
Abstract
Significance:
The role of melatonin (MEL) in plants has gained significant relevance due to its involvement in a wide range of physiological functions, particularly in response mechanisms to both abiotic and biotic stresses.
Recent Advances:
Recent progress highlights the significance of the biosynthetic pathway of MEL in plants, which surpasses that of animals. The discovery of specific plant MEL receptors has revealed new signaling mechanisms. Studies also show that applying exogenous MEL offers benefits under stress conditions and helps maintain the organoleptic qualities of fruits and vegetables during postharvest storage.
Critical Issues:
This review explores MEL’s biochemistry, emphasizing its dual role as both an antioxidant and a signaling molecule. It examines how MEL interacts with phytohormones, its role in regulating the metabolism of reactive oxygen and nitrogen species, and its influence on plant growth and stress tolerance. The potential of MEL-based biotechnological applications for enhancing crop resilience and postharvest quality is also discussed.
Future Directions:
Future research should prioritize molecular mechanisms, high-throughput approaches, and translational studies to bridge the gap between fundamental science and agricultural practices. MEL’s role as a sustainable solution in agriculture offers exciting possibilities for addressing global food security challenges. Antioxid. Redox Signal. 43, 151–188.
Introduction: Melatonin, a Short History
After an extraordinary effort involving the maceration and extraction of 250,000 bovine pineal glands, a few milligrams of a white powder were recovered. Aaron B. Lerner identified this substance as N-acetyl-5-methoxytryptamine, which he named melatonin (MEL). The name was derived from its relationship to serotonin (“-tonin”) and its effects on amphibian melanocytes (“mela-”). Lerner’s groundbreaking discovery was published in two concise reports in 1958 and 1959 (Lerner et al., 1959a; Lerner et al., 1958b), which, despite their brevity, marked a pivotal moment in the understanding of this molecule’s significance.
Initially, MEL was recognized for its role in circadian rhythms and photoperiodic regulation of reproduction in seasonally breeding mammals (Reiter, 1993). Over time, the scope of its physiological significance expanded dramatically. The notion that MEL is exclusively produced by the pineal gland was debunked with its discovery in insects, protists, plants, and numerous extrapineal mammalian tissues. Remarkably, MEL levels in these tissues remained unchanged following pinealectomy (Acuña-Castroviejo et al., 2014). These findings highlighted its evolutionary ubiquity, as MEL was identified in prokaryotes and archaea, organisms that predate vertebrates by billions of years (Hardeland, 2015; Kim et al., 2024; Dubbels et al., 1995). The evolutionary timeline of MEL suggests its origin precedes vertebrates, which date back roughly 520 million years. Prokaryotes capable of synthesizing MEL were likely engulfed by primitive eukaryotic cells, eventually evolving into mitochondria and chloroplasts, as posited by the endosymbiotic theory (Zhao et al., 2019). These organelles retained MEL synthesis due to its protective role as a multifunctional antioxidant, particularly in counteracting reactive oxygen species (ROS) (Venegas et al., 2012; Suofu et al., 2017; Back et al., 2016). Today, MEL is understood to be synthesized in mitochondria and chloroplasts, as well as potentially in other organelles, across all species in the plant and animal kingdoms. This ancient molecule plays a universal role in maintaining cellular homeostasis, bridging the evolutionary gap between the simplest organisms and complex vertebrates (Manchester et al., 1995; Lee et al., 2022; Lee and Back, 2023; Kwon et al., 2024) (Fig. 1).

As far as it can be envisaged, MEL presumably evolved independently at different times to protect against organismal oxidative stress. Cyanobacteria, which are believed to have evolved before proteobacteria, developed oxygenic photosynthesis (Sánchez-Baracaldo et al., 2022). Since this process generates toxic oxygen derivatives, MEL likely emerged as a scavenger for these reactive species. Over millions of years, photosynthetic bacteria contributed to a dramatic rise in atmospheric oxygen during an epoch known as the Great Oxidation Event, which occurred ∼2.4 billion years ago (Sánchez-Baracaldo and Cardona, 2020). This event eventually led to the use of oxygen as the foundation of aerobic metabolism. As a result, an efficient molecule was needed to neutralize the ROS generated by this metabolic shift, potentially driving a second evolution trend of MEL in proteobacteria. When cyanobacteria and proteobacteria were engulfed as food by early eukaryotes, they carried their radical-scavenging ability with them and eventually evolved into mitochondria and chloroplasts, a key concept of the endosymbiotic theory for organelle evolution (Zimorski et al., 2014).
The chemical structure of MEL remains identical across all domains of life, including bacteria, archaea, and eukaryotes. In all species, its original function, which has persisted to the present day, is believed to be the detoxification of oxidizing species produced during internal metabolism. In eukaryotic kingdoms, MEL continues to function as a potent antioxidant, particularly in plants, where it protects against oxidative stress induced by various environmental stressors.
Throughout evolution, MEL has been co-opted for numerous additional functions, such that in the present-day species, it plays multiple critical roles, as illustrated in Figure 1. Many of these functions likely evolved after the emergence of membrane and nuclear receptors for MEL (Carrillo-Vico et al., 2003), which may have first appeared with the rise of vertebrates ∼520 million years ago during the Cambrian Explosion. Thus, for billions of years, MEL primarily served as a radical scavenger before acquiring its diverse physiological roles.
These discoveries, along with the vast literature that documents MEL’s actions as an antioxidant and to overcome oxidative stress, have revolutionized the functional image of MEL and linked it to numerous fundamental cellular functions; its actions are widespread, diverse, and cell-life sustaining. Indeed, the research related to the function of MEL in plants currently outpaces the research in animals (Corpas et al., 2022). The current review considers exclusively the ability of MEL to function in the control of oxidative/nitrosative stress in plants, but its actions far exceed these boundaries. Thus, this review delves into the history, biosynthesis, and functions of MEL in higher plants, providing a comprehensive overview of its multifaceted roles and potential applications in agriculture.
Biosynthesis of MEL in Higher Plants
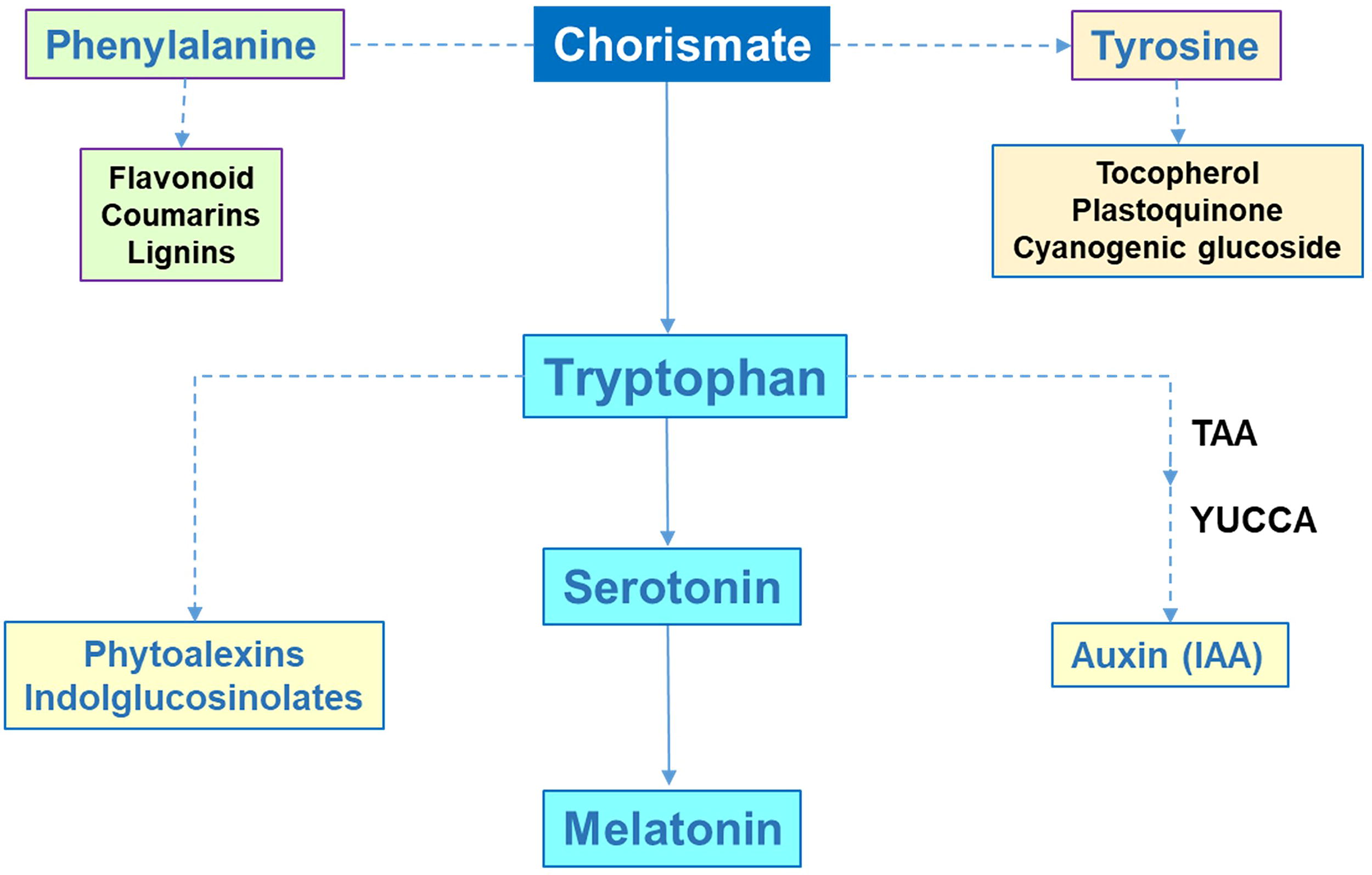
MEL biosynthesis in higher plants involves a series of enzymatic steps, starting from the amino acid tryptophan (Trp) via the shikimate (chorismate) pathway. This pathway is named after shikimic acid was identified, a compound first isolated in 1885 by Johan Fredrik Eykman from the flowers and seeds of the Japanese shikimi, an evergreen shrub closely related to the Chinese star anise (Illicium anisatum) tree. The pathway uses two substrates: phosphoenolpyruvate and erythrose-4-phosphate, with the final product, chorismate, serving as the precursor for the biosynthesis of the aromatic amino acids Trp, tyrosine, and phenylalanine. These amino acids are the base of numerous compounds involved in the secondary metabolism of plants, many of which have signaling functions. In particular, Trp is a precursor for a diverse range of secondary metabolites, including phytoalexins, indole glucosinolates, and the phytohormone indole-3-acetic acid (IAA), which plays a key role in auxin signaling (Radwanski and Last, 1995; Ishihara et al., 2008; Lynch and Dudareva, 2020; Corpas et al., 2021) (Fig. 2). Consequently, this route supports the biosynthesis of MEL.
Since the discovery of MEL in animals, its biosynthetic pathway has been extensively studied. However, some aspects remain unresolved due to the potential reversibility of certain processes (Tan et al., 2016), raising questions about how this impacts our understanding of the signaling pathway, how it affects MEL homeostasis, or how MEL biosynthesis varies across different tissues beyond the pineal gland, such as the gut, skin, and immune cells, among others. Similarly, the MEL biosynthesis pathway in higher plants has also been explored, particularly in rice (Oryza sativa) (Fujiwara et al., 2010; Kang et al., 2011; Park et al., 2013a, 2013b; Byeon et al., 2014a, 2014c, 2015a, 2015b, 2016) and the model plant Arabidopsis thaliana (Byeon et al., 2014b, 2015c; Zuo et al., 2014; Lee et al., 2015, 2016; Yang et al., 2019). However, the pathway is still not fully clarified, especially concerning the genes encoding the involved enzymes and their subcellular localization (Back et al., 2016). Compared to the animal system, MEL biosynthesis in higher plants is more complex, involving certain reversible steps and alternative metabolic routes (Liu et al., 2022; Gao et al., 2023). The biosynthesis mainly consists of four reactions, with Trp as the precursor, derived from the shikimic acid pathway in the chloroplast (Schmid and Amrhein, 1995; Maeda and Dudareva, 2012; Lynch and Dudareva, 2020).
Figure 3 illustrates the working model for MEL biosynthesis in plant cells, including the probable subcellular localization of the enzymes involved. MEL biosynthetic pathway exhibits remarkable complexity, including reversible steps and alternative routes, which remain partially unresolved. The first step involves the decarboxylation of Trp, catalyzed by a cytosolic Trp decarboxylase (TDC; EC 4.1.1.105), which converts Trp to tryptamine using the cofactor pyridoxal-5′-phosphate (PLP). Next, tryptamine is hydroxylated by tryptamine 5-hydroxylase (T5H; EC 1.14.16.4) to form serotonin (5-hydroxytryptamine) (Kang et al., 2007; Park et al., 2012, 2013a; Erland et al., 2016). T5H belongs to the cytochrome P450 enzyme family and is located in the endoplasmic reticulum, where it requires NADPH (Fujiwara et al., 2010). Next, serotonin undergoes acetylation, catalyzed by serotonin N-acetyltransferase (SNAT; EC 2.3.1.87), located in the chloroplast, forming N-acetylserotonin (Yu et al., 2019; Guo et al., 2024). This step is a key regulatory point in MEL biosynthesis (Back, 2021). The final step involves the methylation of N-acetylserotonin, catalyzed by cytosolic N-acetylserotonin O-methyltransferase (ASMT; EC 2.1.1.4) (Lee et al., 2014; Byeon et al., 2016), resulting in the production of MEL. Alternatively, in some plant species, this final step can be catalyzed by caffeic acid O-methyltransferase (COMT; EC 2.1.1.4), a multifunctional enzyme also involved in the biosynthesis of lignin and flavonoids (Byeon et al., 2014; Byeon et al., 2015).
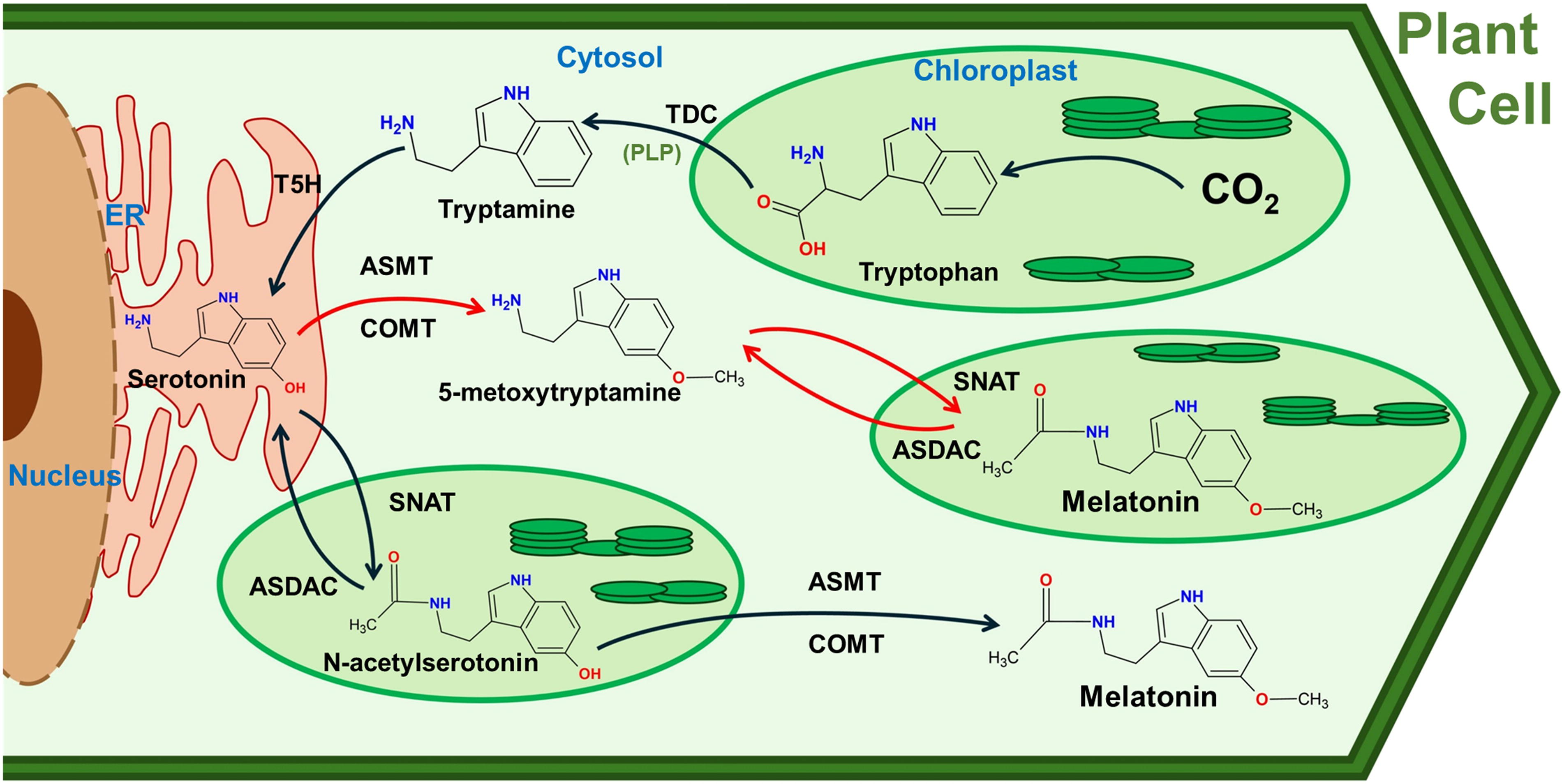
There is also an enzyme capable of catalyzing the deacetylation of both MEL and N-acetylserotonin in the chloroplast to produce 5-methoxytryptamine, known as N-acetylserotonin deacetylase (ASDAC; EC 3.5.1.98) (Lee et al., 2018). These reactions are the reverse of those catalyzed by SNAT activity. Consequently, the presence of this protein in plants is believed to facilitate the reverse reactions in MEL biosynthesis, complicating the understanding of this indolamine’s biosynthetic pathway in plants compared to animals (Lee et al., 2018). Both SNAT and ASDAC proteins are located in the chloroplast, where they play crucial roles in regulating MEL synthesis, which is crucial for maintaining optimal levels of this molecule to support proper plant development.
An important aspect is to understand the precise mechanism of interorganellar transport of MEL and its intermediates, which has neither been fully elucidated yet nor have the specific transporters been identified. However, it can be hypothesized that MEL moves between organelles through various transport mechanisms, such as (i) diffusion: as a small lipophilic molecule, MEL can passively diffuse across membranes; (ii) vesicular transport: membrane-bound vesicles may facilitate the movement of MEL between organelles; or (iii) transporter proteins: potential transporters, such as ATP-binding cassette (ABC) transporters or aquaporins, may mediate MEL transport across membranes. Therefore, an open question remains aimed at identifying specific MEL transporters and understanding how interorganellar MEL transport is regulated in response to stress.
Currently, ongoing research continues to provide additional insights into each stage of MEL’s biosynthesis in different plant species, as well as their regulatory mechanisms, because there is greater diversity than one would expect (Liu et al., 2021; Chen et al., 2022; Xiao et al., 2022; Taboada et al., 2023; Xu et al., 2023; Pham et al., 2024). Advances in molecular techniques, such as CRISPR and subcellular localization studies, could elucidate these pathways further. Highlighting key enzymes like SNAT and their regulatory mechanisms can provide insights into optimizing MEL production for agricultural applications.
Biochemical Properties and Functions of MEL Derivative Metabolites
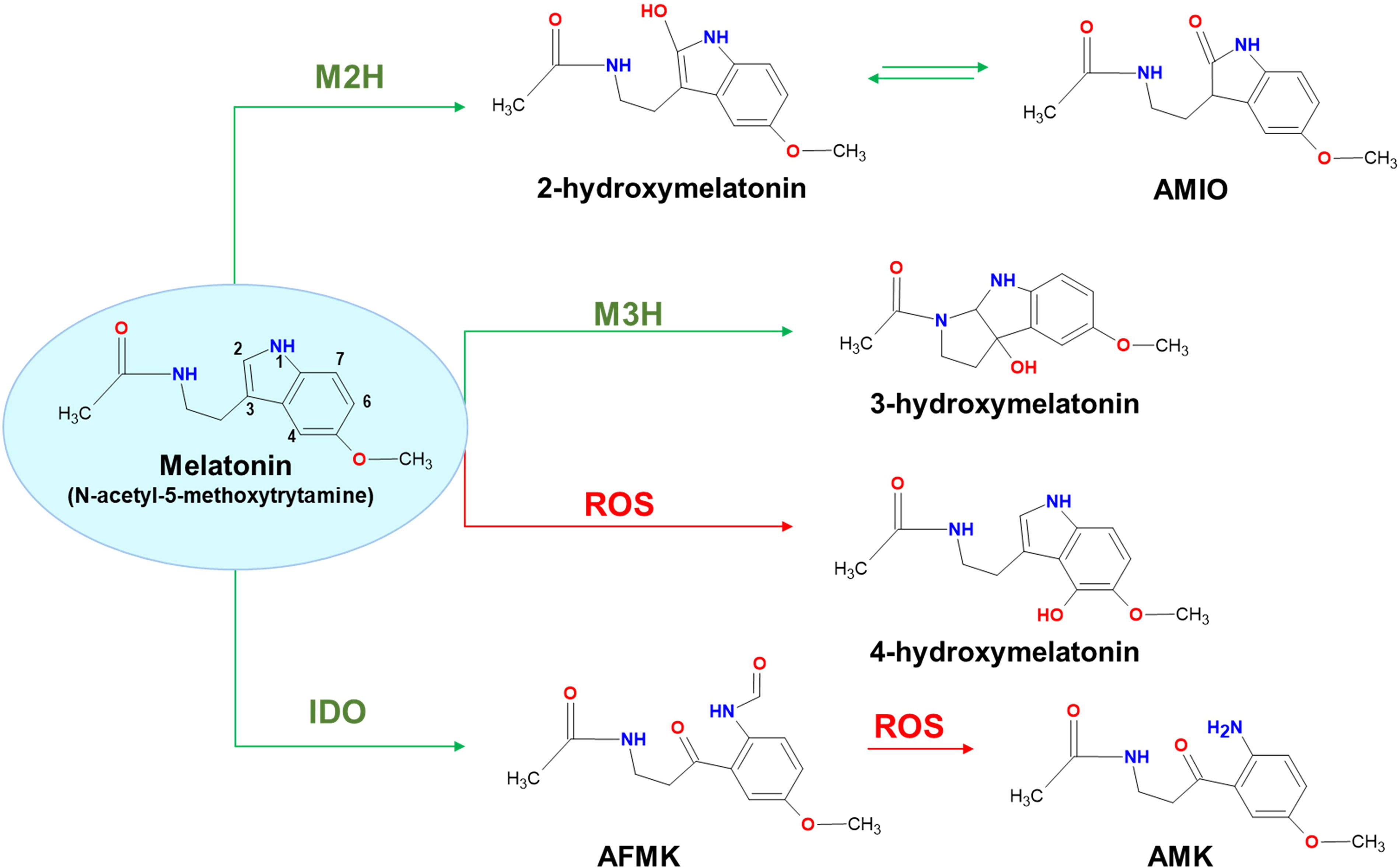
In addition to the intermediary metabolites in the MEL biosynthetic pathway mentioned earlier, various hydroxylated and nitrogenous MEL-derived metabolites with antioxidant capacity to interact with both ROS and reactive nitrogen species (RNS) have also been reported (Manchester et al., 2015; Hardeland, 2017; Corpas et al., 2022). Thus, MEL can scavenge alkoxyl radical (RO•) (Scaiano, 1995), hydroxyl radical (•OH) (Matuszak et al., 1997; Purushothaman et al., 2020), singlet oxygen (1O2) (Matuszak et al., 2003), and nitric oxide (•NO) (Noda et al., 1999). However, it is important to note that MEL content in different plant organs is within the nM range or even lower (Pranil et al., 2021; Verde et al., 2022), which is significantly lower than that of other antioxidant molecules, such as ascorbate and glutathione, which are typically found in the mM range (Airaki et al., 2011; Smirnoff and Wheeler, 2000). However, not only is MEL an effective scavenger, but its derivatives, which are formed when it scavenges, also detoxify reactive species, thereby increasing its effective concentration as an antioxidant. Thus, the relative importance of MEL as a scavenger versus its role as a signaling molecule remains to be determined.
Figure 4 summarizes the main compounds derived from MEL due to its interaction with ROS. The formation of some of these metabolites is catalyzed by specific enzymes: MEL 2-hydroxylase (M2H) generates 2-hydroxymelatonin (2-OHM), melatonin 3-hydroxylase (M3H) produces 3-hydroxymelatonin (3-OHM), and indolamine-2,3-dioxygenase (IDO) catalyzes the formation of N1-acetyl-N2-formyl-5-methoxykinuramine (AFMK). AFMK, the first identified MEL-derived metabolite, interacts with ROS to form N-acetyl-5-methoxykinuramine (AMK) (Hirata et al., 1974; Tan et al., 2001). Additionally, MEL can be hydroxylated by ROS to generate 4-hydroxymelatonin (4-OHM) (Pérez-González et al., 2017). The concentrations of these MEL-derived metabolites and their corresponding enzymes vary depending on the plant species studied (Hardeland, 2017). Such variability might result from differences in enzymatic expression, environmental factors, or developmental stages. In some species, concentrations of 2-OHM have been found to be 368 times higher than MEL, suggesting that MEL is not the final product, but rather an intermediate in a broader pathway (Byeon et al., 2015). This MEL derivative also has a tautomeric form called 2-acetamidoethyl-5-methoxy-indolin-2-one (AMIO) (Hardeland, 2017, 2019).
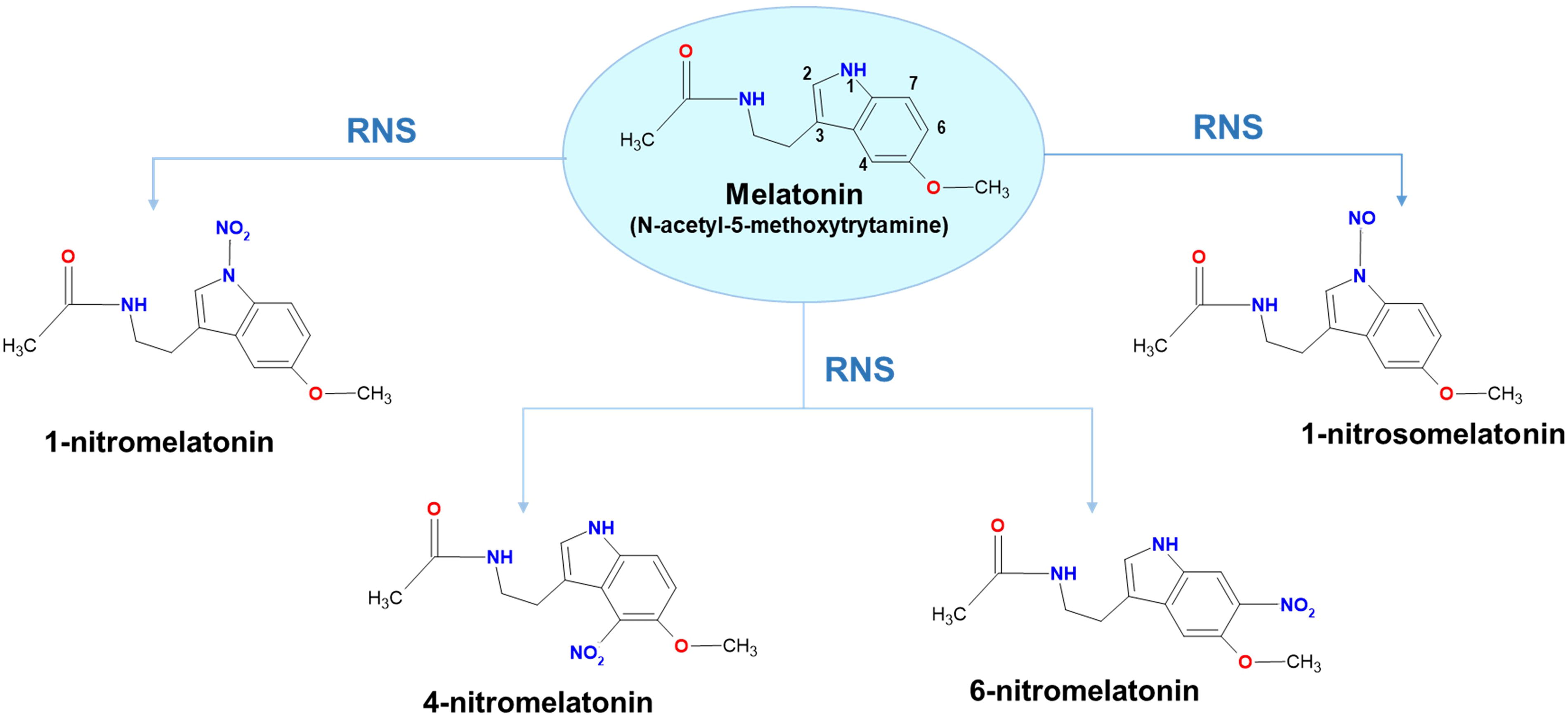
Regarding the interaction of MEL with RNS, it has been proposed that MEL can incorporate a nitro group to form either 1-nitromelatonin (1-NO2M), 4-nitromelatonin (4-NO2M), or 6-nitromelatonin (6-NO2M), but also a nitroso group to form 1-nitrosomelatonin (1-NOM) (Mukherjee, 2019; Singh et al., 2021; Corpas et al., 2022; Martínez-Lorente et al., 2022; Hussain et al., 2024). These interactions underline MEL’s potential role in mitigating nitrosative stress and enhancing plant resilience. Figure 5 outlines the main compounds derived from MEL due to its interaction with RNS.
MEL is also known for its ability to repair oxidized DNA (Galano et al., 2011, 2016; Pérez-González et al., 2019), as well as its capacity to modulate the gene expression and activity of antioxidant enzymes such as catalase, superoxide dismutase (SOD), ascorbate peroxidase (APX), or glutathione reductase (GR), in addition to increasing the content of antioxidant metabolites including ascorbate and glutathione (Wang et al., 2012; Khan et al., 2020; Sun et al., 2020; Siddiqui et al., 2020; Ahmad et al., 2020). Research has identified various mechanisms through which MEL activates the DNA damage response system, including the repair of oxidative DNA damage. For example, MEL induces poly(ADP-ribose) polymerase (PARP), a key enzyme in the base excision repair (BER) that detects and signals DNA strand breaks, facilitating their further repair (Zhao et al., 2022; Li et al., 2023). Overall and considering all the above indicated, MEL is recognized as a broad-spectrum antioxidant (Reiter, 1996; Zhang and Zhang, 2014; Johns and Platts, 2014; Tan et al., 2002; Poeggeler et al., 2002; Anisimov et al., 2006; Bai et al., 2020). Despite the well-established interaction of MEL with ROS and RNS, recent reports suggest that MEL’s direct free radical scavenging ability may be overestimated (Boutin et al., 2024).
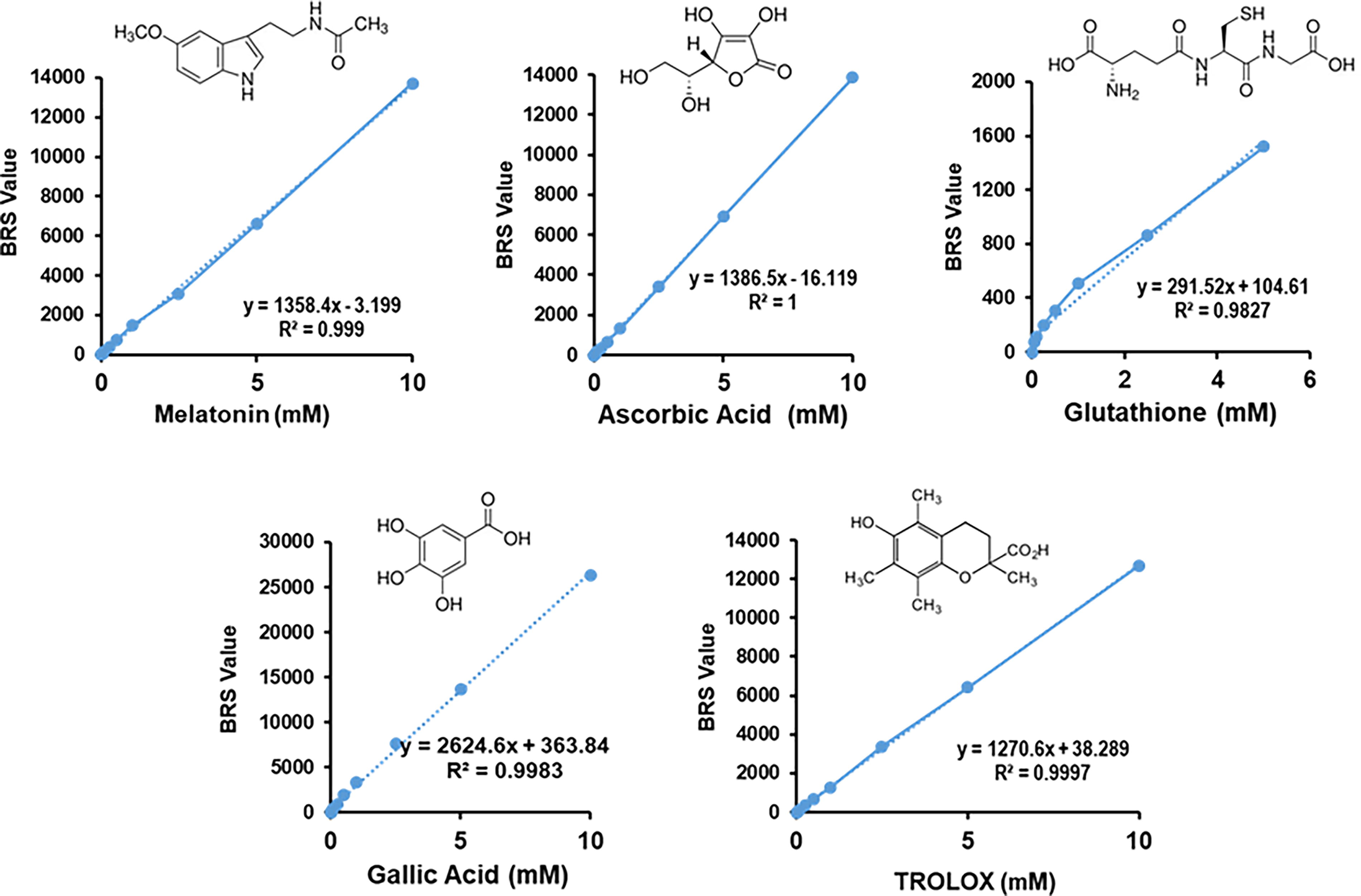
This critique suggests a need for detailed studies focusing on MEL’s in situ concentrations and interactions within specific subcellular contexts. Factors influencing antioxidant capacity include proximity to pro-oxidative events and the combined effects of all antioxidant compounds present in cellular extracts. Furthermore, due to the extreme difficulty with measuring the in vivo scavenging ability of an antioxidant, it is important to consider factors that can influence these contradictory views, such as the in situ MEL concentration in a specific subcellular compartment and its proximity to the production site of the generated pro-oxidative event. Figure 6 presents a recent comparative analysis of the total antioxidant capacity (TAC) of MEL against well-known antioxidants, including natural compounds like vitamin C, glutathione, and gallic acid, as well as the reference compounds like Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid), which is a water-soluble analog of vitamin E (Corpas et al., 2024). Thus, it can be observed that the values obtained for a specific concentration are very similar to those for ascorbic acid or Trolox. For example, at concentration of 1 mM, MEL has an antioxidant capacity of 1500, whereas 1 mM ascorbate has an antioxidant capacity of 1300 (Corpas et al., 2024). Being this true, it is important to also consider that in a cellular context, as it occurs with crude extracts, the antioxidant capacity results from the combined effects of all compounds with such potential and their relative concentrations. These factors may vary depending on the physiological conditions or in response to specific adverse conditions. In any case, this issue can be the focus of future research using thermodynamic, physicochemical, and molecular approaches.
Plant MEL Receptor
Some authors refer to MEL as a new phytohormone (Hardeland, 2014; Ludwig-Müller and Lüthen, 2015). As such, a possible receptor, called CAND2/PMTR1 (G protein-coupled receptor candidate 2/phytomelatonin receptor), has been identified in A. thaliana (Wei et al., 2018; Cheng et al., 2024), unveiling its signaling potential in plants. This MEL receptor (MelR) was believed to have a regulatory function in stomatal closure, which in turn regulates the entry of Ca2+ ions and the production of H2O2. However, the exact subcellular location of this receptor was questioned (Lee and Back, 2020). More recently, homologs to the Arabidopsis MelR have been preliminarily studied in other plant species, including maize (Zea mays) (Wang et al., 2022), alfalfa (Medicago sativa L.) (Yu et al., 2021), tobacco (Nicotiana benthamiana) (Kong et al., 2021; Banerjee and Roychoudhury, 2024), cassava (Manihot esculenta) (Bai et al., 2022), and rice (O. sativa) (Barman et al., 2023). Thus, through molecular approaches using both defective and overexpressing mutants of this MelR, it has been demonstrated in different plant species that this receptor mediates the action of MEL under physiological conditions as well as in response to abiotic and biotic stresses (Kong et al., 2021; Yu et al., 2021; Cheng et al., 2024). In tobacco (Nicotiana tabacum), the MelR has been shown to function upstream of ROS generation in guard cells (Banerjee and Roychoudhury, 2024). In tomato plants, the MYC2 transcription factor mediates the crosstalk between jasmonic acid and MEL by regulating MelR expression during wound-induced adventitious root regeneration (Liu et al., 2025).
In silico analysis of the rice MelR (OsPMTR) using transmembrane predictor software indicates the presence of seven transmembrane domains containing protein resembling G-protein-coupled receptors (Barman et al., 2023). Additionally, there is a long N-terminal loop and a long transmembrane domain at the C-terminal (Fig. 7A).
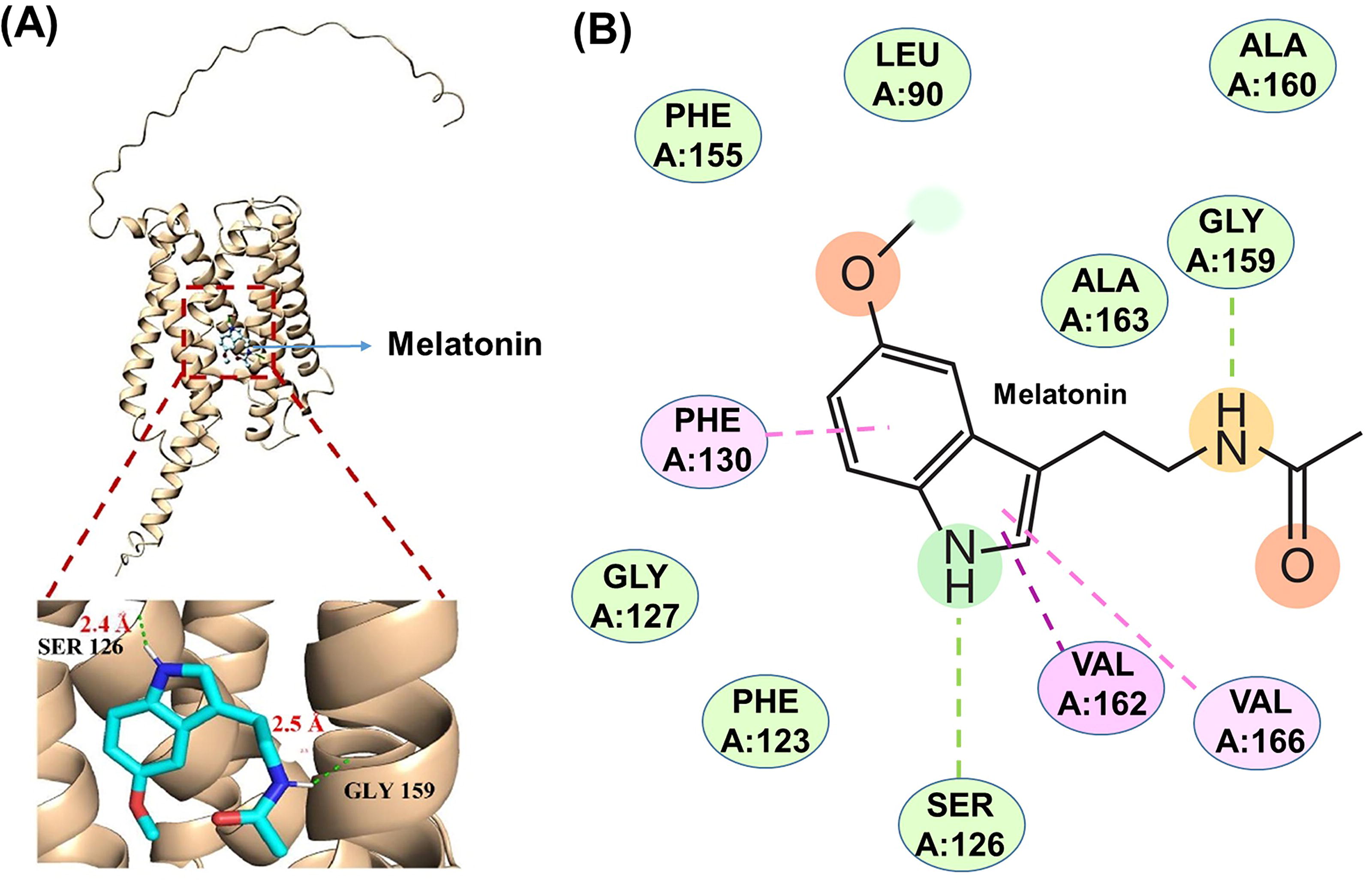
Docking analysis of the MelR interaction with MEL indicates that residues Ser126 and Gly159 establish hydrogen bonds with distances of 2.4 and 2.5 Å, respectively. A more detailed analysis of the MEL-receptor interaction highlights the role of the amino acids Phe130, Val162, and Val166 to stabilize various types of π (pi) interactions. These include (i) conventional hydrogen bonds with Ser126 and Gly159; (ii) π-π stacking interactions with Phe130, which are noncovalent π interactions (orbital overlap) between the π bonds of the aromatic rings in MEL; (iii) π-alkyl interactions, which occur between a π-electron cloud of an aromatic group (Phe130) and the electron cloud of an alkyl group (Val162 and Val166); (iv) π-sigma interactions, which involve the interaction between positive and negative charge sites on different atoms; and (v) van der Waals interactions, which occur when adjacent atoms come close enough for their outer electron clouds to almost interact (Fig. 7B). Although further studies are needed to clarify the signaling cascade that triggers the interaction between MEL and its receptor, it is clear that this opens new avenues of research into the biochemical and molecular mechanisms of MEL action in higher plants.
MEL and Gene Regulation: Transcription Factors and microRNA
Another area of great interest, where continued progress is being made, is the identification of how MEL exerts its regulatory mechanisms involving transcription factors (TFs) and target genes (Kaya and Ugurlar, 2023). Table 1 summarizes representative examples of MEL’s target TFs and genes, as well as the effects exerted in different plant species. Most gene identifications have been made when MEL is added exogenously, commonly at micromolar concentrations that are nonphysiological. However, this does not rule out the possibility that endogenous MEL exerts fine control over those TFs and target genes. Remarkably, MEL influences a wide range of genes involved in its own biosynthesis (Wei et al., 2016), the biosynthesis of anthocyanins (Zhang et al., 2016), abscisic acid (ABA), and genes related to resistance to various stresses, including antioxidant systems (Chen et al., 2017; Chang et al., 2021). Additionally, MEL affects multiple organs, including roots, leaves, flowers, and fruits.
Examples of How Melatonin, Either Endogenous or Exogenously Applied, Affects the Expression of Different Genes and/or Transcription Factors
ABA, abscisic acid; ASMT, N-acetylserotonin O-methyltransferase; AXR3, AUXIN RESISTANT 3; ABC, ATP-binding cassette; AZF2, Arabidopsis Cys2/His2 zinc-finger protein 2; bHLH, basic helix-loop-helices TF; C3H12, CCCH-type zinc-finger protein; C4H, cinnamic acid 4-hydroxylase; CAMTA1, calmodulin binding transcription activator 1; CBFs, C-repeat-binding factors; CHI, chalcone isomerase; CHS, chalcone synthase; CNGC2, cycle nucleotide-gated ion channel 2; COR15a, cold-regulated gene; DFR, dihydroflavonol 4-reductase; DREB1s, drought response element-binding 1 factors; DREBs, drought response element-binding factors; F3′H, flavonoid 3′-hydroxylase; F3H, flavanone 3-hydroxylase; FLC, flowering locus C; GAE6, UDP-glucuronate 4-epimerase 6; GST, glutathione S-transferase; HMA4, heavy metal ATPase 4; HSFA1s, class A1 heat-shock factors; IAA17, indole-3-acetic acid inducible 17; LDOX, leucoanthocyanidin dioxygenase; MYB, myeloblastosis TF; MYC2, myelocytomatosis oncogene; PAL, phenylalanine ammonia-lyase; PCR2, Plant cadmium resistance 2; PDR8, pleiotropic drug resistance 8; PME61, pectin methylesterase 61; RBOH, respiratory burst oxidase homolog; ROS, reactive oxygen species; SAG12, SENESCENCE-ASSOCIATED GENE 12; SEN4, SENESCENCE 4; SNAT, serotonin N-acetyltransferase; SOS1, Salt Overly Sensitive1; TDC, tryptophan decarboxylase; T5H, tryptamine 5-hydroxylase; UFGT, UDP-glucose: flavonoid 3-O-glucosyltransferase; WD40, TF containing WD repeat motifs, which is composed of 44–60 amino acid residues; ZAT, zinc finger of Arabidopsis thaliana TF.
Figure 8 illustrates a model of how exogenous MEL exerts its effects through different target genes, which also interact with each other as well as with diverse TFs. Thus, in watermelon (Citrullus lanatus L.) seedlings, exogenous MEL enhances cold tolerance through two complementary pathways, involving an increase in both H2O2 and cytosolic Ca2+ content. This occurs via the upregulation of the respiratory burst oxidase homolog (RBOH) D and cyclic nucleotide-gated channel (CNGC) 2 genes, respectively, which ultimately activate the C-repeat-binding factor (CBF) pathway (Chang et al., 2021). Additionally, these two pathways can reciprocally be activated, thus potentiating the effect of MEL. These CBF TFs, along with dehydration-responsive element-binding (DREB) factors (Zhou et al., 2020), belong to the APETALA2/ethylene-responsive element-binding factor (AP2/ERF) family, which is involved in freezing tolerance and cold acclimation (Hu et al., 2020). The CBF proteins usually contain a highly conserved DNA-binding domain (AP2 domain) and a putative nuclear localization signal sequence in the N-terminal region. It is also important to note that the expression of CBFs is regulated by gibberellic acid (GA), jasmonic acid (JA), ABA, ethylene (ETH), and brassinosteroids (BRs) (Barrero-Gil and Salinas, 2017). Consequently, MEL could serve as an additional regulatory component in this pathway, potentially interacting with other phytohormones (Ameen et al., 2024; Jahan et al., 2024).

MicroRNAs (miRNAs) are a class of noncoding single-stranded RNAs, ∼22 nucleotides in length, that function as posttranscriptional regulators of gene expression in both plants and animals (Dong et al., 2022). There is now evidence of the interaction between MEL and some miRNAs to regulate the expression of certain target genes in higher plants (Petijová et al., 2020). The first indications of miRNA involvement in MEL mediation were obtained in watermelon. Using high-throughput miRNA and mRNA sequencing approaches, MEL was found to exert differential regulation of a group of miRNAs that influence the expression of genes that affect signal transduction, including CDPK (Ca2+-dependent protein kinase), bHLH (basic helix-loop-helix motif), WRKY, MYB, and DREB, as well as genes involved in detoxification of ROS and protection against environmental stress like low temperature or heavy metal, such as LEA (late embryogenesis abundant) and MDHAR (monodehydroascorbate reductase). This regulation confers watermelon (Cucumis melo L.) plants with higher tolerance to cold. Among the downregulated miRNAs, miR159-5p, miR858, miR8029-3p, and novel-m0048-3p were reported (Li et al., 2016). Similarly, in alfalfa (M. sativa L.) plants exposed to cadmium stress, it has been found that the exogenous application of MEL enhances their tolerance. Thus, among the different target genes, MEL was observed to downregulate the expression of miR398a and miR398b, which allowed the upregulation of their target gene that codes for the antioxidant enzyme CuZn-SOD (Gu et al., 2017). Complementary studies on how MEL confers cadmium tolerance in radish (Raphanus sativus) plants led to the identification of additional miRNA target genes that affect cadmium transport, including ABC transporter genes such as miR162a-targeted ABCB21, miR172c-targeted ABCG39, and miR156j-targeted YSL7 (yellow stripe-like transporter 7) (Xu et al., 2020). In tomato plants, MEL has been found to delay leaf senescence in darkness due to reduced starch degradation, which is attributed to the induction of miR171b expression. This miRNA inhibits the α-glucan water dikinase (GWD) gene, the first gene in the starch degradation pathway (Wang et al., 2022).
In Chinese cabbage (Brassica rapa) exposed to higher than normal salinity, greater tolerance has been linked to miR168a, which targets the O-methyltransferase 1 (OMT1) gene. This gene encodes a protein that catalyzes the transfer of a methyl group onto N-acetylserotonin, producing MEL (Shamloo-Dashtpagerdi et al., 2022). Furthermore, it was found that miR168a expression differs among B. rapa genotypes, suggesting that it may contribute to the different salinity tolerance of these genotypes. During the germination of maize (Z. mays) seeds exposed to elevated saline concentrations, the application of exogenous MEL improves the germination percentage. Analysis of the differential expression of microRNAs identified 17 differentially expressed miRNAs (9 upregulated and 8 downregulated). Some of these miRNAs were associated with mechanisms of response to salinity, such as miR159a/c, miR167a/e/g, and miR171d. Additionally, MYB TFs were identified as targets of miR159a, miR167g, miR319a, and novel_183, along with genes from the TCP (TEOSINTE BRANCHED1, CYCLOIDEA, PROLIFERATING CELL FACTOR 1 and 2), AP2, bHLH, C2H2 (Cys2-His2), ERF, and LSD (lesion-simulating disease) TF families (Xu et al., 2024).
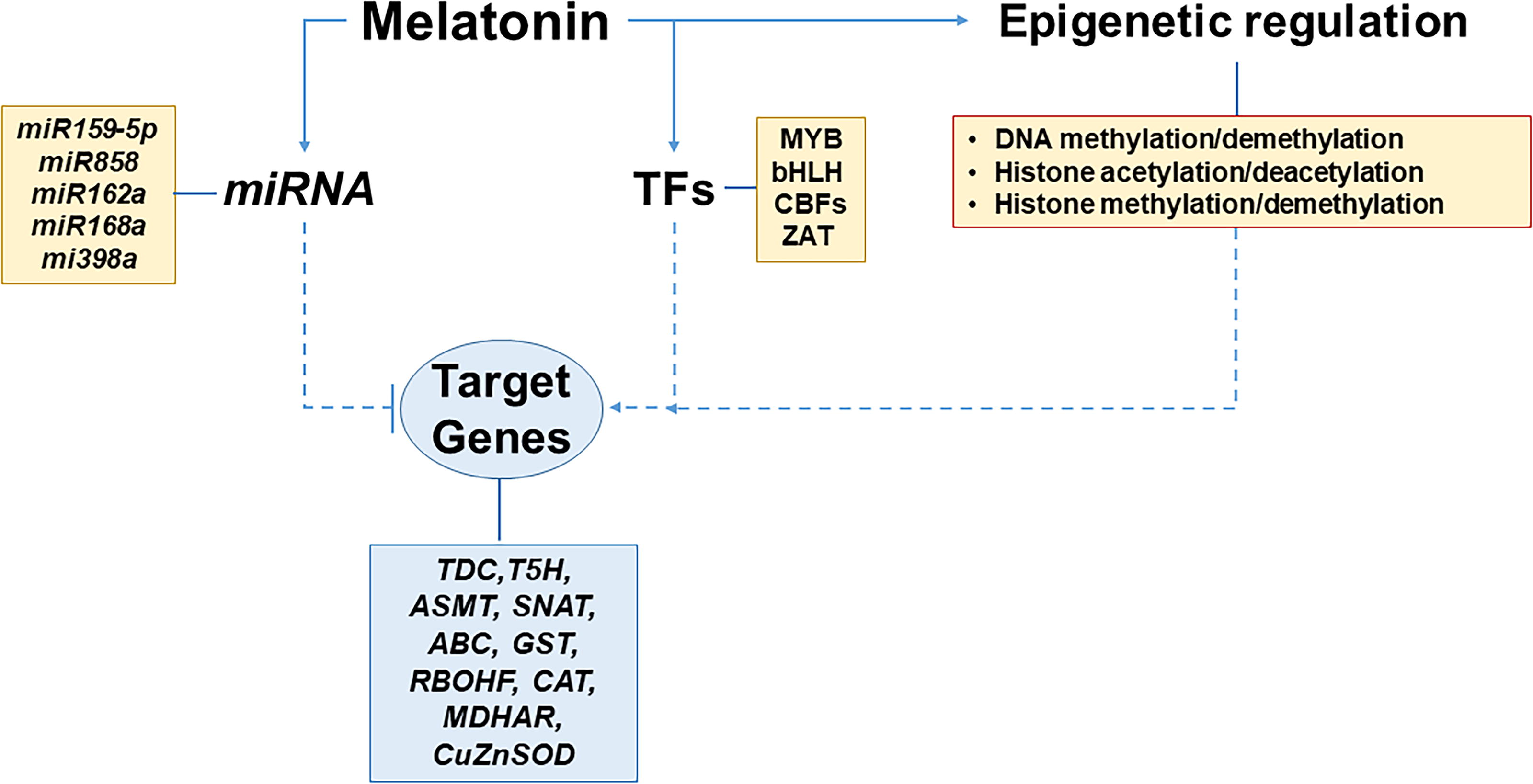
In biotic stress, the involvement of miRNA and MEL has also been observed. In rice infected with the rice black-streaked dwarf virus (RBSDV), a MEL receptor was identified as a target gene of miR-315-5p. Its downregulation protects against viral infection in the early stages, while its upregulation promotes viral infection (Zhang et al., 2021). Additionally, MEL influences gene expression at the epigenetic level by regulating DNA methylation and demethylation, histone acetylation and deacetylation, as well as histone methylation and demethylation (Tang et al., 2021; Shan et al., 2022; Zhao et al., 2023). Figure 9 depicts a model of how these mechanisms, where MEL is involved, operate.
Function of MEL in Plant Development and Growth
MEL’s role in regulating plant growth
Because MEL and auxin share a similar molecular structure, their potential interactions have been widely investigated. It appears that MEL plays a broader developmental role, regulating processes such as flowering, fruit ripening, seed germination, and overall plant growth (Byeon et al., 2014; Khan et al., 2023; Arnao and Hernández-Ruiz, 2020). Studies have suggested that MEL may influence plant development by interacting with redox networks or modulating the activity of other plant hormones. Evidence indicates that MEL works in tandem with phytohormones like auxins to enhance its effects (Arnao and Hernández-Ruiz, 2018, 2020).
Since its discovery in plants (Dubbels et al., 1995; Hattori et al., 1995), MEL has been recognized as essential for controlling various aspects of plant development. Early research by Hernández-Ruiz et al. (2005) highlighted MEL’s direct role in promoting growth by demonstrating its ability to extend the coleoptiles in grasses like canary grass (Phalaris canariensis), wheat (Triticum aestivum), barley (Hordeum vulgare), and oats (Avena sativa). Similarly, vegetative growth has been observed in other plants such as soybean (Glycine max), maize (Z. mays), cucumber (Cucumis sativus), and Arabidopsis when treated with MEL (Hernández-Ruiz et al., 2004; Arnao and Hernández-Ruiz, 2007; Posmyk et al., 2009; Wei et al., 2015).
Although MEL promotes growth, questions remain regarding the concentrations used in these studies, which often exceed physiological levels found naturally in plants. Reduced MEL levels, as seen in Arabidopsis and rice plants with downregulated SNAT1 or SNAT2 genes, have been linked to stunted growth and reduced biomass (Hwang and Back, 2018; Lee and Back, 2019). Conversely, increasing MEL levels through genetic modifications, such as expressing sheep SNAT in rice, has resulted in faster seedling growth and earlier flowering, though with unexpected reductions in grain production (Byeon et al., 2014). Similar to the plant hormone auxin, particularly IAA, MEL has been shown to stimulate the elongation of plant structures like hypocotyls when applied in micromolar concentrations. In fact, MEL’s auxin-like activity has been found to be 63% more effective than IAA in promoting plant growth. However, at higher doses, it acts as an inhibitor, as observed in studies on etiolated lupin (Lupinus albus) seedlings. Transgenic rice seedlings overexpressing serotonin N-acetyltransferase (SNAT) have demonstrated a twofold increase in root growth, correlating with significantly higher endogenous MEL levels compared to wild-type plants, suggesting a strong link between MEL concentration and growth rates (Park et al., 2012).
MEL’s control of seed germination
One of MEL’s notable effects is its ability to enhance seed germination. It protects seeds during germination by stabilizing cell membranes and preventing oxidative damage to lipids, thus improving seed vitality and germination rates (Posmyk et al., 2009). Additionally, MEL influences the cell’s metabolic pathways, particularly gluconeogenesis, aiding in the conversion of amino acids into soluble sugars to fuel germination (Kobylińska et al., 2018).
Seeds of various plant species contain significant levels of endogenous MEL, indicating its role as a natural phytohormone involved in seed physiology. The concentration of MEL varies widely among different plant species and even among different seed types within the same species, suggesting species-specific roles in seed development and germination. Endogenous MEL levels fluctuate during seed development, with higher concentrations often observed during critical phases such as maturation and germination.
During germination, MEL supports sugar metabolism by modifying the gluconeogenesis pathway, facilitating the conversion of amino acids into soluble sugars, which provides energy for seedling growth (Kobylińska et al., 2018). Moreover, studies on soybeans demonstrate that MEL significantly boosts seedling development and yield by improving photosynthetic efficiency and glucose metabolism (Wei et al., 2015). Similarly, MEL enhances wheat yield by increasing the number of ears per plant and strengthening the root system, which improves carbon uptake (Ye et al., 2020).
The previously discussed positive effects of MEL on seed germination were found to be dose-dependent. For instance, early research by Posmyk et al. (2008, 2009) on red cabbage and cucumber seeds revealed that low exogenous MEL enhanced seed germination, while high MEL concentrations inhibited it. The researchers suggested that MEL protected membrane structures from peroxidation during stress, but excessive MEL (around 4 μg/g fresh weight in cucumber) triggered oxidative changes in proteins such as S-sulfenylation. In Arabidopsis, low concentrations of exogenous application of MEL (10 or 100 μM) had no significant impact on seed germination, while higher concentrations (500 or 1000 μM) reduced it. Similar inhibitory effects of high MEL concentrations on seed germination have been reported in several plant species, such as cucumber (Zhang et al., 2013), cotton (Xiao et al., 2019), and melon (C. melo L.) (Castañares and Bouzo, 2019). However, more recent studies have indicated that 1000 μM MEL can enhance seed germination in maize under salt stress (Hussain et al., 2024), and 500 μM MEL improved seed germination in rice under drought stress (Li et al., 2022), suggesting that MEL’s effects may be specific to the type of abiotic stress. Furthermore, a study showed that adding 300 μM MEL to the cultivation solution increased the germination rate of two wheat varieties but decreased it in another, highlighting that MEL’s effects on seed germination may also vary based on genotype (Zhang et al., 2022).
High MEL concentrations may inhibit seed germination by increasing endogenous ABA levels, which can promote seed dormancy or suppress seed germination, depending on ABA-sensitive factors, like ABA overly sensitive 5 (ABO5) (Huang et al., 2017), or reduce seed germination via suppressing the GA synthesis (Lv et al., 2021). MEL can be pro‐oxidant at higher doses and can induce oxidative changes in protein and amino acids (Anisimov et al., 2006), while MEL also positively regulates the internal ROS levels in seeds, which are crucial to stimulate the breakage of seed dormancy and initiation of seed germination under stressful conditions (Arnao and Hernández-Ruiz, 2014; Li et al., 2022).
Abiotic stresses can adversely affect seed quality and germination potential, leading to reduced crop yields. MEL has been shown to mitigate the negative impacts of these stresses by enhancing seed vigor and promoting germination. For example, tomato seeds accumulate high levels of MEL during the early germination phase, which improves their resilience (Okazaki and Ezura, 2009). Cabbage seeds treated with exogenous MEL showed increased germination rates from 72% to 88%, while cucumber seeds treated with MEL in cold conditions exhibited germination rates as high as 98%, compared to just 4% without treatment (Posmyk et al., 2008).
MEL also mitigates inhibitory effects caused by stressors such as polyethylene glycol (PEG) during germination. Cotton and other crops, as well as Arabidopsis, have exhibited enhanced germination under various stress conditions when treated with optimal concentrations of MEL. For example, in cotton seeds, a 20 µM MEL dose was most effective at promoting germination, while in rice, 100 µM MEL improved heat tolerance and boosted germination rates under high temperatures (Yu et al., 2022a; Xiao et al., 2019).
In summary, MEL demonstrates a remarkable ability to improve seed germination by regulating stress responses, supporting metabolic pathways, and interacting with other hormones like auxins and gibberellins. This highlights its potential as a powerful biostimulant in sustainable agriculture.
MEL’s role in controlling root development
MEL structural similarity to IAA, a key auxin, has prompted extensive research into MEL’s role in shaping root architecture. Over the past few decades, studies have revealed that MEL promotes both lateral and adventitious root formation across various plant species, including L. albus, rice, cucumber, and Arabidopsis (Arnao and Hernández-Ruiz, 2007; Chen et al., 2019). For instance, Arabidopsis plants treated with MEL exhibited up to three times more lateral roots compared to untreated controls (Pelagio-Flores et al., 2012). Thus, MEL’s rhizogenic (root-promoting) effects have been well-documented across various species, including rice, pomegranate (Punica granatum), cherry rootstocks, cucumber, and others. For instance, MEL has been observed to stimulate the development of lateral and adventitious roots from pericycle cells in lupin plants, a pattern consistent with findings in several other species (Sarropoulou et al., 2012a; Zhang et al., 2013).
On a molecular level, MEL enhances nitrogen uptake and metabolism, stimulating enzymes critical for nitrogen absorption. This leads to improved crop productivity, as demonstrated in wheat, where MEL treatment significantly boosted yield by optimizing nitrogen utilization by roots (Qiao et al., 2019). MEL also supports carbon and nitrogen balance, improving maize growth by regulating the activity of enzymes involved in these metabolic pathways, such as Rubisco, nitrate reductase, nitrite reductase, glutamine synthase, glutamate 2-oxoglutarate transferase, and NADH-glutamate dehydrogenase (Erdal, 2019). Additionally, and due to its role in regulating ROS in plants, MEL influences the production of H2O2 and other ROS, which act as signaling molecules to stimulate lateral root formation. Thus, it has been shown that MEL promotes O2 •− production by the RBOH system, triggering H2O2 generation, thus enhancing lateral root development in crops like tomato and alfalfa (Chen et al., 2018, 2019). Studies also show that MEL’s effects on root development vary with concentration. Low doses promote root growth, while higher concentrations can inhibit it. In commercial cherry rootstocks, a 1 µM MEL treatment improved root number and length, while a 10 µM concentration hindered rooting (Sarropoulou et al., 2012b). Similar dose-dependent effects were observed in lupin, where low MEL levels stimulated adventitious and lateral roots, but higher doses were inhibitory.
Under stress conditions, MEL enhances root resilience. For example, MEL application increased root biomass and antioxidant activity in rape seedlings under drought stress, aiding osmotic adjustment and improving survival rates (Zeng et al., 2018). Similarly, in apple rootstocks, MEL mitigated low nitrate stress by reshaping root structure and promoting nitrogen uptake through the regulation of nitrogen transport genes (Du et al., 2021).
The interplay between MEL, auxins, and NO plays a critical role in its regulatory functions in root growth and development. NO modulates auxin responses, influencing lateral root formation, root hair growth, and gravitropism. MEL interacts with these signaling pathways, enhancing their combined effects on root development (Sanz et al., 2015). For example, studies on lupin roots showed that MEL induced root tropism effects, resulting in negative tropism that caused roots to grow away from the MEL application site (Arnao and Hernández-Ruiz, 2017).
Through its involvement in ROS signaling, hormonal crosstalk, and stress mitigation, MEL serves as a vital regulator of root growth, further demonstrating its potential as a tool for enhancing crop resilience and productivity in sustainable agriculture.
The role of MEL in flowering
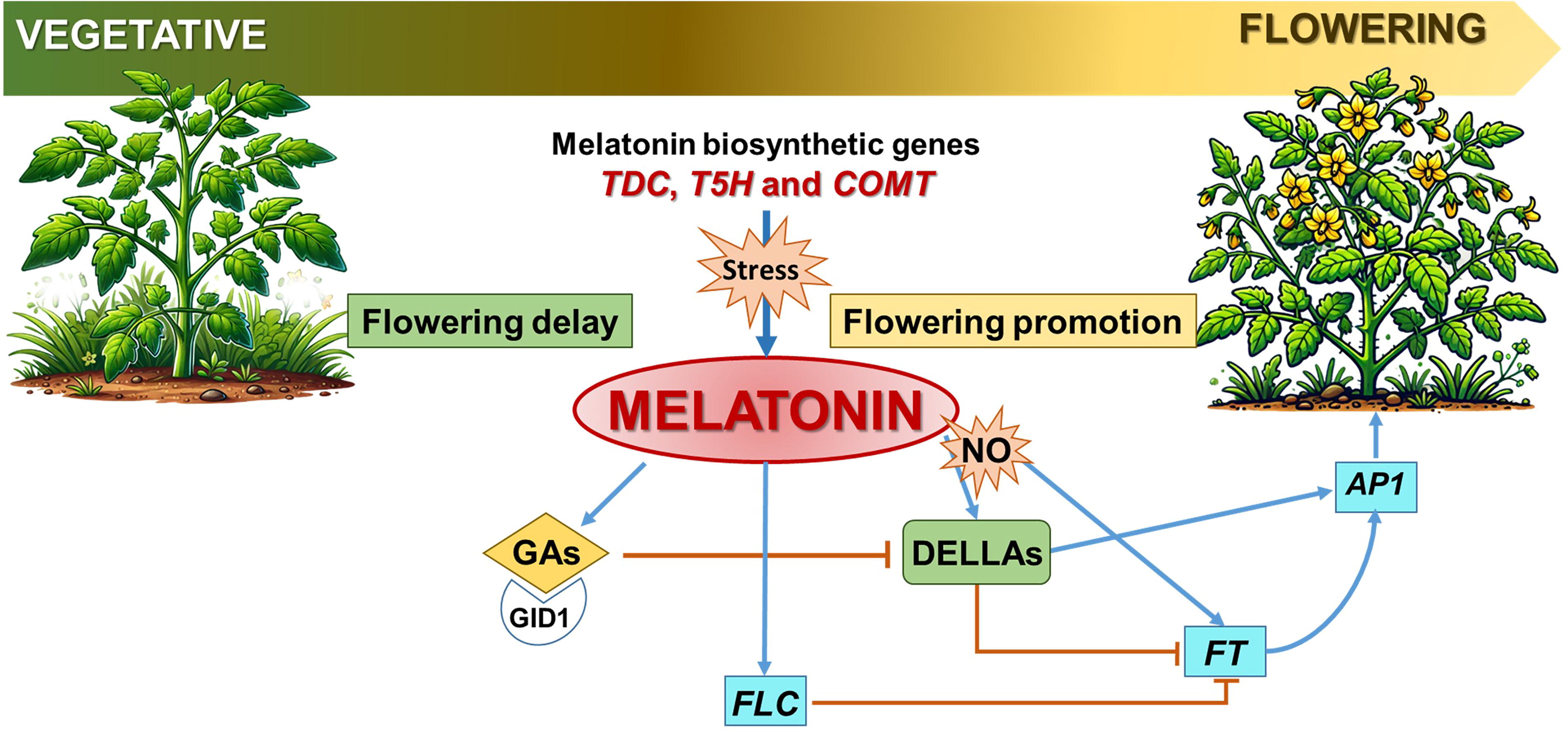
The transition from vegetative growth to flowering is a critical phase in the life cycle of plants, and MEL has emerged as an influential factor in this process. Several studies have shown that GA signaling, a plant hormone essential for reproductive development, interacts with MEL signaling to regulate flowering. Studies in rice have shown that MEL biosynthesis increases significantly, up to sixfold, in the panicle (flower structure) compared to the flag leaves during flowering. This surge correlates with the activation of MEL biosynthesis enzymes like TDC, T5H, and COMT, suggesting MEL’s involvement in floral development (Park et al., 2013a). According to Shi et al. (2016b), snat2 plants with low endogenous MEL levels have reduced GA content, thus showing a link between MEL and GAs.
One of the first studies to link MEL to flowering was conducted by Shi et al. (2016b), which identified that MEL regulates floral transition through its impact on DELLA proteins and flowering locus C (FLC) in Arabidopsis. It is well known that DELLA proteins act as growth repressors, while GA promotes growth by inducing DELLA degradation. Thus, exogenous MEL treatments were found to stabilize DELLA proteins, slowing the transition to flowering without altering DELLA transcript levels. This indicates that MEL influences flowering independently of endogenous GA accumulation (Mukherjee, 2019). MEL’s role in flowering extends beyond hormone regulation. For instance, Murch and Saxena (2002) demonstrated that MEL acts as a protective agent during flower development in Hypericum perforatum L. High concentrations of serotonin and MEL were detected at specific stages of flower development, which coincided with the highest regenerative potential of isolated anthers. This suggests that MEL helps plants adapt to stress and ensures successful reproduction. In Devil’s Trumpet (Datura metel L.) flowers, MEL content was highest in the early stages of bud development but decreased steadily as the buds matured, indicating its dynamic role throughout floral maturation (Murch et al., 2009). Similarly, MEL increased GA levels in cotton seedlings and induced parthenocarpy (fruit development without fertilization) in pears (Xiao et al., 2019; Liu et al., 2018).
While MEL generally promotes vegetative and reproductive growth, its effects on flowering can vary depending on environmental conditions and stressors. For example, MEL delays flowering in some cases by regulating stress responses while simultaneously enhancing the expression of GA biosynthesis genes in others (Wang et al., 2022; Kolář et al., 2003). Furthermore, in apple trees, exogenous MEL has been shown to delay flowering, especially under specific light conditions such as higher far-red and blue light levels (Zhang et al., 2019). This dual role highlights the complexity of MEL’s interactions with plant developmental pathways.
In summary, MEL not only facilitates flowering by interacting with key hormones and biochemical pathways but also acts as a protective agent against environmental stress during reproductive development. Its multifaceted role underscores its potential application in improving crop resilience and optimizing flowering in agricultural systems (Fig. 10).
The role of MEL in fruit development, ripening, and senescence
MEL has a pivotal role in regulating fruit growth, ripening, and delaying senescence through its antioxidant properties and interactions with other plant hormones. Different studies have shown that MEL concentrations vary throughout fruit development, often peaking during critical phases to mitigate oxidative stress and support proper growth (Fig. 11). Early research highlighted MEL’s presence in Datura metel fruits during the initial 10 days postanthesis, with the highest concentrations found in the ovule. Similar trends have been observed in cherry (Prunus avium) fruits, where MEL levels peaked during embryo development and endocarp lignification. This accumulation correlated with increased ROS, suggesting MEL’s role in maintaining redox balance during these developmental stages (Zhao et al., 2013).

In tomatoes, MEL levels rose significantly in the final stages of fruit maturation, coinciding with color changes from green to red. This pattern was also observed in Merlot grapes and pepper cultivars, where higher MEL levels in mature fruits and seeds helped protect these organs from environmental damage (Okazaki and Ezura, 2009; Korkmaz et al., 2014). Mature seeds of hemp (Cannabis sativa) contained 4–10 times more MEL than other plant tissues, further emphasizing its protective role during reproductive phases (Allegrone et al., 2019).
The interaction between MEL and ethylene plays a crucial role in the ripening processes of fruit. Thus, during early ripening, MEL may enhance ethylene production by promoting precursor availability or enzyme activity; however, in later stages, it may inhibit ethylene synthesis, delaying senescence and prolonging shelf life. In tomatoes, treatment with MEL at a concentration of 50 µM significantly enhanced key ripening characteristics, including lycopene content, fruit texture, and flavor. This treatment also upregulated several genes related to ethylene signaling, specifically ethylene receptors (ETR4) and TFs (EIL1, EIL3, and ERF2), as well as the Never Ripe (NR) gene. As a result, there was an increase in levels of sucrose, glucose, and citric acid, which are vital for flavor and overall fruit quality. Overall, tomato plants treated with MEL exhibited a 13% increase in both fruit yield and quality compared to the control group (Sun et al., 2015; Liu et al., 2016).
Similarly, in grape berries, MEL stimulated ethylene production, which in turn promoted polyphenol accumulation, enhancing antioxidant activity and hastening ripening (Xu et al., 2018). However, MEL can also delay ripening and senescence under certain conditions. For example, in pears and bananas, MEL reduced ethylene production and the reactive oxygen burst, delaying ripening and extending shelf life. This effect was mediated by the downregulation of ethylene biosynthesis enzymes such as 1-aminocyclopropane-1-carboxylic acid (ACC) synthase (ACS) and ACC-oxidase (ACO) (Zhai et al., 2018; Hu et al., 2017).
MEL’s antioxidant capacity makes it an effective agent for prolonging postharvest fruit quality. For example, MEL-treated peaches exhibited reduced weight loss, decay, and respiration rates, along with higher firmness, total soluble solids, and ascorbic acid content. These effects are attributed to MEL’s ability to mediate antioxidant pathways, reducing oxidative stress markers like malondialdehyde (MDA) and preserving fruit texture and flavor (Gao et al., 2016). In strawberries, MEL enhanced polyphenol levels, antioxidant activity, and ATP content, further delaying senescence and maintaining fruit quality during storage (Aghdam and Fard, 2017).
In plums, MEL treatment increased ascorbic acid and phenolic content while reducing pericarp browning and oxidative damage, effectively extending the fruits’ postharvest lifespan (Bal, 2019). Similarly, MEL delayed litchi fruit senescence by modulating redox balance and repairing oxidatively damaged proteins. Thus, there are experimental data indicating that MEL can maintain the cellular redox balance and protect protein structures, which reduces protein carbonylation. Furthermore, MEL binds to transition metals like iron and copper, lowering their participation in Fenton reactions that generate reactive hydroxyl radicals (Mayo et al., 2003). In bananas, concentration-dependent MEL treatments (200–500 µM) significantly slowed ripening by suppressing ethylene production and maintaining fruit quality (Hu et al., 2017). Tomatoes treated with 50 μM MEL exhibited significant changes in fruit ripening characteristics compared to untreated tomatoes, including enhanced lycopene levels, increased fruit softness, improved flavor, and elevated ethylene signaling and biosynthesis enzyme activity by the upregulation of the expression of genes related to ethylene receptors and transducing elements (Sun et al., 2015). Following MEL treatment, sucrose and glucose levels increased, while soluble galactose concentrations rose as the fruits ripened, stimulating ethylene production and promoting ripening.
A recent study has shown that treating Hami melon fruit with 0.5 mM MEL can delay its softening. This treatment has an inhibitory effect on the activity of enzymes that are involved in the structure of the cell wall, such as polygalacturonase, cellulose, β-galactosidase, β-glucosidase, and xyloglucan endotransglycosylase. Additionally, MEL helps preserve the quality of the fruit by maintaining the carbohydrate metabolism, resulting in a high content of starch and soluble sugars. These sugars include sucrose, sorbitol, glucose, fructose, maltose, and galactose, which all contribute to maintaining the fruit’s firmness (Wang et al., 2024). In summary, MEL is a key regulator in fruit development and ripening, balancing oxidative stress, modulating hormone pathways, and delaying senescence. Its application in agriculture offers promising solutions for improving fruit quality and extending shelf life, aligning with sustainable farming practices.
MEL treatment has been shown to significantly delay senescence and improve the postharvest quality of various fruits by reducing degradation markers, slowing ripening, and preserving key nutrients. For example, in peaches, MEL administration reduced weight loss, decay incidence, and respiration rate while preserving firmness, total soluble solids, and ascorbic acid levels, demonstrating its potential for postharvest quality preservation (Gao et al., 2016). Similarly, MEL delayed senescence and enhanced antioxidant activity in strawberries while increasing the expression of MEL biosynthesis genes, boosting endogenous MEL levels (Aghdam and Fard, 2017; Liu et al., 2018). Furthermore, in grapes, MEL increases ethylene production, promoting polyphenol accumulation and antioxidant activity, which are vital for fruit quality (Xu et al., 2017). In contrast, MEL treatment in pears, plums, and litchi has been found to reduce ethylene production, suppress senescence, and maintain fruit quality during storage (Zhai et al., 2018; Liu et al., 2018; Bal, 2019).
In bananas, MEL treatment, particularly at 200 and 500 μM, slowed ripening, with the effects being concentration-dependent (Hu et al., 2017). These effects suggest that MEL’s influence on ethylene biosynthesis enzymes (such as ACO and ACS) can either promote or delay ripening depending on the fruit type. In some fruits like tomatoes and grapes, MEL facilitates ripening by increasing ethylene production, while in others, such as pears, strawberries, and bananas, it downregulates ethylene biosynthesis, extending shelf life and delaying senescence (Liu et al., 2018). Thus, the efficacy of MEL in delaying senescence and extending postharvest fruit quality is dependent on factors such as the fruit’s physiological state, MEL concentration, and treatment duration. Overall, MEL holds promise as a method for improving postharvest fruit shelf life and maintaining quality across a variety of species.
Table 2 provides a summary of the MEL content found in various plant species, including seeds, vegetables, and fruits. The values can vary due to several factors, such as the plant cultivar, environmental conditions, the year of harvest, and the analytical techniques used for measurement. These techniques include liquid chromatography–electrospray ionization mass spectrometry, high-performance liquid chromatography with a fluorescence detector, ultra-performance liquid chromatography–mass spectrometry, and enzyme-linked immunosorbent assay.
Melatonin Content Across Different Seeds, Vegetables, and Fruits
The values in nanograms per gram of fresh weight (FW) provided are indicative, as melatonin levels can be influenced by various factors, including plant cultivar, environmental conditions, harvest year, and the analytical techniques used for measurement.
aIndicate the content of MEL expressed in ng · g−1 dry weight.
MEL, melatonin.
Crosstalk between MEL and other phytohormones in the control of plant growth, seed germination, and fruit ripening
MEL interacts with various phytohormones, influencing growth, development, and stress responses in plants. Initially, researchers focused on the relationship between MEL and auxins, given their structural similarities. However, recent studies have highlighted MEL’s interplay with a wide range of plant hormones, including gibberellins (GAs), cytokinins (CKs), ABA, and ethylene, as well as more recently discovered signaling molecules like JAs, salicylic acid (SA), brassinosteroids, polyamines, and strigolactones (Arnao and Hernández-Ruiz, 2018; Khan et al., 2022).
MEL and auxins
MEL shares functional and structural characteristics with IAA, the principal auxin, influencing three main areas: growth, rooting, and gravitropism. Research on lupin plants revealed that while high levels of MEL inhibit growth similar to auxins, lower concentrations promote it. For example, MEL enhances vegetative growth by regulating auxin receptor activity and auxin-responsive genes (Tan et al., 2019). MEL also significantly boosts root formation, including lateral and adventitious roots, by interacting with auxin pathways. In Arabidopsis, MEL treatment increased lateral roots up to threefold without affecting root hair density, and overexpressed MEL lines showed more lateral roots compared to wild types (Pelagio-Flores et al., 2012; Zuo et al., 2014).
One key mechanism involves the interaction of MEL with auxin-related genes, particularly those encoding YUCCA flavin monooxygenases (YUCs) and TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA) proteins, which are central to indole-3-acetic acid (IAA) biosynthesis (Wang et al., 2016). These authors demonstrated that MEL treatment in Arabidopsis led to a differential expression of these genes, resulting in altered endogenous IAA levels. Additionally, MEL influences auxin transport by downregulating auxin influx carriers such as AUX1/LAX, which are critical for root development and gravitropic responses (Swarup and Péret, 2012; Weeda et al., 2014). In rice, MEL was found to modulate the expression of various auxin-related TFs, including WRKY, NAC, MYB, and bHLH, contributing to the reconfiguration of root architecture (Liang et al., 2017).
MEL and GAs
MEL’s influence on GAs has been particularly evident in germination processes. In salt-stressed cucumber seedlings, MEL upregulated the expression of genes responsible for GA biosynthesis, including several GA oxidases such as GA 20-oxidase (GA20ox) and GA 30-oxidase (GA3ox), which catalyze the conversion of GA precursors into bioactive GAs by oxidizing C-20 and C-30 of the GA molecule, respectively, thus leading to enhanced germination and early seedling growth (Zhang et al., 2014). Similar effects were observed in Brassica napus, where MEL increased GA levels, leading to stronger seedling development under stress (Tan et al., 2019). Additionally, MEL was shown to regulate GA signaling components, including DELLA proteins, promoting growth and reproductive development (Shi et al., 2015a, 2015b, 2015c, 2015d).
MEL and CKs
MEL works synergistically with CKs to regulate plant growth and stress tolerance. MEL increases CK levels by upregulating genes involved in CK biosynthesis. In drought-stressed creeping bentgrass, the combined action of CKs and MEL improved photosynthetic efficiency, chlorophyll content, and relative water content, enhancing drought resilience (Ma et al., 2018). In sweet cherries, MEL and CKs were linked to improved fruit ripening, highlighting their combined role in plant development (Tijero et al., 2019). A recent study on two-week-old seedlings of A. thaliana exposed to high light conditions (600 μmol · m−2 · s−1 for 24 h) has demonstrated that MEL works synergistically with CKs by either activating or repressing genes involved in CK synthesis and signaling. Key genes in this process include IPT3, IPT5, and LOG7 (Lonely Guy 7), which encode isopentenyltransferases (IPT) and phosphoribohydrolase (PRH), respectively. Additionally, CKs influence the expression of genes related to MEL metabolism, specifically ASMT, SNAT, COMT, and CAND2 (Bychkov et al., 2023).
MEL and ABA
MEL generally antagonizes ABA, a hormone associated with stress and dormancy. It reduces ABA levels by downregulating key biosynthesis enzymes, such as 9-cis-epoxycarotenoid dioxygenase (NCED), while upregulating ABA catabolism genes like CYP707 that encodes ABA 8’-hydroxylase (Li et al., 2015). This dynamic lowers ABA content, thereby alleviating dormancy and promoting growth. For example, MEL decreased ABA levels in cucumbers, apples, and Chinese cabbage, facilitating seed germination and early development (Arnao and Hernández-Ruiz, 2021). However, MEL may increase ABA levels under certain conditions, as observed in barley and Elymus nutans, suggesting that the response varies by plant species and stress type (Li et al., 2016; Fu et al., 2017). Additionally, Zhang et al. (2017) found that MEL treatment in perennial ryegrass under heat-induced senescence conditions resulted in decreased ABA content, increased CK biosynthesis gene expression, and improved photosynthetic efficiency, delaying senescence.
MEL and ethylene
MEL’s interaction with ethylene is complex, influencing both ripening and senescence. It stimulates ethylene production in some cases, such as in tomato fruits, where MEL promotes ethylene biosynthesis and improves ripening by enhancing sugar and acid content (Sun et al., 2015). Conversely, MEL suppressed ethylene in pears, strawberries, and bananas, delaying ripening and extending postharvest shelf life (Zhai et al., 2018; Hu et al., 2017). The regulatory effect depends on factors like MEL concentration, fruit physiology, and environmental conditions.
Overall, MEL acts as a plant growth regulator by fine-tuning phytohormone interactions to optimize growth, development, and stress tolerance. Figure 11 illustrates a working model summarizing MEL’s role in key physiological processes of higher plants, including seed germination, seedling growth across developmental stages, root development, senescence, and fruit ripening. It also highlights its interactions with various phytohormones and environmental factors. This model provides insights into melatonin’s multifunctional role in plant physiology and its potential applications in agriculture.
MEL: Abiotic and Biotic Stresses
Since the beginning of human civilization, plants have played a fundamental role as crops, medicines, and sources of biomaterials. Crops serve as a primary food source for humans, and their successful development is crucial for meeting the demands of an anticipated population increase in the future (Zhu et al., 2022). However, environmental factors, including climate change and human activities, can reduce crop yields. These stressors are generally divided into two main categories: abiotic and biotic stresses (Lal et al., 2023; Moustafa-Farag et al., 2019). Abiotic stressors include salinity (Zhao et al., 2021b), drought (Tiwari et al., 2021), high light exposure (Roeber et al., 2021), extreme temperatures (Ahad et al., 2023; Zioutopoulou et al., 2021), and heavy metal exposure (Zulfiqar et al., 2019), among others. Conversely, biotic stressors involve pathogens such as bacteria (Greco et al., 2021; Peeters et al., 2013), fungi (Doehlemann et al., 2017), viruses (Tiwari et al., 2022), and nematodes (Montarry et al., 2021), herbivorous insects (Liu et al., 2023), among others.
Although each stress factor, or a combination of these, affects each plant species differently (Mittler, 2006; Suzuki et al., 2014; Mareri et al., 2022; Pardo-Hernández et al., 2024a, 2024b), among the main negative effects that they can cause in the plant are modulations at transcriptomic (Hirayama and Shinozaki, 2010; Kreps et al., 2002), metabolomic (Ramakrishna and Ravishankar, 2011), biochemical (Wahab et al., 2022) and physiological levels (Vezza et al., 2018; Yildiztekin et al., 2018), which usually result in a reduction of photosynthetic rate and plant growth, modification of stomatal movement, enhanced lipid peroxidation and increased levels of ROS and RNS, as well as modulations in phytohormones levels (Mittler, 2002; Gill and Tuteja, 2010; Gong et al., 2020; Gogoi et al., 2024; Rizhsky et al., 2002; Yamamoto et al., 2001; Yang et al., 2008). Nevertheless, there are numerous scientific studies in which attempts have been made to increase tolerance to these stresses from the perspective of gene modification (Kumar et al., 2020; Zhan et al., 2021), addition of chemical stimulants (Zulfiqar et al., 2022), phytohormones (Bittner et al., 2022; Wang and Komatsu, 2022) or with microorganisms (Enebe and Babalola, 2018; Kumar and Nautiyal, 2022). Among these methods, MEL, both endogenously and exogenously applied, has proved to be a novel means of alleviating these negative effects in plants such as oxidative membrane damage, electrolyte leakage, and inefficient photosynthesis (Aghdam et al., 2023; Altaf et al., 2021; Arnao & Hernández-Ruiz, 2021; Colombage et al., 2023; Karumannil et al., 2023; Moustafa-Farag et al., 2019; Sun et al., 2021).
A major factor that makes MEL such a promising molecule for dealing with plant stresses is its ability to scavenge the reactive species generally overproduced in plant stresses (Ahammed et al., 2024; Gu et al., 2022; Hernández-Ruiz et al., 2023; Jiang et al., 2023; Pardo-Hernández et al., 2020; Tan et al., 2000; Zhao et al., 2021a) although the mechanism of action is not totally clear yet (Ahmad et al., 2023; Hassan et al., 2022; Jahan et al., 2023). In turn, it has been demonstrated crosstalk between MEL and the rest of the phytohormones in plants that can enhance plant survival and manage stresses (Khan et al., 2022; Sati et al., 2024; Sharif et al., 2018; Sharma et al., 2020; Zeng et al., 2022). In addition, an increase in both enzymatic and nonenzymatic antioxidant systems has been observed when MEL is applied to plants, enhancing their tolerance to stress (Imran et al., 2021; Jan et al., 2023; Magri and Petriccione, 2022; Zhao & Hu, 2023; Wang et al., 2023).
MEL has been applied exogenously to plants at any development stage by different methods such as immersion (Fan et al., 2022; Sun et al., 2015), spraying (Erdal, 2019; Xia et al., 2021; Zahedi et al., 2020), irrigation (Park et al., 2021; Wang et al., 2013) or even injection (Mansouri et al., 2021), being previously demonstrated that the plants accumulate the exogenous MEL and it is safe for animals and humans (Tan et al., 2007). The application of exogenous MEL enhances the response of plants to several stresses (Table 3); for example, in tomato (Solanum lycopersicum L.), several beneficial effects have been reported (Altaf et al., 2022a). Among these, in managing salinity stress, MEL application by irrigation has led to an increase in the tolerance of salt stress in tomato seedlings, resulting in increased plant growth and upregulation of the ROS scavenging system, and also a decrease in chlorophyll degradation and MDA content (Siddiqui et al., 2019a; Zhou et al., 2016). Additionally, a pretreatment or a spray application of MEL in tomato seedlings has exhibited more tolerance to drought stress by enhancing the antioxidant enzymes again and decreasing the oxidative damage, as well as minimizing the reduction of photosynthesis efficiency and improving fruit quality and yield (Altaf et al., 2022b; Ibrahim et al., 2020). Also, the enhanced activity of key photosynthetic enzymes and the reduction of H2O2 and MDA by spraying exogenous MEL on tomato seedlings can alleviate the negative effects of acid rain and high-temperature stresses (Debnath et al., 2018; Jahan et al., 2021a). Apart from these, the infiltration with exogenous MEL of tomato fruits has shown an increased methyl jasmonate (MeJA) content and upregulation of JAs defense signaling, leading to an enhanced defense resistance to the pathogen fungus Botrytis cinerea (Liu et al., 2019).
Representative Examples of How the Exogenous Melatonin Treatment of Plants Manages Some Abiotic and Biotic Stresses
APX, ascorbate peroxidase; CAT, catalase; GA, gibberellin; GR, glutathione reductase; NOS, nitric oxide synthase-like activity; NR, nitrate reductase; PAL, phenylalanine ammonia-lyase; POD, peroxidase; RBOH, respiratory burst oxidase homologs; SOD, superoxide dismutase.
Not only is the exogenous application of MEL relevant to improving plant survival against stresses, but endogenous MEL also plays a fundamental role. Table 4 summarizes some examples of how endogenous MEL levels can affect the mechanism of response against some abiotic and biotic stresses. This fact has been demonstrated in multiple studies where some of the genes involved in MEL biosynthesis have been inhibited or overexpressed. In the model plant A. thaliana, knockdown mutant and SNAT overexpressing line have shown that MEL promotes UV-B stress resistance (Yao et al., 2021). Also in this plant species, ASMT- and SNAT-overexpressing lines, which have more endogenous MEL content, are more resistant to B. cinerea by acting synergistically with antioxidant systems and increasing JA levels (Zhu et al., 2021). Moreover, a double mutant snat1snat2 in Arabidopsis has revealed that the biosynthesis of MEL is associated with the auxin signaling pathway, being implicated in the tolerance to salt stress (Miao et al., 2024). Furthermore, in Solanaceae plants such as N. benthamiana seedlings that overexpressed the ASMT gene of grapevine enhanced the MEL content, improving its resistance under osmotic and salt stresses (Yu et al., 2022b), and in overexpressing COMT1 Solanum lycopersicum L. plants, a boost in the photosynthetic process and higher tolerance to elevated CO2 atmosphere and heat stress was observed (Hasan et al., 2023).
Representative Examples of How the Endogenous Melatonin of Plants Accomplishes Some Abiotic and Biotic Stresses
KO, knockout.
Not only is MEL protective of plants against stresses, but its derivatives, especially 2-hydroxymelatonin (2-OHM) treatments, also do so. It is noteworthy that exogenous application of 2-OHM has been shown to mitigate the negative effects of cadmium stress in cucumber seedlings by enhancing the photosynthetic rate and the activity of SOD, catalase, and APX, and also upregulating the transcription of ethylene-regulating genes (Shah et al., 2020). Moreover, in rice seedlings, drought and cold combined stress effects can be alleviated by the 2-OHM treatment due to elevated proline content, decreased levels of electrolyte leakage, better preservation of chloroplast and mitochondrial structures, and the upregulation of genes related to the tolerance of several abiotic stresses (Lee and Back, 2016). Figure 12 presents a graphical representation of the beneficial effects of exogenous MEL, administered through various methods, in counteracting nitro-oxidative stress induced by different environmental factors, both abiotic and biotic. Thus, the exogenous application of MEL has several beneficial effects, including increased activity of antioxidant enzymes, improved photosynthetic parameters, enhanced production of endogenous MEL, and reduced levels of ROS and lipid peroxidation. Together, these effects contribute to greater abiotic and biotic stress tolerance and improved plant growth.

Additionally, MEL was found to induce systemic resistance by activating pathogenesis-related (PR) genes and enhancing antioxidant enzyme activity, which provides protection against postharvest fungal pathogens in tomato fruits (Arnao and Hernández-Ruiz, 2017). MEL plays a crucial role in plant immunity by modulating the expression of key defense-related genes and interacting with signaling molecules such as SA and JA. It acts as a signaling molecule that triggers innate immune responses against various pathogens, including bacteria and fungi (Lee et al., 2014; Qian et al., 2015). One of the main mechanisms through which MEL regulates plant immunity is by inducing the expression of PR genes, which are typically activated by SA. This suggests that MEL functions upstream in the SA-mediated immune response. Studies in Arabidopsis have demonstrated that MEL application increases PR1, PR2, and PR5 gene expression, enhancing resistance to Pseudomonas syringae infection (Lee et al., 2015). Additionally, MEL upregulates mitogen-activated protein kinase (MAPK) cascades, which are essential for transducing immune signals and activating SA-dependent defense pathways (Lee and Back, 2016, 2017).
Beyond SA signaling, MEL also interacts with JA, another critical hormone in plant defense. While SA is typically associated with systemic acquired resistance against biotrophic pathogens, JA is involved in induced systemic resistance against necrotrophic pathogens and insect herbivores. MEL has been shown to coregulate the expression of JA biosynthesis genes, such as lipoxygenase (LOX), allene oxide synthase (AOS), and 12-oxo-phytodienoic acid reductase (OPR), leading to increased JA accumulation under pathogen attack (Zhu and Lee, 2015). This suggests that MEL fine-tunes the balance between SA- and JA-mediated defense pathways, depending on the nature of the stressor. Furthermore, MEL enhances NO production, which in turn modulates both SA and JA signaling pathways, contributing to a well-coordinated immune response (Shi et al., 2015a, 2015b, 2015c, 2015d).
At the gene regulation level, MEL-induced immunity involves the upregulation of TFs that control SA and JA responses. Studies have shown that MEL treatment increases the expression of WRKY and ERF TFs, which are key regulators of SA- and JA-dependent defense genes (Arnao and Hernández-Ruiz, 2015, 2017). The crosstalk between MEL, SA, and JA ensures a dynamic and flexible immune response, allowing plants to effectively combat various pathogens while minimizing growth trade-offs. Overall, MEL functions as a central regulator in plant immunity, orchestrating SA- and JA-mediated gene expression to enhance stress resilience and disease resistance.
Likewise, MEL plays a role in the interaction between beneficial microorganisms and plants. It enhances nodulation efficiency in rhizobia-legume symbiosis by regulating genes involved in root architecture and nitrogen fixation (Wang et al., 2024). Additionally, MEL mitigates oxidative stress in root nodules, ensuring a stable environment for nitrogen-fixing bacteria. On the other hand, in arbuscular mycorrhizal symbiosis, MEL promotes spore germination and mycelial growth, enhancing nutrient uptake (e.g., phosphorus and nitrogen) and improving resistance to various stresses (Wei et al., 2023).
Biotechnological Applications of MEL
Due to the significant and wide mode of action of MEL in plant physiology, it is clearly conceivable to think about the use of this phytohormone as a cornerstone for biotechnological purposes to improve most plant developmental stages, including germination, growth, and fruit ripening. This is of special interest to impact crop production but also to gain knowledge that could be exploited in the future.
Thus, seeds can be directly treated with MEL to benefit germination and favor the establishment of the epicotyl and radicle primordia once the seed coat bursts and the dormancy breaks down (Arnao and Hernández-Ruiz, 2017, 2019; Aghdam and Arnao, 2024; Mukherjee et al., 2024). Seed dormancy is an evolutionary adaptation that avoids germination when the environmental conditions are not suitable to ensure seedling survival (Fenner and Thompson, 2005; Black et al., 2006). During seed germination, MEL not only operates as a modulatory molecule in the newest tissues by virtue of its hormonal potential but also as a former antioxidant under circumstances where the oxygen concentration increases, as a consequence of the exposure of plant cells to an aerobic environment after disappearing the coat protection (Bailly, 2004; Pulido et al., 2009; Diaz-Vivancos et al., 2013; Gomes and Garcia, 2013; Soln et al., 2022). Furthermore, seed priming with MEL can enhance the germination rate, especially for seeds stored in unfavorable conditions (Cao et al., 2019; Yu et al., 2021; Kolupaev et al., 2024a).
The exogenous application of MEL is also a biotechnological tool to improve plant growth and development due to its dual role as an antioxidant and regulator of different responses in plant cells (Arnao and Hernández-Ruiz, 2017, 2019; Corpas et al., 2022; Aghdam and Arnao, 2024; Mukherjee et al., 2024). Additionally, the external MEL stimulates the proper internal biosynthesis, thus leading to a cascade of signals that could improve the plant’s health status. This may also contribute to fostering priming events, which can be useful when plants undergo harmful situations imposed by either biotic or abiotic actors (Ulhassan et al., 2019; Sun et al., 2021; Ali et al., 2022, 2023; Alfalahi et al., 2024; Ameen et al., 2024; Lucas et al., 2024; Sati et al., 2024). Accordingly, the use of MEL to counteract the effect of diverse stress agents and to improve the quality of edible products obtained from several horticultural products comprising leaves (spinach, pak choi, cabbage), florets (broccoli, cauliflower), and roots/tubercules (cassava, potato) has been recently reviewed by Mukherjee et al. (2024).
In the case of fruits, the treatment with MEL after postharvest has been widely used in diverse crops, independently of whether fruits are climacteric or nonclimacteric. Climacteric fruits are those whose ripening is associated with higher levels of respiration and of the gaseous phytohormone ethylene, whereas nonclimacteric fruits do not follow that pattern, and a multifaceted involvement of other phytohormones takes place (Palma et al., 2019; Farcuh et al., 2022; Corpas et al., 2023). As a consequence of the MEL application to edible fruits, different effects that improved their quality have been reported: They include delayed ripening (Sun et al., 2016; Yan et al., 2022, others), lowering damage and injury as a result of chilling practices (Aghdam et al., 2019; Liu et al., 2019; Darré et al., 2024; Wang et al., 2022; others), or potentiating the tolerance against infections triggered by pathogens (Sharafi et al., 2019; Li et al., 2022a, 2022b; others), and other species. Nevertheless, other reports have revealed opposite effects in the same fruit species (Shan et al., 2022), so this potential controversy needs to be further explored to get a clear role of MEL in fruit physiology. Perhaps the effects depend on the species, the cultivar, the MEL concentration, and the mode of application, but also the surrounding and environmental conditions. These issues are targets for future research.
Additionally, the preharvest treatment of plants with MEL has been widely followed to improve the quality of diverse fruits, including, among others, tomato (Li et al., 2023), pomegranate (Lorente-Mento et al., 2021), pear (Yan et al., 2023), kiwifruit (Peng et al., 2023), sweet cherry (Michailidis et al., 2021; Carrion-Antoli et al., 2021), apricot (Medina-Santamarina et al., 2021), Japanese plum (Cortés-Montaña et al., 2023), and eggplant (Song et al., 2023). This is somehow some sort of priming strategy more complex than postharvest and implies the application of MEL in whole plants in the field, where handling by farmers and the interactions with the neighboring conditions should be taken into account. In Figure 13, a simple scheme of the options to apply MEL to seeds and whole plants through either the aerial parts or roots and by pre- and postharvest practices is depicted.

The treatment with MEL can also be envisaged as an alternative biotechnological strategy to common culture uses involving pesticides and fertilizers, based on the vigor promoted to plants by this phytohormone. Thus, it has been reported that exogenous treatment with MEL prevents pests (Sun et al., 2021; Hernández-Ruiz et al., 2023), and the hormone has also been used instead of pesticides, as it mediates the response and signaling events under biotic stress conditions (Tiwari et al., 2022; Giraldo-Acosta et al., 2023). Additionally, the combination of MEL with pesticides has been proposed to reduce the amount of chemicals used in agriculture (Zhang et al., 2017).
Alternatively, the promotion of the effects exerted by MEL can be triggered by the use of Trp as a precursor of this phytohormone, as indicated in section II above. This strategy allows the activation of the own plant metabolism, which can be considered as a sort of “training/learning” process, thus facilitating the plant cells to execute their machinery not only under physiological conditions but also under unfavorable situations provoked by biotic and/or abiotic/environmental agents. The treatment with Trp has been widely developed as a tool to benefit plant growth and against plant stress in diverse crops (Park et al., 2009; Rao et al., 2012; El Sayed et al., 2014; Korkmaz et al., 2020; Mosa et al., 2021; Gondek and Mierzwa-Hersztek, 2021; Hussain et al., 2024).
From an anthropological point of view and due to the key role of MEL in our physiology, one might think about using plants as a source of such regulatory chemicals for human consumption to promote health and prevent certain disorders. Unlike other active principles (vitamin C/ascorbate; provitamin α/β-carotene; vitamin E/α-tocopherol) that are present in plant tissues at high levels, and we include them in our diet through fruit and vegetables, MEL is only contained in very little amounts in plant materials. Nevertheless, although the intake of MEL-containing plant products influences our MEL levels less than the light–dark cycle, it has been reported that the diet content of this hormone is significant in modulating daily MEL fluctuations. The effect is not only direct but it cannot be discarded as a stimulatory effect on the MEL biosynthesis once we have ingested the hormone from edible plants (Peuhkuri et al., 2012) (Fig. 14). The MEL content in edible plant products varies considerably (Peuhkuri et al., 2012; Meng et al., 2017; Wu et al., 2021), so a diet rich in a diversity of these foods can contribute to improve our health status and prevent a number of disorders (Peuhkuri et al., 2012; Gomes Domingos et al., 2019; Muñoz-Jurado and Escribano, 2024). For example, a study involving 1277 schoolchildren and university students found that those who consumed foods rich in MEL, such as strawberries, tomatoes, pomegranates, cherries, mangoes, and peanuts among others, experienced improved sleep quality and lower levels of depression (Borisenkov et al., 2023).

With this view, the industry devoted to the provision of MEL is called to have a variety of products depending on the target sectors, including pharmacology and biomedicine, academic research, and agrochemistry, as depicted in Figure 15. For example, MEL-loaded nanocarriers have indeed emerged as a significant advancement in drug delivery systems, enhancing MEL’s stability, bioavailability, and targeted delivery (Chuffa et al., 2021). Beyond medical applications, recent research indicates that these nanocarriers hold promise in the agro-industrial sector as well.
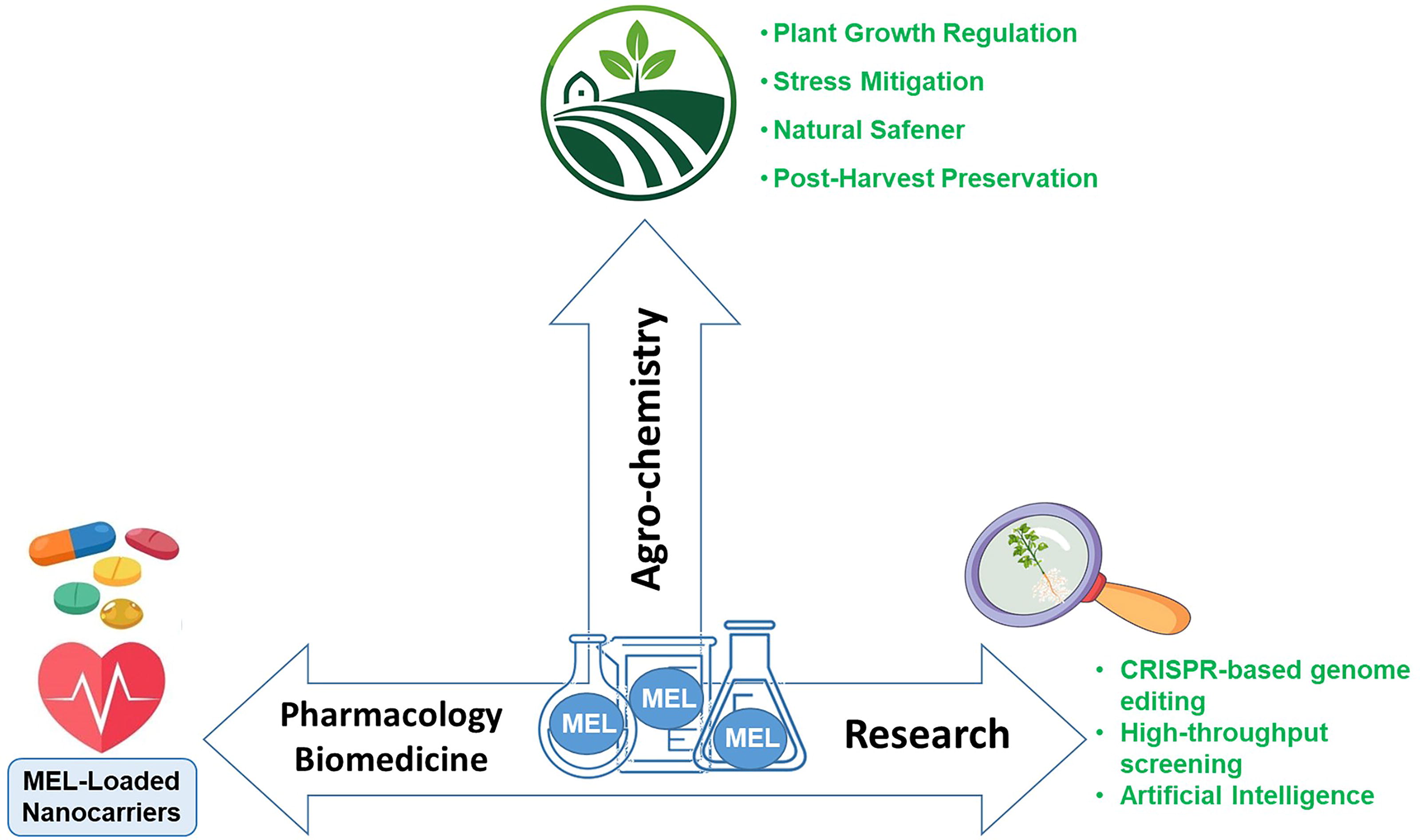
Conclusions and Future Perspective
Since the discovery in 1995 that MEL is produced endogenously in plant cells, significant advancements have been made regarding its role in plant metabolism and physiology. Initial studies on MEL highlighted its antioxidant properties, but subsequent research has uncovered its roles in regulating physiological functions, particularly under stress conditions. The MEL biosynthetic pathway exhibits remarkable complexity, including reversible steps and alternative routes that remain partially unresolved. Advances in molecular techniques, such as CRISPR and subcellular localization studies, could further elucidate these pathways. Highlighting key enzymes like serotonin N-acetyltransferase (SNAT) and their regulatory mechanisms can provide valuable insights into optimizing MEL production for agricultural applications. The identification of MEL receptors has revealed its signaling potential in plants. Additionally, MEL influences TFs and microRNAs, which regulate stress–response and developmental genes. Integrating high-throughput sequencing and mutant studies can deepen our understanding of these regulatory networks. In this context, MEL’s potential in agriculture includes enhancing crop resilience, improving postharvest quality, and mitigating stress impacts. Translating these findings into cost-effective solutions requires collaboration between researchers and industry stakeholders. Emphasizing case studies and pilot projects can demonstrate MEL’s value in real-world settings. Future directions in MEL-related research should focus on: Elucidating the molecular mechanisms behind the MEL biosynthetic pathway: Employ advanced tools like omics technologies and CRISPR to identify unknown enzymes and regulatory pathways in MEL biosynthesis. Understanding the mechanisms of MEL transporters and the transport of their biosynthetic intermediates across cellular compartments and between cells. Optimizing agricultural applications: Develop standardized protocols for MEL treatments, considering crop-specific responses, dose-dependent effects, and environmental conditions. Bridging research and practice: Conduct translational studies to validate laboratory findings in field conditions and assess the economic feasibility of MEL-based interventions. Expanding the research scope: Explore MEL’s role in nontraditional crops and its potential in addressing emerging agricultural challenges such as climate change. Combining recent methodological approaches, such as CRISPR-based genome editing, high-throughput screening, and artificial intelligence, could accelerate large-scale gene discovery and be useful for the precise functional validation of MEL’s regulatory mechanisms, as well as improve stress resilience and enhance crop productivity in higher plants.
An intriguing area of study is the integration of MEL with other signaling molecules, such as the gasotransmitters NO and hydrogen sulfide, as well as H2O2. These molecules are interconnected and can exhibit either synergistic or antagonistic effects, depending on the specific physiological process or in response to adverse environmental conditions (Kolupaev et al., 2024b; Corpas et al., 2024). Their interactions may occur at the transcriptomic level or through reversible posttranslational modifications, such as S-nitrosation or persulfidation. MEL’s multifaceted roles in plant physiology and stress tolerance position it as a promising tool for sustainable agriculture.
By integrating fundamental research with practical applications, MEL-based strategies can enhance crop productivity and resilience, contributing to global food security. Continued interdisciplinary collaboration will be key to unlocking its full potential.
Footnotes
Acknowledgments
The authors would like to thank reviewing editors: Alexandre Boscari, Natalia Bykova, Robert Hancock, Jingjing Huang, Alberto Iglesias, Yuriy E. Kolupaev, Chandan K. Sen, and Hideo Yamasaki.
Authors’ Contributions
Conceptualization: F.J.C. Writing—original draft: F.J.C., J.T., R.M.R., R.J.R., and J.M.P. Writing—review and editing: F.J.C., J.T., R.M.R., R.J.R., and J.M.P. All authors discussed and commented on the article, equally contributed to making the figures, provided critical feedback, and helped shape the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
F.J.C.’s and J.M.P.’s research is funded by grants from the European Regional Development Fund, cofinanced by the Ministry of Science and Innovation of Spain (PID2023-146153NB-C21 and CPP2021-008703). J.T. acknowledges funding through contract PREP2023-001833 from the Ministry of Science and Innovation, Spain, R.M.R.’s research is supported by the “Fundación Séneca—Agencia de Ciencia y Tecnología de la Región de Murcia” under grant reference 22527/PI/24, Spain.