
Abstract
Aims:
Silicosis is a lung disease caused by inhalation of silica particles. Both silica-induced oxidative stress and aberrant activation of the Wnt/β-catenin signaling pathway are potential targets in the treatment of pulmonary fibrosis. Dickkopf-1 (Dkk1), an inhibitor of the Wnt/β-catenin signaling pathway, plays regulatory roles in cell fate determination and immune responses. Our previous study demonstrated that adenoviral vector-mediated Dkk1 gene transfer alleviated the silica-induced mouse silicosis. However, the mechanism of therapeutic action of Dkk1 in silicosis is yet completely understood; together with the drawbacks of adenoviral vectors in gene therapy, we investigated the therapeutic effect and mechanisms of Dkk1 by employing an adeno-associated virus (AAV) vector in a silicosis mouse model.
Results:
The AAV vector could efficiently transduce the Dkk1 gene in silicotic lung during both the early and the late phases of disease, resulting in an alleviation of silicotic lesions, improvement of pulmonary compliance, and radiological findings. Mechanistic studies further demonstrated that the transduction of Dkk1 inhibited the silica-activated Wnt/β-catenin signaling and reduced the silica-induced reactive oxygen species-producing enzyme NADPH oxidase 4, oxidative stress regulator nuclear factor erythroid 2-related factor 2, and signaling molecules binding immunoglobulin protein and C/EBP homologous protein. In addition, shRNA-mediated downregulation of Dkk1 exacerbated the progression of silicosis in mice, whereas the treatment of ROS scavenger
Innovation and Conclusion:
This study provides an insight into the mechanism by which Dkk1 inhibits the silica-induced Wnt signaling and oxidative stress to mitigate the pathogenesis of lung silicosis and evidence of the potential of AAV-mediated Dkk1 gene transfer as an alternative approach in silicosis treatment. Antioxid. Redox Signal. 42, 529–546.
Introduction
Silicosis is an interstitial lung disease caused by inhalation of crystalline silica dust, which remains a significant public health concern in occupational settings in many developing countries, despite the incidence is considerably decreased over years (Handra et al., 2023). Up to date, the silicosis remains incurable, and it is therefore imperative to discover novel targets for the treatment of this disease. Wnt/β-catenin signaling is a critical developmental pathway that is involved in a variety of cellular processes contributing to embryonic development, tissue regeneration, and stem cell fate determination (Aros et al., 2021; Liu et al., 2022; Logan and Nusse, 2004). In addition, accumulating evidence suggests that Wnt signaling also exhibits abilities to regulate oxidative stresses and immune responses (Karimaian et al., 2017; Mukherjee and Balaji, 2019; Pai et al., 2017; Zhou et al., 2019). However, an aberrant Wnt/β-catenin signaling activity has been implicated in the pathogenesis of many diseases, including idiopathic pulmonary fibrosis (IPF) (Chanda et al., 2019; Chilosi et al., 2003) and silicosis (Cai et al., 2022; Dai et al., 2016; Ma et al., 2020; Wang et al., 2015). Therefore, blocking the Wnt/β-catenin signaling has shown a great promise in the treatment of both IPF and silicosis (Cai et al., 2022; Dai et al., 2022; Dai et al., 2016; Gottardi and Konigshoff, 2013; Wang et al., 2015).
Using AAV-mediated Dkk1 inhibits aberrant Wnt signaling and oxidative stress exhibits a promise of treatment of silicosis, his study provides an insight into the mechanism of the regulation of oxidative stress by targeting Wnt signaling for the treatment of silicosis and evidence of proof-of-principle for gene therapy of silicosis using AAV vectors.
Dickkopf-1 (Dkk1) is an antagonist of the Wnt/β-catenin signaling pathway (Jaschke et al., 2020), which plays a crucial role in regulating Wnt/β-catenin signaling activity and immune responses during inflammation and injury repair (Park et al., 2021). A growing number of studies have demonstrated that Dkk1 is involved in the pathogenesis of a variety of diseases (Huang et al., 2018; Tian et al., 2003), and thus, it is a potential therapeutic target of many disorders (Chu et al., 2021; Fulciniti et al., 2009; Tao et al., 2022; Ye et al., 2023), such as IPF (Pfaff et al., 2011) and experimental silicosis (Cai et al., 2022). We have previously demonstrated that adenoviral vector-mediated intrapulmonary Dkk1 gene transduction could significantly alleviate the pathogenesis of silicosis in mouse lung (Cai et al., 2022). However, mechanisms underpinning the experimental therapeutic effect have yet to be fully investigated. In addition, gene therapy using adenoviral vectors faces many challenges including a transient expression of transgene and the generation of neutralizing antibodies against adenovirus and induces a wide variety of immune responses (Zhang et al., 2023). To overcome these drawbacks of adenoviral vectors, adeno-associated virus (AAV) vectors may be one of the best solutions, owing to its ability to mediate long-term and stable expression of transgene but induce mild immune responses, which make the AAV a safer and more promising vector for gene therapy over adenovirus vectors (Naso et al., 2017).
Alveolar type 2 (AT2) cells are essential for the preservation of the architecture and physiology of normal lung by producing secrete pulmonary surfactant and exerting functions of stem/progenitor cells for maintaining the integrity of alveolar epithelium (Liu et al., 2024), for which the Wnt signaling is required (Nabhan et al., 2018; Nabhan et al., 2023). The impairment of AT2 cell self-renewal and differentiation of AT2 into AT1 contributed to the pathogenesis of IPF (Jiang et al., 2020). The mutation of the surfactant associated protein C (SFTPC or SPC) gene, an AT2 cell type-specific marker, is associated with the development of human spontaneous pulmonary fibrosis (Brasch et al., 2004). The spontaneous lung fibrosis phenotype was reproduced in mice expressing mutant Sftpc (Nureki et al., 2018). Of note, a previous study has demonstrated that the exposure of silica and other pulmonary toxicants could induce AT2 cell hypertrophy and hyperplasia (Miller and Hook, 1990), indicating that AT2 cells are a potential cell type of target for the treatment of silicosis. For gene therapy by targeting AT2 cells, the SPC promoter has been shown to specifically facilitate transgene nuclear import and expression in AT2 cells only (Degiulio et al., 2010), suggesting that the SPC promoter-derived Dkk1 transduction can modulate Wnt signaling of AT2 cells in a cell type-specific manner.
It has been well documented that oxidative stress is a key molecular process in pulmonary fibrosis (Chilosi et al., 2003; Estornut et al., 2021; Otoupalova et al., 2020) and silicosis (Jalali et al., 2016; Vallyathan et al., 1997). The induction of oxidative stress and production of reactive oxygen species (ROS) in response to an inhalation of environmental pollutants such as silica dust, coal, and polystyrene microplastics are one of the major contributors causing lung injury and the pathogenesis of silicosis (Barsan et al., 2021; Kurt et al., 2023; Vallyathan et al., 1997). In contrast, the reduction of oxidative stress by administration of phytomedicines with antioxidative activity, such as quercetin (Yao et al., 2023), curcumin (Barsan et al., 2021), and antioxidants melatonin (Li et al., 2022) and metformin (Li et al., 2023), ameliorates the pathogenesis and pulmonary fibrosis in experimental silicosis models. Thus, targeting oxidative stress is a holistic approach for the treatment of lung diseases such as IPF and silicosis (Estornut et al., 2021). Given the fact that oxidative stress is responsible for DNA damage and cell-programmed deaths and is linked to Wnt/β-catenin signaling (Dong et al., 2021; Karimaian et al., 2017; Zhou et al., 2019), in the present report, by using AAV-mediated intrapulmonary Dkk1 gene transduction in silicosis mice, we therefore tested a hypothesis that Dkk1 regulated silica-activated Wnt/β-catenin and oxidative stress signaling pathways and subsequently alleviated the pathogenesis of experimental silicosis.
Results
AAV-mediated Dkk1 gene transfer restricts silicosis development and progression, along with reduced silica-induced oxidative stress in mouse lung with an early phase of silicosis
To investigate whether targeting Wnt/β-catenin signaling by AAV-mediated Dkk1 gene transfer could restrict the development of silica-induced silicosis in mice, AAV was intrapulmonarily delivered to silicosis mice at 2 weeks post silica challenge, when the mice were at an early phase of silicosis development. The mice were analyzed for pathological and molecular changes at 4 weeks (28 days) (Fig. 1A, Supplementary Fig. S1A) post the challenge. Since Wnt signaling is found selectively activated in different stem cell populations in the lung (Nabhan et al., 2023), the AAV5SPC-Dkk1 was employed to tissue specifically transduce Dkk1 expression in AT2 cells. The body weight (BW) was not significantly altered between mice received different AAV vectors (Supplementary Fig. S1B), although a reduced ratio of wet lung/BW was observed in silicosis mice that were administrated with AAV5CMV-Dkk1 or AAV5SPC-Dkk1, compared with the AAV5CMV-Luc control vector (Supplementary Fig. S1C). Histopathological assessments demonstrated that the AAV-mediated Dkk1 transduction at 2 weeks post silica challenge significantly reduced the silicotic nodule formation and extracellular matrix (ECM) deposition in the lungs of silicosis mice evaluated at 4 weeks (Fig. 1B, C), compared with the AAV luciferase vector. Biochemistry analysis showed the transduction of Dkk1 increased the circulating Dkk1 protein concentration (Fig. 1D), coupled with the reduction of Wnt3a concentration (Fig. 1E), compared with the AAV luciferase control vector. The changes of both circulating Dkk1 and Wnt3a proteins were in line with the abundances of Wnt3a ligand and active β-catenin (ABC), and the mediator of Wnt/β-catenin signaling in the lung tissues was determined by immunoblotting (IB) assay (Fig. 1F, G) and/or immunohistochemical staining (IHC; Fig. 1H). In addition, both IB (Fig. 1F, G) and IHC (Fig. 1H) assays showed the reduction of epithelial mesenchymal transition (EMT) markers
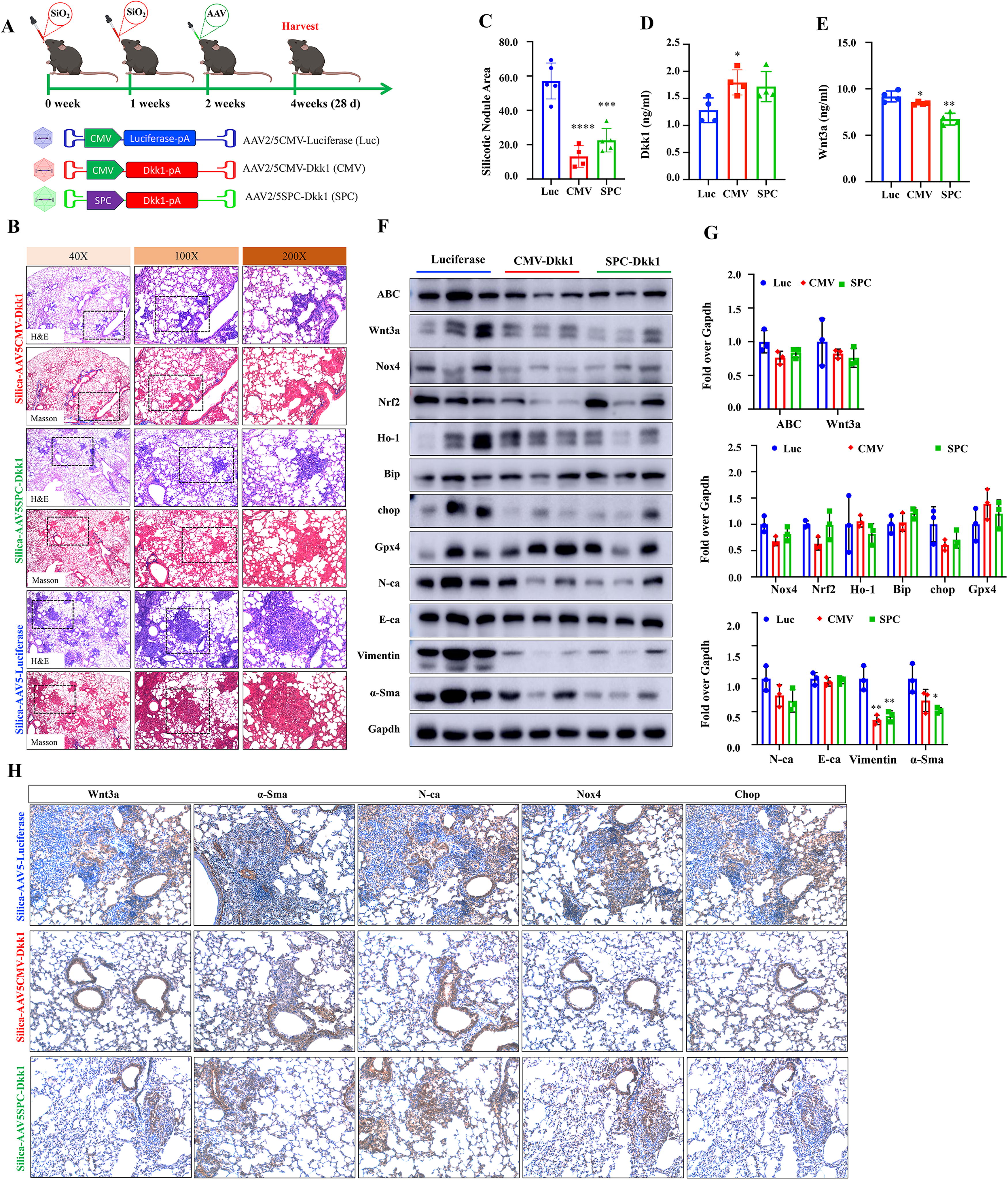
Notably, the IB assay also showed that less abundant signaling components related to oxidative stress, including NADPH oxidases (Nox4), nuclear factor erythroid 2-related factor 2 (Nrf2), heme oxygenase 1 (Ho-1), endoplasmic reticulum (ER), heat shock protein 70, chaperone binding immunoglobulin protein (BiP), and C/EBP homologous protein (Chop), but more abundant lipid hydroperoxidase glutathione peroxidase 4 (Gpx4), in the lung tissues of silicosis mice infected with either AAV5CMV-Dkk1 or AAV5SPC-Dkk1, compared with AAV5CMV-Luc at 2 weeks after the infection (4 weeks post the silica challenge; Fig. 1F, G), highlighting the involvement of underlying mechanisms by which the AAV-mediated Dkk1 transduction reduces the silica-induced oxidative stress, subsequently restricts the development of silicosis.
To investigate whether the delivery of AAV-mediated Dkk1 gene transfer at an early phase of silicosis could inhibit the disease progression of silicosis in mice, the mice were delivered with AAV at 2 weeks and analyzed at 8 weeks (56 days) post the challenge (Fig. 2A, Supplementary Fig. S1A). Histopathological analysis showed that the Dkk1 transduction significantly reduced the silicotic nodule formation (Fig. 2B, C) and hydroxyproline contents (Fig. 2D). However, unlike an increased circulating Dkk1 and reduced Wnt3a seen in mice at 2 weeks post the Dkk1 transduction (Fig. 2D, E), the circulating Dkk1 was not altered (Fig. 2E), whereas the plasma Wnt3a level was increased in silicosis mice at 6 weeks post the infection of Dkk1 vectors, particularly the AAV5SPC-Dkk1, compared with the Luc vector (Fig. 2F). The abundance of ABC and Wnt3a proteins was reduced in the lung tissues infected with AAV5CMV-Dkk1, but they were not changed in the lung transduced by AAV5SPC-Dkk1, as determined by IB assay (Fig. 2G, H). The IB assay also showed a reduction of EMT markers N-ca, E-ca, α-SMA and vimentin, and Nox4, Chop, Ho-1, and BiP in the lungs of silicosis mice infected with AAV5CMV-Dkk1, compared with AAV5CMV-luc (Fig. 2G, H). Of note, the administration of the AAV5SPC-Dkk1 vector did not significantly alter the abundance of the above-examined proteins (Fig. 2G, H).
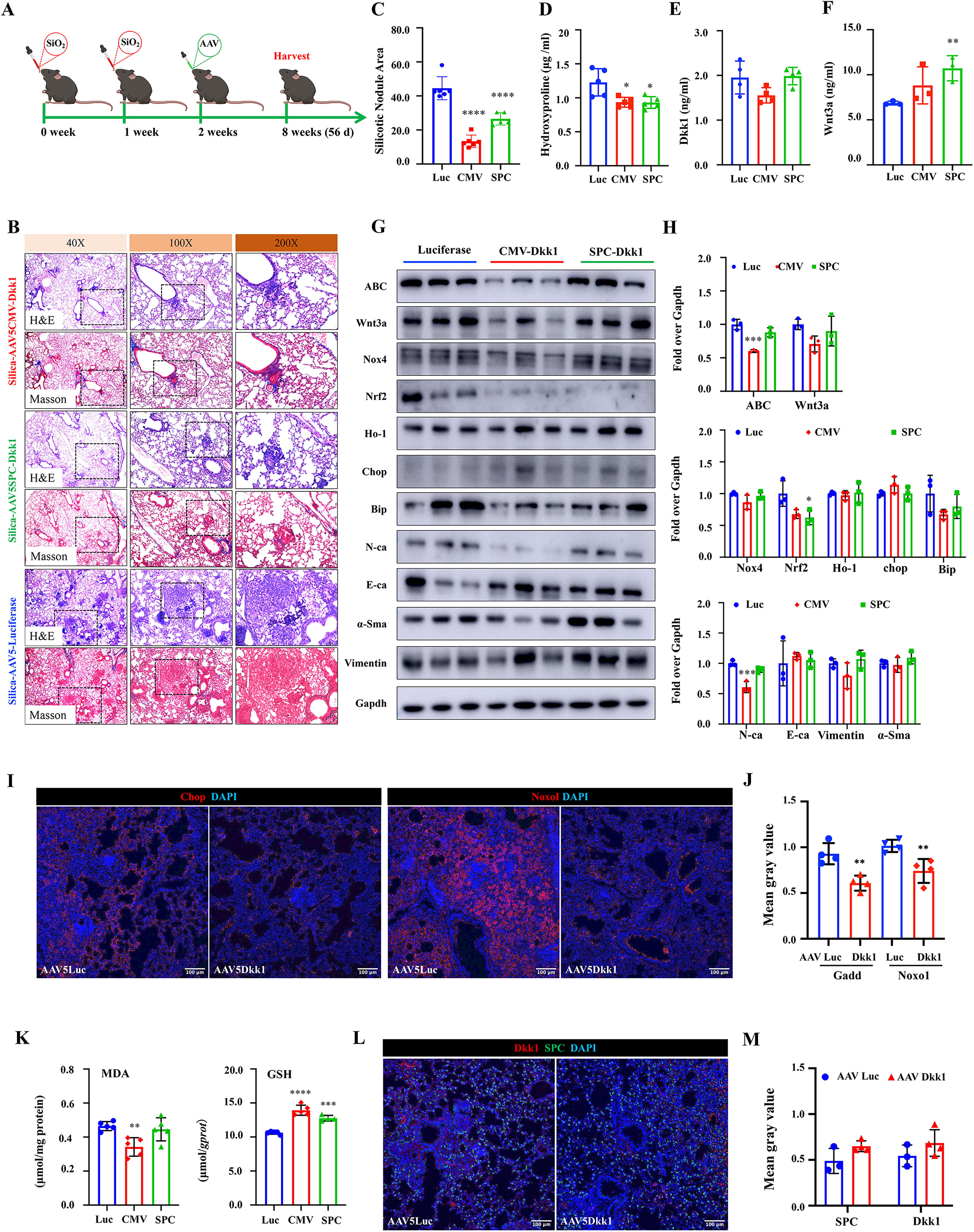
The inhibitory role of Dkk1 transduction in oxidative stress was also corroborated by accessing the abundance of Chop and NADPH oxidase organizer 1 (Noxo1) using an immunofluorescence (IF) staining assay (Fig. 2I, J) and malondialdehyde (MDA) and glutathione (GSH; Fig. 2K). Indeed, a decreased abundance of Chop, Noxo1, and MDA but an increased GSH was observed in the lung tissues of silicosis mice infected with either AAV5CMV-Dkk1 or AAV5SPC-Dkk1, compared with AAV5CMV-Luc at 6 weeks (Fig. 2I–K). Interestingly, IF staining further demonstrated the increased Dkk1 protein in the lung of mice transduced with the Dkk1 gene by AAV vectors, and part of the Dkk1 protein was colocalized in SPC-positive AT2 cells (Fig. 2L, M, Supplementary Fig. S2).
These results clearly suggest that the administration of the Dkk1 AAV vector at an early phase of silicosis (2 weeks post silica challenge) inhibits that oxidative stress restricts the development (examined at 4 weeks post the challenge) and the progression (examined at 8 weeks post the challenge) of disease. Importantly, the Dkk1-mediated inhibition of the silica-induced Wnt signaling and oxidative stress in AT2 cells could efficiently restrict the development but not the progression of silicosis.
AAV-mediated Dkk1 gene transduction mitigates disease progression in mouse lung with late phase of silicosis
Next, we sought to investigate experimental therapeutic effects of AAV Dkk1 vectors that were administrated at the late phase of silicosis disease (4 weeks post silica challenge). Mice were challenged with 3 doses of silica at 1 week interval, followed by intrapulmonary administration of AAV vectors at 4 weeks after the silica challenge, and the experiment was terminated at 8 weeks for analysis (Fig. 3A). Unlike a reduced ratio of wet lung/BW seen in AAV administration in an early phase of disease, both AAV5CMV-Dkk1 and AAV5SPC-Dkk1 vector had no impact on the weight gain (Fig. 3B) and the wet lung/BW ratio (Fig. 3C) as compared with the control vector. Histopathological analysis demonstrated the AAV-mediated Dkk1 transduction could reduce silicotic nodule formation and ECM deposition in the lungs of silicosis mice (Fig. 3D, E). However, only the AAV vector with CMV promoter-derived ubiquitous Dkk1 expression led a significantly reduced silicotic nodule area in mice evaluated at 8 weeks after the challenge, and there were no significant changes in mice infected with AAV vector with SPC promoter-derived Dkk1 expression in AT2 cells, compared with the luciferase vector (Fig. 3D). Molecular studies revealed a significantly reduced Wnt3a and ABC in the lung tissues of mice infected with AAV5CMV-Dkk1 and AAV5SPC-Dkk1 vector, compared with the control Luc vector, despite the infection of AAV5SPC-Dkk1 vector resulted in less extent reduction of ABC and Wnt3a in comparison with the AAV5CMV-Dkk1 (Fig. 3F, G). The IB assay also showed the reduction of EMT markers N-ca and α-SMA and markers of oxidative stress (Fig. 3F, G). The decrease of oxidative stress signaling molecules including Nox4, Nrf2, Ho-1, BiP, and Chop was significant in the lung tissues of silicosis mice infected with AAV5CMV-Dkk1, compared with theAAV5CMV-Luc and AAV5SPC-Dkk1 vectors (Fig. 3F, G). The Dkk1-mediated inhibition of silica-induced oxidative stress markers was further confirmed by the less abundant Chop and Noxo1 proteins in the lung tissues by immunofluorescence (IF) assay (Fig. 3H, I). In consistence with above molecular findings, IF staining also revealed less abundant α-SMA and vimentin in the lungs of silicosis mice received AAV5SPC-Dkk1 at an early phase of disease compared with that of control vector (Supplementary Fig. S3A), while the administration of AAV5SPC-Dkk1 failed to reduce α-SMA and vimentin in the lungs of silicosis mice received this vector at the late phase of disease (Supplementary Fig. S3B).
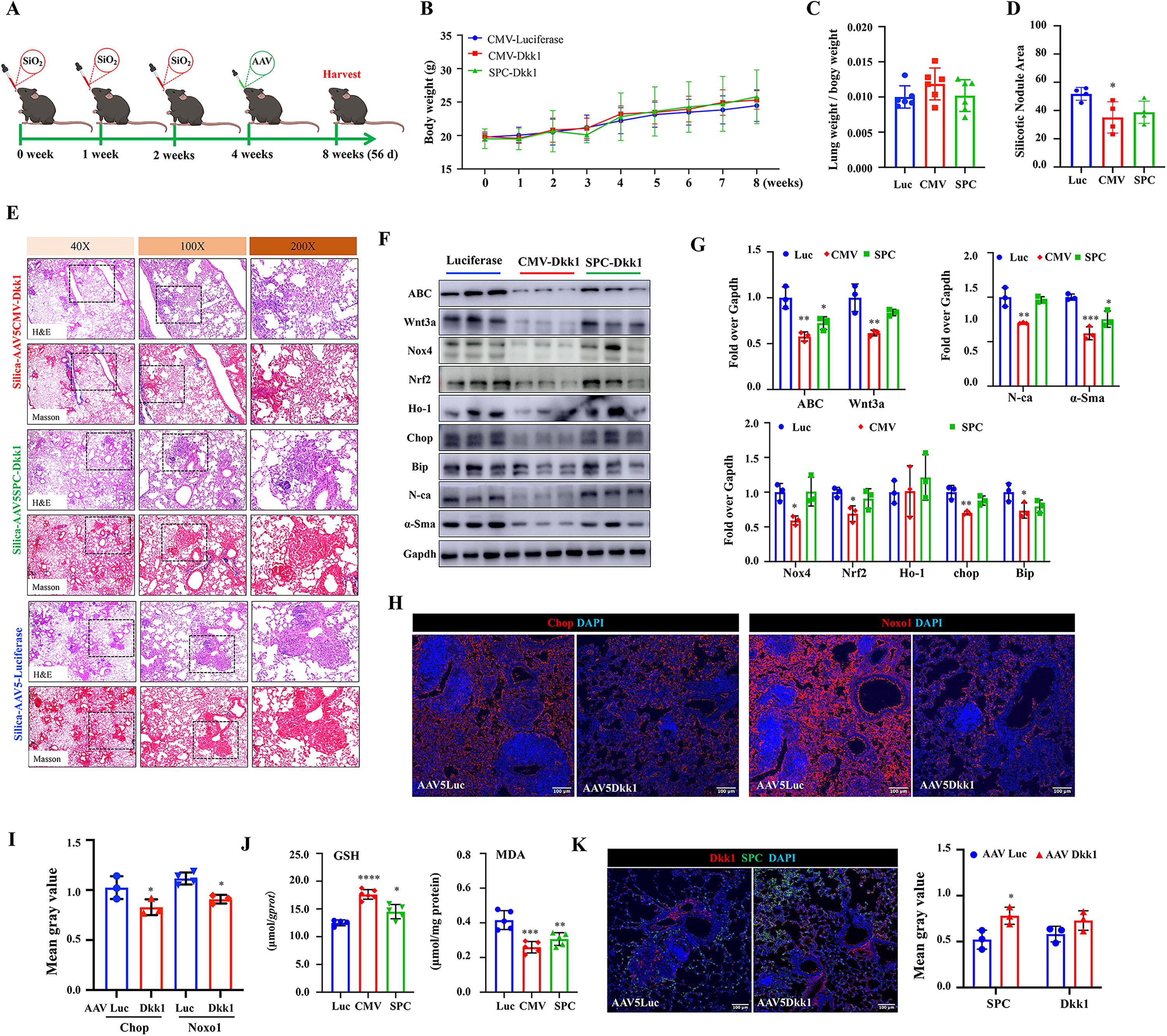
Furthermore, the significant increase of content of GSH and decrease of MDA were also observed in the lung tissues of silicosis mice infected with either AAV5CMV-Dkk1 or AAV5SPC-Dkk1, compared with AAV5CMV-Luc (Fig. 3J). In addition, the IF staining showed the increased Dkk1 protein in the lung of mice transduced with the Dkk1 gene by AAV vectors (Fig. 3K, Supplementary Fig. S2). These data indicate that the AAV-mediated Dkk1 expression at the late phase of silicosis ameliorates the disease progression at least in part by inhibiting the oxidative stress signaling activity.
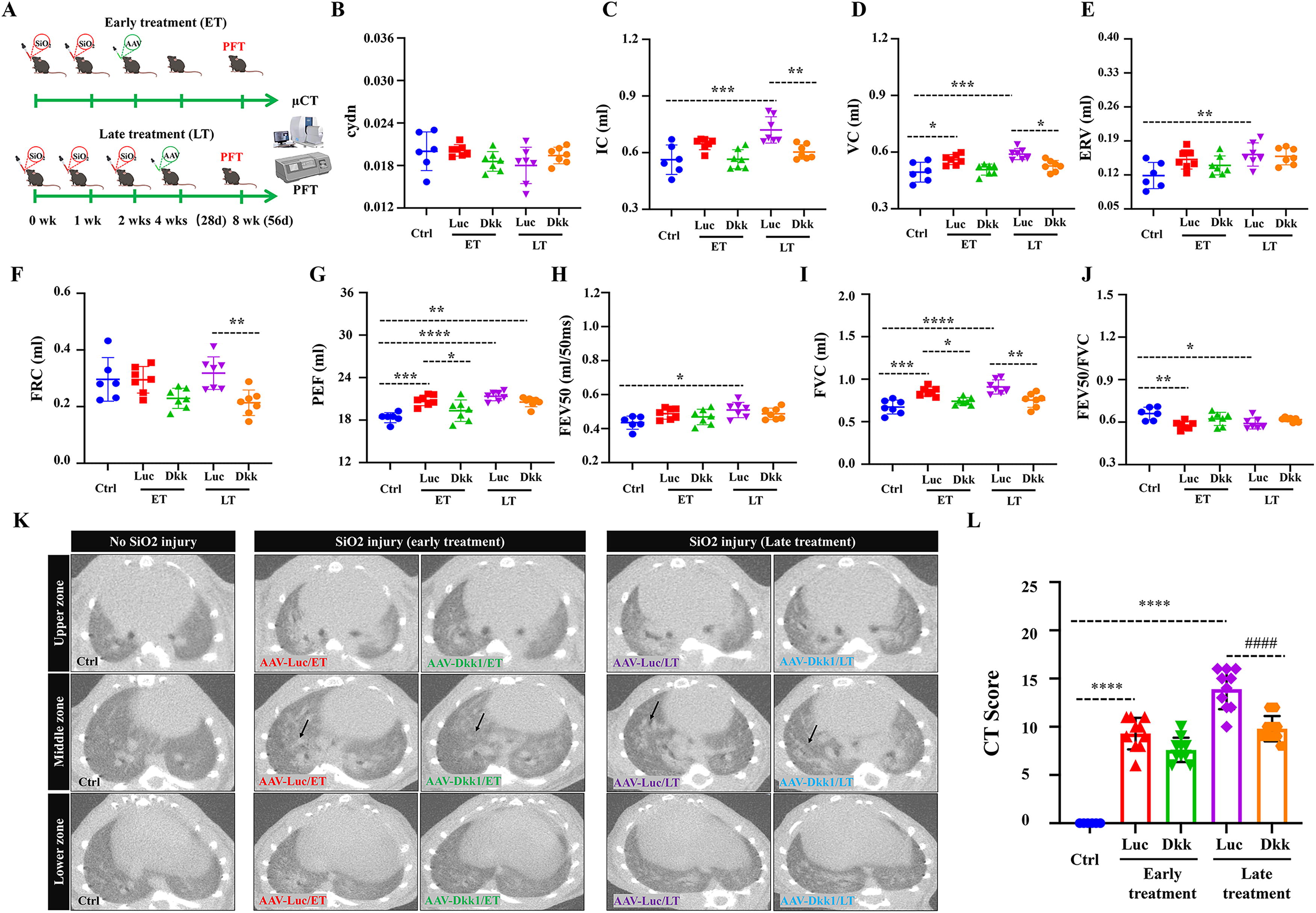
AAV-mediated Dkk1 gene transfer improves pulmonary function and microCT findings in silicosis mouse lung
To examine whether the AAV-mediated Dkk1 gene transduction could improve lung function and structure in silicosis mice, silicosis mice treated with AAV5CMV-Dkk1 and AAV5CMV-Luciferase at early phase (ET) and late phase (LT) of diseases were conducted for assessments of pulmonary function test (PFT) and microcomputed tomography (microCT) findings at 8 weeks post the first silica challenge (Fig. 4A). Consistent with the above histopathological findings, treating mice with the AAV5CMV-Dkk1 vector at the late phase of disease significantly improved the inspiratory capacity (IC) (Fig. 4C), vital capacity (VC) (Fig. 4D), functional residual capacity (FRC) testing (Fig. 4F), and forced vital capacity (FVC) (Fig. 4I), compared with the control AAV vector. While only the peak of expiratory flow (PEF) (Fig. 4G) and FVC were significantly improved in mice treated with the AAV5CMV-Dkk1 vector at an early phase of disease (Fig. 4I), there was no significant improvement of other tested parameters of lung function, such as compliance (Cdyn) (Fig. 4B), expiratory volume (ERV) (Fig. 4E), forced expiratory flow in 50 s (FEV50) (Fig. 4H), and FEV50/FVC ratio (Fig. 4J) in mice infected with the AAV5CMV-Dkk1 vector compared with the control AAV, although a trend of improvement was detected in mice transduced with Dkk1 (Fig. 4). MicroCT findings revealed that the administration of AAV5CMV-Dkk1 reduced the CT score of consolidation and reticular opacity in silicosis mice, particularly the treatment of the AAV5CMV-Dkk1 vector at the late phase of disease (LT), and significantly reduced the CT score in comparison with the AAV luciferase control vector (Fig. 4K, L). These results suggest that AAV-mediated Dkk1 gene transduction is a promising therapeutic approach for silicosis treatment.
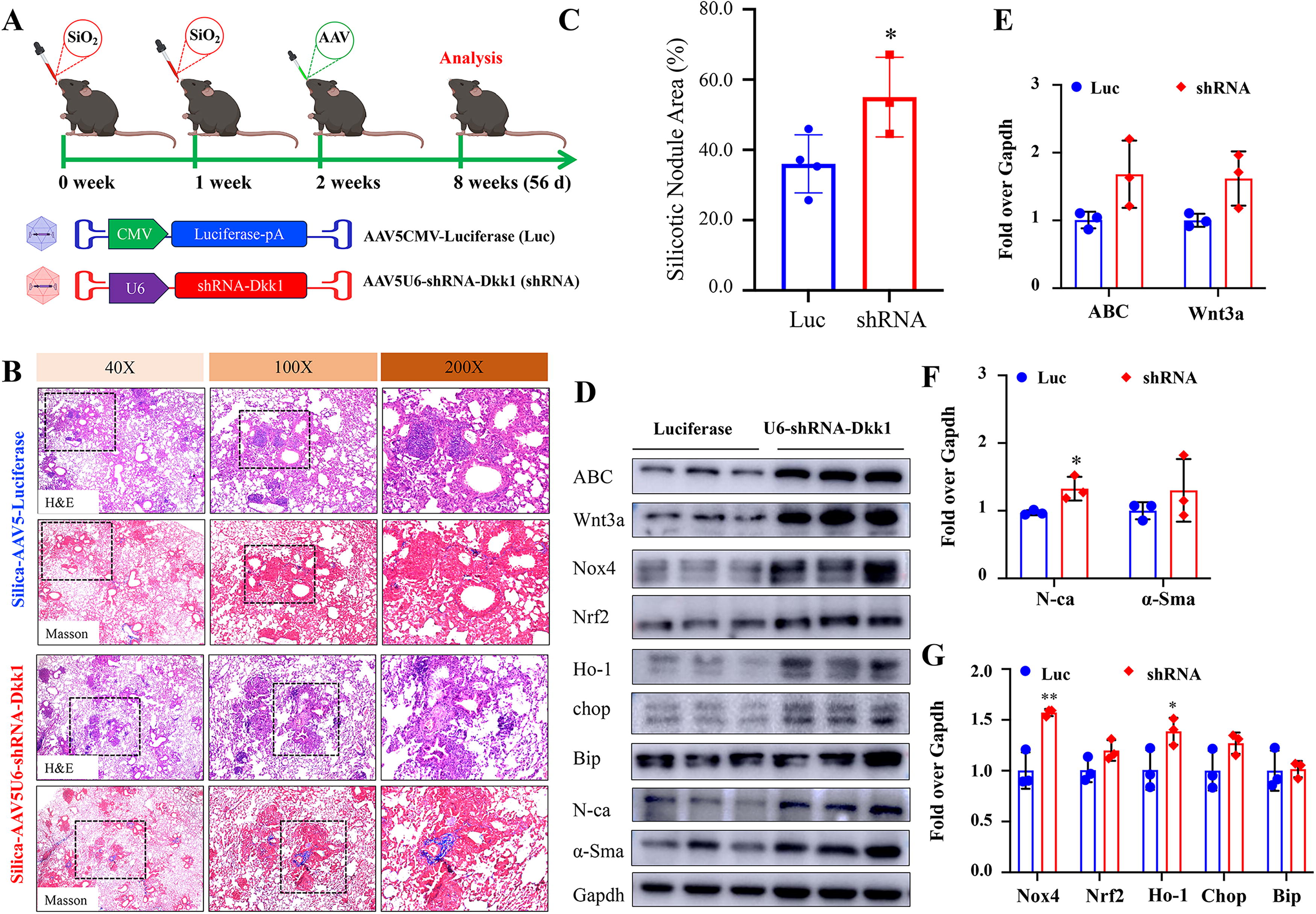
AAV-mediated shRNA knockdown of Dkk1 expression at an early disease phase exacerbates the progression of silicosis in mouse lung
We next examined the impact of knockdown of endogenous Dkk1 expression on the development and progression of silicosis in mouse lung. The knockdown of Dkk1 expression by AAV-mediated shRNA-Dkk1 (AAV5U6-shRNA-Dkk1) led to exacerbation of silicosis progression when the vector was administered at an early phase of disease (2 weeks post silica challenge) and evaluated at 8 weeks post silica challenge (Fig. 5A). The shRNA-mediated knockdown of Dkk1 resulted in histopathological changes of increased ECM deposition and silicotic nodule area in silicosis mouse lung (Fig. 5B, C), coupled with the increased abundance of Wnt signaling ligand Wnt 3a and mediator ABC, EMT marker N-ca, ROS producer Nox4, and Ho-1, a marker of oxidative stress and cellular stress responses, as compared with the control AAV vector (Fig. 5D–G). However, the knockdown of Dkk1 had no impact on the development of silicosis when the shRNA AAV vector was administered at an early phase of disease (2 weeks post silica challenge) and evaluated at 4 weeks post silica challenge, as accessed by the changes of histopathology and the abundance of signaling components of Wnt signaling, markers of EMT, and oxidative stress (Supplementary Fig. S4). Similarly, the knockdown of Dkk1 at the late phase of silicosis that was induced by 3 doses of silica challenge (4 weeks post the first challenge) did not alter the disease progression either, with no significant change of histopathology and the expression of the above-tested signaling components (Supplementary Fig. S5), although there was a trend of increase of silicotic nodule area in mice received AAV5U6-shRNA-Dkk1 relative to those given AAV5CMV-Luciferase (Supplementary Fig. S5C). These results imply that the persistent activation of Wnt signaling and oxidative stress plays significant roles in the progression of silicosis in mouse lung.
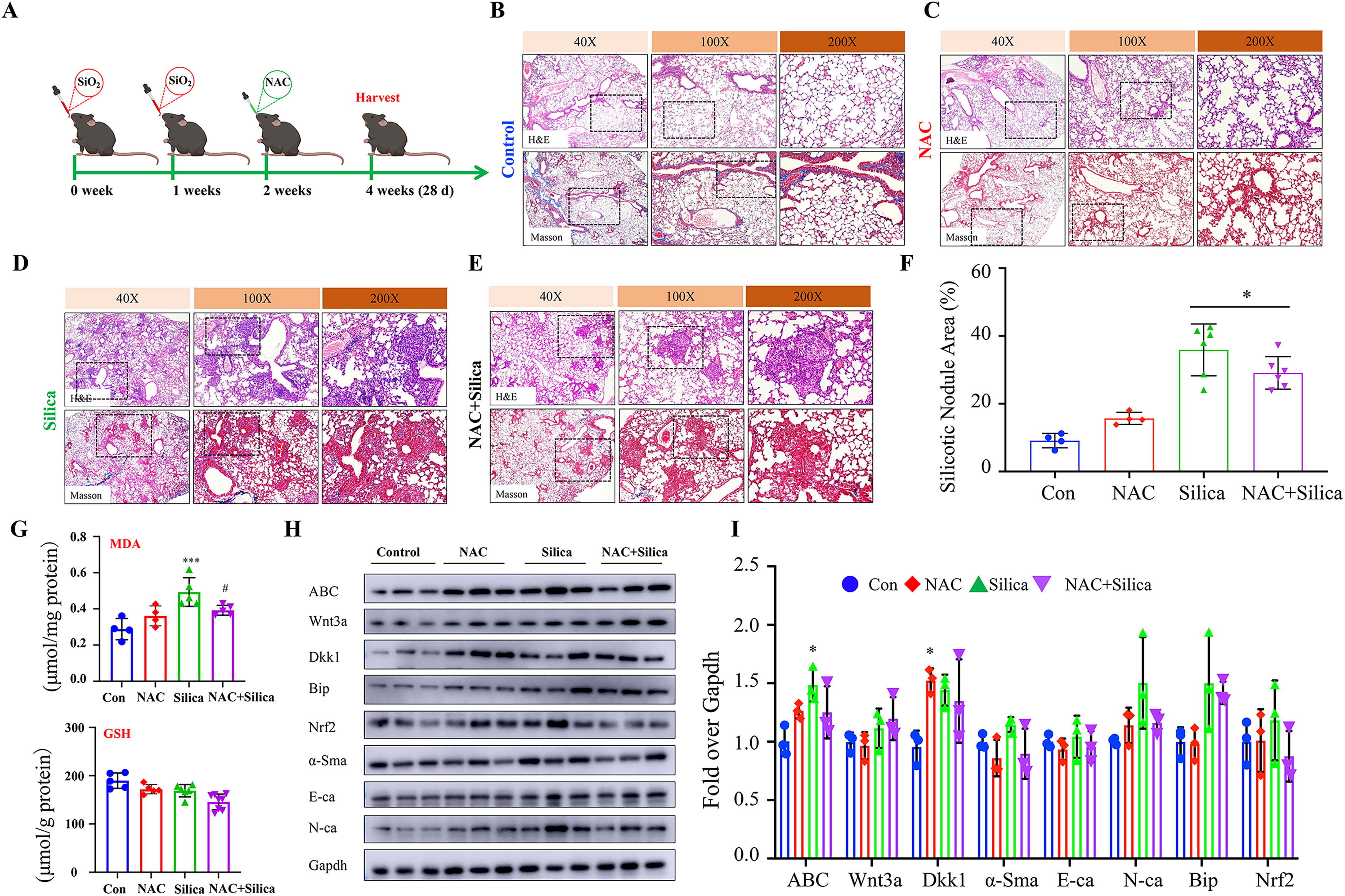
ROS scavenger n -acetyl-d -cysteine mitigates the pathogenesis of silicosis in mouse lung
To further validate the oxidative stress or ROS is a potential target of Dkk1 for the treatment of silicosis, the therapeutic effect of ROS scavenger
EMT of epithelial cells and inflammatory response of macrophages are key contributors in the pathogenesis of fibrotic diseases including pulmonary fibrosis and silicosis (Byrne et al., 2016, Malaviya et al., 2020, Moss et al., 2022). To test whether Dkk1 had impacts on silica-induced epithelial cell EMT and macrophage inflammation, the changes of markers of EMT, myofibroblast, and inflammatory signaling were evaluated in human epithelial cell line A549 cells and RAW264.7 mouse macrophage-like cells exposed to silica particles and/or recombinant DKK1 protein. To further demonstrate the antioxidative effect of DKK1 protein in cells in response to silica stimulation, NAC was served as an antioxidant control. Indeed, the addition of DKK1 protein significantly inhibited the silica-induced α-SMA and N-ca and oxidative stress marker BiP in A549 cells. The degree of action was comparable to that of NAC as determined by IB assay (Supplementary Fig. S6A). The IF assay also showed both DKK1 and NAC were able to reduce the silica-induced Chop and BiP in A549 epithelial cells (Supplementary Fig. S6B). As expected, DKK1 and NAC also exhibited the potential to reduce silica-activated inflammatory signaling by inhibiting NF-κB-p65 and oxidative stress in murine macrophage-like Raw264.7 cells (Supplementary Fig. S6C, S6D). As expected, DKK1 and NAC could restore the silica-inhibited GSH production and inhibit the silica-induced ROS generation in both A549 and RAW264.7 cells (Supplementary Fig. S6E). These results suggest that DKK1 inhibits silica-induced oxidative stress, inflammation, and EMT in the lung epithelial cells and macrophages.
Discussion
Pulmonary silicosis is an irreversible lung disease caused by the excessive inhalation of silica particles (SiO2), which is characterized by silicotic nodule that is the anatomical hallmark with well-demarcated, rounded fibrotic lesions concentrated in the upper lung lobes (Handra et al., 2023). Wnt/β-catenin signaling is reactivated in both human IPF (Chilosi et al., 2003; Lam et al., 2014), while abundance of both Dkk1 and Wnt3a is elevated in the plasmas and lungs of silicosis patients and experimental silicosis (Cai et al., 2022; Dai et al., 2016; Ma et al., 2020; Wang et al., 2015); therefore, Wnt/β-catenin signaling antagonists or inhibitors are considered as promising targets for the treatment of fibrotic diseases such as IPF (Henderson et al., 2010; Nabhan et al., 2023; Villanueva, 2023) and silicosis (Cai et al., 2022; Dai et al., 2016; Wang et al., 2022).
DKK1 is a well-characterized Wnt inhibitor and has been correlated with chronic pulmonary diseases including IPF and chronic obstructive pulmonary disease (Dai et al., 2022; Pfaff et al., 2011). Our previous proof-of-concept study of intrapulmonary mouse Dkk1 gene transfer using adenoviral vector (AdDkk1) demonstrated the potential mechanism of Dkk1 in preventing the development and progression of disease in silica-induced silicosis mice, in part through a mechanism by which Dkk1 inhibited the silica-activated Wnt/β-catenin and transforming growth factor-β (TGF-β) signaling, a well-known signaling pathway driving fibrotic diseases (Cai et al., 2022), and NOX4 (Ma et al., 2020), a main inducible source of ROS production (Lee et al., 2010). However, the underlying mechanism between the Wnt signaling and TGF-β or Nox4 of the therapeutic effect of Dkk1 in silicosis mice remains elusive. Given the facts of disadvantages of adenoviral vectors and advantages of AAV for gene therapy from clinical standpoints (Carneiro et al., 2020; Zhang et al., 2023), as well as the involvement of other mechanisms such as oxidative stress in the pathogenesis of silicosis (Vallyathan et al., 1997; Yao et al., 2023), we interrogated the antioxidative effect and mechanism of AAV-mediated Dkk1 gene transfer (AAV-Dkk1) in ameliorating the pathogenesis of silicosis in mouse. Our results demonstrated that intrapulmonary delivery of AAV-Dkk1 significantly alleviated the development and progression of silicosis, coupled with the inhibition of Wnt signaling and reduction of silica-induced oxidative stress and the improvement of lung function and CT findings in silicosis mice. This study not only provides additional evidence of potential of targeting Wnt inhibitor DKK1 in the treatment of silicosis using AAV-mediated DKK1 gene transfer, but also increases our understanding the mechanism by which interactions among the Wnt signaling, oxidative stress and TGF-β in the pathogenesis of silicosis.
Wnt/β-catenin signaling is reactivated in IPF and silicosis lungs of both patients and experimental animal models (Cai et al., 2022; Chilosi et al., 2003). The aberrant activation of Wnt signaling can result in undesirable effects in the homeostasis of the lung. Targeting the Wnt signaling thus becomes an attractive option in developing therapeutic agents for the treatment of fibrotic diseases such as IPF and silicosis (Cai et al., 2022; Dai et al., 2016; Villanueva, 2023; Wang et al., 2015). Despite the Wnt activation may have beneficial effects on alveolar epithelial repair following injury during fibrogenesis in distal lung, the persistent or chronic activation of Wnt signaling in alveolar tissue may lead epithelial stem cell exhaustion or senescence (Lehmann et al., 2020; Yang et al., 2022), subsequentially drives fibroblast expansion and differentiation to myofibroblasts, a hallmark of fibrosis in silicosis and IPF lungs (Cao et al., 2018, Famili et al., 2016; Sun et al., 2014).
Subsets of AT2 cells have been identified as stem cells responsible for maintaining alveolar epithelial homeostasis and alveolar epithelial regeneration, and Wnt signaling is required for self-renewal and differentiation (Nabhan et al., 2018). Interestingly, the Wnt activation is different among epithelial, mesenchymal, and endothelial cells in alveolar tissues, and the balance of Wnt activation in distinct cell types is thus critical to normal lung injury repair (Nabhan et al., 2018; Nabhan et al., 2023). For example, AT2 cells require Wnt activation cell proliferation, but the activation in fibroblast can drive fibroblast pathological expansion and myofibroblast differentiation in alveolar space (Cao et al., 2018; Nabhan et al., 2023; Sun et al., 2014). Therefore, the selective activation of Wnt signaling in AT2 cells but inhibition in mesenchymal cells by modulating cell type-specific Wnt ligand receptor frizzled (FZD) could ameliorate fibrogenesis in IPF mouse lung (Nabhan et al., 2023; Villanueva, 2023).
Given the importance of Wnt activation in stem cell self-renewal and tissue regeneration, and potential side effects that caused by broadly targeting Wnt signaling in the lung with AAV5CMV-Dkk1 vectors, AAV5SPC-Dkk1 vector was employed to test the experimental therapeutic effect of Dkk1-targeted Wnt signaling in AT2 cells in this report. The AAV5SPC-Dkk1 vector was able to tissue specifically transduce Dkk1 expression in AT2 cells. When AAV vectors were administrated at an early phase of silicosis development (2 weeks post the silica challenge), both AAV5SPC-Dkk1 and AAV5CMV-Dkk1, a vector that using CMV promoter to derive the ubiquitous expression of Dkk1 in every transduced cell, could significantly ameliorate histopathological changes in the lungs (Figs 7,1). This finding was in concurrence with an exacerbation of progression of silicosis in mice that were shRNA-mediated inhibition of endogenous Dkk1 (Fig. 5). While the vector was administrated at the late phase of disease (4 weeks post the silica challenge when the silicosis was well established), only the administration of AAV5CMV-Dkk1 but not AAV5SPC-Dkk1 showed the ability to significantly mitigate the progression of silicosis in mice (Fig. 3). These results imply that the inhibition of aberrant Wnt activation in AT2 cells at the early or initial stage of disease can restrict silicosis development and subsequently inhibit the disease progression. However, in the late stage of silicosis, the potency of AT2 stem cells is exhausted during the disease progression, which most likely was caused by the persistent Wnt activation (Lehmann et al., 2020). In this context, the transduction of Dkk1 may have a negative impact on the stemness restoring in AT2 cells, as the Wnt signaling is blocked by Dkk1. This may explain that the transduction of AAV5SPC-Dkk1 and AAV5U6shRNA-Dkk1 failed to alter the pathological progression of silicosis lung when it was administrated at the late phase of disease. Unlike SPC promoter, CMV is a ubiquitous promoter able to derive Dkk1 expression in any AAV5CMV-Dkk1-transduced cells, including fibroblasts and endothelial cells, in which Dkk1 inhibited Wnt activation and consequently inhibited fibroblast proliferation and differentiation to myofibroblast. This may explain the mitigation of silicosis pathogenesis introduced by the administration of AAV5CMV-Dkk1 at the late stage of disease in this study (Fig. 3) and the ineffectiveness of the shRNA-mediated downregulation of Dkk1 and transduction of AAV5SPC-Dkk1 at the late stage of silicosis (Figs. 4–5).
Wnt/β-catenin signaling has been closely linked to oxidative stress during the development and progression of many diseases (Dong et al., 2021; Karimaian et al., 2017; Ma et al., 2023; Zhou et al., 2019). In this regard, oxidative stress has been recognized as a critical molecular process of fibrosis, and thus, it is a promising therapeutic target of fibrotic diseases (Estornut et al., 2021, Kurt et al., 2023, Li et al., 2022, Mokra et al., 2023; Roksandic Milenkovic et al., 2022; Yao et al., 2023). In this context, oxidative stress is a trigger of Wnt/β-catenin signaling pathway (Fang et al., 2023; Karimaian et al., 2017), while the activation of Wnt signaling in turn induces oxidative stress and inflammation (Dong et al., 2021; Vikram et al., 2014), forming a positive feedback loop to promote EMT and disease progression (Ma et al., 2020). Herein, targeting Wnt signaling may resolve oxidative stress that is induced by the activation of Wnt signaling.
Accumulating evidences reveal that silica particles induce oxidative stress, which plays a key role in the pathogenesis of silicosis in experimental animals (Barsan et al., 2021; Ma et al., 2023) and silicosis patients (Kurt et al., 2023). We previously also found the exposure of silica dust could induce ER-stress and ferroptosis in silicosis mouse lung in vivo and macrophages in vitro (Ma et al., 2023). In the present study, inhibiting Wnt/β-catenin signaling by AAV-mediated Dkk1 gene transduction reduced the expression of oxidative stress signaling components Ho-1, Chop and BiP, and ROS production but increased the levels of ferroptosis inhibitor Gpx4 and GSH in silicosis mouse lung. Indeed, targeting oxidative stress and ROS product by intrapulmonary delivery of ROS scavenger NAC significantly alleviated the silica-induced silicosis in mice (Huang et al., 2019; Li et al., 2021; Fig. 6). These data strongly suggest that the Dkk1 mitigated the pathogenesis in silicosis mice was in part by inhibiting the silica-induced oxidative stress and ROS production in mouse lung.
AAV is currently the most promising viral vector in gene therapy for pulmonary diseases, owing to its advantages in safety of low immune response and long-term of gene expression (McLachlan et al., 2022). Three AAV vectors, Luxturna® (Spark Therapeutics, Inc.) for individuals with an inherited retinal dystrophy, Zolgensma® (Novartis AG) for the treatment of spinal muscular atrophy in pediatric patients <2 years old, and Hemgenix® (CSL Behring LLC) for the treatment of hemophilia B, have been granted by the U.S. Food and Drug Administration (Srivastava, 2023). However, no AAV “drug” has been approved for pulmonary diseases in clinical practice, although AAV-mediated cystic fibrosis transmembrane conductance regulator gene therapy for cystic fibrosis disease has been tested in clinic trials since 1996 (Flotte et al., 1996, Guggino and Cebotaru, 2020). Many facts may contribute the difficulty of gene transduction in the lung using viral vectors. For instance, the physical and chemical barriers of lung structure and diversified cell composition lead to inefficient viral vector delivery in airways, and the natural immune protection against external viruses may also elicit inflammatory response to viral vectors. Moreover, increased mucus production, cough clearance, and proinflammatory base state are often existed in many pulmonary disease states, which can exacerbate the difficulty of pulmonary viral gene delivery in practices (Banuls et al., 2020; McCarron et al., 2023). However, in contrast, lung is an organ constantly exposed to the external environment for gas exchanges, which increases the accessibility of lung for direct local gene delivery through airway routes. The local targeted delivery has many advantages, including increase of dose control and local retention of vector, minimum of invasive procedure, and rapid onset transgene expression (Katz et al., 2019). In this report, AAV vectors were delivered through intratracheal route to silicosis mouse lung, and efficient Dkk1 gene transduction was observed by IB and IF assays. Furthermore, the AAV-mediated Dkk1 gene transfer led to a significant mitigation of silicosis with an improvement of lung function in silica-injured mouse lungs, suggesting that targeting Wnt signaling with DKK1 using AAV-mediated gene transfer is a promising approach for the treatment of silicosis.
Conclusion
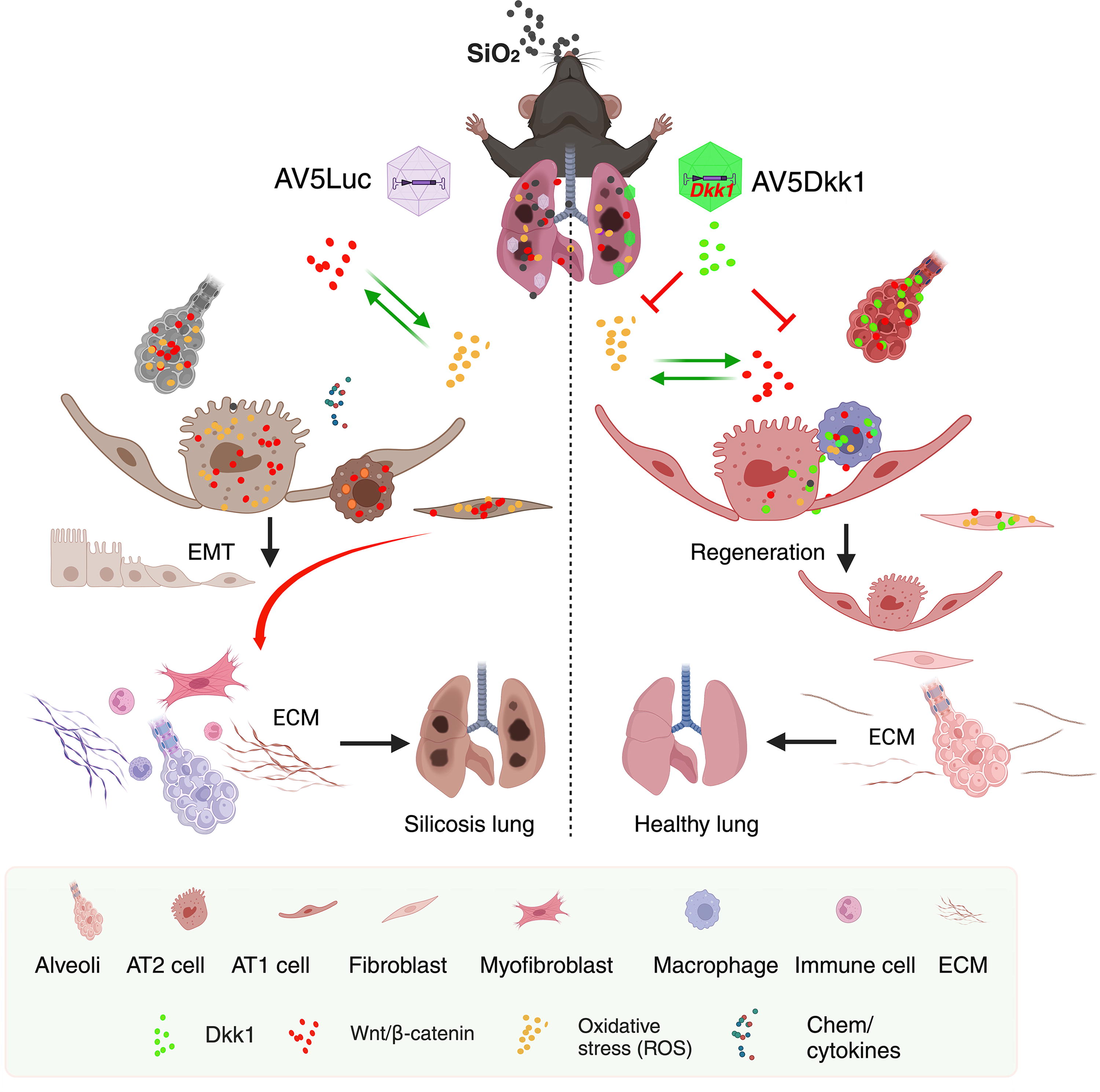
AAV-mediated Dkk1 gene transfer could alleviate the development and progression of silicosis, whereas tissue-specific expression of Dkk1 in AT2 cells showed functional effect of restriction on silicosis mice at an early disease state. Mechanical study further demonstrated that targeting Wnt signaling with Dkk1 could reduce the silica-induced ROS production and oxidative stress, which subsequently mitigate the development and progression of silicosis in the lung (Fig. 7).
Materials and Methods
Preparation of silica (SiO2) particles
Silica dioxide (∼99% SiO2, 0.5–10 µm particle size, S5631) and silica (SiO2, mesoporous, 2 µm particle size, 806587) were from Sigma-Aldrich. Silica particles were baked at 200°C for 2 h for removing endotoxin prior to being suspended in saline at a concentration of 50 mg/mL. The silica solution was dispersed for 15 min in a water bath solicitor before the use (Cai et al., 2022; Ma et al., 2023; Yang et al., 2022).
Generation of silica-induced silicosis mouse model and intrapulmonary AAV infection
All animal protocols approved using Laboratory Animal Committee of College of Life Science at Ningxia University, in accordance with guidelines of the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NXULS20180123–3). The 6- to 8-week-old male and female C57BL/6N mice were purchased from Beijing Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). Serotype 5 AAV vectors (AAV5) were used for transgene delivery in mouse lung. AAV5CMV-luciferase (Luc) was an AAV vector contained a CMV promoter-derived luciferase gene; AAV5CMV-Dkk1 (CMV) was an AAV vector contained a CMV promoter-derived murine Dkk1 gene; AAV5SPC-Dkk1 (SPC) was an AAV vector contained a mouse surfactant protein C (SPC) promoter-derived murine Dkk1 gene; and AAV5U6-shRNA-Dkk1 was an AAV vector contained a mouse U6 Pol III promoters driven targeting sequence (GAACCACACTGACTTCAAA) of murine Dkk1 cDNA (GenBank: JN966751.1). All AAV vectors were produced by Shanghai GeneChemCo. Ltd. To generate silicosis mouse model, mice were challenged with 2 or 3 doses of silica dust (50 µL of 50 mg/mL in saline) at one week interval by intratracheal instillation (Cai et al., 2022; Ma et al., 2023). For intrapulmonary AAV infection, silicosis mice were intratracheally instilled 25 µL AAV5 vectors (containing 1 × 1011 viral particle/mouse) containing 200 µmol of doxorubicin (Sigma) at 14 or 28 days post the first dose of silica challenge as previously described (Liu et al., 2009). The lung tissues were harvested at 28 or 56 days post the challenge. The reagents used in this study are listed in Supplementary Table S1.
Measuring Wnt3a and Dkk1 protein concentration
The concentration of Wnt3a and Dkk1 proteins in mouse serum was measured by commercial enzyme-linked immunosorbent assay kits per manufacturer’s instructions from Shanghai MLBIO Biotechnology. The concentration of those proteins was calculated according to the standard curve and presented as nanograms per milliliter. All experiments were technically triplicated for the sample from each mouse.
Hydroxyproline content in the lung tissues
In total, 50 mg of mouse lung tissue was homogenized for total protein isolation according to the experiment protocol. The supernatant of collected lysates from each mouse lung was triplicated for hydroxyproline measurement. Hydroxyproline content was measured by commercially available hydroxyproline assay kit (Shanghai Jiang Lai Biology Institute) according to the manufacturer’s instructions. The hydroxyproline content was presented as microgram (µg) per milligrams total protein of wet lung tissues.
Lung histopathological analysis
Mouse lung tissues were fixed in 10% neutral-buffered formalin solution before they were processed for paraffin embedding, section, and staining. Sections of 5-µm thickness were used for lung histopathological analysis by hematoxylin and eosin (H&E) staining and Masson’s trichrome staining. All images were captured by Olympus optical microscope (Olympus TH4-200). Fibrotic (silicotic nodule) area of H&E images was quantified by using identical threshold settings FIJI Image J software. The percentage and size of silicotic nodules relative to the entire lung lobe were calculated (Cai et al., 2022).
Measurement of MDA
Mouse lung tissues were lysed in RIPA lysis buffer and centrifuged at 12,000 rpm for 20 min at 4°C, and the resultant supernatants were collected. The supernatant was subjected to the protein contents for the measurement of MDA. The content of MDA of lung of mouse was measured using an MDA assay kit (Beyotime). Briefly, the above supernatants were mixed with thiobarbituric acid and boiled for 15 min, prior to being further centrifuged at 10,000 g for 20 min to remove debris, and the supernatants containing MDA product were read the absorbance of 532 nm. The concentration was calculated by comparison with the standard curve generated with MDA provided from the kit according to the manufacturer’s instruction.
Measurement of reduced GSH
Mouse lung tissues were lysed with RIPA lysis buffer, cell cultures were lysed with 1% TritonX-100 in PBS for 30 min at 4°C before they were centrifuged at 12000 rpm for 20 min at 4°C, and the supernatants were collected as total protein for analysis. A total of 100 μL supernatant was employed for measuring GSH using GSH Assay kit by measuring optical density at 405 nm (OD405nm) value per the manufacturer’s manual (Jiancheng Institute of Biotechnology). The reduced GSH was normalized by protein concentrations. All experiments were performed with biological triplicates, and data are representative of at least three independent experiments.
Measurement of ROS
Cells cultured in 96-well plate before the intercellular ROS was assessed by a ROS Assay Kit (Beyotime). Briefly, the cells were first incubated with phenol red-free DMEM containing 10 μM 2′,7′-Dichlorofluorescindiacetate (DCFH-DA) in dark at 37°C for 30 min, the reagent solution was then removed, and the cells were washed with PBS for three times before 100 µL of PBS was added in the well. The intercellular ROS was ascertained by reading the value of OD488–525nm according to the manufacturer’s instruction. All experiments were performed with biological triplicates, and data were representative of at least three independent experiments.
Cell culture and treatments
Human lung epithelial cell line A549 cells and mouse mononuclear macrophage-like cell line RAW264.7 cells were purchased from American Type Culture Collection. A549 cells were cultured in high-glucose DMEM basal medium (HyClone) supplemented with 10% fetal bovine Serum (Gibco), penicillin (1000 IU/mL), and streptomycin (1000 IU/mL). RAW264.7 cells were cultured in high-glucose DMEM basal medium supplemented with 10% Newborn Calf Serum (Gibco, USA), penicillin (1000 IU/mL), and streptomycin (1000 IU/mL). Cells were cultured in incubator at 37°C with 5% CO2. Cells were treated with silica particle at a concentration of 100 µg/cm2 (A549 cells) or 50 µg/cm2 (RAW264.7) (Cai et al., 2022; Ma et al., 2023), in the presence or absence of 100 ng/mL of recombinant human DKK1 protein (DK1-H5221, ACRO Biosystems) or 10 mmol/L ROS scavenger NAC (Sigma) for 48 h (A549 cells) or 12 h (RAW264.7 cells).
IF staining
Mouse lung tissues and cell lines were fixed in 4% paraformaldehyde solution and embedded in optimal cutting temperature (OCT) compound. The IF staining was performed using OCT sections or cell culture chamber slides. Sections/slides were air-dried and permeabilized in 0.2% Triton X-100/PBS solution for 30 min at room temperature (RT), prior to being blocked by 5% donkey serum in PBS for at least 2 h. The blocked sections/slides were probed with primary antibodies to proteins of interest in 1% donkey serum dilutant buffer at 4°C for overnight. Sections were washed three times for 5 min in PBS and incubated with appropriate fluorescence-labeled secondary antibodies in 1% donkey serum dilutant buffer for 2 h at RT. Finally, the slides were washed three times and mounted with medium contain DAPI. The images acquired by Zesis 880 Confocal Microscope and processed by using FIJI Image J software (http://rsb.info.nih.gov/ij/).
Immunohistochemical staining
For IHC staining, paraffin sections were deparaffinized and rehydrated. Following antigen retrieval in citrate buffer, sections were blocked by 5% donkey serum in PBS for at least 2 h before being probed with primary antibodies in 1% donkey serum dilutant buffer at 4°C overnight. Sections were then washed three times with PBS-tween 20 (0.1%) and then incubated with biotin- or horseradish peroxidase (HRP)-conjugated secondary antibodies for 30 min at RT. The antibody binding was then developed with DAB substrate (Zsbio) for HRP-conjugated secondary antibodies or incubated with an Avidin-Biotin Complex Staining Kit (ABC Kit; Vector Laboratories) per the manufacturer’s instructions. Slides were then counterstained with hematoxylin, dehydrated, mounted, and finally visualized under an upright microscope.
Western blotting analysis
Mouse lung tissues were lysed in RIPA lysis buffer and centrifuged at 12000 rpm for 20 min at 4°C, and the resulted supernatants were collected as total protein for IB assay. About 30 µg proteins were resolved by 8%−12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. Membranes were blocked in 5% nonfat milk in Tris-buffered saline (pH 7.4) for 2 h at RT and probed with antibody to protein of interest. The binding of antibody was developed by using enhanced chemiluminescence reagent (Advansta). The relative expression of protein was semi-quantified by optical densitometry using Image J Software. The ratio between the net intensity of each sample was normalized by the GAPDH internal loading control and calculated as a densitometric arbitrary unit (AU). The AU was used as an index of the relative expression of the protein of interest. The information of primary antibodies used in the present study is listed in Supplementary Table S2.
Mouse PFT and microCT
The 6- to 8-week-old male and female C57BL/6N mice (n = 10) were obtained from Guangzhou Medical University. The pulmonary function of mice was measured by an automated system to compute spirometry-like parameters (Buxco PFT, Data Sciences International). Both intubation and tracheostomy were performed with an 18G intravenous catheter. Mouse was first anesthetized by inhaling isoflurane and mounted on supine position in a scanning chamber. The mouse scanned by microCT instrument (50 µm voxel size) using a Super Nova CT instrument (Super Nova CT, PINGSENG Healthcare, SNC-100). The algorithms of the axial and coronal images of mouse lung were reconstructed and analyzed according to the manufacturer’s instructions with an AVATAR 1.5.0 three-dimensional finite element (PINGSENG Healthcare). The severity of the image findings was semi-quantified with an assessment of the following criteria according to previous studies (Choi et al., 2014; Lee et al., 2008). Lesions including honeycombing, reticular opacity and consolidation, and ground-glass opacity were measured at left and right upper, middle, and lower levels. The severity of image findings was further semi-quantitatively scored on a scale from 0 to 3 (0 = finding was absent, 1 = single scattered changes, 2 = larger single changes or several minor changes, and 3 = uniform or substantial changes) and presented as the sum of scores of left and right lungs of 3 levels of above 3 categories of lesion, according to methods previously documented (Choi et al., 2014; Lee et al., 2008, Nurmi et al., 2018). After the microCT scanning procedure, the mouse was further deep anesthetized by administration of ketamine/xylazine, intubated, and connected to a ventilator for testing the airflow and pressure during a succession of forced pulmonary tests to generate spirometry-like parameters. The weight and age correlated positively with forced expiratory flow in 50 s (FEV 50) were recorded. Moreover, PEF, FVC, IC, tidal volume, compliance, VC, FRC testing, expiratory volume, and total lung capacity were also recorded.
Statistical analysis
All experiments were performed with biological triplicates, and data represent at least three independently performed experiments. Data are presented as mean ± standard deviation. All analyses were assessed using GraphPad Prism version 8 software (GraphPad Software Inc.). Statistical difference was analyzed by the one-way analysis of variance (ANOVA) or two-way ANOVA comparisons. p-Values of <0.05 were considered statistically significant.
Footnotes
Authors’ Contributions
X.L., J.C., and J.Y. conceived and designed the experiments; J.M., J.W., R.S., Z.W., Z.H., J.X., S.W., J.Y., and W.H. conducted the experiments and acquired data; J.M., J.W., L.Y., and Q.C. collected and analyzed data; J.M. and J.W. drafted the article; X.L., J.C., and J.Y. interpreted data and critically revised the article. All authors read and approved the final version of the article.
Ethics Statements
Studies involved with the live vertebrate animals were carried out following protocols approved by the Research Ethics Committee of the Institutional Animal Care and Use Committee of Ethics Committee at College of Life Science of Ningxia University (NXULS20180123-3). All animal protocols were adhered to the National Institutes of Health’s requirements for activities involving the care and use of animals and in compliance with the ARRIVE guidelines.
Data Availability Statement
The data generated during this study can be found within the published article and its supplementary files, and additional data are available from the corresponding author on reasonable request.
Author Disclosure Statement
The authors declare there are no competing financial interests in relation to the work described in this study.
Funding Information
This work was financially supported by grants from
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Table S1
Supplementary Table S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.