
Abstract
Significance:
Reduction–oxidation (redox) regulation is an important biological phenomenon that provides a balance between antioxidants and the generation of reactive oxygen species and reactive nitrogen species under pathophysiological conditions. Structural and functional changes in glycans are also important as post-translational modifications of proteins. The integration of glycobiology and redox biology, called glyco-redox has provided new insights into the mechanisms of epithelial–mesenchymal transition (EMT)/mesenchymal–epithelial transition (MET), cancer, and various diseases including Alzheimer’s disease, chronic obstructive lung disease, type 2 diabetes, interstitial pneumonitis, and ulcerative colitis.
Recent Advances:
Glycans are biosynthesized by specific glycosyltransferases and each glycosyltransferase is either directly or indirectly regulated by oxidative stress and redox regulation. A typical example of glyco-redox is the role of N-glycan referred to as core fucose in superoxide dismutase 3. This glycan was found to be involved in the growth inhibition of cancer cell lines.
Critical Issues:
The significance of glyco-redox in EMT/MET, cancer, and various diseases was found in major N-glycan branching glycosyltransferases β1,4N-acetylglucosaminyltransferase III, β1,4N-acetylglucosaminyltransferase IV, β1,6N-acetylglucosaminyltransferase V, β1,4-acetylglucosaminyltransfearfse VI, β1,6-acetylglucosaminyltransferase IX, α-1,6 fucosyltransferase, and β-galactoside α-2,6-sialyltransferase 1. Herein, we summarize previous reports on target proteins and how this relates to oxidative stress. We also discuss the products of these processes and their significance to cancer and various diseases.
Future Direction:
A clear-cut understanding of the significance of glyco-redox in relation to prevention, diagnosis, and therapeutics is required. These studies will open a new road toward glycobiology and redox biology. Antioxid. Redox Signal. 41, 910–926.
Introduction
Our group has reported the purification and gene cloning of many glycosyltransferases involved in the biosynthesis of major N-glycan branching. Before explaining the significance of glyco-redox, pathophysiological changes of glycans in relation to oxidative stress will be described. Understanding these changes involves determining the structures and functions of these enzymes in relation to their pathophysiological significance (Takamiya et al., 2021; Taniguchi et al., 2016). Similar reports have recently provided more convincing evidence (Khoder-Agha and Kietzmann, 2021; Lim et al., 2022). Our group previously reported many redox-related studies on glutathione-related enzymes γ-glutamyl transpeptidase (γ-GTP) (Ikeda and Taniguchi, 2005), glutathione peroxidase (Fujii and Taniguchi, 1999), glutathione reductase, and nitric oxide synthases as well as studies focused on related enzymes (Kaneto et al., 1995), superoxide dismutase (SOD) (Taniguchi, 1992), and thioredoxin reductase (Fujiwara et al., 1999). Based on our experiences both in glycosylation and redox research, the present review is focused on the interplay with glycans and redox regulation against oxidative stress and also on the significance of the epithelial–mesenchymal transition/mesenchymal–epithelial transition (EMT/MET) processes and cancer in relation to N-glycan branching glycosyltransferases and their enzymatic products (Takahashi et al., 2022; Takahashi et al., 2016b; Taniguchi et al., 2021).
Glycosyltransferases of N-Glycan Branching and Their Biosynthesized Glycans: Structure and Function
The involvement of glycosyltransferases in the biosynthesis of N-glycans is described in the pioneering work of Harry Schachter and his group (Brockhausen et al., 1988). In the Golgi, only after actions by β1,2N-acetylglucosaminyltransferase I (GnT-I) and mannosidases, β1,2N-acetylglucosaminyltransferase II (GnT-II) will add GlcNAc for further glycan modifications by branching enzymes including β1,4N-acetylglucosaminyltransferase III (GnT-III), IV, V, VI, IX, α-1,6 fucosyltransferase (Fut8), and β-galactoside α-2,6-sialyltransferase 1 (ST6Gal1) (Fig. 1).
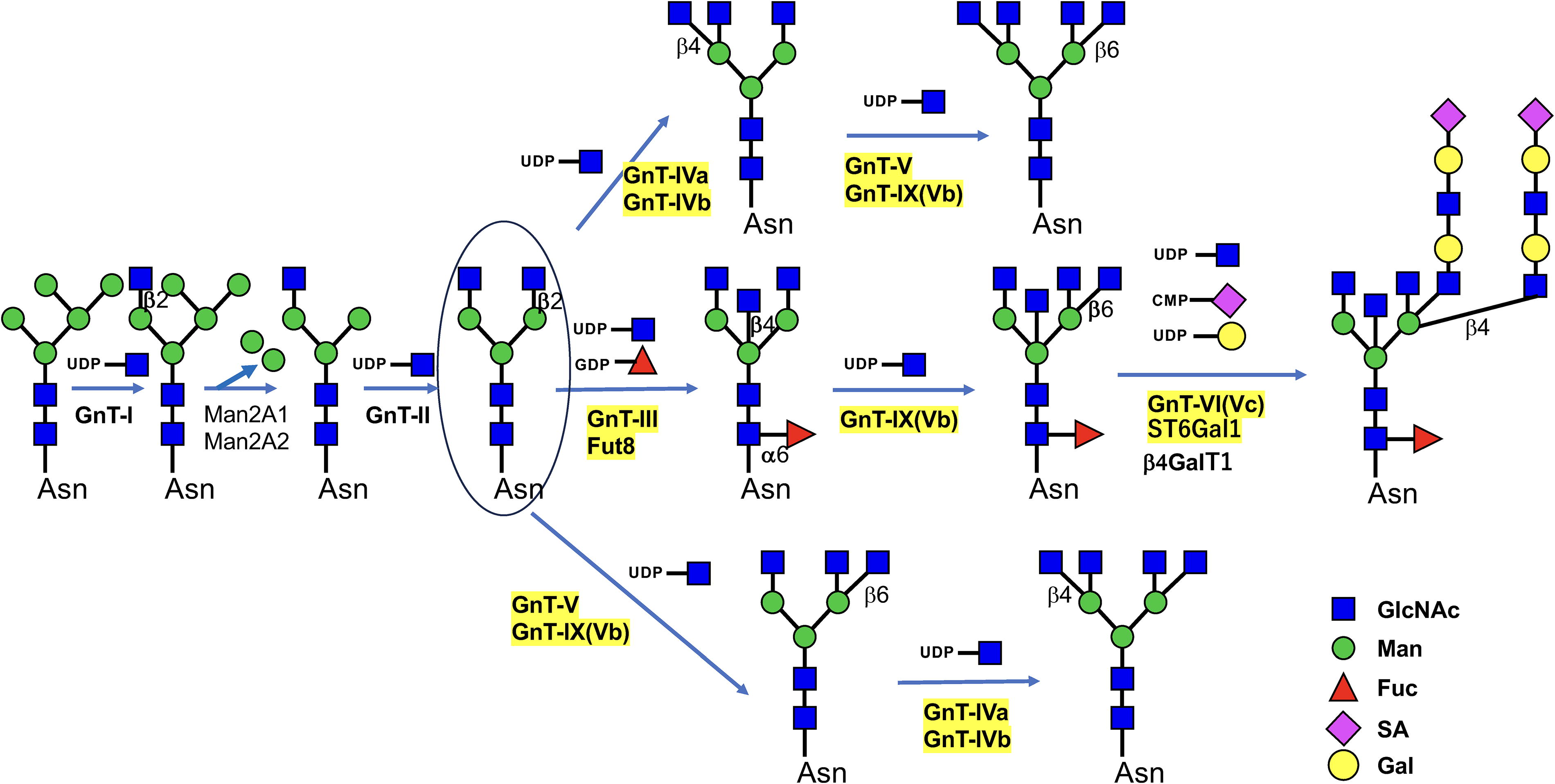
Our group used biochemical techniques and affinity chromatography for the purification and gene cloning of major N-glycan branching glycosyltransferases including GnT-III (Nishikawa et al., 1992); GnT-IVa, b (Oguri et al., 1997); GnT-V (Gu et al., 1993); GnT-VI, GnT-IVc (Sakamoto et al., 2000; Taguchi et al., 2000); and, Fut8 (Uozumi et al., 1996). An exception was GnT-IX, GnT-Vb, which was identified via homology cloning (Inamori et al., 2003). Regarding the purification and gene cloning of GnT-V, our group and the Pierce group purified the enzyme independently from a human lung cancer cell line (Gu et al., 1993) and from a rat kidney (Shoreibah et al., 1992), respectively. All of these glycosyltransferases are involved in the biosynthesis of N-glycan branching (Stanley et al., 2009), but GnT-IX (also referred to as GnT-Vb) is also involved in O-glycosylation (Abbott et al., 2006). GnT-VI (GnT-IVc) was originally found in a hen oviduct (Brockhausen et al., 1989), but a recent study has shown that it is related to prostate cancer and a stem cell marker (Mak et al., 2011). This review focuses on GnT-III, IV, V, VI (IVc), IX (Vb), Fut8, and ST6Gal1, as highlighted in yellow in Figure 1.
The above-mentioned glycosyltransferases are known to generate various structural and functional changes in many glycoproteins (Taniguchi et al., 2006). Various cell-surface receptors have been modified using glycosyltransferases (Sato et al., 2001; Takahashi et al., 2016a; Takahashi et al., 2016b), and we and other researchers have focused on evaluation of the target proteins of glycosyltransferases that have been identified using mutant mice, including glycosyltransferase gene-knockout (KO) mice and glycosyltransferase gene-transgenic mice. For in vitro analyses, gene-edited cells (glycosyltransferase-KO or-overexpressing cells) have also been used. Lectins are good research tools for use in recognizing specific glycan structures, and treatment with glycosidases and use of inhibitors targeting glycosyltransferases are often extremely helpful in analyzing target proteins and their glycan structures. Techniques such as high-performance liquid chromatography and mass spectrometry (MS) are currently being used to identify target proteins and their structures.
Some representative examples of target molecules of N-glycan-branched glycosyltransferases are listed in Table 1. Many of the direct target molecules are plasma membrane proteins, receptor molecules, and adhesion molecules such as integrins and E-cadherins. As shown in Table 1, many of the target proteins are known to be altered by oxidative stress. These target proteins are modified directly or indirectly by their respective glycosyltransferases.
Target Molecules Toward N-Glycan Branching Glycosyltransferases and Their Implication in Oxidative Stress
Due to the limited number of citations, the list is not comprehensive and only one of the representative hallmarks is described.
AFP, α-fetoprotein; AKT, AKT serine/threonine kinase; BACE1, β-site amyloid precursor protein cleaving enzyme; CAR-T, chimeric antigen receptor-T cell; EGFR, epidermal growth factor receptor; ERK, extracellular signal-regulated kinase; IgG, immunoglobulin; JNK, c-Jun terminal kinase; GLUT2, glucose transporter2; γ-GTP, γ-glutamyl transpeptidase; iNOS, inducible nitric oxide synthese; eNOS, endothelial nitrix oxiside synthes; LEF1, β-catenin lymphoid enhancer binding factor-1; PD1, program death protein 1; PDGF, platelet-derived growth factor; PDL1, program death protein ligand; PI3K, phosphoinositide three kinase; PTPRβ, protein tyrosine phosphatase β; Ras, rat sarcoma viral oncogene; SPARC, secreted proteins that are acidic and rich in cysteine; SP-D, surfactant protein D; STAT3, signal transducer and activator of transcription; TCR, T cell receptor; TIMP-1, tissue inhibitor of metalloproteinase 1; TGF-β, transforming growth factor β; VEGF, vascular endothelial growth factor; VEGFR, vascular matrix metalloproteinase; Wnt, wingless, the Drosophila melanogaster segment polarity-gene and integrase-1.
Glycosyltransferases are known to be regulated by the levels of nucleotide glycans as donor substrates, glycoproteins as acceptor substrates, antagonism between enzymes, and by signal transduction pathways. Each glycan is highly associated with various diseases or biological functions, and diagnostic biomarkers and disease situations are shown in Table 2.
Pathophysiological Changes of N-Glycan Branching Glycosyltransferases and Hallmarks
Glycosyltransferases involved in N-glycan branching and their implications for pathophysiological significance, except for GnT-IX that is implicated in O-glycosylation. The list consolidates the current research on N-glycan branching glycosyltransferases in relation to diseases, but it may not be comprehensive, and the classification(s) may be subject to further research.
ADCC, antibody-dependent cellular cytotoxicity; CDG, congenital disorders of glycosylation; COPD, obstructive pulmonary disease; MUC, mucin; O-GT, O-GlcNAc transferase; TET3, ten-eleven translocation 3.
Our previous studies showed that glycan branching enzymes and their enzymatic products are altered in various diseases including Alzheimer’s disease (AD), chronic obstructive lung disease (COPD), and cancer (see review in [Taniguchi et al., 2022]). Moreover, many studies have shown that oxidative stress plays a significant role in the onset of AD (Christen, 2000; Plascencia-Villa and Perry, 2021; Simpson and Oliver, 2020), COPD (Barnes, 2020), and various cancers (Hayes et al., 2020).
There are various target molecules of GnT-III that are related to cancer metastasis via adhesion to molecules such as E-cadherin and EMT-mediating proteins and are related to amyloid-β (Aβ) production in AD, which is highly implicated in oxidative stress.
Previous studies showed that the brains of AD patients have increased oxidative stress and that the increased oxidative stress might play a role in the pathogenesis of neuron degeneration (Markesbery, 1997; Shuvaev et al., 2001). Expression of the GnT-III gene is increased in the brains of AD patients as compared with patients with mild cognitive impairment. Liquid chromatography–electrospray ionization–mass spectrometry (LC/ESI/MS) analysis has shown that β-site amyloid precursor protein-cleaving enzyme 1 (BACE1), a protease essential for Aβ production in AD, is also modified by GnT-III (Kizuka and Taniguchi, 2018). BACE1 from Neuro2A cells contains Asn153 and Asn223 as bisecting GlcNAc-modified sites. These mutations almost cancelled the enhanced BACE1 degradation in GnT-III gene (Mgat3) KO mouse embryonic fibroblasts, indicating that bisecting GlcNAc on BACE1 indeed regulates its degradation. In tumors, GnT-III is known to exert suppressive effects on cancer metastasis in mice (Yoshimura et al., 1996). However, while normal ovary GnT-III transcription and enzyme activity were found to be very low, analysis with a lectin capture strategy coupled to nano-ESI-RPLC-MS/MS showed that the expression of GnT-III mRNA is amplified in ovarian cancer tissue compared with that in normal ovarian tissue (Abbott et al., 2008). Attachment of bisecting GlcNAc to the Fc glycan is known to induce antibody-dependent cellular cytotoxicity (ADCC) (Shields et al., 2002). Expression of GnT-IVa is significantly reduced in pancreatic β-cells of patients with type 2 diabetes (Ohtsubo et al., 2011), while GnT-IVa expression is high in peripheral white blood cells of type 2 diabetes patients (López-Orduña et al., 2007). This indicates that GnT-IVa is transcriptionally controlled in diverse ways according to the cell type. In pancreatic β-cells, GnT-IVa is transcriptionally controlled by class O of forkhead box transcription factors (forkhead box A2; FOXA2) and hypoxia-inducible factor 1α (HIF-1α) (Gunton et al., 2005), indicating that intracellular distribution is regulated by cellular redox balance. This also suggests that GnT-IVa is a typical model of glyco-redox regulation. The membrane sub-domain distribution of glucose transporter 2 is associated with the glucose transport activity of β-cells, in which the GnT-IVa-dependent formation of the N-glycan-galectin lattice plays an important role. This provides a novel pathophysiological insight into the mechanism of β-cell failure in the pathogenesis of type 2 diabetes (Ohtsubo et al., 2013). The association of GnT-V and its products with cancer metastasis have been reported by several authors (Chakraborty and Pawelek, 2003; Shao et al., 1999), but the target proteins were unclear at that time (Chakraborty and Pawelek, 2003; Guo et al., 2014), and the involvement in EMT and angiogenesis is described below (Saito et al., 2002). GnT-VI (GnT-IVc) was revealed to play the role of a stem cell marker (Mak et al., 2011) as described above. GnT-IX (GnT-Vb) modifies the glycation of laminin (Abbott et al., 2006). Fut8 KO mice show growth retardation and develop emphysema after birth (Wang et al., 2005). These mice are suitable models for human COPD (Gao et al., 2012; Kobayashi et al., 2013), and Fut8 expression is decreased in patients with COPD (Gao et al., 2012) and patients with idiopathic pulmonary fibrosis (Sun et al., 2021).
Behavior abnormalities have been observed in Fut8-heterozygous KO mice (Fukuda et al., 2011). Congenital disorders of glycosylation FUT8 mutation have been detected in humans with various symptoms including dysmorphisms, failure to thrive, respiratory abnormality (Ng et al., 2020), and glaucoma (Schweigert and Areaux, 2019). The core fucose has been shown to play an important role in ulcerative colitis (UC) in both mouse models (Cantero-Recasens et al., 2022; Fujii et al., 2016) and patients (Cantero-Recasens et al., 2022). Levels of core fucosylation were increased in T cells from mice with colitis compared with mice without colitis as well as in the inflamed mucosa of patients with infectious bowel disease compared with the levels in noninflamed tissues or tissues from control patients (Cantero-Recasens et al., 2022). Core fucosylation of T cell receptors is required for T cell signaling and for production of inflammatory cytokines and the induction of colitis in mice (Fujii et al., 2016). FUT8 modifies the biophysical properties of mucous by controlling the levels of cell surface MUC1 and both the quantity and quality of secreted mucin and mucin 5AC. These changes suggest that mucus viscosity facilitates bacterial–epithelial interactions that lead to inflammation and UC progression (Cantero-Recasens et al., 2022). FUT8 facilitates invasion and tumor dissemination, in part due to the reduced cleavage of L1 adhesion molecules by core fucosylation, and acts as a driver for melanoma metastasis (Agrawal et al., 2017). ST6Gal1 is changed in many cancers such as pancreatic cancer (Bhalerao et al., 2023), prostate cancer (Scott et al., 2023), and lung cancer (Yuan et al., 2018), and this enzyme modulates many signaling molecules and target proteins (Gc et al., 2022).
A schematic drawing depicting the interplay between oxidative stress and glycan changes in relation to diseases is shown in Figure 2. An imbalance between production of reactive oxygen species/reactive nitrogen species (ROS/RNS) and levels of antioxidants results in oxidative stress, which either directly or indirectly causes the occurrence of various oxygen stress-related diseases, such as AD, COPD, and type 2 diabetes, as described above. Interstitial pneumonitis (Chakraborty and Pawelek, 2003) and UC (Arab et al., 2014) are also related to oxidative stress. As a direct effect of oxidative stress, nonenzymatic glycosylation, which results in free radical formation and in the formation of various advanced glycation end products (AGEs) occurs. These reactions have been described as glycoxidation reactions. A relationship between AGEs and AD has also been reported (Shuvaev et al., 2001). Due to space limitations, we have not included this modification, but it is depicted in Figure 3.


ROS and Antioxidant Defense Systems and Their Involvement in Glycan Changes in Diseases
ROS and RNS are generated in cellular components such as the mitochondrial transport system and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase. The generation of ROS/RNS and antioxidants for their removal is important, and oxidative stress eventually occurs if levels of oxidants reach high levels (Sies, 2015).
As mentioned above, oxidative stress causes a number of diseases that involve alterations in glycans. Among these diseases, in addition to AD, interstitial pneumonia, COPD, and UC have been shown to increase the risk of developing cancer.
The compartment sites for the production of ROS and RNS, as well as sites for production of antioxidants, within cells are known. For example, the generation of superoxide occurs during oxidative phosphorylation in mitochondria and involves a number of enzymatic sources within the cytosol and at the cell membrane. Involvement of ROS in the cleavage of glycans was first reported in hyaluronan that was degraded by hydrogen peroxide and hydroxyl radical generated by hydrogen peroxide (Roberts et al., 1989). It is well known that diverse types of antioxidant systems play pivotal roles in redox regulation. For example, under the inflammatory conditions, the electron transport system in neutrophiles and macrophages generates many forms of ROS and RNS including NADPH oxidases (NOX1-4, Duox1,2) (Babior, 2004), xanthine oxidase (Chung et al., 1997), and nitric oxide synthases (Rochette et al., 2013). These ROS and RNS serve as second messengers that regulate various signaling pathways such as nuclear factor-kappa B (Morgan and Liu, 2011), nuclear factor-erythroid 2-related factor 2 (Ma, 2013), histone deacetylase (HDAC) (Shimazu et al., 2013), ten-eleven translocation (TET) (Zhang et al., 2017), FOXA2 (Kittappa et al., 2007), and HIF (Dewhirst, 2009), and all of these are involved in various biological events such as cell proliferation, apoptosis, autophagy, migration, DNA damage, inflammation, and drug resistance (Sies and Jones, 2020). For example, the lungs are exposed to various environmental irritants such as cigarette smoke that causes the production of ROS. Cigarette smoke promotes the recruitment of activated neutrophils and macrophages and produces both ROS and RNS and therefore causes accumulation of damage to the surface area of lungs, which eventually result in acute and chronic inflammation such as that seen in acute respiratory distress syndrome (McCord et al., 1994) and in COPD (Barnes, 2020).
It is notable that 15 genes were selected from RNA sequence data of glycan synthesis-related genes and antioxidant response genes; the 15 genes include Fut8 and GnT-III. After treatment with 5-hyroxy-4-phenylutenolide and sulphoraphane, Fut8 mRNA expression was increased in the human keratinocyte cell line HaCaT, while the expression of GnT-III was decreased (Kyunai et al., 2023). Recently Fut8 was found to be highly correlated to the enzymatic function of an antioxidant enzyme SOD3 that is a good example for glyco-redox (Ohkawa et al., 2023). There are three kinds of SOD in mammals: SOD1, SOD2, and SOD3. SOD1 is also referred to as Cu-Zn-SOD, while SOD2 is referred to as Mn-SOD, and they are localized in the cytosol and mitochondria, respectively. SOD3, on the other contrary, is referred to as extracellular SOD and is localized in extracellular spaces (Marklund, 1990). SOD3 has one N-glycan that is required for enzyme activity and secretion capacity, and it has also shown suppressive activity against cancer (Ota et al., 2017; Ota et al., 2016). SOD3 contains a heparin-binding domain with a cluster of six positively charged amino acids in the carboxy terminus of the protein that creates a strong binding affinity to collagen, heparin, and other negatively charged matrix elements (Marklund, 1990). Among the cluster of six positively charged amino acids in the heparin-binding domain, arginine is important because both animal and human genetic studies have indicated that a functional mutation (R213G) results in a marked decrease in affinity for the negatively charged extracellular matrix and a marked increase in its circulating levels. It is well known that tobacco smoke is the most common contributor to COPD, but 35% of heavy smokers do not develop COPD due to a genetic polymorphism. In fact, it has been shown that persons who possess the R231G variant of SOD3 have a large amount of SOD3 in lung extracellular fluid and plasma, which protects against lung inflammation and COPD even if they are heavy smokers (Bentley et al., 2008; Oberley-Deegan et al., 2009). As shown in Figure 3, SOD3 has a heparin-binding domain and binds to endothelial cells, but a mutant without this domain is released into the plasma or extracellular space of alveoli. In addition, SOD3 has one N-glycan with core fucose and sialic acids, but their functions in binding to endothelial cells have not been elucidated. We found that a core fucose glycan in SOD3 is also increased in the sera of patients with lung cancer (Ohkawa et al., 2023). An elevated level of the core fucose structure has been found in non-small cell lung cancer (NSCLC) cell lines (Kondo et al., 2022; Ohkawa et al., 2023). Moreover, non-core fucosylated and sialylated SOD3 increases extracellular excretion and scavenging activity and cell growth suppression (Ohkawa et al., 2023). The presence of terminal sialic acids in the N-glycans of SOD3 enhanced both the secretion and furin-mediated C-terminal cleavage of SOD3 (Ota et al., 2017; Ota et al., 2016). The involvement of Fut8 and ST6Gal1 in SOD3 function is summarized in Figure 2.
Interplay Between N-Branching Glycans and Redox Regulation
Redox regulation was originally defined as oxidation–reduction of thiol groups (Halliwell and Gutteridge, 2015) via ROS. Modifications of thiol groups have been reported. Redox balance in apoptosis, autophagy, endoplasmic reticulum stress, and mitochondrial dysfunction has been implicated in various diseases. The role of redox regulation in various glycans including O-GlcNAcylation (Martinez et al., 2017) was recently reviewed (Khoder-Agha and Kietzmann, 2021). In this review, we focus on N-glycan branching and their regulation by redox signaling.
HIF-1 and hypoxic regulation of glycosylation has been proposed to occur via N-glycan-related glycosyltransferases and their donor nucleotide-related transporters (Shirato et al., 2011). The Mgat3-encoded GnT-III activates Notch signaling in epithelial ovarian cancer (Allam et al., 2017). Mgat5-encoded GnT-V is regulated by Ets-1 (E-twenty-six family) (Kang et al., 1996; Ko et al., 1999) and plays a key role in cancer metastasis and hypoxic situations (Silva-Filho et al., 2017). ST6Gal1 is also regulated by HIF-1 (Jones et al., 2018), while GnT-IX(Vb) is regulated by a combination of specific chromatin modifiers including HDAC-1, O-GT, and TET3 and by transcription factor neuroD1 (Kizuka et al., 2014). Exposure to hypoxia (1% O2) and treatment with CoCl2 resulted in significant suppression of the expression of macrophage scavenger receptor 1 mRNA accompanied by an increase of the expression of HIF-1α protein in the nucleus. Therefore, HIF-1 and hypoxic regulation of glycosylation has been proposed to occur via N-glycan-related glycosyltransferases and their donor nucleotide-related transporters (Shirato et al., 2011). The expression of Mgat3-encoded GnT-III is high and activates Notch signaling in epithelial ovarian cancer. (Allam et al., 2017).
Involvement of Glycans and Redox Regulation in EMT/MET and Cancer
EMT/MET has been reported to play roles in wound healing, embryonic development, and cancer progression (Terry et al., 2017; Yang and Weinberg, 2008). Glycans are also known to play a key role in EMT/MET (Li et al., 2014; Pinho et al., 2012; Takahashi et al., 2022; Takahashi et al., 2016b; Taniguchi et al., 2021; Terao et al., 2011; Xu et al., 2012; Zhang et al., 2019). The significance of redox regulation in EMT and its molecular basis was a factor in proposing its therapeutic utility (Abbott et al., 2008; Jiang et al., 2017). Oxidative stress also promotes the expression of transforming growth factor-β1 (TGF-β1). Inhibition of TGF-β1 signaling as well as treatment with SOD3 prevents the oxidative stress-driven EMT-like changes in COPD and interstitial pneumonitis, and interventions also inhibited EMT-like changes (Gorowiec et al., 2012). In fact, compelling evidence highlights ROS as crucial conspirators in EMT engagement (Giannoni et al., 2012). Those studies have identified a link between oxidative stress, TGF-β1, and EMT in the lung epithelium and chronic lung diseases such as COPD and interstitial pneumonitis.
Figure 4 shows the role of N-glycan branching glycosyltransferases in the EMT/MET process by adding recent findings to our previous studies (Taniguchi et al., 2021; Takahashi et al., 2016b). E-cadherin is a glycoprotein, and partial loss of E-cadherin has been associated with carcinoma progression and a poor prognosis in various forms of human and mice tumors. Glycan changes in E-cadherin were first found in B16 melanoma cells with high GnT-V activity. These changes have the capacity for experimental metastatic lung cancer, whereas transfection of GnT-III into B16 melanoma cells suppresses metastasis in lung cancer (Yoshimura et al., 1995). These facts support the EMT/MET process that occurs via the modification of E-cadherin glycans. Several reports have provided information concerning on the glycans of E-cadherin and integrin in various cancer cell lines (Iijima et al., 2006; Osumi et al., 2009; Pinho et al., 2012; Zhao et al., 2008). When E-cadherin is modified with bisecting GlcNAc, turnover is delayed and release from the cell surface is decreased, resulting in elevation of expression at the cell-cell border of GnT-III transfectants (Yoshimura et al., 1996). Furthermore, cell aggregation is enhanced in GnT-III transfectants, indicating that glycosylated E-cadherin is biologically functional (Iijima et al., 2006). Introduction of GnT-III also results in downregulation of the tyrosine phosphorylation of β-catenin, as well as downregulation of the intracellular signaling pathway, suggesting its involvement in cell motility and in the suppression of cancer metastasis (Kitada et al., 2001).

The bisecting GlcNAc structure inhibits EMT in breast cancer (Tan et al., 2018). TGF-β1 downregulates the expression of GnT-III in EMT-like changes in human MCF-10A and mouse GE11 cells (Xu et al., 2012). On the other contrary, the expression of GnT-V was slightly enhanced by TGF-β1 treatment. It is of particular interest that the overexpression of GnT-III influenced the EMT-like changes induced by TGF-β1, which was confirmed by cell morphological changes in phase contrast as well as by the immunochemical staining patterns of E-cadherin and actin. In addition, GnT-III modification prolonged E-cadherin turnover on the cell surface when examined via biotinylating and pulse-chase experiments. GnT-III expression consistently inhibited the β-catenin translocation into the cytoplasm and nucleus, which is normally caused by cell–cell contact. Furthermore, a trans-well assay showed that GnT-III expression suppresses TGF-β1-induced cell motility (Kitada et al., 2001). Taken together, these observations are the first observations that clearly demonstrate that GnT-III affects cell properties, which in turn influences EMT-like changes and suggests a molecular mechanism for the inhibitory effects of GnT-III on cancer metastasis. The EMT process is also related to GnT-V, which also modulates E-cadherin glycans by downregulating functionality (Guo et al., 2003). The specific site of E-cadherin glycans was confirmed to be important in the treatment of gastric cancer and in the modulation of E-cadherin function by GnT-III as well as that by GnT-V, Fut8 (Osumi et al., 2009), and ST6Gal1 (Bhalerao et al., 2023; Krick et al., 2021; Lu et al., 2014). GnT-V and FUT8 are highly activated in most tumor cells.
Lin et al. showed that Fut8 plays a key role in EMT in renal interstitial fibrosis by blocking TGF-β and platelet-derived growth factor signaling pathways (Lin et al., 2011), and FUT8 is upregulated during EMT via the transactivation of β-catenin/lymphoid enhancer-binding factor-1 in NSCLCs (Chen et al., 2013). Regarding GnT-V and EMT, overexpression of GnT-V promotes EMT, and suppression of GnT-V expression downregulates EMT. However, one study showed a low level of GnT-V expression impairs TGF-β/Smad signaling and promotes EMT, migration and invasion in lung cancer cells (Li et al., 2014). In NSCLC GnT-V activity downregulates cancer promotion (Dosaka-Akita et al., 2004). These results suggest that the effect of GnT-V on EMT and cancer progression might be dependent on the tissues.
Our group found that when fucosylated immunoglobulin G1 (IgG1) is measured by a specific antibody against core fucose in IgG, it is highly downregulated in the serum of lung cancer as well as in interstitial pneumonitis and gives rise to fibrosis in lung tissue and C-C chemokine receptor type 2 regulates partly this reaction (Ohkawa et al., 2023). A decrease in the concentration of core fucose in IgG1 indicates an increase in a core fucosylated (non-core fucose) IgG1 and suggests that an adaptive response against fibrosis could occur via an upregulation of ADCC activity in patients with interstitial pneumonia; Fut8 is highly activated in many cancers and normally promotes the EMT process. However, the decreased expression of Fut8 in lung cancer and interstitial pneumonia may be an adaptive response to reverse from EMT to MET.
St6Gal1 also promotes TGF-β-mediated EMT in breast cancer cells (Lu et al., 2014; Takamiya et al., 2021; Takahashi et al., 2016b; Tao et al., 2019). These studies suggest that GnT-V and Fut8 mostly play a role in promoting the EMT process but are also implicated in the MET process.
Conclusions
This mini review summarizes findings regarding glyco-redox, particularly findings regarding ROS and N-glycan branching glycosyltransferases in EMT/MET and various diseases such as AD, COPD, cancer, and UC, and perspectives also are discussed.
It might be possible to use antioxidants for controlling the EMT/MET processes, and new types of antioxidants for treating cancer and other diseases might be discovered in the near future.
The interplay of glycans and redox regulation is designated as the glyco-redox concept, and it plays a pivotal role in various pathophysiological events including AD, COPD, fibrosis, and cancer.
Footnotes
Acknowledgments
The authors would like to acknowledge many colleagues in our past and present laboratories who contributed to glycobiology and redox research. Due to page limitation, I must apologize that I could only cite limited numbers of references. I would like to dedicate this article to the late Dr. John Gutteridge, a scientist who made great contributions in this field who was a visiting professor in Osaka University Medical School when N.T. was a chair of the Department of Biochemistry. We acknowledge Mr. Stewart Chisholm who edited English language of our article. This article is supported by a personal contribution (20211201 and 21K06083 for M.T.). Finally, N.T. acknowledge Dr. Mikko Laukkanen who invited him to contribute a mini-Forum Issue in this journal.
Authors’ Contributions
N.T. and M.T. conceived and wrote most of this article, and Y.O. provided useful data and suggestions. T.K., J.A., and Y.H. helped to research the references and draw figures. All authors read and approved this article.
Data Availability Statement
The data that support the findings of this review are available on request.
Author Disclosure Statement
The authors declare no conflicts of interest associated with this study.
Funding Information
This study was supported in part by Grants-in Aid for Scientific Research © (23K06734 to N.T. and 23K06420 to Y.O.) from the Japanese Society for Promotion of Science and public grants from Osaka Prefecture (52000090 and 52000093) and the Osaka Adult Disease Foundation to N.T.