such as ascorbic acid (vitamin C) (Yamasaki et al., 2023), glutathione (Bersuker et al., 2019), and polyphenolic phytochemicals (Sakihama and Yamasaki, 2021), may be needed to intercept the delocalized (ambient or environmental) ROS that have leaked into the bulk aqueous phase under the condition where the localized regulated ROS generation is transformed into a delocalized or an uncontrolled one. Consequently, it could be inferred that the localization of reactions determines the role of RONSS in each biological system, either as harmful (when delocalized) or beneficial (when localized). In this context, cationic and/or amphipathic molecules, which can potentially influence not only the surface potential but also the assembly of microdomains such as the lipid rafts, may disrupt signaling mechanisms associated with RONSS. These disrupting mechanisms may be involved also in the pathogenesis of infectious diseases in both animals and plants.
In 2022, a new giant sulfur bacterium was discovered in the sulfurous waters of mangrove forests in the Guadeloupe Archipelago, found in sunken leaves (Levin, 2022; Volland et al., 2022). This bacterium tentatively named Thiomargarita magnifica has a thin layer of cytoplasm surrounding a large vacuole with stores of nitrate. Its length measures around a centimeter, making it visible to the naked eye, while most bacterial cells are around 2 μm in size. Just as the Nobel laureates Hodgkin and Huxley used the squid's giant axon to investigate how nerves generate and propagate signals (Schwiening, 2012), the giant sulfur bacteria may offer a unique opportunity to explore the spatiotemporal issues or localization mechanisms involved in RONSS metabolism within the interactive system of a bacterium (Fig. 3).
Conclusions
Proton (H+) concentration or pH of water plays crucial roles in biology, fundamentally underpinning the functioning of living systems. In biochemistry, enzyme activities involved in metabolism are altered by pH changes, in turn affecting the overall metabolic pathways in cells. Consequently, researchers use biological buffers to stabilize pH in their experiments. Among these buffers, “Good's buffers” have been widely used for an extended period in in vitro experiments within the life sciences. Good's buffers are a series of buffering agents with pK a values within the physiological pH range, developed by Good et al. (1966).
Hicks and Gebicki (1986) clarified the reactivity of 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethane-1-sulfonic acid (HEPES), N-[1,3-dihydroxy-2-(hydroxymethyl)propan-2-yl]glycine (Tricine), and Tris (2-amino-2-hydroxymethylpropane-1,3-diol), organic buffers with •OH, and reported their rate constants as 5.1 × 109, 1.6 × 109, and 1.1 × 109 M −1s−1, respectively. This indicates that these organic buffers are efficient scavengers of •OH. In addition to their pH-buffering actions, the antioxidant properties of Good's buffers often provide a significant advantage in preventing oxidative damage to cells during experiments and in protecting biomolecules from functional loss during isolation procedures, as recently demonstrated in the cryopreservation of red blood cells (Liu et al., 2022). Grady and coworkers (1988) suggested that Good's buffers with a piperazine ring, such as HEPES, which may produce nitroxide radicals in the presence of H2O2, should be avoided in studies of redox processes in biochemistry. The antioxidant behaviors of Good's buffers have not been fully recognized.
Consequently, using Good's buffers without careful consideration may lead to misinterpretation of results or could even result in artifactual outcomes in studies exploring cellular events associated with RONSS chemistry. We recommend that the selection of Good's buffers be approached with caution, taking into account their reactivity against reactive molecular species, as well as their charge status and membrane permeability, which may affect local pH or proton availability (Ort et al., 1976). Such consideration is crucial in experimental designs aimed at exploring the physiological functions of RONSS.
Despite the significant role of protons in biological activities, there have been a relatively limited number of studies that have confirmed the impact of proton availability on RONSS-related biological functions to date. A notable example of proton significance is seen in the therapeutic use of antacids to suppress excessive stomach acid production. As illustrated in Figure 1, NO can be produced nonenzymatically from inorganic nitrite (NO2 −) in aqueous solutions. Swallowed salivary nitrite—derived from dietary nitrates (NO3 −) in vegetables—is converted to NO under the strongly acidic conditions of gastric juice (Lundberg et al., 2008; Yamasaki et al., 2023). NO, along with the sulfur-containing compound thiocyanate (SCN−), produced from nitrite is considered to have a potent antipathogenic activity, inactivating swallowed pathogens (Yamasaki et al., 2023).
The strong acidity of gastric juice, approximately pH 1 in fasting conditions, is maintained by an H+/K+-ATPase (proton pump) in the gastric parietal cells (Sachs et al., 1988). Inhibition of this pump by antacid drugs or proton pump inhibitors, such as pantoprazole (Protonix), could impair the antipathogenic functions of gastric juice, potentially increasing the risk of oral infections (Zhou et al., 2017).
In the mid-20th century, researchers discovered that Ca2+ induces a sudden increase in the permeability of mitochondria, a phenomenon referred to as “permeability transition” (PT) (Halestrap, 2009). This mitochondrial PT can trigger organelle swelling, leading to cell death, a process associated with ischemia-reperfusion injury (Bernardi et al., 2023). This process is facilitated by a hypothetical channel often called the permeability transition pore (PTP), which is thiol-sensitive and significantly inhibited at pH levels below 7 (Halestrap, 2009). Several components, such as the F0F1-ATPase (the proton pump in mitochondria) and adenine nucleotide translocase, have been proposed as potential candidates for PTP (Bernardi et al., 2023). Given their nonselectivity for ions and solutes smaller than 1.5 kDa, protons may be translocated by PTP to dissipate the pmf, acting as endogenous uncouplers with a gating mechanism.
Mild uncoupling has been suggested to confer protection to cardiomyocytes against ischemia-reperfusion injury (Modriansky and Gabrielova, 2009). As discussed above, mild uncoupling can be characterized as a state in which transmembrane proton translocation is reduced, allowing proton exchange between the bulk aqueous phase on one side and the aprotic interior of the lipid bilayer. To regulate localized RONSS reactions within membranes via proton homeostasis, living organisms—depending on their evolutionary positions—may have developed various proton transfer mechanisms to control the spatiotemporal availability of protons or local pH. These mechanisms could involve weak acids (Takahashi and Asada, 1988), H+-translocating ATPase (Hisabori et al., 2013; Mitchell, 1961; Shima and Takagi, 2009), proton channels including UCPs (Modriansky and Gabrielova, 2009), cation channels (Lavorato et al., 2017), anion channels (Han et al., 2003), and PTPs (Bernardi et al., 2022).
We propose that the spatiotemporal translocation of protons into aprotic microenvironments significantly impacts RONSS-related chemical reactions, an effect that has been overlooked. This perspective could illuminate the pharmacological actions of chemical uncouplers (Childress et al., 2018; Modriansky and Gabrielova, 2009; Shrestha et al., 2021), polyphenols (Modriansky and Gabrielova, 2009; Sakihama and Yamasaki, 2021), and potentially some classes of AMPs (Andres et al., 2022; Manzini et al., 2014; Pastore et al., 2020) in therapeutic applications.
Footnotes
Acknowledgments
We thank Drs. George K. Iwama, Ichiro Terashima, Kintake Sonoike, Hiroshi Takagi, Toru Hisabori, and Yasuko Sakihama for their critical reading of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the JSPS KAKENHI grant number JP 23K05493 to Hideo Yamasaki.