
Abstract
Aim:
Spinal cord injury (SCI) is a catastrophic injury characterized by oxidative stress. Glutathione peroxidase 3 (GPx3) is an antioxidant enzyme that protects against immune responses in various diseases. However, the effects of GPx3 in SCI remains unclear. This study aimed to investigate the role of GPx3 in SCI and its underlying mechanisms.
Results:
We injected adeno-associated viruses to overexpress GPx3 in mice. Primary microglia and BV2 cells were used as in vitro models. We knocked down or overexpressed GPx3 in BV2 cells. Additionally, BV2 cells transfected with siIRAK4 were used to perform rescue experiments. A series of histological and molecular biological analyses were used to explore the role of GPx3 in SCI. Overexpression of GPx3 inhibited oxidative stress in mice, improving functional recovery after SCI. Similarly, LPS+ATP stimulation decreased GPx3 expression in microglia. Silencing of GPx3 elevated the generation of reactive oxygen species, increased the expression of IRAK4 and pro-inflammatory factors, and promoted pyroptosis in microglia. However, overexpression of GPx3 reversed these results. Moreover, silencing of IRAK4 alleviated these phenomena, which were upregulated by GPx3 deficiency.
Innovation and Conclusion:
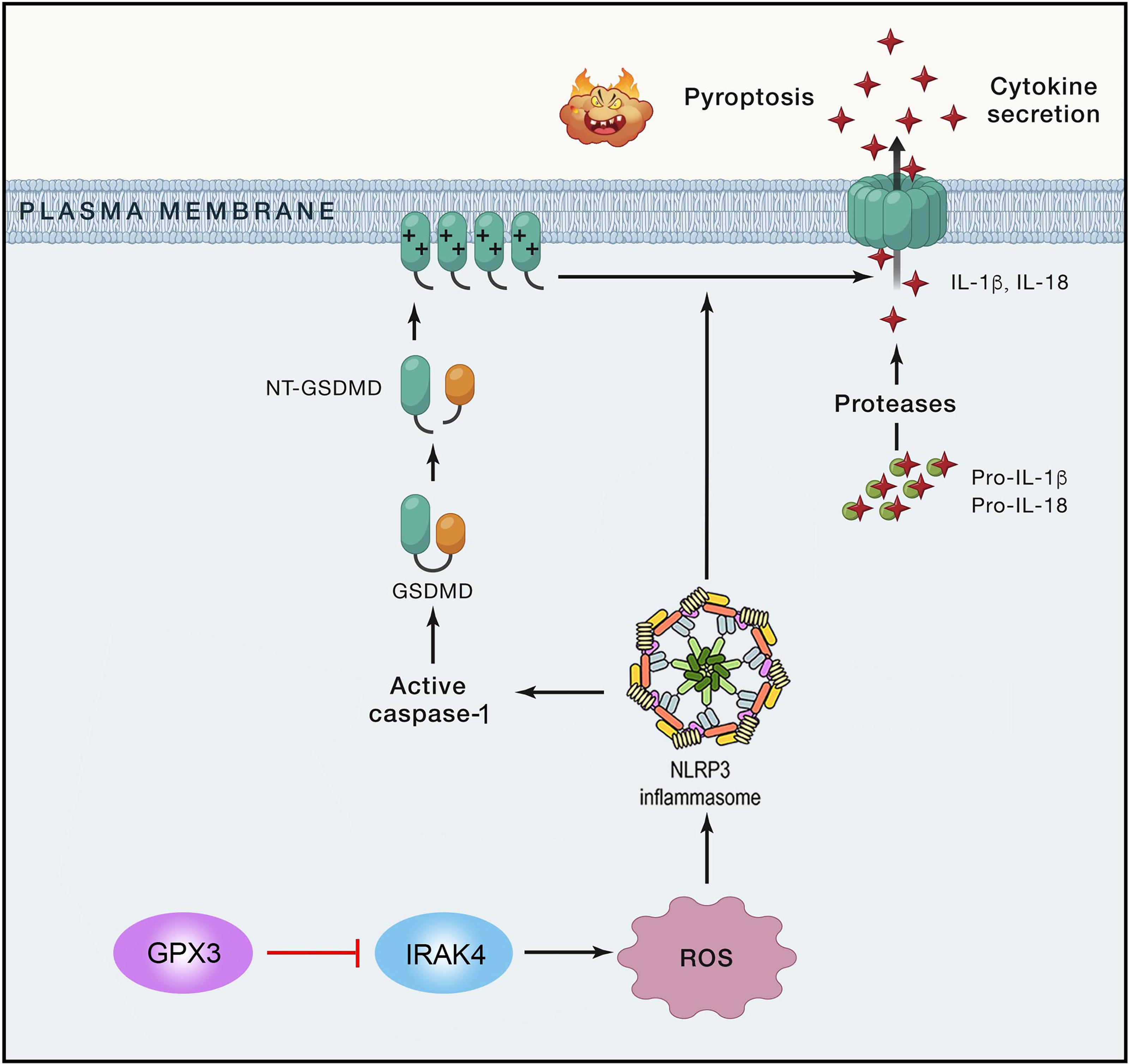
Our results demonstrated that GPx3 plays a critical role in SCI by inhibiting microglial pyroptosis via the IRAK4/ROS/NLRP3 signaling pathway. Antioxid. Redox Signal. 42, 711–729.
Introduction
Spinal cord injury (SCI) is a devastating traumatic injury in the central nervous system (CNS) that leads to loss of sensory and motor functions (Liu et al., 2021a). Approximately 10.4–83.0 new cases of SCI are reported annually per million people (Chang et al., 2020). SCI can be classified as primary and secondary injuries. Primary injury is physical damage to the spine that cannot be repaired, whereas secondary injury is manageable and includes an inflammatory response, immune response, and oxidative stress (Wang et al., 2021).
Neuroinflammation is recognized as a pivotal mechanism in secondary injury (Liu et al., 2021c). Secondary injury is characterized by increased oxidative stress and reactive oxygen species (ROS) production. SCI results in increased oxidative stress and ROS production, which in turn triggers an inflammatory response. Microglia are resident macrophages that participate in active innate immune defenses in the CNS. After SCI, neuroinflammation is primarily mediated by microglia, which exacerbate the inflammatory response by producing excessive ROS (Liao et al., 2020). Given that neuroinflammation is a hallmark of secondary injury, alleviation of microglia-mediated inflammation may be a feasible therapeutic option for SCI.
The IL-1R-associated kinase (IRAK) family is involved in TLR signaling and consists of four functionally and structurally related members, IRAK1–4. IRAK4 is a critical target that is involved in the innate immune response process (Su et al., 2020). IL-1β binding recruits adapter proteins, which in turn recruit IRAK family members by directly interacting with IRAK4. IRAK4 autophosphorylates and activates the downstream kinases IRAK1 and IRAK2. Studies have reported that IRAK4 binds to upstream and downstream signal transduction effectors, initiates signal transduction cascades, and participates in inflammatory signaling pathways (Suzuki et al., 2002; Zarrin et al., 2021).
Innovation
The main work of this study explores the role of Glutathione peroxidase 3 (GPx3) in spinal cord injury (SCI) and its underlying mechanisms. Our results demonstrate that that GPx3 significantly reduced microglia-mediated oxidative stress and neuroinflammation following SCI. The specific mechanisms may be closely related to the activation of the IRAK4/ROS/NLRP3 signaling pathway and the inhibition of pyroptosis.
Pyroptosis, a recently recognized inflammatory form of programmed cell death, induces a robust inflammatory response and releases abundant pro-inflammatory mediators (Shi et al., 2015). Our previous study showed that oxidative stress and neuroinflammation in SCI induced pyroptosis (Liu et al., 2020). Numerous studies have shown that inflammasomes play a pivotal role in pyroptosis. The term inflammasome was first used in 2002 by Martinon to describe a novel macromolecular complex that can cause inflammation (Martinon et al., 2002). The precursor caspase-1 is cleaved into caspase-1 upon inflammasome activation, which further facilitates the maturation of the pro-inflammatory cytokines IL-1β and IL-18. Of the four types, the NLRP3 inflammasome has been extensively studied in CNS diseases. They are abundantly present in microglia and are essential for initiating the innate immune system and inflammatory response (Ising et al., 2019). Numerous studies indicate that lysosomal rupture, mitochondrial damage, ROS production, and ion channel gating might be functions of the NLRP3 inflammasome (Yu et al., 2017; Zhou et al., 2011; Hofbauer et al., 2021). Of these, ROS has been considered a classical mechanism of NLRP3 inflammasome activation.
The glutathione peroxidase (GPx) family comprises important peroxide decomposition enzymes that are widely distributed in mammals and constitute the first line of defense against ROS production (Martins et al., 2020). Eight distinct isozymes belonging to the GPx family (GPx1–8) catalyze the reduction of hydrogen peroxide, hydroperoxides, and lipid hydroperoxides. Although they are phylogenetically similar, they differ in their antigenicity, structure, and subcellular localization (Jelinek et al., 2018; Zhang et al., 2020). Of these, GPx3 can eliminate all complex hydroperoxides and plays an important role in antioxidant defense by scavenging free radicals. The human GPx3 gene, which comprises five exons spanning 10 kb in the 5q32 region of chromosome 5, encodes a 23 kDa protein that assembles into a homotetramer. Each monomer contains a conserved tetrad composed of glutamine, tryptophan, asparagine, and selenocysteine. Selenocysteine is present in the enzyme’s active site. GPx3 can prevent immune system dysfunction and its absence results in oxidative damage and immunosuppression (Avissar et al., 1994; Olson et al., 2010). Recent studies have suggested that aberrant GPx3 expression is closely related to the development of several neurological diseases such as ischemia and Alzheimer’s disease (de Carvalho et al., 2020; Zimetti et al., 2021). In the above studies, GPx3 was considered a disease biomarker; however, its role in disease pathology and specific underlying mechanisms remain elusive. To date, no study has demonstrated the roles of GPx3 in the pathophysiology of SCI and its underlying mechanisms.
This study aimed to investigate the role of GPx3 in SCI and its related mechanisms both in vivo and in vitro. Thus, our study confirmed the potential therapeutic role of GPx3 in SCI. To the best of our knowledge, this is the first study to demonstrate that GPx3 attenuates ROS-dependent NLRP3 inflammasome activation and inhibits microglial pyroptosis, thereby reducing neuroinflammation following SCI.
Results
GPx3 levels are decreased in SCI mice
To identify GPx3′s role in the pathogenesis of SCI, we examined the expression levels of GPx3 in mice with SCI at different time points. Immunofluorescence and western blotting revealed that the expression of GPx3 in the spinal cord tissue decreased after SCI at each time point (Fig. 1A, B). The most significant decrease was found at one-week post-injury. Hence, the time point of one week after SCI was selected for subsequent experiments. Enzyme-linked immunosorbent assay (ELISA) results suggested that GPx3 expression decreased significantly in the SCI group compared with that in the sham group (Fig. 1C, D). The ELISA results for GPx3 levels in primary microglia were consistent with those observed in vivo (Fig. 1E). These results supported that GPx3 plays a crucial role in SCI.

Overexpression of GPx3 promotes functional recovery and alleviates structural disorder after SCI
Motor function recovery is a critical index for assessing the therapeutic efficacy. Thus, in the present study, the functional recovery of the forelimbs was assessed using rearing tests and FLS in the present study.
In the rearing test, the preoperative rearing usage rate was approximately 50%. The usage rate of the ipsilateral forelimb in the SCI+Adeno-associated virus (AAV)-GPx3 group was higher than that in the SCI+AAV-NC group at all time points post injury. The utilization rates of the ipsilateral fore limb in SCI+AAV-GPx3 group were significantly higher than in SCI+AAV-NC group at 10 weeks (34.454 ± 6.018 vs. 25.87 ± 6.51, respectively) and 12 weeks (35.747 ± 5.174 vs. 26.393 ± 5.086, respectively) (Fig. 2A).
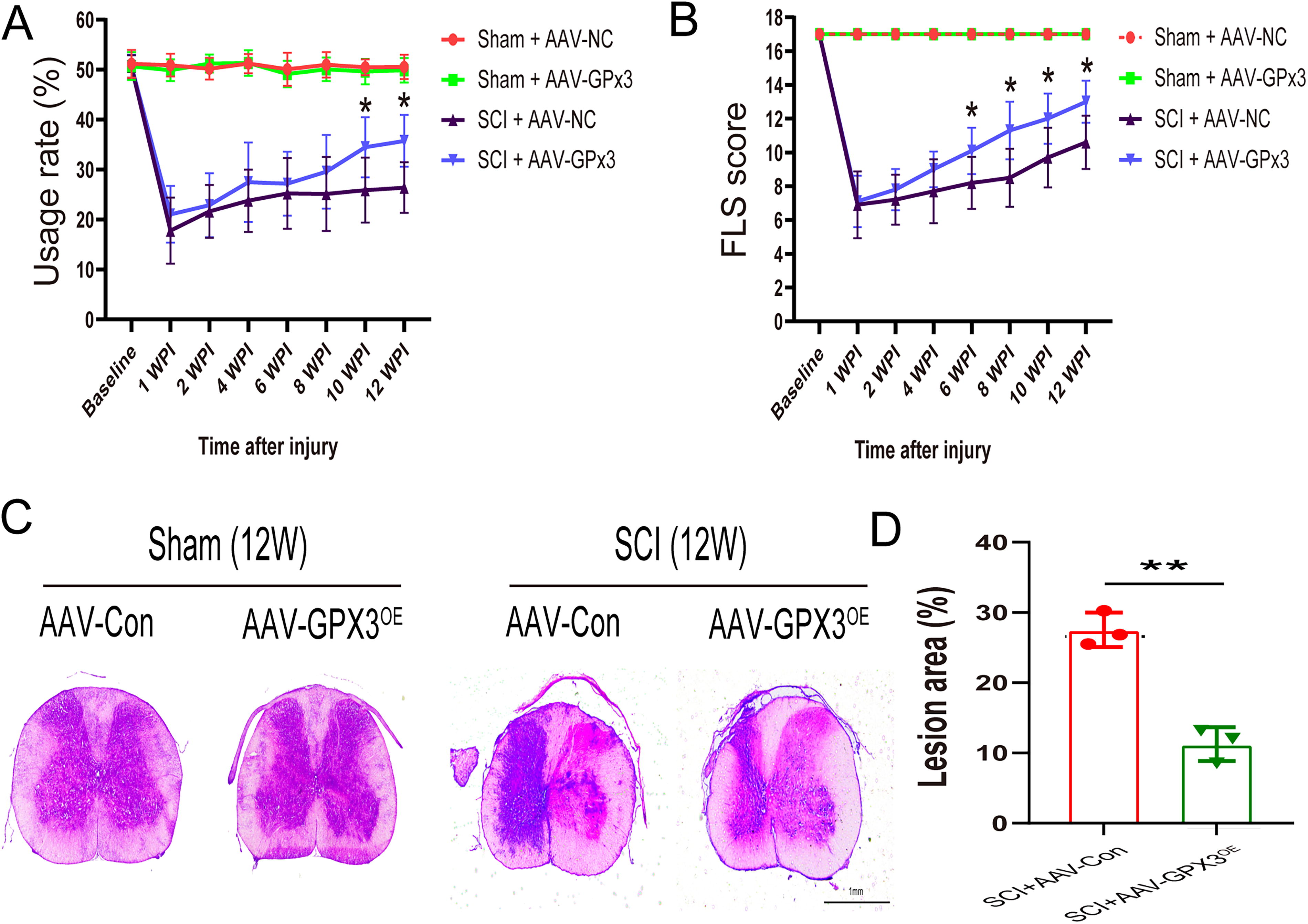
The mean baseline FLS was 17. Similar to the rearing test, the SCI+AAV-GPx3 group obtained higher scores than the SCI+AAV-NC group at all time points post-injury (6 weeks: 10.1 ± 1.370 vs. 8.2 ± 1.549, respectively; 8 weeks: 11.3 ± 1.702 vs. 8.5 ± 1.716, respectively; 10 weeks: 12 ± 1.491 vs. 9.7 ± 1.767, respectively; and 12 weeks: 13 ± 1.247 vs. 10.6 ± 1.578, respectively) (Fig. 2B).
To assess histological changes, hematoxylin and eosin (H&E) staining was conducted to compare the lesion area between the SCI+AAV-GPx3 and SCI+AAV-NC groups. As shown in Figure 2C–D, the lesion area decreased after AAV-GPx3 treatment at 12 weeks post-injury (from 27.541 ± 2.442 to 11.265 ± 2.354). These results demonstrate that AAV+GPx3 treatment resulted in better recovery of forelimb motor function and structural disorders.
Overexpression of GPx3 inhibits microglia activation and oxidative stress after SCI
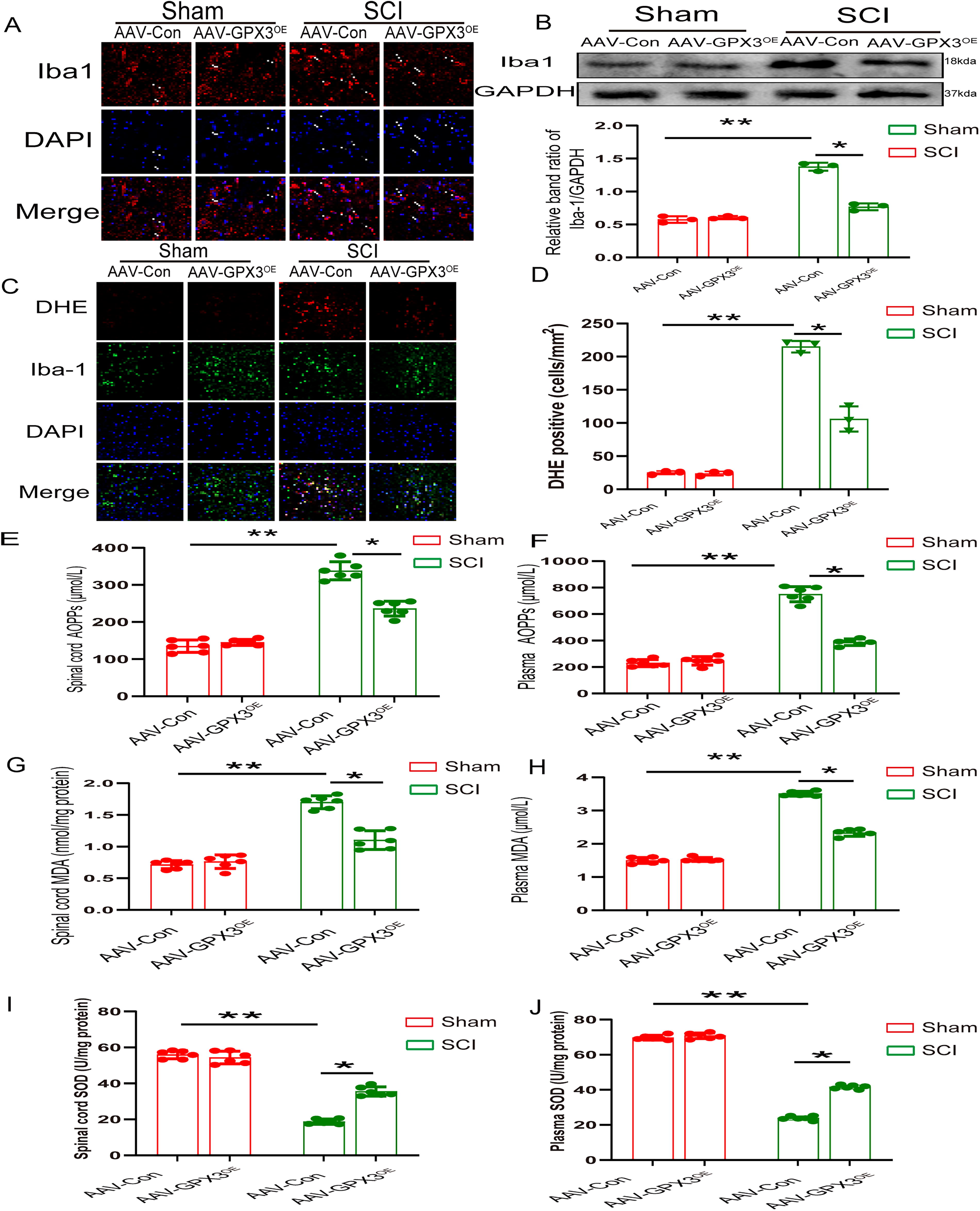
SCI is characterized by microglial activation (Li et al., 2020). Immunofluorescence and western blot analyses revealed that the number of activated microglia in the SCI+AAV-GPx3 group was significantly lower than that in the SCI+AAV-NC group (Fig. 3A, B). Oxidative stress is a crucial pathogenic factor in SCI. ROS are regarded as the major contributors to oxidative stress. Therefore, we measured ROS levels using dihydroethidium (DHE) staining. Immunofluorescence analyses revealed fewer DHE-positive cells in the SCI+AAV-GPx3 group than in the SCI+AAV-NC group (106 ± 19.079 vs. 215 ± 8.718) (Fig. 3C and D). AOPPs, measurement of malondiadehyde (MDA), and superoxide dismutase (SOD) are important indicators of oxidative stress. AOPPs levels significantly decreased in the spinal cord tissue and plasma of the SCI+AAV-GPx3 group than those in the SCI+AAV-NC group (tissue: 338.326 ± 36.362 vs. 236.525 ± 28.99, respectively; plasma: 749.868 ± 79.957 vs. 387.406 ± 36.173, respectively) (Fig. 3E and F). A similar pattern was observed for the MDA levels. Compared with SCI+AAV-NC group, the MDA levels in the spinal cord tissue and plasma of the SCI+AAV-GPx3 group were significantly decreased (tissue: 1.715 ± 0.14 vs. 1.115 ± 0.159, respectively; plasma: 3.486 ± 0.069 vs. 2.324 ± 0.139, respectively) (Fig. 3G and H). In contrast, the SOD levels significantly increased in the spinal cord tissue and plasma of the SCI+AAV-GPx3 group than those of the SCI+AAV-NC group (tissue: 18.864 ± 1.613 vs. 35.604 ± 1.889, respectively; plasma: 23.582 ± 1.616 vs. 42.13 ± 1.223, respectively) (Fig. 3I and J). These data implied that AAV-GPx3 treatment attenuated oxidative stress after SCI.
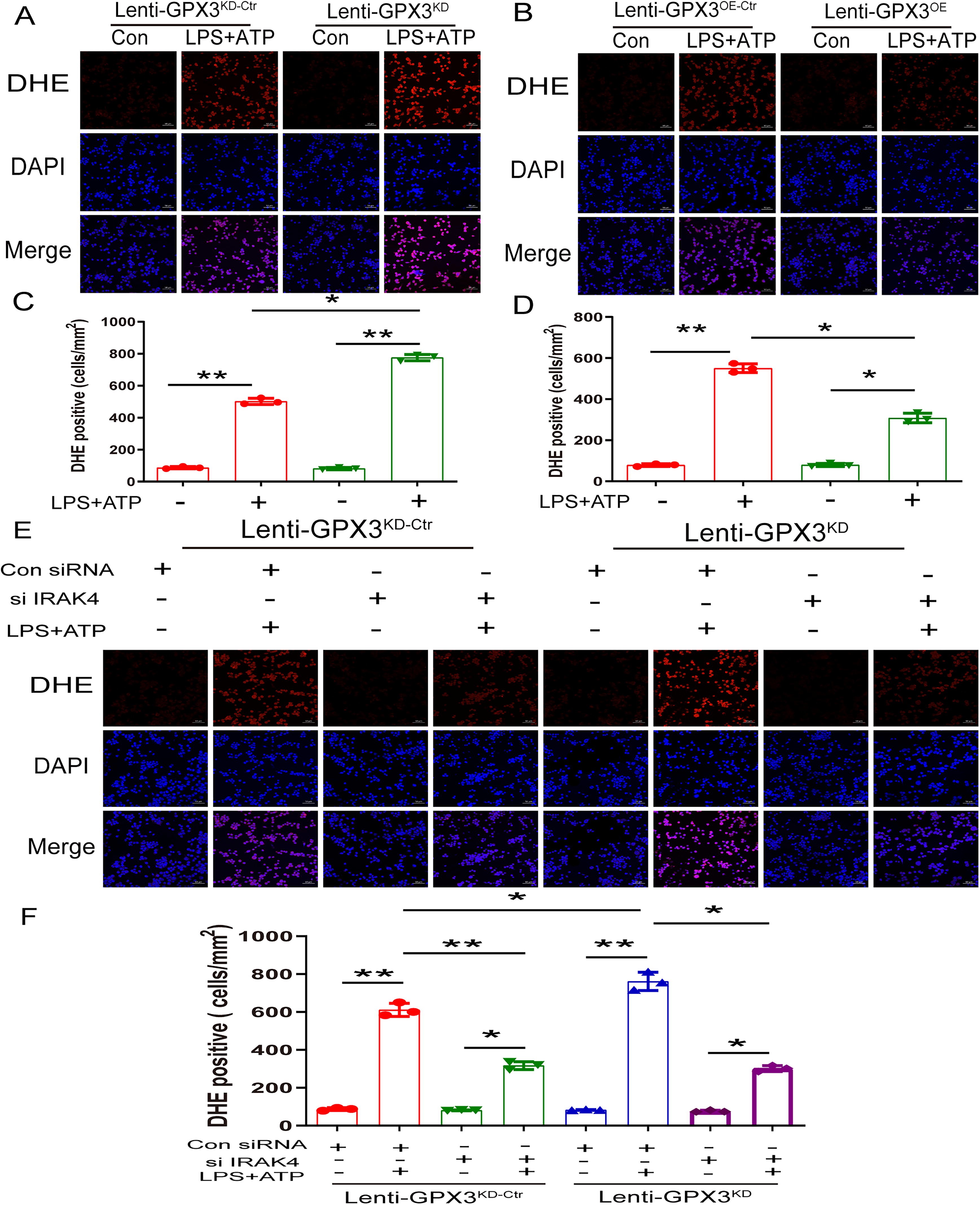
GPx3 reduce intracellular ROS through IRAK4 in BV2 cells
To verify the role of GPx3 in ROS generation in BV2 cells, we silenced or overexpressed GPx3 using lentiviruses. DHE staining results showed that silencing of GPx3 amplified ROS generation (from 502.333 ± 19.296 to 776 ± 19.698), whereas GPx3 overexpression reduced ROS generation (from 551 ± 21.656 to 308.333 ± 23.159) (Fig. 4A–D). To detect whether GPx3 deficiency increases ROS generation through IRAK4, scrambled and shBAP31 BV2 cells were transfected with IRAK4 siRNA. As shown in Figure 4E and F, IRAK4 silencing reduced ROS production. Overall, these results show that GPx3 reduced ROS production via IRAK4 in BV2 cells.
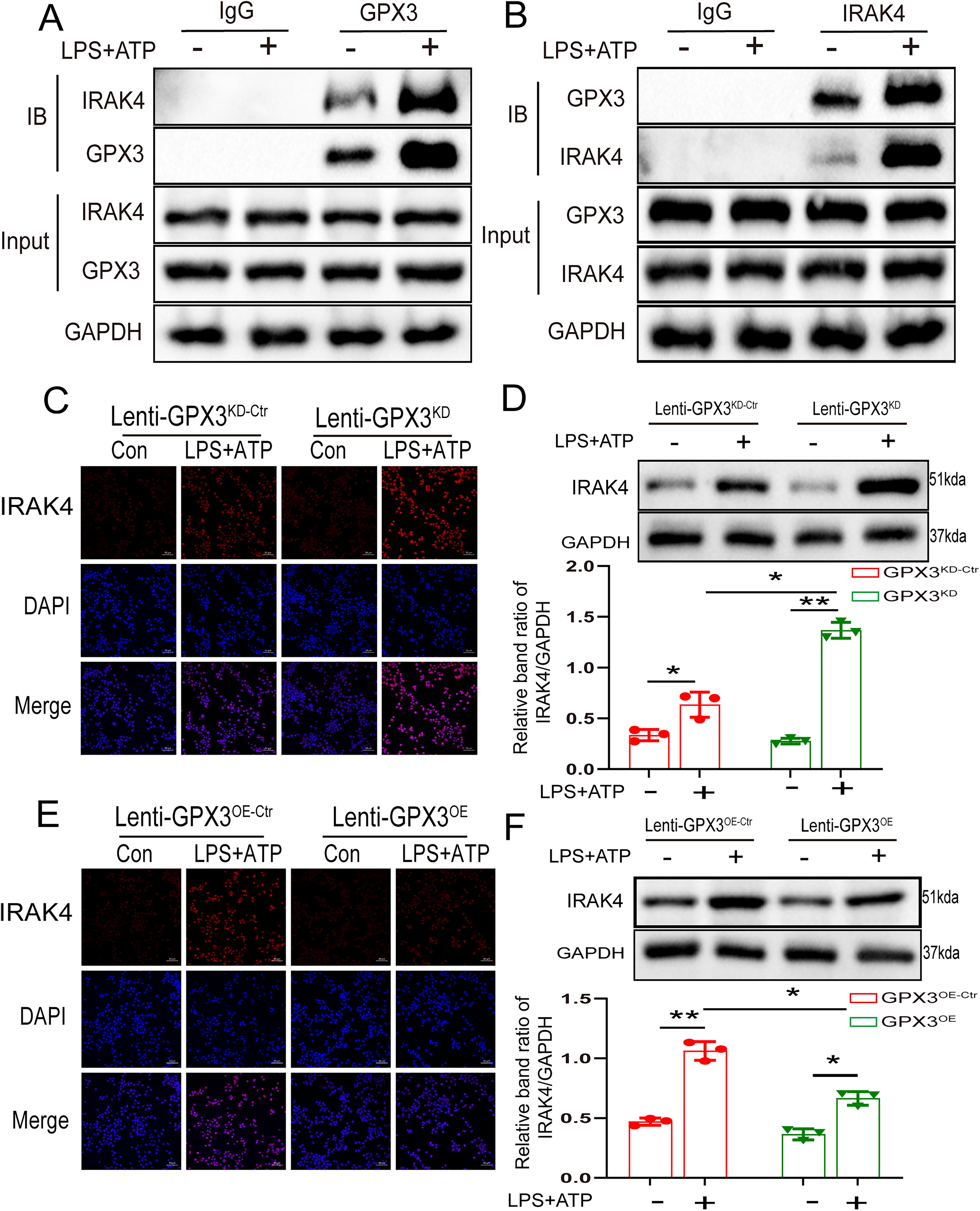
GPx3 binds to and reduces IRAK4 expression
The interaction between GPx3 and IRAK4 was verified using a coimmunoprecipitation experiment during LPS + ATP stimulation. The results showed that the anti-GPx3 antibody and anti-IRAK4 antibodies pulled each other down (Fig. 5A and B). To study the mechanism underlying GPx3-dependent IRAK4 regulation, we performed immunofluorescence and western blot assays and found that the protein levels of IRAK4 increased markedly in GPx3 knockdown BV2 cells (Fig. 5C and D). Correspondingly, overexpression of GPx3 significantly decreased the levels of IRAK4 (Fig. 5E and F). Collectively, these results suggest that GPx3 nteracted with IRAK4 and inhibited its expression.
GPx3 inhibits microglial pyroptosis
IL-1 and IL-18 are released as a consequence of pyroptosis, an inflammatory form of programmed necrosis (Man et al., 2017). Immunofluorescence experiments were conducted to detect the expression of IL1β and IL18. The results revealed that the expression of IL1β increased markedly in GPx3 knockdown BV2 cells (Fig. 6A). Correspondingly, GPx3 overexpression significantly decreased the expression of IL1β (Fig. 6B). Furthermore, western blotting revealed that the protein levels of IL1β and IL18 increased markedly in GPx3 knockdown BV2 cells (Fig. 6C–E). In contrast, overexpression of GPx3 significantly decreased the levels of IL1β and IL18 (Fig. 6F–H). To further characterize microglial pyroptosis, we performed Propidium iodide (PI) staining assays and lactate dehydrogenase (LDH) release. GPx3 deficiency increased the number of PI-positive cells (from 59.659 ± 2.847% to 78.699 ± 4.061%) (Fig. 6I and K). In contrast, GPx3 overexpression decreased the number of PI-positive cells (from 64.908 ± 5.332% to 44.34 ± 4.669%) (Fig. 6J and L). Similar results were obtained in LDH release results. GPx3 deficiency significantly enhanced the release of LDH (from 59.127 ± 3.675% to 72.344 ± 5.54%) compared to that in scrambled BV2 cells (Fig. 6M). Overexpression of GPx3 significantly reduced c (from 69.021 ± 11.53% to 45.468 ± 6.643%) compared to that in scrambled BV2 cells (Fig. 6N). TEM results revealed that bubbles and large pores in the membrane were more frequent observed in GPx3 knockdown BV2 cells. While overexpression of GPx3 decreased the membrane bubbles and pores (Fig. 6O and P). These findings indicated that GPx3 inhibited pyroptosis in BV2 cells.

GPx3 inhibits the activation of NLRP3 inflammasome
The NLRP3 inflammasome comprises NLRP3, ASC, and pro-caspase-1. Once the NLRP3 inflammasome is activated, pro-caspase-1 is autocatalytically cleaved, resulting in the processing and release of the pro-inflammatory cytokines IL-1 and IL18 (Chou et al., 2021). When the NLRP3 inflammasome is activated, GSDMD is cleaved, yielding the N-terminus of GSDMD as a pyroptosis effector molecule. Immunofluorescence revealed that the expression of NLRP3 and GSDMD-NT increased in GPx3 knockdown BV2 cells (Fig. 7A and B). However, overexpression of GPx3 decreased the expression of NLRP3 and GSDMD-NT (Fig. 7C and D). Immunoblot experiments were used to further corroborate these results, and it was observed that the protein levels of the NLRP3 inflammasome, cleaved-caspase 1, and GSDMD-NT increased drastically in GPx3 knockdown BV2 cells (Fig. 7E–I). Overexpression of GPx3 markedly decreased the expression of the NLRP3 inflammasome, cleaved caspase 1, and GSDMD-NT (Fig. 7J–N). These results suggest that GPx3 inhibited microglial pyroptosis via the NLRP3 inflammasome.
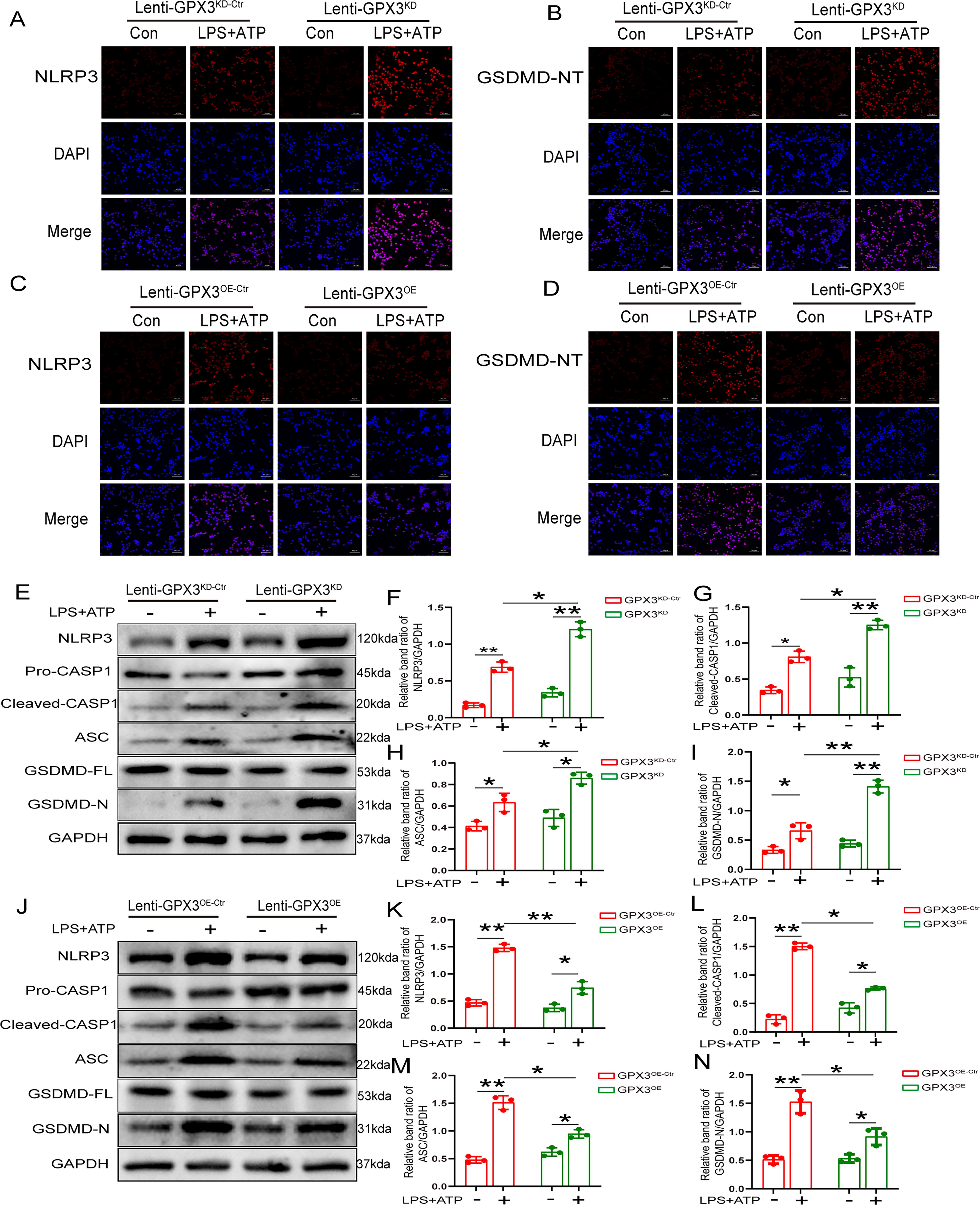
GPx3 inhibits pyroptosis via IRAK4/NLRP3 axis in BV2 cells
To identify the specific molecular mechanism underlying GPx3 inhibition in microglial pyroptosis, a series of rescue experiments were performed. We performed immunofluorescence staining and found that the expression of NLRP3, GSDMD-NT, and IL-1β was increased in GPx3 knockdown BV2 cells, whereas silencing IRAK4 inhibited the upregulation of NLRP3, GSDMD-NT, and IL-1β induced by GPx3 deficiency (Fig. 8A–C). Western blotting analysis revealed similar trends. The protein levels of NLRP3, GSDMD-NT, and IL-1β were dramatically increased in GPx3 knockdown BV2 cells, whereas silencing of IRAK4 inhibited the upregulation of NLRP3, GSDMD-NT, and IL-1β induced by GPx3 deficiency (Fig. 8D–G). We further investigated the role of pyroptosis using rescue experiments. To address this, PI staining and LDH release assays were performed. Their results agree with the abovementioned immunofluorescence and immunoblot results. PI-positive cells increased in GPx3 knockdown BV2 cells, whereas IRAK4 silencing decreased the number of PI-positive cells upregulated by GPx3 deficiency (Fig. 8H and I). Moreover, the release of LDH was markedly increased in GPx3 knockdown BV2 cells, whereas silencing IRAK4 inhibited the increase in LDH release induced by GPx3 deficiency (Fig. 8J). These results confirmed that GPx3 inhibits microglial pyroptosis via the IRAK4/NLRP3 signaling pathway.
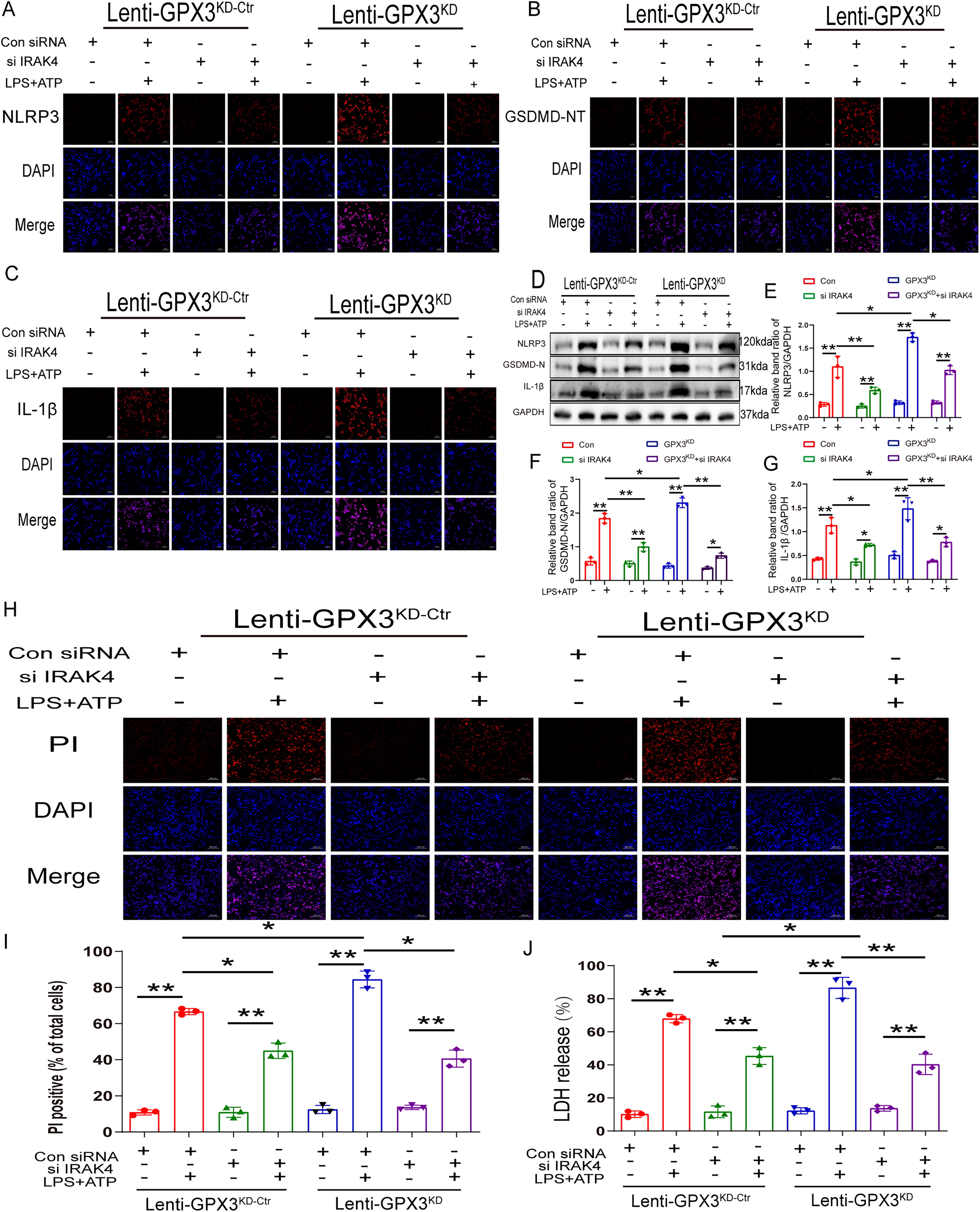
Overexpression of GPx3 inhibits IRAK4 and NLRP3 inflammasome-dependent pyroptosis after SCI
We further attempted to determine how GPx3 inhibits neuroinflammation and pyroptosis following SCI. Immunostaining revealed that overexpression of GPx3 decreased the fluorescence intensity of IRAK4, NLRP3, caspase p10, and IL-1β after SCI (Fig. 9A–D). Similarly, western blotting showed that GPx3 overexpression considerably decreased the protein levels of IRAK4, NLRP3, caspase p10, GSDMD-NT, IL18, and IL-1β after SCI (Fig. 9E–L). Pyroptosis is distinguished based on the formation of membrane pores. To observe the changes in the cell membrane post SCI, a transmission electron microscope was used to observe the ultrastructure of the microglia. Bubbles and large pores were observed in the plasma membrane of the microglia in the SCI+AAV-NC group, whereas the sham+AAV-NC group exhibited a linear and intact membrane. Membrane pores were less frequent in the AAV-GPx3-treated group (Fig. 9M). Taken together, these results indicated that GPx3 inhibits IRAK4- and NLRP3 inflammasome-dependent pyroptosis in microglia after SCI.

Discussion
The present study indicates a potential therapeutic role of GPx3 in SCI. We found that overexpression of GPx3 could alleviate neuroinflammation after SCI, and its mechanism was closely related to the inhibition of microglia-mediated oxidative stress and NLRP3-dependent pyroptosis. These therapeutic effects were mediated by inhibition of the IRAK4/NLRP3 signaling pathway in vivo and in vitro. Silencing IRAK4 attenuated the adverse effects of GPx3 deficiency. To the best of our knowledge, this is the first study to describe the precise mechanisms underlying the therapeutic benefit of GPx3 in SCI.
The GPX (glutathione peroxidase) family constitutes a critical antioxidant enzyme system that plays a regulatory role in oxidative stress responses both within and outside of cells.GPX1, an antioxidant enzyme, safeguards neurons post-SCI by combating ROS, curbing oxidative stress, and preventing cell death, with its levels dynamically changing post-injury (Li et al., 2020). GPX2 is primarily expressed in the intestine and gastrointestinal epithelial cells, regulating local antioxidant balance and maintaining intestinal homeostasis (Marzullo et al., 2019). GPX3 is the main plasma antioxidant enzyme capable of regulating peripheral and central ROS levels (de Carvalho et al., 2020). GPX4 is the only member of the GPX family that can directly reduce phospholipid hydroperoxides and plays an important role in inhibiting lipid peroxidation (Krause, 2018).
SCI are typically divided into primary and secondary injuries. Primary injury is caused by mechanical impact on the spine. Because primary injury is irreversible, basic, and clinical research have focused on developing therapeutic strategies for secondary injuries (Wang et al., 2018). Secondary injury is characterized by inflammation, oxidative stress, necrosis, and apoptosis of the spinal cord. This secondary injury can be modulated through various mechanisms. In the central nervous system, nitric oxide induces cellular damage in neurodegenerative diseases by regulating synaptic plasticity, participating in oxidative stress responses (Calabrese et al., 2007; Zhang et al., 2008) and influencing vascular regulation. Of these, oxidative stress, one of the severe pathological responses, has critical effects on inflammation after SCI (Zhou et al., 2020; Tran et al., 2018; Fan et al., 2019). Numerous studies have indicated that GPx3 exerts a strong antioxidant effect. However, the potential protective mechanism of GPx3 against SCI remains unclear.
Together with selenoprotein P, GPx3 is the only extracellular enzyme in the GPx family that is extensively secreted into the plasma and accounts for more than 97% of all plasma selenium in mice and humans (Gong et al., 2020). GPx3 can also be found in other tissues and the surrounding extracellular environment, playing a crucial role in regulating ROS and preventing hydroperoxide-mediated inflammatory responses in the body’s extracellular environment. Studies have suggested that the GPx3 deficiency results in ROS accumulation, including hydrogen peroxide and organic hydroperoxides (Carvalho et al., 2022; Nirgude and Choudhary, 2021). To investigate the expression pattern of GPx3 in SCI, spinal cord, plasma, and primary microglia were harvested. Consistent with previous studies, we found that GPx3 levels decreased in the above samples after SCI. The expression of GPx3 in the spinal cord increased as the injury time increased after SCI. These findings implied that GPx3 is a potential therapeutic target for SCI. We aimed to better understand the protective mechanism of GPx3 based on the expression pattern of GPx3 after SCI. We overexpressed GPx3 in the local spinal cord of mice using AAV and silenced and overexpressed GPx3 in BV2 cells using lentiviruses.
Behavioral test is regarded as the gold standard for evaluating functional recovery in patients with SCI. Therefore, in this study, the rearing test and FLS scores were used to assess the left forelimb motor function in mice with SCI. We found that the overexpression of GPx3 in SCI mice improved the motor function of the ipsilateral forelimb. Behavioral test outcomes showed that the mice from the AAV-GPx3 group improved significantly by 10 and 12-weeks post-injury. This is most likely because SCI is a fatal injury that causes serious neurological impairment that is difficult to improve within a short time period. Moreover, H&E staining at 12 weeks post-injury demonstrated that GPx3 significantly relieved the structural disorder of the spinal cord. These results indicated that GPx3 is a potential therapeutic molecule for the treatment of SCI.
In the present study, ROS levels in the spinal cord were assessed by DHE staining. The fluorescence intensity of DHE staining showed that GPx3 robustly eliminated ROS. Furthermore, AOPPs and MDA levels in the plasma and spinal cord were determined to further verify the antioxidant role of GPx3. AOPPs are recognized as novel markers of oxidative stress that initiate ROS bursts (Ding et al., 2016). As a byproduct of lipid peroxidation, higher MDA levels indicate oxidative stress (Ott et al., 2014). Our results indicated that AOPPs and MDA levels rapidly decreased in the AAV-GPx3 group, corroborating the antioxidant effect of GPx3. Microglia are the resident immune cells in the CNS and play a role in preserving immunological homeostasis and defense. They have exceptional functional flexibility and the ability to develop in response to CNS damage and acute diseases. SCI induces massive activation of microglia, which leads to neuroinflammation through oxidative stress and the production of proinflammatory cytokines (Lin et al., 2015). Our data showed that GPx3 overexpression inhibited microglial activation in the spinal cord. These findings showed that GPx3 reduced microglia-mediated oxidative stress following SCI.
In the present study, we found a correlation between GPx3 and IRAK4 expression in primary microglia. Overexpression or silencing of GPx3 can induce the downregulation or upregulation of IRAK4. We further investigated how GPx3 and IRAK4 interact using molecular docking analysis. Co-IP results indicated that GPx3 directly binds with IRAK4. Our data suggest that IRAK4 acts downstream of GPx3. IRAK4 is a serine-threonine kinase that mediates the Toll-like receptor and IL-1 signaling pathways, which regulate innate immunity and inflammation (Smith et al., 2019). IRAK4 is recruited to the receptor complex and phosphorylates IRAK1 to initiate autophosphorylation of IRAK1. Thus, IRAK4 is a key target in the signal transduction pathways of inflammation (Wu et al., 2022). A study by Ankita Singh revealed a significant role of IRAK4 in the accumulation of ROS (Singh et al., 2016). In line with this study, we found that the overexpression or silencing of GPx3 induced the upregulation or downregulation of ROS production in BV2 cells. Silencing IRAK4 alleviates the ROS production upregulated by GPx3 deficiency. These results suggest that GPx3 reduced ROS production via IRAK4.
Pyroptosis is a type of inflammasome-mediated programmed cell death that is triggered by caspase-1 activation. Additionally, pore formation in the cell membrane, swelling, and rupture are characteristics of pyroptosis (Shi et al., 2017; Shi et al., 2015). The occurrence of pyroptosis in microglia promotes the secretion of IL1β and IL18 which further aggravates neuroinflammation (Voet et al., 2019). Compared to other CNS diseases, pyroptosis in SCI has not been extensively studied. Therefore, in the present study, we focused on the regulation of microglial pyroptosis following SCI. Western blot and fluorescence experiments were conducted to assess the expression of IL1β and IL18 in microglia. We found that GPx3 effectively reduced their expression, thereby alleviating pyroptosis-induced neuroinflammation. LDH release and PI uptake experiments are regarded as classical markers of pore formation in the plasma membrane and are widely used to confirm pyroptosis. TEM provides a direct visual way to observe the morphological features of proptosis in microglia. These results indicate that GPx3 reduces membrane pores and inhibits pyroptosis in microglia. These findings indicate that GPx3 suppresses pyroptosis and neuroinflammation in microglia. However, the specific mechanism of how GPx3 inhibits microglial pyroptosis needs to be further investigated.
Our earlier study showed that the NLRP3 inflammasome is essential for microglia (Liu et al., 2021b). Inflammasomes are a group of protein complexes that trigger inflammatory responses leading to pyroptosis. The NLRP3 inflammasome, which is composed of NLRP3, ASC, and pro-caspase-1, has been the subject of many studies (Zheng and Kanneganti, 2020; Huang et al., 2021). Pathogen-associated molecular patterns and damage-associated molecular patterns are essential for NLRP3 inflammasome activation, which in turn activates downstream signaling pathways with pattern recognition receptors on the cell surface. ROS are believed to be key components in the activation of the NLRP3 inflammasome. Upon activation, NLRP3 recruits ASC and cleaves pro-caspase-1 into its active forms, which process pro-IL1β and pro-IL18 into their mature forms (IL1β and IL18) ultimately leading to pyroptosis (Briard et al., 2020; Mills et al., 2017; Wada and Makino, 2016). Consistent with this view, our results confirmed that GPx3 inhibits the expression of NLRP3 inflammasomes, including NLRP3, ASC, and caspase-1. Meanwhile, the maturation of IL1β and IL18 were also suppressed by GPx3 in BV2 cells. Rescue experiments were conducted to elucidate the critical role of IRAK4 in NLRP3-dependent pyroptosis signaling pathway. Silencing IRAK4 using siRNA inhibited NLRP3 inflammasome activation and microglial pyroptosis induced by GPx3 deficiency. To strengthen the conclusions of our in vitro experiments, GPx3 overexpressing mice models were established. In vivo experiments demonstrated that AAV-GPx3 treatment significantly inhibited the activation of the NLRP3 inflammasome and the expression of IRAK4 and inflammatory cytokines. Classical morphological alterations of pyroptosis in microglia were mitigated by AAV-GPx3 treatment. Therefore, we confirmed our hypothesis that GPx3 exerts its suppressive effects by inhibiting the pyroptosis signaling pathway.
Collectively, our results provide the first evidence that GPx3 significantly reduced microglia-mediated oxidative stress and neuroinflammation following SCI. The specific mechanisms may be closely related to the activation of the IRAK4/ROS/NLRP3 signaling pathway and the inhibition of pyroptosis, which may provide a possible innovative approach for the treatment of SCI.
Experimental limitations
The current study had several limitations. First, we selected the only NLRP3 inflammasome to explore the relationship between GPx3 and pyroptosis. However, it is unclear whether other inflammasomes are involved in the process of GPx3-mediated pyroptosis inhibition. Secondly, the therapeutic effects of GPx3 at other time points should be investigated. Third, multiple microglia-mediated mechanisms participate in the pathology of SCI. However, this study focused only on pyroptosis in SCI neuroinflammation.
Methods and Materials
Animals
Adult male C57 mice (8 weeks old) were purchased from the Laboratory Animal Center of Southern Medical University and housed at the Laboratory Animal Center of Nanfang Hospital. The mice were on a 12-h light–dark cycle with ad libitum access to food and water. All animal experiments were approved by the Laboratory Animal Care and Use Committee of Nanfang Hospital, Southern Medical University, and all animal procedures were performed in accordance with the National Guidelines for the Care and Use of Animals.
Preparation of adeno-associated virus particles and stereotaxic surgery
Adeno-associated virus 9 (AAV9) engineered to overexpress mouse GPx3 under the microglial-specific CD68 promoter and a control AAV were provided by GeneChem Biotech (Shanghai, China). The titers for AAV9 and control AAV (AAV-NC) particles were 2.06 × 1012 and 6.26 × 1012 vg/mL, respectively.
To investigate the in vivo effects of GPx3, a cervical spinal cord injection was administered four weeks before the C5 contusion. AAV injection was performed as described previously (Brommer et al., 2021). Immediately after being anesthetized, mice were transferred to a stereotaxic frame, and a 10 μL Hamilton Neuros syringe, coupled to a micro syringe pump controller (RWD), was inserted to inject AAV-GPx3 (2.06 × 109 vg; 1 μL) at a rate of 200 nL/min into the C5 spinal cord at a depth of 1 mm at each corner of the lesion site. The syringe was left in the C5 level of the spinal cord for 5 min before being slowly withdrawn to limit reflux. Blood vessels and nerves were avoided during the injection.
Cervical spinal cord injury
Mice underwent C5 hemicontusion as previously described (Huang et al., 2022). Briefly, the mice were anesthetized using isoflurane (3% for induction and 1.5%–2% for maintenance), a partial laminectomy was performed at the C5 section to expose the left side of the spinal cord, and a custom-designed clamp was used to stabilize the C4–C6 vertebral columns. An impactor tip (diameter = 1.0 mm) with a set displacement of 1.2 mm at 300 mm/s was pointed to the left side of the C5 segment and then triggered.
Animal experimental designs
Experiment 1: To ascertain the GPx3 expression pattern following SCI, 32 mice were randomly assigned to four groups (eight mice per group): Sham, 1-week post-SCI, 6 weeks post-SCI, and 12 weeks post-SCI groups.
Experiment 2: To evaluate the effect of GPx3 on functional recovery and histological outcome, 48 mice were randomly divided into four groups (12 mice per group) at 12 weeks post injury: Sham+AAV-NC, Sham+AAV-GPx3, SCI+AAV-NC, and SCI+AAV-GPx3 groups.
Experiment 3: To investigate the protective mechanism of GPx3 after SCI, 36 mice were randomly divided into four groups (nine mice per group) at 1 week post injury: Sham+AAV-NC, Sham+AAV-GPx3, SCI+AAV-NC, and SCI+AAV-GPx3.
Behavioral assessment
All mice were acclimatized to the testing room prior to behavioral testing. A rearing test and a forelimb locomotor scale (FLS) were used in this study. As mice are nocturnal, all behavioral tests were conducted at night. Blinded scoring was used to ensure that the observers did not know the groups. Different groups of mice were assessed at week 1 and every 2 weeks following the injury.
Rearing test
Mice were placed separately in a 20-cm diameter transparent cylinder and recorded for 15 min. Climbing movements (touch of left forelimb, right forelimb, and both forelimbs) or all climbing movements within 15 min were examined framed-by-frame for each mouse. Subsequently, the utilization rate of the ipsilateral forelimb was calculated.
Forelimb motor function assessment
Based on the joint’s range of motion, body weight support, and balance between the trunk and forelimbs, forelimb motor function was assessed using a 0–17-point scale derived from the Beattie and Bresnahan (BBB) locomotor rating scale (Sandrow et al., 2008). The mice were placed in an open field and videotaped for 5 min.
ROS measurements
To evaluate oxidative stress levels in the spinal cord, spinal cord samples (length, 1.0 cm) separated at the C5 level were collected one-week post-SCI from each group of mice. The tissue samples were processed and dissected at 4°C, flash frozen in liquid nitrogen, and stored at −80°C. DHE staining was performed according to manufacturer’s protocol. Briefly, 10 μm thick frozen spinal cord sections were incubated with 2 μmol/L fluorescent DHE dye at 37°C for 30 min in a humidified chamber in the dark. Co-staining of DHE and microglia was performed using immunofluorescence staining. After DHE staining, the sections were incubated with a mouse monoclonal antibody against ionized calcium-binding adapter molecule 1 (Iba-1) (1:100, Santa Cruz Biotechnology, USA) and fluorescently labeled using a mouse immunoglobulin G labeling kit (Zenon Alexa Fluor 488, Invitrogen, USA). The sections were observed under a Leica confocal microscope (Zeiss, Wetzlar, Germany) to detect red fluorescence through a 535 nm long-pass filter.
Measurement of malondialdehyde, superoxide dismutase, and advanced oxidation protein products
MDA, SOD, and AOPPs are common indicators of oxidative stress. MDA, a secondary product of lipid peroxidation, was measured using the method described by Ohkawa et al. (Ohkawa et al., 1979). The SOD concentration was measured using a commercial kit (Jiancheng, Nanjing, China). Samples were collected and processed, reaction buffer was prepared, and appropriate amounts of reaction buffer and sample extracts were added to 96-well plates.Nitrite were added to initiate the reaction. Absorbance was measured at 550 nm. The SOD activity was calculated using a standard curve. To detect the levels of MDA, SOD, and AOPPs, C5 sections of cervical cord tissue and plasma were collected one week after SCI. The absorbance of the supernatant was measured at 532 nm. AOPPs are dityrosine-containing and cross-linking protein products recognized as biomarkers of oxidative stress. Under acidic conditions, AOPPs were detected immediately in the supernatant, and the absorbance was measured at 340 nm. The levels of MDA and AOPPs are expressed as MDA (nmol)/protein (mg) or mol/L.
Co-immunoprecipitation assay
Co-immunoprecipitation (Co-IP) was performed using a Pierce Classical IP Kit (Thermo Scientific) according to the manufacturer’s protocol. Briefly, primary microglia were seeded in 6-well plates. The cells were collected and lysed in IP lysis buffer. Proteins (600 μg) were immunoprecipitated using protein A/G magnetic beads. The subsequent steps were performed according to the manufacturer’s instructions. The final samples were analyzed by western blot using the indicated antibodies.
Enzyme-linked immunosorbent assay
C5 sections of the cervical cord tissues and plasma were collected one week after SCI, and the supernatant of the primary microglia was collected after stimulation for 24 h. After centrifugation, the supernatant was subjected to ELISA. The GPx3 content was measured using an ELISA kit according to the manufacturer’s instructions. GPx3 concentrations were calculated using standard curves.
Primary microglia cultures
To investigate the specific mechanism underlying the effects identified in the in vivo experiment and to simulate the natural microenvironment of the spinal cord, we used primary microglia from mice to perform the in vitro experiments. Primary microglial cells were isolated according to our previously published procedures (Murgoci et al., 2020). Briefly, postnatal mouse pups (day, P0–P1) were collected and transferred into 0.125% trypsin–EDTA at 37°C for 15 min; subsequently, trypsinization was stopped with 10% fetal bovine serum (FBS). Mouse brains were triturated and passed through a 70 μm nylon mesh cell strainer. Cells were then cultured in Dulbecco’s modified Eagle’s medium/F-12 nutrient mixture (DMEM/F12) containing 10% FBS and incubated in a CO2 incubator at 37°C for 14 days. On day 14, confluent mixed glial cell cultures were agitated for 3 h at 250 rpm to separate the microglia, and the microglial cells were cultured in fresh DMEM/F12 containing 10% FBS and 1% penicillin–streptomycin and maintained in a 5% CO2 incubator under a humidified atmosphere of 95%. Microglia were treated with LPS (1 μg/mL) for 24 h and ATP (5 mM) for 1 h.
Murine BV2 culture
Immortalized murine BV-2 cells, which are derived from primary mouse microglial cells, were used as an alternative model because they have roughly the same inflammatory profile as primary microglia. Thus, BV2 cells were used for gain-of-function or loss-of-function experiments in this study. BV2 microglial cells were purchased from Shanghai Cell Research Center (Shanghai, China). The cells were cultured in DMEM (4.5 g/L glucose) with 10% FBS and 1% penicillin–streptomycin and maintain at 37°C under 5% CO2. The BV2 cells were digested with trypsin and passaged for additional experiments when they reached approximately 90% confluence. The cells were treated with LPS (1 μg/mL) for 24 h and ATP (5 mM) for 1 h to induce pyroptosis.
Lentiviral vector construction
The GPx3 shRNA and negative control sequences were designed and synthesized by Obio Co., Ltd. (Shanghai, China). Cells were treated with puromycin (1 μg/mL) (InvivoGen, San Diego, CA, USA) to produce stably transfected cells.
Full length GPx3 cDNA-containing or negative control viruses were procured from Obio Co., Ltd. (Shanghai, China). Stably transfected cells were selected using puromycin (1 μg/mL).
Small interfering RNA transfection
IRAK4-targeting and scrambled non-targeting (siNC) siRNAs were purchased from Tsingke (Beijing, China). BV2 cells were seeded in 6-well plates (SORFA, China). Lipofectamine 2000 (Invitrogen) was used for siRNA transfection, in accordance with the manufacturer’s instructions. Lipofectamine 2000 reagent was mixed with Opti-MEM (Gibco) for 5 min; subsequently, the respective siRNAs were added to the solutions for 20 min at room temperature. After incubation, the mixtures were diluted in serum-free medium and added to each well. The final concentration of the siRNAs was 20 nM. All experiments were performed 48 h after transfection.
H&E staining
C5 sections of the cervical spinal cord were collected 12 weeks post-SCI. Frozen sections of the spinal cord tissues (thickness, 20 µm) were stained with H&E. H&E staining was performed according to the manufacturer’s instructions using a staining kit (Solarbio, Beijing, China). A light microscope (Leica, Germany) was used to image the H&E-stained spinal cord slices.
Transmission electron microscopy
BV-2 cells were fixed in 2.5% glutaraldehyde at 4°C for 24 h, and post-fixed in 1% osmium tetroxide. The cells were then dehydrated in an ethanol gradient and then acetone. C5 sections of the cervical spinal cord were collected one-week post-SCI, cut into 6 mm pieces, fixed in 2.5% glutaraldehyde for 24 h, and post-fixed in 1% osmium tetroxide for 3 h. After dehydration in an ethanol gradient (50%, 70%, 90%, and 100%) for 10 min each, the samples were embedded in resin and cut to a thickness of 60–90 nm. The cells and sections were stained with uranyl acetate and lead citrate and observed under an electron microscope (HT7700; Hitachi, Japan).
Western blot analysis
BV2 cells and the C5 sections of the cervical spinal cords (1-week post-injury) were lysed with ice cold lysis buffer (pH 8.0, 50 mM Tris-HCl, 150 mM NaCl, 1% Triton-X 100, 100 µg/mL PMSF). Protein concentration was determined using BCA assay (Pierce BCA Protein Assay Kit, Thermo Scientific), and 30 µg protein per sample was loaded in each lane, separated by 10% SDS-PAGE, and transferred to a PVDF membrane. Membranes were then blocked in 5% BSA at room temperature for 1 h, followed by incubation with specific primary antibodies at room temperature for 1 h (anti-GPx3, 1:1000, Santa Cruz Biotechnology, USA; anti-Iba1, 1:1000, Santa Cruz Biotechnology; anti-IRAK4, 1:1000, Cell Signaling Technology, Inc.; anti-NLRP3, 1:1000, Cell Signaling Technology, Inc.; anti-caspase p10, 1:1000, Abcam, UK; anti-IL1β, 1:1000, Cell Signaling Technology; anti-IL18, 1:1000, Abcam; anti-GSDMD, 1:1000, Abcam; anti-IRAK4, 1:1000, Cell Signaling Technology, Inc.). Thereafter, the membranes were incubated with secondary antibodies (anti-mouse and anti-rabbit IgG, 1:5000; Abmart, China) for 1 h at room temperature. Enhanced chemiluminescence reagent (Merck Millipore) was used to examine the blots. Each experiment was repeated thrice, and band intensities were measured using a Gel-Pro Analyzer (Media Cybernetics, Sarasota, USA).
Immunofluorescence staining
For spinal cord samples, the mice were euthanized at one-week post-injury and intracardially perfused with PBS, followed by 4% paraformaldehyde (PFA). C5 sections of cervical cord tissue were post-fixed in 4% PFA for 6 h and then dehydrated in sucrose gradients of 12%, 18%, and 24% for 24 h each. The spinal cords were embedded and frozen in Tissue-Tek O.C.T. compound (Sakura Finetek, USA). Spinal cord sections of 10 µm thickness was cut using a Leica CM1950 cryostat. Sections were incubated with primary antibodies overnight at 4°C. (anti-Iba1, 1:200, Santa Cruz Biotechnology; anti-IRAK4, 1:400, Cell Signaling Technology, Inc.; anti-NLRP3, 1:400, Cell Signaling Technology, Inc.; anti-caspase p10, 1:200, Abcam; anti-IL1β, 1:200, Cell Signaling Technology, Inc.). After three PBS washes, the tissue sections were incubated for 2 h with the following secondary antibodies: Alexa Fluor 555 goat anti-rabbit IgG and Alexa Fluor 647 donkey anti-mouse IgG (Abcam), followed by counterstaining with DAPI for 10 min. Images were captured using a confocal microscope (LSM980, ZEISS, Germany).
BV2 cells were seeded in 12-well plates (NEST Biotechnology, Wuxi, China). After reaching 80% confluence, the cells were rinsed with PBS, fixed with 4% PFA for 30 min at room temperature, and washed thoroughly with PBS. Thereafter, the cells were incubated with primary antibodies at 4°C overnight (anti-IRAK4, 1:400, Cell Signaling Technology, Inc.; anti-NLRP3, 1:400, Cell Signaling Technology, Inc.; anti-GSDMD, 1:200, Abcam; anti-IL1β, 1:200, Cell Signaling Technology, Inc.) and then incubated with Alexa Fluor 594-labeled goat anti-mouse IgG antibody (Abcam) for 2 h. Subsequently, after washing, nuclei were stained with DAPI for 5 min. Images were captured using a confocal microscope (LSM980, ZEISS).
Propidium iodide staining
The BV2 cells were seeded in 24-well plates and allowed to reach 80% confluence. Following treatment, the cells were washed thrice with cold PBS. Thereafter, the cells were incubated with DAPI and PI (Beyotime, Shanghai, China) at 37°C. After 15 min, the cells were observed using a confocal microscope (LSM980, ZEISS).
Lactate dehydrogenase (LDH) release assays
An LDH kit (Beyotime) was used to assess cell death. BV2 cells were seeded at the desired confluence in 96 well plates and subjected to different treatments. A microplate reader was used to measure the LDH activity in the culture medium and cellular lysate. The LDH release % was estimated as: [(infected cell LDH release − spontaneous LDH release)/(total LDH release − pontaneous LDH release)] × 100.
Statistical analysis
All experiments were independently repeated in triplicates. Statistical analysis data are presented as means. Results are presented as means ± SEMs and analyzed by SPSS 20.0 (IBM, NY, USA) and GraphPad Prism 9 (GraphPad Software, CA, USA) software. The mean differences between multiple groups were tested using ANOVAs. Differences were considered statistically significant at p < 0.05.
Footnotes
Acknowledgment
The authors are particularly grateful to Professor Qingan Zhu for his valuable suggestions and advice.
Availability of Data and Materials
All data supporting the conclusions of this article are provided in the text and figures.
Ethics Approval and Consent to Participate
All experimental procedures were approved by the Experimental Animal Ethics Committee of Southern Medical University and carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Authors’ Contributions
Z.Z., L.W., J.C., and Z.L. conceived the project and designed the studies. Z.L. performed most experiments, analyzed data, and wrote the article. Z.Z. and L.W. revised the article. K.T., H.M., J.C., X.X., and P.Z. conducted parts of the animal surgery and performed the experiments. C.L., Z.H., X.Y., R.D., and P.L. contributed to article preparation. All authors have read and approved the final version of the article.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by the China Postdoctoral Science Foundation [2022M711515], Guangdong Basic and Applied Basic Research Foundation [No.2021A1515111183], Science and Technology Program of Guangzhou (No.2025A04J4097) and Director Foundation of NanFang Hospital [2021C001].