
Abstract
Aim:
Azithromycin (AZM) is widely used to treat mycoplasma infection in pregnancy. However, there is no adequate evaluation of its side effect on the placenta. In this study, using human placental syncytiotrophoblasts and a mouse model, we investigated whether AZM use in pregnancy might adversely affect placental function and pregnancy outcome.
Results:
Transcriptomic analysis of AZM-treated human placental syncytiotrophoblasts showed increased expression of endoplasmic reticulum (ER) stress-related genes and decreased expression of genes for hormone production and growth factor processing. Verification studies showed that AZM increased the abundance of ER stress mediators (phosphorylated eIF2α, activating transcription factor 4 [ATF4], and C/EBP Homologous Protein [CHOP]) and decreased the abundance of enzymes involved in progesterone and estradiol synthesis (STS, CYP11A1, and CYP19A1) and insulin-like growth factor binding protein (IGFBP) cleavage (PAPPA and ADAM12) in human placental syncytiotrophoblasts. Inhibition of ER stress blocked AZM-induced decreases in the expression of CYP19A1, CYP11A1, PAPPA, and ADAM12, suggesting that the inhibition of AZM on those genes’ expression was secondary to AZM-induced ER stress. Further mechanism study showed that increased ATF4 in ER stress might repressively interact with C/EBPα to suppress the expression of those genes, including CEBPA itself. Mouse studies showed that AZM administration decreased fetal weights along with increased ER stress mediators and decreased levels of insulin-like growth factor, estrogen, and progesterone in the maternal blood, which could be alleviated by inhibition of ER stress.
Innovation and Conclusion:
These findings first support the fact that AZM, often used during pregnancy, may affect fetal growth by inhibiting crucial enzymes for estrogen and progesterone synthesis and disrupting crucial proteases for IGFBP cleavage via inducing ER stress in placental syncytiotrophoblasts. Antioxid. Redox Signal. 42, 16–35.

Introduction
Azithromycin (AZM) is a broad-spectrum macrolide antibiotic. It inhibits not only certain bacteria but also atypical bacteria such as chlamydiae, legionella, mycoplasma, and mycobacteria (Araujo et al., 1988; Heidary et al., 2022). Clinical data show that mycoplasma has the highest frequency of colonization in the amniotic cavity in preterm deliveries with intra-amniotic infection (Keelan et al., 2016). Therefore, AZM is widely prescribed for the treatment of mycoplasma infection in pregnancy with preterm premature rupture of membranes (Tanaka et al., 2019), as well as a measure of adjunctive prophylaxis in cesarean delivery (Tita et al., 2017). Although AZM use in pregnancy has generally been thought to be safe for the fetus (Administration, 2018), there are no adequate well-controlled studies on pregnant women (Antonucci et al., 2022). Studies in animals have indicated that AZM use in pregnancy may potentially carry multiorgan toxicity development in the fetus (Liu et al., 2020; Lu et al., 2023), retard fetal growth (Karabulut et al., 2008), and even increase the risk of spontaneous abortion (Muanda et al., 2017). In this context, it is necessary to carry out further evaluation of the potential side effects of AZM use in pregnancy to ensure its safety on the fetus.
The placenta plays a vital role in pregnancy maintenance and fetal development by serving not only as an interface for the delivery of vital nutrients and oxygen to the fetus (Maltepe and Fisher, 2015) but also as an endocrine organ that generates a bulk of peptide (Murphy et al., 2006) and steroid hormones (Pepe and Albrecht, 1995). Those functions of the placenta are accomplished mostly by the syncytial layer, a highly polarized multinucleated epithelium covering the outer surface of the villi, which derives from the underneath mononucleated cytotrophoblasts through syncytialization. In addition, the syncytial layer is also a source of proteolytic enzymes, including pregnancy-associated plasma protein A (PAPP-A) and A2 (PAPP-A2) (Conover and Oxvig, 2023; Monget and Oxvig, 2016; Nishizawa et al., 2008; Zhabin et al., 2003) and the secretory short isoform of a disintegrin and metalloprotease 12 (ADAM12-S) (Irwin et al., 2000; Jiang et al., 2012; Loechel et al., 2000; Shi et al., 2000), which cleave circulating insulin-like growth factor binding proteins (IGFBPs) (Zhu et al., 2022, Duan et al., 2010), in pregnancy. Proteolytic cleavage of IGFBPs is necessary to liberate insulin-like growth factors (IGFs) from IGFBPs so that the bioavailability of IGFs can be increased for the regulation of fetal growth (Jones and Clemmons, 1995; Han et al., 1996).
Innovation
AZM is widely used to treat mycoplasma infection in pregnancy. However, there is no adequate evaluation of its side effects on the placenta. In this study, we first revealed that AZM can cause ER stress in placenta syncytiotrophoblasts, thereby suppressing the expression and function of C/EBPα, which subsequently decreases the expression of enzymes related to estrogen and progesterone synthesis and IGFBP cleavage, leading to the inhibition of fetal growth.
Despite the crucial role of the placenta in pregnancy, there is not much literature regarding the potential adverse effect of AZM use in pregnancy on the placenta. In this study, we screened the biological pathway that might be adversely influenced by AZM in human placental syncytiotrophoblasts with transcriptome sequencing and found that the pathway related to the unfolded protein response (UPR) of the endoplasmic reticulum (ER) was enhanced, whereas the pathways related to steroid hormone synthesis and proteolytic cleavage of IGFBPs were attenuated.
The ER is an important organelle for protein folding, maturation, and trafficking. Physiological activities and environmental stimuli can lead to the accumulation of misfolded proteins, thereby evoking the UPR, also known as ER stress. Optimal ER stress, by altering transcriptional and translational activities, is necessary for the restoration of ER homeostasis (Hetz, 2012; Hetz et al., 2020). However, failure in restoration can be a cause of cell apoptosis (Han et al., 2013). Three key signal pathways can lead to ER stress, i.e., PRKR-like ER kinase (PERK), inositol requiring enzyme 1 (IRE1), and activating transcription factor 6 (ATF6) pathways. For instance, activation of the PERK pathway can reduce the protein load on the ER by suppressing translations through phosphorylation of the eukaryotic translation initiation factor-2α (eIF2α) (Wek et al., 2006). However, certain genes may gain a selective expression advantage in ER stress, such as the gene encoding the activating transcription factor 4 (ATF4) (Quirós et al., 2017). Given those characteristics of the UPR, and considering the potential enhancement of ER stress and attenuation of pathways related to steroid hormone synthesis and IGFBP cleavage by AZM in syncytiotrophoblasts, we hypothesized that AZM exposure might attenuate the gene expression related to steroid hormone synthesis and IGFBP cleavage by evoking ER stress in placental syncytiotrophoblasts, which might consequently disturb fetal growth. In this study, we examined the safety of AZM use in pregnancy on the placenta and fetus by addressing this hypothesis in primary human placental trophoblasts and a mouse model.
Results
Effect of AZM exposure on trophoblast viability and syncytialization
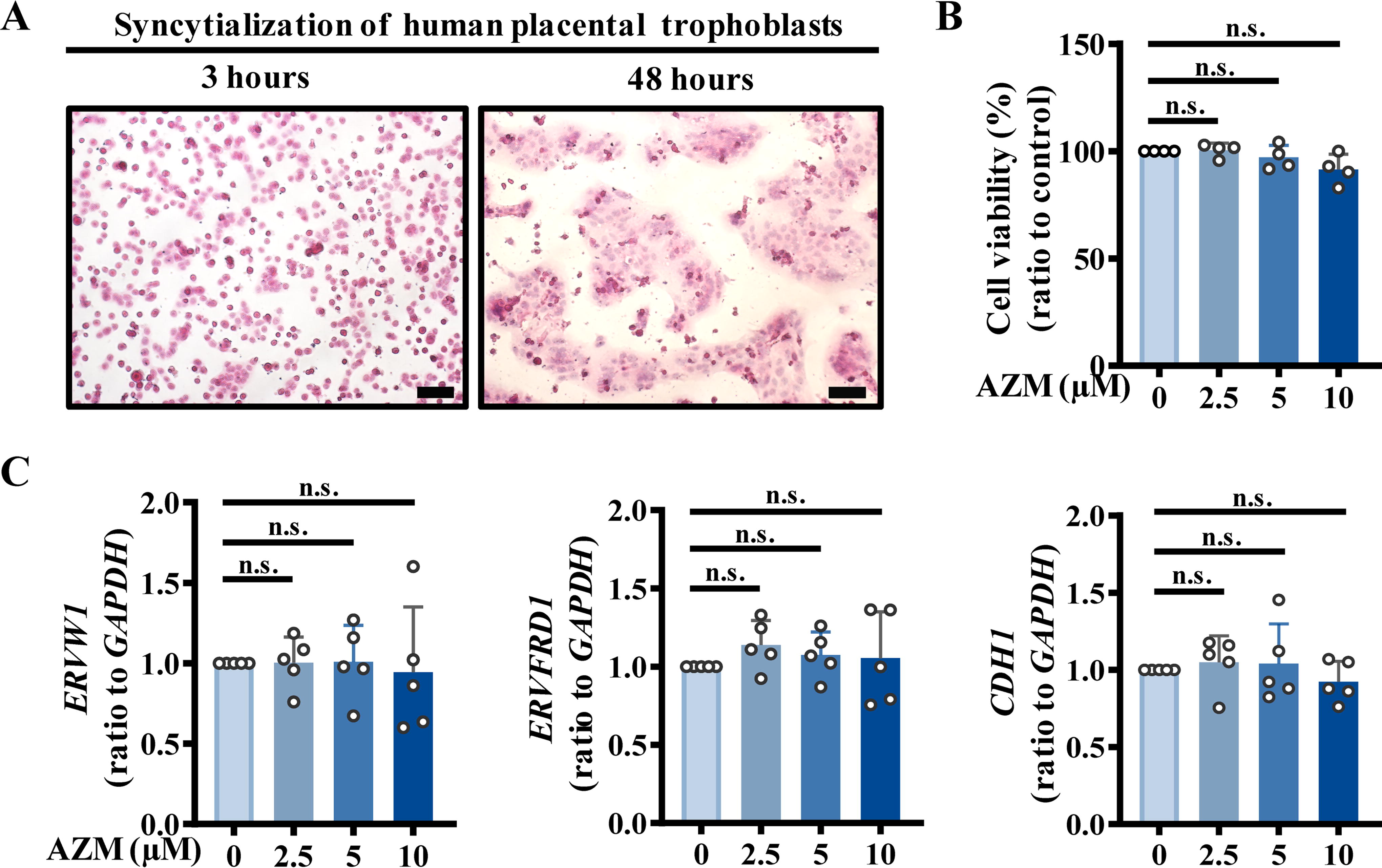
As has been well illustrated by us and others in previous studies (Kliman et al., 1986; Li et al., 2013; Wang et al., 2012), isolated primary human placental cytotrophoblasts were syncytialized progressively with incubation time when cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) (Fig. 1A). First, we examined whether AZM exposure might affect trophoblast viability and syncytialization. Assay with Cell Counting Kit-8 (CCK-8) showed that AZM treatment (2.5, 5, 10 μM; 24 h) (Fig. 1B) and (2.5 μM; 48 h) (Supplementary Fig. S1) did not affect the viability of syncytiotrophoblasts. However, treatment with AZM (2.5 μM) for 72 h significantly inhibited the cell viability (Supplementary Fig. S1), indicating that long-term culture may increase the vulnerability of cultured trophoblasts to AZM cytotoxicity. To avoid interference by conditions of cultured trophoblasts per se, AZM treatment for 24 h was used in subsequent studies. Exposure of trophoblasts to AZM (2.5, 5, 10 μM; 24 h) during syncytialization did not affect the syncytialization process with no alteration in the classical syncytialization markers, including ERVW1 (encoding endogenous retrovirus group W member 1, syncytin-1), ERVFRD1 (encoding endogenous retrovirus group FRD member 1, syncytin-2), and CDH1 (encoding calcium-dependent adhesion protein 1, E-cadherin) (Fig. 1C).
Bioinformatic analysis of the biological pathways potentially influenced by AZM in human placental syncytiotrophoblasts
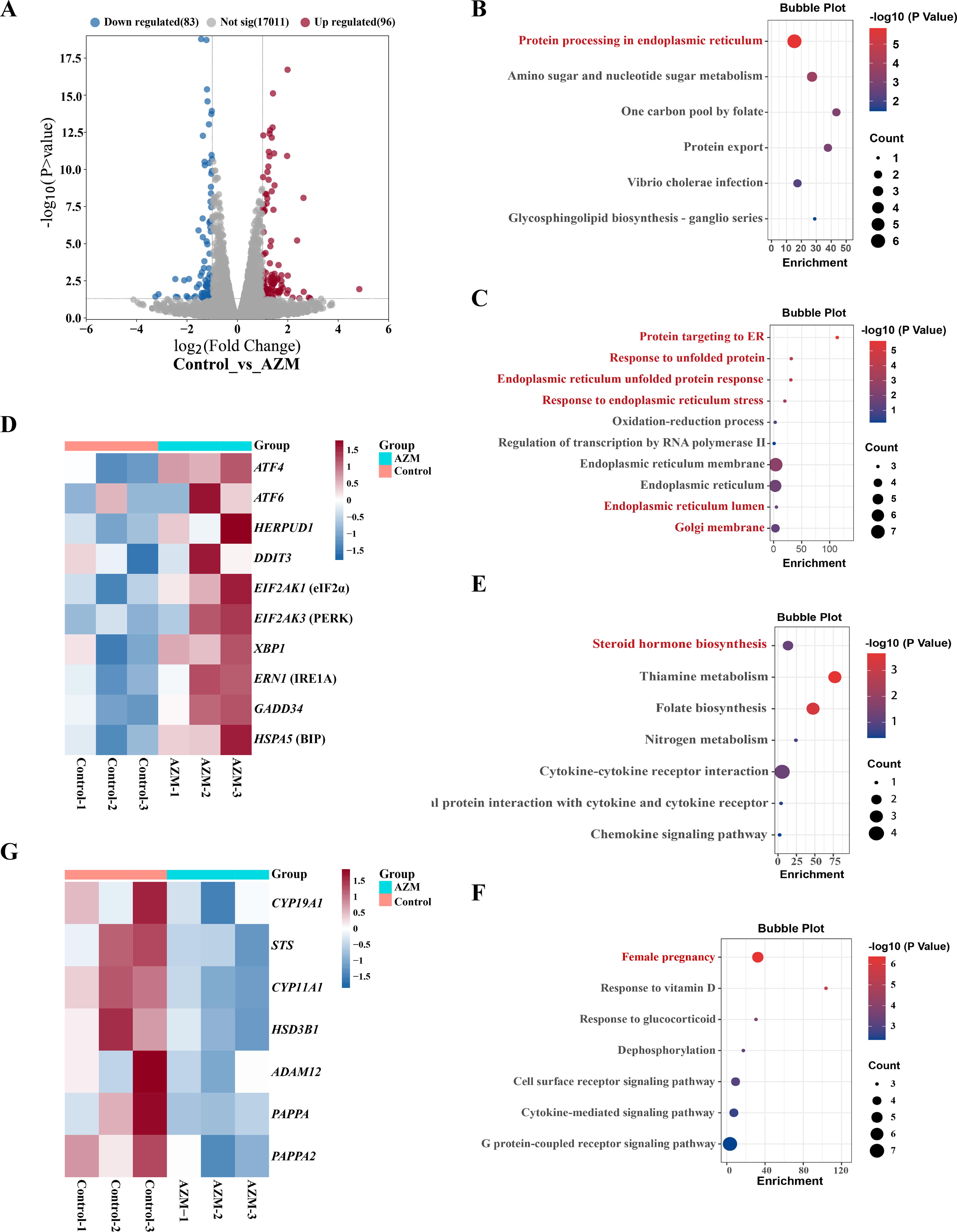
To screen the biological pathway potentially influenced by AZM exposure, transcriptome sequencing was performed in syncytiotrophoblasts after exposure to AZM (10 μM; 24 h). Results showed that there were 96 upregulated genes and 83 downregulated genes (fold change ≥2 or ≤0.5, adjusted p value <0.05) in syncytiotrophoblasts after exposure to AZM (Fig. 2A and Dataset S1). Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses were applied to identify the potential biological pathways influenced by AZM, which showed that the upregulated genes were enriched in the biological pathways related to ER stress (Fig. 2B and C), along with increased expression of heat shock protein family 70 member 5 (HSPA5) (encoding BiP), EIF2AK3 (encoding PERK), EIF2AK1 (encoding eIF2α), and ATF4 (encoding ATF4) in the PERK pathway and X-box binding protein 1 (XBP1) (encoding X-box binding protein 1, XBP1) in the IRE1 pathway (Fig. 2D), but not the genes in the ATF6 pathway (Supplementary Fig. S2A and B). Concomitantly, the common downstream genes underlying ER stress, including HERPUD1 (encoding Homocysteine-responsive endoplasmic reticulum-resident ubiquitin-like domain member 1, HERPUD1) (Schulze et al., 2005), DNA damage inducible transcript 3 (DDIT3) (encoding C/EBP Homologous Protein, CHOP) (Dalton et al., 2013), and GADD34 (encoding growth arrest and DNA damage-inducible protein, GADD34) (Choy et al., 2015) were also upregulated by AZM treatment (Fig. 2D). Analysis of the downregulated genes by AZM showed that these genes were enriched mainly in the pathways related to steroid hormone synthesis and female pregnancy (Fig. 2E and F). Specifically, the genes related to steroid hormone synthesis, including STS (encoding steroid sulfatase, STS), cytochrome P450 family 11 subfamily A member 1 (CYP11A1) (encoding cholesterol side-chain cleavage enzyme, P450scc), HSD3B1 (encoding 3 beta-hydroxysteroid dehydrogenase isomerase type 1, 3β-HSD1), and cytochrome P450 family 19 subfamily A member 1 CYP19A1 (encoding aromatase), and the genes related to IGFBP proteolytic cleavage, including PAPPA (encoding PAPP-A), PAPPA2 (encoding PAPP-A2), and a disintegrin and metalloprotease 12 (ADAM12) (encoding ADAM12-S), were downregulated by AZM treatment (Fig. 2G).
Induction of ER stress by AZM in human placental syncytiotrophoblasts
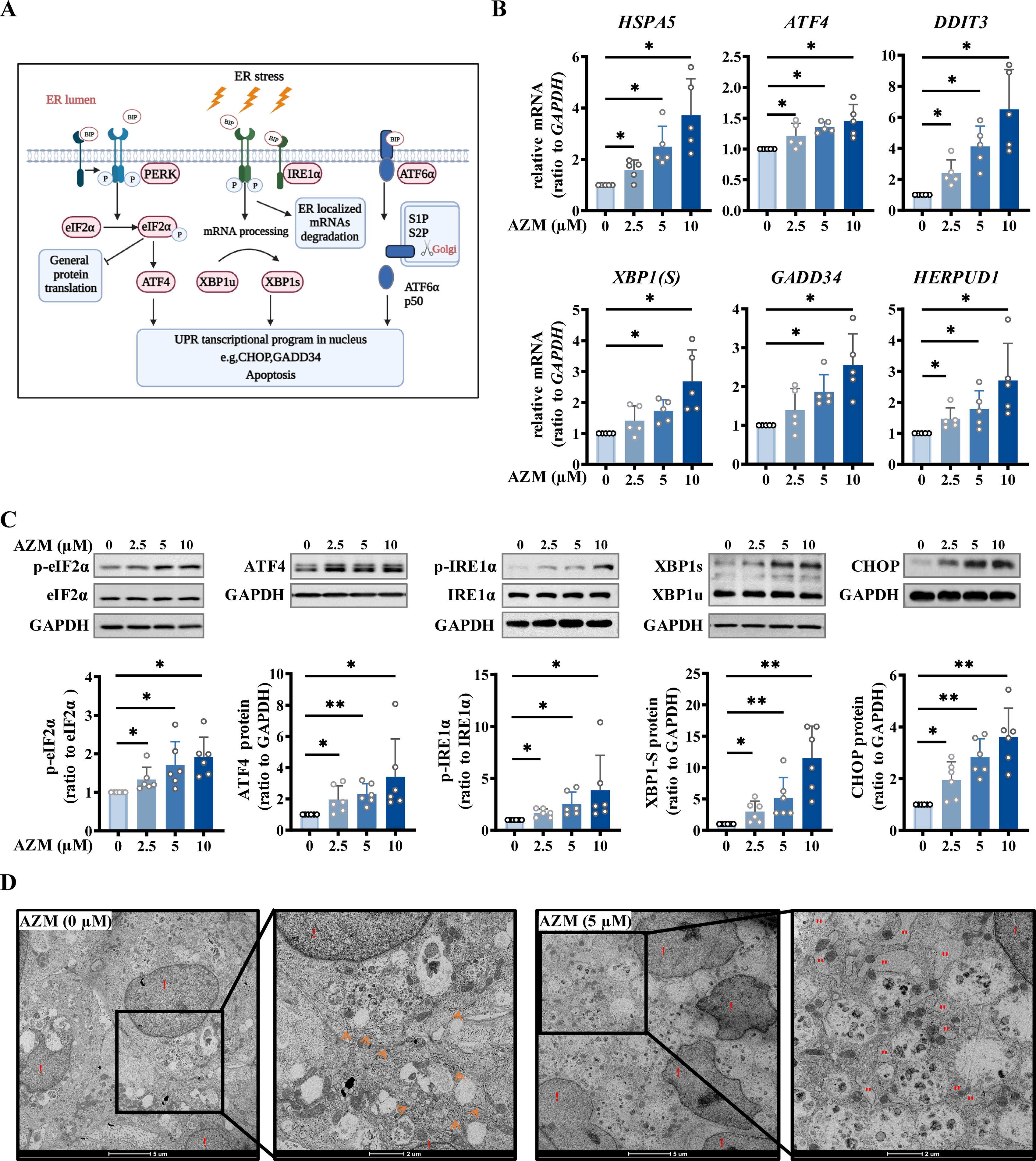
We went further on to confirm whether AZM exposure could indeed induce ER stress in syncytiotrophoblasts. Measurement with quantitative real-time PCR (qRT-PCR) showed that AZM (2.5, 5, 10 μM; 24 h) increased the mRNA transcripts related to ER stress in a concentration-dependent manner, including ATF4 and GADD34 of the PERK pathway, XBP1 of the IRE1 pathway, and the common downstream genes HSPA5, DDIT3, and HERPUD1 (Fig. 3A and B). Consistently, Western blotting analysis showed that AZM treatment (2.5, 5, 10 μM; 24 h) also increased the proteins related to ER stress in a concentration-dependent manner, including phosphorylated eIF2α (p-eIF2α) and ATF4 of the PERK pathway, phosphorylated IRE1α (p-IRE1α) and XBP1s of the IRE1 pathway, and the common downstream protein C/EBP Homologous Protein (CHOP) (Fig. 3C). Notably, significant induction was observed for all of these genes by AZM at 2.5 µM. In addition to 24 h, treatment of the cells with AZM (2.5 µM) for 48 h also increased the protein abundance of ATF4 and CHOP (Supplementary Fig. S3 A and B). Moreover, examination with transmission electron microscopy showed that enlarged ER lumens could be seen in syncytiotrophoblasts after exposure to AZM (5 μM; 24 h), a morphological signature of a dysfunctional ER (Fig. 3D). Since apoptosis may pursue ER stress (Hetz, 2012), we further examined whether AZM treatment affected cell apoptosis in syncytiotrophoblasts, which showed that neither the pro-apoptosis proteins (B-Cell CLL/Lymphoma 2 Associated X and cleaved-caspase-3) nor the antiapoptosis protein (B-Cell CLL/Lymphoma 2) was altered in syncytiotrophoblasts by AZM (2.5, 5, 10 μM; 24 h) (Supplementary Fig. S4A and B). Consistently, the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay also revealed no signs of cell apoptosis in syncytiotrophoblasts upon AZM (2.5, 5, 10 μM; 24 h) treatment (Supplementary Fig. S4C). These data suggest that AZM exposure could induce ER stress through activation of the PERK and IRE1 pathways in placental syncytiotrophoblasts but with no indication of apoptosis.
Inhibition of enzymes involved in estradiol and progesterone synthesis and IGFBP cleavage by AZM in human placental syncytiotrophoblasts
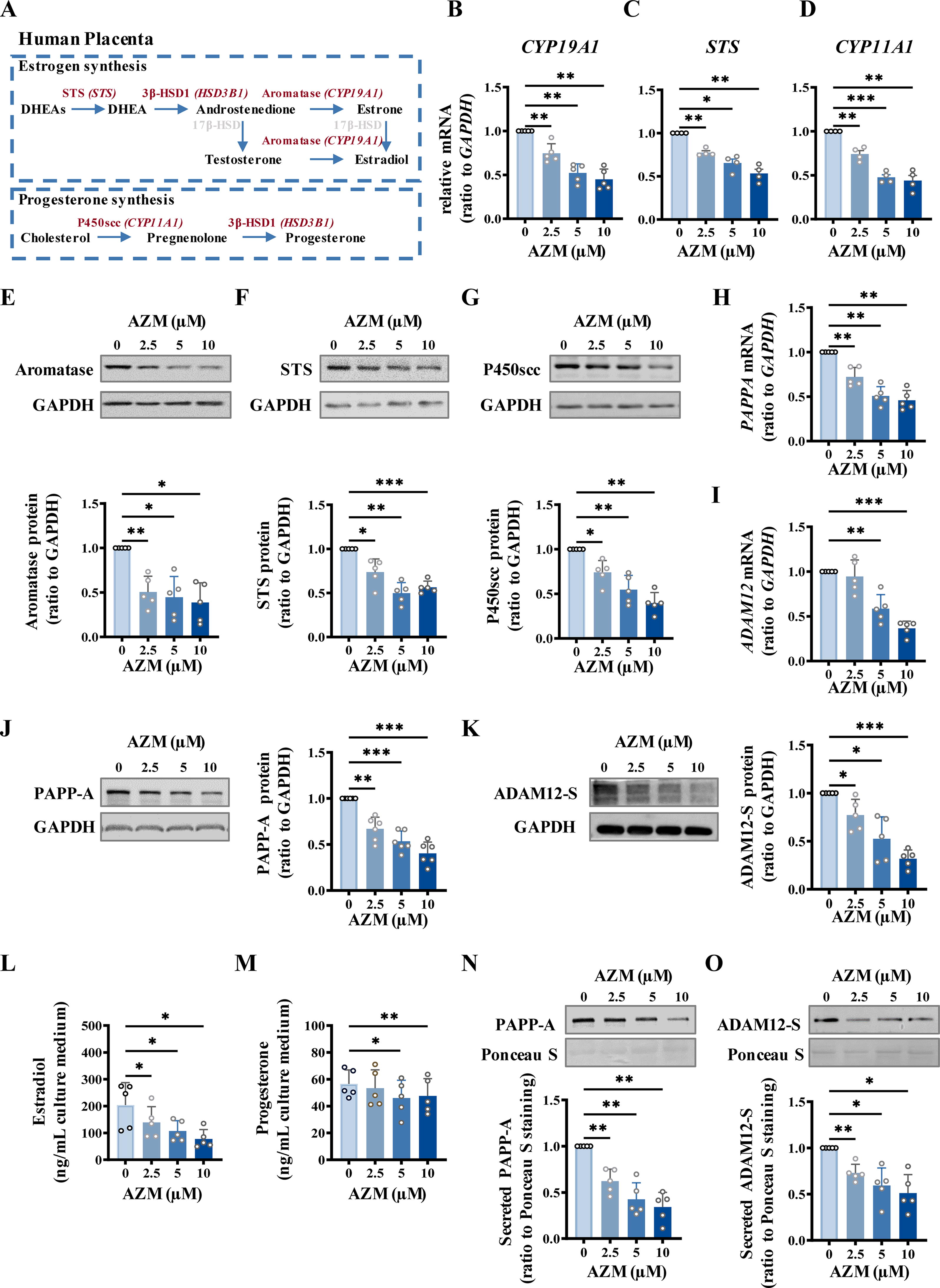
Next, we validated the suppression of gene expression by AZM in syncytiotrophoblasts. Analysis with qRT-PCR and Western blotting showed that the enzymes involved in estrogen and progesterone synthesis (STS, CYP19A1, and CYP11A1) and the proteases involved in IGFBP cleavage (PAPP-A and ADAM12-S) were all decreased at both mRNA and protein levels by AZM (2.5, 5, 10 μM; 24 h) (Fig. 4A–K), although the decreased expression of HSD3B1 and PAPPA2 was confirmed only at the mRNA level (Supplementary Fig. S5A–D). Consistently, the productions of estradiol (Fig. 4L), progesterone (Fig. 4M), PAPP-A (Fig. 4N), and ADAM12-S (Fig. 4O) by syncytiotrophoblasts were also significantly reduced by AZM (2.5, 5, 10 μM; 24 h). Again, these suppressive effects of AZM manifested in a concentration-dependent manner, with the significant suppression observed mostly at 2.5 µM of AZM.
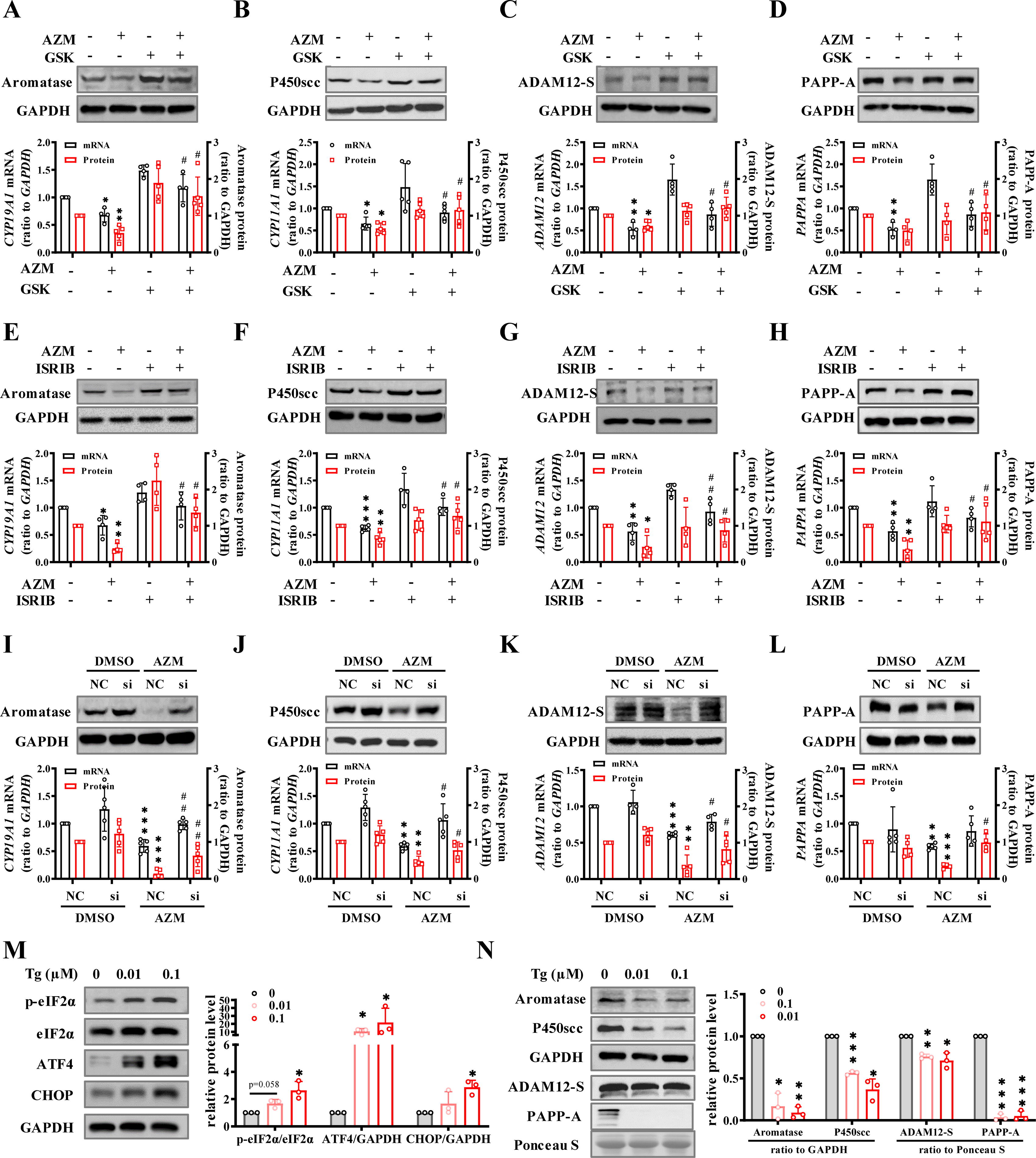
Involvement of ER stress in the inhibition of estradiol, progesterone, PAPP-A, and ADAM12-S synthesis by AZM in human placental syncytiotrophoblasts
Considering that ER stress is often accompanied by altered transcription and translation (Han et al., 2013), we anticipated that the suppression of gene expression by AZM might be secondary to ER stress induced by AZM. This notion was supported by the findings that the downregulation of CYP11A1, CYP19A1, ADAM12, and PAPPA mRNA and protein by AZM (5 μM; 24 h) could be blocked by either PERK inhibitor (GSK2656157, GSK, 2.5 μM) or p-eIF2α inhibitor (integrated stress response inhibitor, ISRIB, 0.1 μM) (Atkins et al., 2013; Schoof et al., 2021) (Fig. 5A–H), or by siRNA-mediated knockdown of ATF4 expression in syncytiotrophoblasts (Fig. 5I–L), but not by blocking IRE1α RNase activity of the IRE1 pathway with 4μ8c (2.5 μM) (Sato et al., 2017) (Supplementary Fig. S6), suggesting the involvement of the PERK rather than the IRE1 pathway in the downregulation of those gene expressions. Of interest, thapsigargin (TG) (0.01, 0.1 μM; 24 h), a widely used ER stressor (Lytton et al., 1991), not only increased ER stress mediators, including p-eIF2α, ATF4, and CHOP (Fig. 5M), but also decreased the abundance of P450scc, aromatase, ADAM12-S, and PAPP-A (Fig. 5N) in a concentration-dependent manner in syncytiotrophoblasts, suggesting that the suppression of gene expression related to steroid hormone synthesis and IGFBP cleavage may be a common effect in ER stress irrespective of the nature of the inducer as long as the PERK pathway is activated.
Involvement of C/EBPα in the suppression of gene expressions related to steroid hormone synthesis and IGFBP cleavage by AZM-induced ER stress in syncytiotrophoblasts
The CCAAT/enhancer binding protein (C/EBPα) is a member of the C/EBP family, which comprises six members of the basic-leucine zipper transcription factors. Our recent study has shown that C/EBPα expression is increased greatly during syncytialization, which is implicated in the enhancement of gene expression involved in steroid hormone synthesis (Zhu et al., 2023). In this study, we found that siRNA-mediated knockdown of C/EBPα significantly suppressed the basal expression of CYP11A1, CYP19A1, ADAM12, and PAPPA at both mRNA and protein levels (Fig. 6A and B), whereas overexpression of C/EBPα attenuated the suppression of these genes by AZM (5 μM; 24 h) in syncytiotrophoblasts (Fig. 6C). Moreover, chromatin immunoprecipitation assay (ChIP) showed that the enrichment of C/EBPα to the promoters of CYP11A1, CYP19A1, ADAM12, PAPPA, and CEBPA itself was also significantly decreased by AZM (5 μM; 24 h) in syncytiotrophoblasts (Fig. 6D). These data suggest that C/EBPα is a common transcription factor driving the expression of not only CYP11A1, CYP19A1, ADAM12, and PAPPA but also CEBPA itself in syncytiotrophoblasts. Interestingly, we found that both AZM (2.5, 5, 10 μM; 24 h) and TG (0.01, 0.1 μM; 24 h) decreased C/EBPα mRNA and protein abundance in a concentration-dependent manner in syncytiotrophoblasts (Fig. 6E–H). Moreover, in addition to CYP11A1, CYP19A1, ADAM12, and PAPPA expression (Fig. 5), inhibition of PERK with GSK (2.5 μM; 24 h), inhibition of p-eIF2α with ISRIB (0.1 μM; 24 h), or siRNA-mediated knockdown of ATF4 could also reverse the suppression of C/EBPα expression by AZM (5 μM; 24 h) in syncytiotrophoblasts (Fig. 6I–K). Co-immunoprecipitation (Co-IP) assay showed that ATF4 could heterodimerize with C/EBPα in syncytiotrophoblasts (Fig. 6L). These data suggest that the suppression of aromatase, P450scc, ADAM12-S, and PAPP-A expression by AZM-induced ER stress is possibly a consequence of the initial suppression of C/EBPα through interaction with the elevated ATF4 in syncytiotrophoblasts.

Inhibition of fetal growth by AZM in pregnant mice
Administration of AZM (200 mg/kg, once a day for three days) to pregnant mice by gavage on gestational days (GD) 15.5, 16.5, and 17.5 significantly decreased the fetus weight (GD18.5) but not the placenta weight (Fig. 7A and B) irrespective of sex along with an increased abundance of ER stress mediators, including p-eIF2α and ATF4 in the placenta (Fig. 7C), and enlarged ER lumen in the syncytiotrophoblast of the placental labyrinth layer (Fig. 7D). Enzyme-linked immunosorbent assay (ELISA) showed that AZM administration also decreased ADAM12 (Fig. 7E) and PAPP-A (Fig. 7F) abundance in the placenta and free IGF-1 (Fig. 7G), estradiol (Fig. 7H), and progesterone (Fig. 7I) levels in the maternal blood. Moreover, the concentration of free IGF-1 in serum correlated positively with the concentrations of ADAM12 and PAPP-A in the placenta (Supplementary Fig. S7A and B). Coadministration of ISRIB (10 mg/kg, once a day for 3 days) with AZM ameliorated these alterations induced by AZM, including the fetal weight and ER stress mediators in the placenta (Fig. 7B-1). These data suggest that ER stress in the placenta may contribute, at least in part, to the disturbed fetal growth by AZM use in pregnancy.

Discussion
Although AZM is classified as a Class B antibiotic by the FDA of the USA for use during pregnancy (Administration, 2018), the safety of AZM use during pregnancy has not been a topic without controversies. Animal studies have pointed to AZM use in pregnancy as a possible disruptor of fetal development, and even as a risk factor for spontaneous abortion (Karabulut et al., 2008; Liu et al., 2020; Lu et al., 2023; Muanda et al., 2017). In this study, we demonstrated that AZM use in pregnancy could cause ER stress in human placental syncytiotrophoblasts resulting in the suppression of genes associated with steroid hormone synthesis and IGFBP cleavage. These findings were recapitulated in vivo in the mouse along with the consequence of reduced fetal body weight. This growth-retarding effect of AZM on the fetus is in line with previous studies in the mouse, which showed that AZM use in pregnancy decreased the crown-rump length and the birthweight of the offspring (Karabulut et al., 2008; Liu et al., 2020; Lu et al., 2023). Notably, we demonstrated for the first time that AZM exposure might cause ER stress in both human and mouse placenta, which is in line with the findings in other tissues (Mansour et al., 2021; Moriya et al., 2013; Yu et al., 2023). Moreover, we found that fetus growth-retarding effect of AZM might be associated with ER stress in placental syncytiotrophoblasts.
The ER is a cellular organelle that is responsible for the processing and trafficking of newly synthesized proteins. Compounds that disrupt protein glycosylation such as tunicamycin (Wu et al., 2018), disulfide bond formation such as dithiothreitol (Keestra-Gounder et al., 2016), and chaperone function such as TG, or brefeldin A which induces retrograde protein transport from the Golgi to the ER (Oh-Hashi et al., 2021), can all induce ER stress. In this study, we found that TG carried effects similar to AZM on the enzymes involved in steroid hormone synthesis and IGFBP cleavage in syncytiotrophoblasts, suggesting that ER stress may impair these biological pathways in syncytiotrophoblasts irrespective of the nature of the inducers as long as the PERK pathway is activated, since our study demonstrated that only the PERK pathway appeared to be involved in those effects induced by AZM. This speculation requires further proof from studies using other ER stress inducers, particularly those that specifically activate the PERK pathway. In this study, we demonstrated that neither apoptosis nor cell viability was affected by AZM exposure, indicating that the observed suppression of gene expression in syncytiotrophoblasts by AZM was unlikely due to cell apoptosis or cell death.
At present, three key signal transduction pathways, i.e., PERK, IRE1, and ATF6 pathways, have been recognized to mediate ER stress. Of interest, we found that AZM treatment could activate the PERK and IRE1 pathways but not the ATF6 pathway in syncytiotrophoblasts. Furthermore, it appeared that only the PERK pathway but not the IRE1 pathway was responsible for the inhibition of gene expressions related to steroid hormone synthesis and IGFBP cleavage by AZM in syncytiotrophoblasts. Our study seems to be consistent with previous studies showing that environmental xenobiotics, such as cadmium (Kim et al., 2021; Zhu et al., 2021), polybrominated diphenyl ethers (Zhao et al., 2022), perfluorooctanoate acid (Du et al., 2022), and cyfluthrin (Ni et al., 2022), could all impair fetal and placental growth by inducing ER stress through activation of the PERK pathway in the mouse placenta. It is intriguing why only the PERK pathway is involved in the suppression of these genes in syncytiotrophoblasts. Interestingly, ATF4, the transcription factor of the PERK pathway, was reported to be capable of heterodimerizing with C/EBPα at the C/EBP binding site in the promoters of the myeloid-specific genes, thus creating a transcriptionally less active heterodimer than the C/EBPα homodimer, resulting in the attenuation of the transcriptional activity of C/EBPα (Gombart et al., 2007). In this study, we also found a possible suppressive interaction between ATF4 and C/EBPα, a common upstream transcription factor driving the expression of not only the enzymes related to steroid hormone synthesis and IGFBP cleavage but also C/EBPα itself in syncytiotrophoblasts. Our findings might explain why only activation of the PERK pathway was involved in those adverse effects of AZM in syncytiotrophoblasts.
Currently, the precise molecular pathways by which AZM triggers ER stress are not fully understood. In the study by Moriya et al., it is hypothesized that AZM might interfere with the ubiquitin-proteasome system, leading to an accumulation of misfolded proteins in multiple myeloma cells (Moriya et al., 2013). The study by Yu et al. showed that AZM (40 μM) exposure induced ER stress in murine fibrosarcoma cells via activation of the P-eIF2α/ATF4/CHOP pathway following accumulation of reactive oxygen species (Yu et al., 2023). In addition, Mansour et al. showed that AZM treatment was associated with a significantly lower concentration of glutathione (GSH) in the heart tissue (Mansour et al., 2021). When GSH is depleted, the ER may become more susceptible to oxidative stress, leading to the accumulation of misfolded proteins (Haynes et al., 2004).
IGFs include IGF-I and -II, which are the major regulators of fetal growth (Agrogiannis et al., 2014). However, circulating IGFs are mostly bound by IGFBPs, thereby sequestering IGFs from their receptors (Allard and Duan, 2018; Han et al., 1996). To increase the bioavailability of IGFs, levels of ADAM12-S and PAPP-A increase with gestational age in the maternal blood so that increased IGFBP cleavage can be achieved (Chełchowska et al., 2016; Jiang et al., 2012). Lines of evidence have indicated that reduced ADAM12 and PAPPA levels in pregnancy are associated with fetal growth restriction (Gaccioli et al., 2018; Pihl et al., 2008; Sifakis et al., 2018; Zhu et al., 2022), further highlighting the crucial role of ADAM12- and PAPPA-mediated IGFBP cleavage in the regulation of fetal growth. Therefore, we believe that the fetal growth restriction observed in this study was attributed, at least in part, to the suppression of ADAM12-S and PAPP-A productions by AZM in placental syncytiotrophoblasts.
Progesterone and estradiol are crucial steroid hormones in pregnancy, which are implicated in pregnancy maintenance and maternal adaptation to pregnancy (Pepe and Albrecht, 1995). Previous studies have demonstrated that ER stress caused by tunicamycin and palmitic acid can disrupt the synthesis of these steroid hormones in granulosa cells (Guzel et al., 2017; Park et al., 2014). In the present study, we also found that AZM-induced ER stress could reduce these steroid hormone syntheses in syncytiotrophoblasts. However, unlike humans, the corpus luteum rather than the placenta is the major site of progesterone and estradiol synthesis in mouse pregnancy (Stocco et al., 2007). Nevertheless, AZM administration also decreased progesterone and estradiol levels in the maternal blood in the pregnant mouse, suggesting that AZM may also disturb these steroid syntheses in the corpus luteum possibly by evoking ER stress in the mouse ovary. Previous studies have demonstrated that estradiol and progesterone levels in the maternal blood were significantly reduced in pregnancy with fetal growth restriction (Anelli et al., 2019; Salas et al., 2006), suggesting that adequate estradiol and progesterone are necessary for the normal growth of the fetus. To this end, decreased estradiol and progesterone production may also be a contributing factor to the fetal growth retardation induced by AZM. However, despite the crucial role of progesterone in pregnancy maintenance, we failed to observe preterm birth in the pregnant mouse with AZM administration, which is somewhat in contrast to the observation of abortion in previous studies (Muanda et al., 2017). This discrepancy may be due to the gestational day when AZM was administered. In this study, AZM was given to the pregnant mouse from GD15.5 to GD17.5, a period corresponding to the third trimester of human gestation when pregnancy is well established but rapid growth of the fetus occurs. The gestational period for AZM administration may also account for the observation of the retarded growth of the fetus but not the placenta in this study, since the development of the mouse placenta is accomplished mostly before GD13.5 (Elmore et al., 2022). In this context, it is reasonable to predict that AZM use in early gestation may inhibit placental growth and incur spontaneous abortion as reported in a previous study (Muanda et al., 2017).
Several studies have measured AZM concentrations in the blood and tissues after AZM administration. Ramsey et al. demonstrated that the peak serum concentration of AZM was about 311 ng/mL (about 0.5 μM) after taking 1 g AZM orally, but the concentration of AZM in the placenta could build up to 2130 ng/mL (about 2.84 μM) (Ramsey et al., 2003). Ramaiah et al. found that AZM concentration in the blood and other organs (e.g., lung, spleen, and liver) could reach up to 10 μg/mL or μg/g (about 13.35 μM) at 12 h after intravenous injection of AZM solution with 50 μg/g body weight in mice (Ramaiah et al., 2016). Moreover, in the study by Sutton AL et al., pregnant women received 500 mg of AZM intravenously 15, 30, or 60 min before cesarean delivery, and the maximum concentration was 4500 (±2430) ng/mL (approximately 2.67–10 μM) in the plasma and 961 ng/g in the placental tissue (Sutton et al., 2015). Given the robust tissue penetration ability of AZM, as well as the half-life of 78 h in pregnant women (Salman et al., 2010), the concentration of AZM in the placental tissue may further increase after multiple doses. In the present study, we found that AZM could induce ER stress in the placenta at concentrations within the range of plasma or tissue concentrations that can be achieved clinically, suggesting that our findings in this study may carry clinical implications. In other words, the safety of AZM use in pregnancy, particularly on the placenta and fetus, should be taken seriously. Since findings in the mouse may not truthfully reflect the situation in humans due to inherent species-specific differences in gestation length, it is necessary to carry out large-scale retrospective and prospective clinical studies to further address this issue.
Conclusion
We have demonstrated that AZM use in pregnancy may cause ER stress in placenta syncytiotrophoblasts, thereby suppressing the expression and function of C/EBPα, which subsequently decreases the expression of enzymes related to steroid hormone synthesis and IGFBP cleavage, resulting in the inhibition of fetal growth. Based on the findings in this study, we recommend that caution should be taken for the use of AZM in pregnancy, and AZM use should be limited to situations when the benefits of treatment are expected to outweigh the potential risks during pregnancy. However, it should also be kept in mind that the findings in the mouse may not truthfully reflect the situation in humans due to inherent species-specific differences in gestation length. It is necessary to carry out large-scale retrospective and prospective clinical studies to further address this issue.
Materials and Methods
Isolation and culture of primary human placental trophoblasts
Human placentas were obtained from uncomplicated term (38 to 40 weeks) pregnancies after elective cesarean section without labor with written informed consent following a protocol approved by the Ethics Committee of Ren Ji Hospital, Shanghai Jiao Tong University School of Medicine, and in accordance with Declaration of Helsinki. Placental cytotrophoblast cells were prepared utilizing a modified Kliman’s method (Kliman et al., 1986) as described previously (Wang et al., 2012, Li et al., 2013). Briefly, the villous tissue cut from the maternal side of the placenta was washed thoroughly and then minced for digestion with 0.125% trypsin (Sigma Chemical Co., St. Louis, MO) and 0.03% deoxyribonuclease I (Sigma) in DMEM (Gibco, Grand Island, NY) containing 1% antibiotics (Gibco) at 37°C. The dispersed placental cells from 3 digestions were pooled together for purification with 5%–65% Percoll (GE Healthcare Bio-Sciences, Uppsala, Sweden) discontinuous density gradients. The cytotrophoblasts between density makers of 1.049 and 1.062 g/mL were collected and cultured in DMEM containing 10% FBS (Gibco) and 1% antibiotics (Gibco) at 37°C in 5% CO2/95% air to allow spontaneous syncytialization in vitro. The process of syncytialization was observed with a microscope (Zeiss, Germany) following staining with hematoxylin–eosin.
Cell viability assay
A CCK8 Assay kit (WeiaoBio, Shanghai, China) was used to examine cell viability after AZM treatment. In brief, cells were seeded in a 96-well culture plate for 2 days to allow spontaneous syncytialization. Subsequently, the cells were treated with AZM (2.5, 5, and 10 µM; #S1835, Selleck, Houston, TX) for 24 h or AZM (2.5 µM) for 48 and 72 h. CCK-8 solution was then added to the cells for incubation (2 h) at 37°C in darkness. The optical density at 450 nm was measured with a microplate reader (Infinite® 200 PRO, TECAN, Switzerland). Wells without cells were taken as blank controls.
Transcriptome sequencing of human placental syncytiotrophoblasts with or without AZM treatment
Total RNA was extracted from syncytiotrophoblasts treated with or without AZM (10 μΜ; 24 h) (n = 3 each) using TRIzol reagent (#15596026, Thermo Fisher Scientific, Carlsbad, CA). RNA purity and integrity were determined using a NanoDropND-2000 (Thermo Fisher Scientific) and an Agilent 2200 TapeStation. Extracted RNA meeting the parameters (A260/A280 ratio ≥1.8, A260/A230 ratio ≥2.0, and RNA integrity number value ≥7.0) was further processed. RNA-sequencing libraries were constructed on an Illumina NovaSeq 6000 system using a TruSeq RNA sample preparation kit (Illumina Inc. San Diego, CA) according to a protocol from the manufacturer, and a computational pipeline was used to process the RNA-sequence data. After removing the adaptor and low-quality reads, the clean reads were aligned to the human genome (NCBI; GRCh38) using Hisat v.2.1.0 with default options (Kim et al., 2015). The fragment per kilobase per million mapped reads was used to indicate the abundance of transcripts. Differential expression was analyzed using the DESeq2 algorithm (Anders and Huber, 2010). Adjusted p value <0.05 and fold change ≥2 or ≤0.5 were set as the threshold for significantly differential expression. GO and KEGG pathway enrichment analyses (Kanehisa et al., 2008) of differentially expressed genes were performed using R based on the hypergeometric distribution. The transcriptome sequencing data are available in the Gene Expression Omnibus (GEO) data repository with the accessing number GSE249788.
Treatment of human placental syncytiotrophoblasts
To examine whether AZM affects syncytialization, the cytotrophoblasts were treated with AZM (2.5, 5, and 10 µM; 24 h) 3 h after plating. To examine the effects of AZM and TG, on ER stress, C/EBPα, and enzymes related to steroid hormone synthesis and IGFBP cleavage in syncytiotrophoblast, the syncytialized trophoblasts were treated with AZM (2.5, 5, and 10 µM; 24 h), AZM (2.5 µM; 48 h), or TG (0.01 and 0.1 μM; 24 h, #HY-13433, MCEchem, Princeton, NJ) two days after plating in the DMEM free of phenol red and FBS. To determine whether the PERK pathway was involved in the suppression of C/EBPα and enzymes related to steroid hormone synthesis and IGFBP cleavage by AZM, the syncytiotrophoblasts were treated with AZM (5 μM) in the presence and absence of PERK inhibitor (GSK2656157, 2.5 μM; #S7033, Selleck) or p-eIF2α inhibitor (ISRIB, 0.1 μM; #S7400, Selleck) or IRE1α RNase inhibitor (4μ8c, 2.5 μM; #S7272, Selleck) or siRNA-mediated knockdown of ATF4, for 24 h. The method of siRNA transfection is described below.
To measure estrogen secretion, the syncytiotrophoblasts were treated with AZM for 24 h, and the culture medium was then changed to a fresh medium containing dehydroepiandrosterone sulfate (, 2 μM; #1099872; Abotchem, Shanghai, China), the substrate of aromatase, for further incubation for 24 h. The medium from treated syncytiotrophoblasts was collected for estradiol measurement. To assess progesterone secretion, the syncytiotrophoblasts were treated with AZM for 24 h, and the culture medium was then collected directly for progesterone measurement. Both estradiol and progesterone levels were measured with chemiluminescence assay kits (Beckman Coulter, Pasadena, CA) following protocols provided by the manufacturer.
Transfection of small interfering RNA and overexpression vectors in human placental trophoblasts
The role of ATF4 and C/EBPα in the mediation of AZM-induced inhibition of CYP19A1, CYP11A1, ADAM12, and PAPPA was studied by transfecting cytotrophoblasts with 50 nM siRNA (Gene Pharma Co., Ltd., Shanghai, China) against ATF4 (5′-GUGAGAAACUGGAUAAGAATT-3′) and CEBPA (5′-GCUGACCAGUGACAAUGACTT-3′) using Lipofectamine RNAiMAX reagent (Invitrogen, San Diego, CA) or by transfecting cytotrophoblasts with the GV657 plasmid encoding C/EBPα using Lipofectamine 3000 reagent (Invitrogen) immediately after isolation. Randomly scrambled siRNA (5′-UUCUCCGAACGUGUCACGUTT-3′) and empty GV657 vector served as negative controls. After transfection for 48 h, syncytiotrophoblasts were incubated in DMEM for 24 h in the presence or absence of 5 μM AZM. The average knockdown efficiency of ATF4 was 74% and 90% at mRNA and protein levels, respectively (Supplementary Fig. S8A). The average knockdown efficiency of C/EBPα was 85% and 74% at mRNA and protein levels, respectively (Supplementary Fig. S9A and B). The average overexpression efficiency of C/EBPα was 1400% at the protein level (Supplementary Fig. S9C).
RNA extraction from human placental syncytiotrophoblasts and analysis with qRT-PCR
Total RNA was extracted from syncytiotrophoblasts using an RNA isolation kit (Foregene, Chengdu, China). After the determination of RNA purity and concentration with NanoDrop ND-2000 (Thermo Fisher Scientific), mRNA was reversely transcribed to complementary DNA (cDNA) using a Prime Script RT Master Mix Perfect Real-Time Kit (Takara, Kyoto, Japan). The mRNA abundance related to mRNA related to syncytialization (ERVW1, ERVFRD1, CDH1), the mRNA related to ER stress (HSPA5, ATF4, XBP1(S), HERPUD1, DDIT3, and GADD34), the mRNA related to steroid hormone synthesis, and IGFBP cleavage (CYP19A1, CYP11A1, STS, HSD3B1, ADAM12, PAPPA, and PAPPA2), as well as CEBPA, was measured with qRT-PCR using the transcribed cDNA and Power SYBR Premix Ex TaqTM (Takara) following a previously described protocol (Wang et al., 2015). The primers used for qRT-PCR are listed in Supplementary Table S1.
Protein extraction from human placental syncytiotrophoblasts and analysis with western blotting
Total cellular protein was extracted from syncytiotrophoblasts using the radioimmunoprecipitation assay lysis buffer (Active Motif, Carlsbad, CA) containing inhibitors for proteases (Roche, Basel, Switzerland) and phosphatases (Roche). The total secreted protein in the culture medium was extracted and condensed with a centrifugal filter device (#UFC501096, Millipore, Billerica, MA) following the instructions provided by the manufacturer. After measurement of the protein concentration using a BCA Protein Assay kit (ZJ101, Epizyme Biotech, Shanghai, China), the abundance of aromatase, P450scc, 3β-HSD1, STS, ADAM12-S, PAPP-A, PAPP-A2, eIF2α, p-eIF2α at Ser51, IRE1α, p-IRE1α at Ser724, ATF4, XBP1s, CHOP, ATF6, and C/EBPα was determined with Western blotting following a standard protocol. In brief, 20–30 μg of protein was electrophoresed in a sodium dodecyl sulfate (SDS)–polyacrylamide gel and then transferred onto a nitrocellulose membrane (Millipore). After blocking with 5% nonfat milk, the membrane was probed with primary antibodies against aromatase (1:1000, #ab35604, Abcam, Cambridge, UK), P450scc (1:1000, #ab175408, Abcam), 3β-HSD1 (1:1000, #sc-100466, Santa Cruz, Santa Cruz, CA), STS (1:2000, #ab233233, Abcam), ADAM12 (1:1000, #ab223476, Abcam), PAPP-A (1:500, #sc-365226, Santa Cruz), PAPP-A2 (1:1000, #AF1668, R&D Systems, Minneapolis, MN), p-eIF2α at Ser51 (1:1000, #9721, Cell Signaling, Danvers, MA), p-IRE1α at Ser724 (1:1000, #PTM-20054, PTMab, Hangzhou, China), ATF4 (1:1000, #11815, Cell Signaling), XBP1s (1:1000, #24868-1-AP, Proteintech, Wuhan, China), CHOP (1:500, #2895, Cell Signaling), ATF6 (1:1000, #24169-1-AP, Proteintech), eIF2α (1:1000, #A0764, ABclonal, Wuhan, China), IRE1 (1:1000, #A17940, ABclonal), and C/EBPα (1:1000, #8178, Cell Signaling), respectively, overnight at 4°C. After washing with 1×Tween 20/Tris buffered salt solution, the membrane was incubated with the corresponding secondary antibody conjugated with horseradish peroxidase. The peroxidase activity was developed with a chemiluminescence detection system (Millipore), which was visualized using the digital imaging equipment eBlot Touch Imager™ (eBlot Tec., Shanghai, China). The internal loading control was performed by probing the same blot with the antibody against Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (1:20000, #60004-1, Proteintech). The ratio of the band intensity of the target protein to that of GAPDH was used to indicate the abundance of the target protein in the cell. The ratio of the band intensity of ADAM12-S or PAPP-A to the strongest band stained with Ponceau S was used to indicate the abundance of the secreted target protein in the culture medium. The ratio of phosphorylated protein band density over that of total protein was used to indicate the abundance of phosphorylated protein.
TUNEL assay
The TUNEL assay was performed to detect apoptotic cells upon treatment with AZM (0, 2.5, 5,10 µM; 24 h). Briefly, after treatment with AZM, the cells were washed with PBS and then fixed with 4% paraformaldehyde at room temperature. After permeabilization with 0.3% Triton X-100, cells were subjected to a TUNEL Detection kit (Vazyme Biotech; #A111). After incubating in a humidified chamber at 37°C for 1 hour and washing with PBS, images were captured using a fluorescence microscope (Zeiss, Germany). Treatment of cells with Dnase (2U/mL) served as a positive control.
Transmission electron microscopic examination
To observe the endoplasmic reticulum morphology, the human placental syncytiotrophoblasts with or without AZM (5 μM; 24 h) treatment and mice placenta were processed for observation with a transmission electron microscope. After rapid fixation with 2.5% glutaraldehyde at 4°C for 2 h and postfixation with 1% osmium tetroxide solution for 2 h at 4°C, the cells were processed for dehydration, infiltration, and routine embedding in resin. After solidification, ultrathin sections (70 nm) were cut and stained with lead citrate, which was then observed for ER morphology with a transmission electron microscope (FEI Talos L 120C; Thermo Fisher Scientific).
ChIP assay
The enrichment of C/EBPα at CEBPA, CYP19A1, CYP11A1, PAPPA, and ADAM12 promoters following treatment with or without AZM (5 μM; 24 h) was determined with ChIP assay as described previously (Zhu et al., 2023). Briefly, after crosslinking with 1% formaldehyde, syncytiotrophoblasts were lysed with the SDS lysis buffer containing a protease inhibitor cocktail. The lysed cells were sonicated to shear the chromatin DNA to an optimal size of around 500 bp, which was then immunoprecipitated with the antibody against C/EBPα (Cell Signaling; #8178). An equal amount of preimmune IgG served as negative control. Sheared DNA without immunoprecipitation served as input control. The immunoprecipitate was pulled down with Magna ChIP Protein A + G agarose Magnetic Beads (Millipore) on a magnetic stand (Millipore). After reverse crosslinking, protein and RNA were removed by digestion with proteinase K and ribonuclease, respectively. The sheared DNA was extracted using a DNA Purification kit (Cwbiotech, Beijing, China) for subsequent qRT-PCR with primers aligning the putative C/EBPα binding sites in CEBPA, CYP19A1, CYP11A1, PAPPA, and ADAM12 promoters (Supplementary Fig. S10). The primer sequences for the ChIP assay are illustrated in Supplementary Table S2. The ratio of DNA precipitated by C/EBPα antibody over that of input control was used to indicate the abundance of bound C/EBPα.
CoIP assay
CoIP assay was carried out to examine whether ATF4 and C/EBPα could conjugate after AZM treatment. Nuclear protein was extracted from syncytiotrophoblasts treated with or without AZM (5 µM; 24 h) using a Nuclear Extract Kit (Active Motif). After the determination of protein concentration, 10 µg of nuclear protein was incubated with 1:50 dilution of the antibody against C/EBPα (Cell Signaling; #8178) or preimmune IgG as negative control overnight at 4°C. Then, protein A agarose beads were added for incubation with the above reaction mixture for 2 h to pull down the antibody/antigen complex. The precipitated antibody/antigen/agarose complex was washed adequately and then denatured in Western blotting loading buffer at 95°C for subsequent detection of ATF4 with Western blotting following the same standard protocol as Western blotting.
Animal study
The animal experimentation was approved by the Institutional Review Board of Ren Ji Hospital, Shanghai Jiao Tong University School of Medicine, and followed the ARRIVE guidelines for animal care. C57BL/6 mice aged between 10 and 13 weeks (Charles River, Beijing, China) were mated overnight. The presence of a vaginal plug the next morning was counted as GD 0.5. Pregnant mice were then transferred to individual cages and divided randomly into the control, AZM, ISRIB, and AZM plus ISRIB groups (n = 9 per group). The concentrations of AZM and ISRIB used for gavage were 20 mg/mL and 1 mg/mL, respectively, both of which were dissolved in 0.5% carboxymethyl cellulose sodium (CMC-Na). AZM (200 mg/kg body weight) and ISRIB (10 mg/kg body weight) were administered by gavage once a day for three days on GD15.5, 16.5, and 17.5. The control group received an equivalent volume of 0.5% CMC-Na. Maternal blood was collected from the orbital sinus after anesthesia with 65 mg/kg Zoletil 50 (Virbac, Nice, France), and subsequently, the dam was euthanized by cervical dislocation on GD 18.5. The fetus and placenta were promptly removed and weighed. After weighing, a portion of the placenta was snap-frozen in liquid nitrogen and stored at −80°C for subsequent protein extraction after homogenization with an electric homogenizer (Biheng Bio., Shanghai, China). Another portion of the placenta was fixed in 2% glutaraldehyde, and the labyrinth layer was dissected under a stereoscopic microscope for processing and observation with a transmission electron microscope. The abundance of placental p-eIF2α and ATF4 was quantified with Western blotting with p-eIF2α at Ser51 (1:1000, #9721, Cell Signaling) and ATF4 (1:1000, #11815, Cell Signaling) antibodies. The internal loading control was examined by probing the same blot with a GAPDH antibody (1:10000, #60004-1, Proteintech). ELISA kits were used to measure the abundance of ADAM12-S and PAPP-A in the placenta and estradiol and progesterone in the maternal blood following instructions from the manufacturers. The information of the ELISA kits was as follows: ADAM12 (#MBS177214, MyBioSource, San Diego, CA), PAPP-A (#RK03099, ABclonal, Wuhan, China), estradiol (#DEV9999, Demeditec Diagnostics GmbH, Kiel, Germany), and progesterone (#DEV9988, Demeditec Diagnostics GmbH).
Statistical analysis
All data are presented as mean (standard deviation [SD]). The n number in each experiment represents independent experiments using samples from individual pregnant women or mice. After assessment of the data normality with the Shapiro–Wilk test, a comparison of two groups was conducted using paired or unpaired Student’s t-test or Mann–Whitney U test where appropriate. Multiple group comparisons were conducted by one-way analysis of variance followed by Tukey’s post hoc tests or Kruskal–Wallis test followed by Dunn’s post hoc test where appropriate. Statistical significance was defined as p < 0.05.
Use of electronic laboratory notebook
Electronic laboratory notebook was not used.
Footnotes
Acknowledgments
The authors are grateful to Wen-jia Lei, Lu-yao Wang, Li-jun Ling, and Hao Ying for assistance with the placenta collection.
Data Availability
The raw data of RNA sequencing are available from the NCBI GEO accession GSE249788. The original data and materials presented in the study are available from the corresponding authors upon reasonable request.
Authors’ Contributions
F.P., F.Z., and M.-D.L.: investigation and methodology; F.P., W.-S.W., and K. S.: writing. K.S. and W.-S.W.: conceptualization, supervision, and funding acquisition.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by the
Supplementary Material
Dataset S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Table S1
Supplementary Table S2