
Abstract
The distribution of major phosphate deposits in the Precambrian sedimentary rock record is restricted to periods that witnessed global biogeochemical changes, but the cause of this distribution is unclear. The oldest known phosphogenic event occurred around 2.0 Ga and was followed, after more than 1.3 billion years, by an even larger phosphogenic event in the Neoproterozoic. Phosphorites (phosphate-rich sedimentary rocks that contain more than 15% P2O5) preserve a unique record of seawater chemistry, biological activity, and oceanographic changes. In an attempt to emphasize the potentially crucial significance of phosphorites in the evolution of Proterozoic biogeochemical cycles, this contribution provides a review of some important Paleoproterozoic phosphate deposits and of models proposed for their origin. A new model is then presented for the spatial and temporal modes of occurrence of phosphorites along with possible connections to global changes at both ends of the Proterozoic. Central to the new model is that periods of atmospheric oxygenation may have been caused by globally elevated rates of primary productivity stimulated by high fluxes of phosphorus delivery to seawater as a result of increased chemical weathering of continental crust over geological timescales. The striking similarities in biogeochemical evolution between the Paleo- and Neoproterozoic are discussed in light of the two oldest major phosphogenic events and their possible relation to the stepwise rise of atmospheric oxygen that ultimately resulted in significant leaps in biological evolution. Key Words: Precambrian—Primary productivity—Phosphorus—Biogeochemical evolution—Great Oxidation Event. Astrobiology 10, 165–181.
1. Introduction
M
2. Distribution and Characteristics of Paleoproterozoic Phosphorites
Worldwide occurrences of sedimentary phosphate deposits around 2.0 Ga suggest that phosphogenesis was related to global chemo-oceanographic changes and probably not solely to restricted local conditions. A compilation of Paleoproterozoic phosphate deposits is presented in Table 1, which also lists noteworthy characteristics, including age constraints, host sedimentary rock and associated minerals, type of phosphate occurrence (often related to microbial or diagenetic processes), and estimated tonnage of P2O5 when available. One of the oldest and largest Paleoproterozoic phosphate deposits occurs as stromatolitic phosphorites (with up to 35% wt of P2O5) in the ∼2.0 Ga Jhamarkotra Formation of the Lower Aravalli Group in Rajasthan, India (Banerjee, 1971; Chauhan, 1979). These columnar stromatolites are mainly composed of carbonate fluorapatite with intercolumnar dolomite (Fig. 1a) and form a band of ∼35 m in thickness within the Jhamarkotra dolomite. Like many other rock successions containing Paleoproterozoic phosphorites, the Lower Aravalli Group was deposited in an active rift basin unconformably above the Archean basement and constitutes a marine transgression (Roy and Paliwal, 1981).

Photographs of Paleoproterozoic phosphorites: (
Occurs in a transgressive sequence stratigraphically above a conglomerate unit unconformably sitting on Archean basement.
Fm., formation.
Phosphorites of the Kursk Magnetic Anomaly region in western Russia and Ukraine typically occur in carbonaceous sedimentary rocks from intracratonic rift basins and are often associated with extensive banded iron formations (Nikitina and Shchipanskiy, 1987). These Paleoproterozoic phosphorites often occur as thin beds and nodules, which may reflect a diagenetic origin. In the Susong Group in China, phosphorites have up to 26% P2O5 and occur in dolomites, cherts, and graphitic schists that are part of a transgressive sequence that stratigraphically overlies a conglomerate unit (Longkang and Zhendong, 1988). In the Chinpingshan area of China, Paleoproterozoic apatite-rich deposits are hosted in carbonates that are often manganiferous (Yudin, 1996). High manganese content in such rocks could be related to increased redox states of seawater. Farther east on the Sino-Korean platform, there is a possibly coeval and relatively large micaceous phosphorite with around 20% P2O5 in Youngyu, North Korea (Yudin, 1996). In the Aldan Shield of Siberia and Yakutia in northeastern Russia, Paleoproterozoic phosphorite deposits occur in highly metamorphosed skarn rocks and contain large P2O5 reserves, estimated to 130 million tons (Guliy, 1989; Yudin, 1996). Many of these deposits share similarities in depositional environments and mineralogical characteristics in that they preserve relatively high amounts of organic matter or sulfides, or both, which are likely related to biological processes during deposition and diagenesis.
In North America, several Paleoproterozoic phosphorites occur along the southern shore of the Superior Craton and share similar characteristics. Uraniferous phosphatic sediments occur in Minnesota and contain up to 25% P2O5 in the form of nodular apatites (Ullmer, 1981, 1985; McSwiggen et al., 1986). Uranium (U4+) likely substitutes for Ca2+ during the diagenetic formation of apatite and may have adsorbed onto organic matter in the depositional environment or during diagenesis. Although relatively restricted in extent, phosphorites of Minnesota occur in graphitic and pyritiferous shales or mudstones that stratigraphically overlie a basal conglomerate. Nearby in the Baraga Group in northern Michigan and its equivalent in western Ontario, phosphorites with up to 15% P2O5 occur in argillites and cherts as nodules (Fig. 1b), thin beds, and also as small rounded laminated structures that are possibly stromatolites (Mancuso et al., 1975; Cannon and Klasner, 1976; Hall, 1985). Farther east in Québec and Labrador, contemporary cherty carbonaceous and pyritiferous turbidites also host uraniferous phosphorite beds (Bell and Thorpe, 1986). These three coeval examples of phosphorites along the Paleoproterozoic southern margin of the Superior Craton suggest that regional oceanographic changes contributed to the deposition of phosphorites. In the late Paleoproterozoic Thelon Formation, in the Canadian Northwest Territories, uraniferous fluorapatite occurs occasionally with high levels of sulfate and aluminum and has growth habits that include microcrystalline cement, acicular crystals, and nodules in quartz arenites and arkoses (Miller, 1983; Gall and Donaldson, 2006). These occurrences in North America illustrate the diversity of depositional environments in which phosphorites formed in the Paleoproterozoic.
In Fennoscandia, Paleoproterozoic uraniferous phosphorite beds are found in metamorphosed sedimentary rocks and are of comparable ages to those from North America. Dolomites and apatite-rich gneisses with up to 24% P2O5 host thin phosphorite beds in various schist belts in western and central Finland (Rehtijärvi et al., 1979; Vaasjoki et al., 1980). In the Pechenga area of northwestern Russia, 1.90 to 2.06 Ga phosphorites in organic-rich sediments occur as concretions, beds, oolites, and stromatolites in carbonaceous gritstones, shales, and other sedimentary rocks (Bekasova and Dudkin, 1982; Yudin, 1996). The Pechanga Belt has been geochronologically correlated in the region with the Onega Basin where phosphate concretions occur in organic-rich and sulfidic shales (Melezhik et al., 1999, 2005). Like the Aravalli stromatolitic phosphorites of India, these occurrences in the organic-rich zone of the Ludikovi Group of Fennoscandia are noteworthy examples of the intimate relationship between phosphorites, microbial activity, and diagenesis. In central and northern Finland, fluorapatite bands that are a few centimeters thick occur in organic-rich Paleoproterozoic sulfide facies banded iron formations of the Kainuu Schist Belt (Fig. 1c, Gehör, 1994). Possibly coeval phosphorites in banded iron formations in Swedish Lapland contain up to 18% P2O5 (Parák, 1973). Finally, uraniferous phosphorite bands occur in carbonates and graphitic gneisses in the slightly older Jatuli Group (Laajoki and Saikkonen, 1977; Äikäs, 1980, 1981).
3. Conceptual Models of Phosphogenesis
The origin of phosphorites is still not well understood, and over the years there have been several hypotheses for their origin that include biological, chemical, and volcano-sedimentary influences (see reviews in Kazakov, 1937; Bushinskii, 1966; Cook and Shergold, 1990). Kazakov (1937) recognized a striking similarity between the various phosphorites of the Russian platform, where most occurrences are part of a transgressive sequence and lie stratigraphically above basal conglomerates followed by arenaceous or argillaceous sandstones (see also Table 1). Kazakov (1937) further proposed that the source of phosphorus in phosphorites was mineralized organic matter in deep marine environments and that nutrient-rich water masses ascended to continental shelves to form phosphate deposits.
Many phosphorite deposits in the ocean today occur in areas of upwelling where phosphate, regenerated from the recycling of organic matter in the deep ocean, is brought to coastal areas and triggers blooms of primary productivity such as along the coasts of Namibia (Bremner and Rogers, 1990), Baja California (Jahnke et al., 1983), and Peru-Chile (Veeh et al., 1973). A simplified illustration of the upwelling model is shown in Fig. 2. Cyanobacterial blooms in upwelling areas can produce large quantities of organic matter that concentrate phosphorus in biomass, deplete it from surface waters, and lead to decreased oxygen availability beneath the photic zone. During the decomposition of organic matter in the water column, phosphorus is liberated, and its concentration increases in the anoxic zone (Ruttenberg, 2003). Phosphorus is also regenerated during early diagenesis and can either return to the water column by upwelling or become concentrated in sediments. Depositional environments of phosphorites are varied, and this is reflected by the diversity of host lithologies where they are found (Fig. 2). Rock types hosting phosphate deposits include deep marine facies such as shales and banded iron formations, which are often carbonaceous, sulfidic, or both, and shallow-marine facies such as argillites, limestones, cherts, and sandstones.

Simplified illustration of the upwelling model for phosphogenesis. Sedimentary phosphate accumulations in Precambrian sedimentary rocks are shown in red as ovals (for concretions or nodules), curved lines (for thin beds), stromatolite columns, and small dots (for fine disseminations). Various lithotypes represented include volcanic rock (“v” pattern), granitoids (“+” pattern), iron formations (black), shales (lines), limestone (bricks), sandstone-chert (dotted pattern), and conglomerate (fragmented pattern). Major metabolic pathways in the water column or in sediments are shown in yellow. The water column is color coded to represent redox states as turquoise (oxic photic zone), blue (redox transition zone), and green (anoxic zone). BIFs, banded iron formations; REE, rare Earth elements.
Non-upwelling areas such as estuaries and semi-restricted embayments can also be host to phosphate deposits or authigenic carbonate fluorapatite (Bremner and Rogers, 1990). For instance, diagenetic phosphate minerals have been observed in sediments of the Long Island Sound and Mississippi Delta in the United States (Ruttenberg and Berner, 1993), and phosphorites occur along the eastern and southern coast of Australia (O'Brien and Veeh, 1980). Most organic matter burial in marine sediments occurs in non-upwelling areas where deltaic-shelf-estuarine sediments are deposited. In these environments, pore water solutions in sediments often become enriched in dissolved phosphate due to its release during organic decomposition by anaerobic heterotrophic microbial communities (Berner, 1990).
During early diagenesis, phosphorus can be released from organic matter but also from iron oxides under anoxic conditions, both of which may play a role in phosphogenesis (Ingall and Jahnke, 1994; van Cappellen and Ingall, 1994). Oceanic islands and seamounts can also host sedimentary phosphorites because these sites expose sediments to oxygen minimum zones (see Glenn et al., 1994, and references therein). At the transition between anoxic and oxygenated waters, phosphogenesis can be promoted by microbial communities involved in the anaerobic recycling of organic matter produced by primary producers in the photic zone. Bioturbation and aerobic degradation in immediate subsurface sediments enhance the removal of phosphorus by sediments, whereas anoxic conditions promote the return of dissolved phosphorus to the water column (van Cappellen and Ingall, 1996; Colman and Holland, 2000; Brasier and Callow, 2007). The redox state of sediments during early diagenesis thus strongly influences phosphate return fluxes to the water column. Therefore, the accumulation and preservation of phosphate-rich sediments can be promoted by increased atmospheric oxygen levels that lead to a more oxygenated water column.
Another possible link in the model for phosphogenesis is major glaciations, during which post-glacial oceanic overturn after thawing could drive a rise in sea level and upwelling of phosphorus-rich bottom waters (Arthur and Jenkyns, 1981; Cook and Shergold, 1984; Donnelly et al., 1990; Glenn et al., 1994). As noted in Table 1, many Paleoproterozoic phosphorites occur during marine transgressions, and rising sea levels generally follow major glaciations. Transgressions may favor the deposition of phosphate deposits by creating new restricted basin configurations and increased nutrient input from weathering or upwelling. Massive input of fresh water loaded with nutrients, cations, and anions from chemically weathered continental rocks should result in a stratified water column and favor oceanic overturn. It has also been suggested that the lack of bioturbation in the Precambrian may have permitted shallow water phosphatization and that the introduction of bioturbation in the Cambrian would have contributed to deepening of the anoxic alkaline conditions necessary for phosphate precipitation (Brasier and Callow, 2007).
The formation of sedimentary phosphorites ultimately depends on a significant enrichment of phosphorus from seawater by microbial activity and the recycling of this organic matter in sediments (Fig. 2). Microbial influence on the formation of phosphorites is conspicuous, and different microorganisms probably have a range of involvement from heterotrophy and phosphorus regeneration to active apatite biomineralization. Microbial community composition of modern phosphorites is not well characterized, but there are a few potentially key lineages of bacteria known from these sedimentary environments. The proteobacteria Thiomargarita resides in sediments from upwelling waters off the coast of Namibia, and its abundance correlates with concentrations of phosphate in pore water solutions, which has led to the suggestion that these bacteria cause phosphogenesis (Schulz and Schulz, 2005). Interestingly, Thiomargarita is the largest known bacterium, with cells up to 750 μm in diameter; and, like some of its large-celled close relatives Beggiatoa and Thioploca, it is a chemolithotroph that performs denitrification and sulfide oxidation (Schultz et al., 1999). Modern mats of Beggiatoa from the Santa Barbara Basin in California are also enriched in phosphorus by up to 6 times the level found in planktonic organic matter (Reimers et al., 1990).
Heterotrophic microorganisms that recycle organic matter in sediments use a variety of anaerobic metabolic pathways such as sulfate reduction, denitrification, iron and manganese reduction, methanogenesis, and fermentation. Microbial sulfate reduction is an important metabolic pathway involved in the decomposition of organic matter and can lead to relatively high levels of sulfide in organic-rich sediments thereby resulting in the diagenetic formation of sulfide minerals. A product of microbial sulfate reduction is also bicarbonate (
The detailed mechanism behind the precipitation of authigenic carbonate fluorapatite from seawater or during diagenesis is not well understood, but studies of carbon, oxygen and sulfur isotopes of carbonate, and sulfate in carbonate fluorapatite have suggested that diagenetic processes, especially microbial sulfate reduction and fermentation, play important roles (Kolodny and Kaplan, 1970; McArthur et al., 1980, 1986; Benmore et al., 1983). The precipitation of carbonate fluorapatite ultimately occurs during early or late diagenesis from the decay of organic matter. Precipitates of carbonate fluorapatite can form granules, bands, hardgrounds, concretions, or nodules in muds and other marine sediments but also occur as stromatolites, oolites, peloids, foraminifera, and various fossilized organisms (Glenn et al., 1994). Microbial structures and fossils in sedimentary phosphorites are widespread and occur as filaments, cells, fossilized animals, and organic remains (Chauhan, 1979; Reimers et al., 1990; Xiao et al., 1998, 2007; Steiner et al., 2004).
4.1. Setting the Stage for Paleoproterozoic Phosphogenesis: Rifting, Glaciations, and Oxygenation
A surge in tectonic activity appears to have occurred during the assembly of large continental landmasses in the Neoarchean (possibly one or more supercontinents) and when these began to break up in the earliest Paleoproterozoic (Aspler and Chiarenzelli, 1998; Bleeker, 2003; Zhao et al., 2004; Barley et al., 2005). It has been suggested that two supercontinents formed in the Neoarchean: one included cratons in Zimbabwe, West and South Africa, Western Australia, Brazil, and India; while the other supercontinent, called Kenorland, included cratons in North America, Fennoscandia, and Siberia (Aspler and Chiarenzelli, 1998). The initial protracted breakup of Kenorland may have been related to a mantle plume around 2.5–2.4 Ga (Fig. 3), while the final separation and dispersion of this large continental landmass was also a plume-related event but was completed around 2.1–2.0 Ga (Heaman, 1997; Barley et al., 2005; Bekker et al., 2006; Halls et al., 2008). Rifting of these ancient large pieces of continental crust would have led to the creation of rift zones and new sedimentary basins in the Paleoproterozoic. Increased rifting activity is significant because marine extensional basins are sites where glacially derived diamictites are deposited and subsequently preserved, whereas regional uplift caused by plate collisions is generally not correlated to glacial periods (Eyles, 2008). An increase in chemical weathering rates during this period is expected to have perturbed the carbon-silicate cycle as more Ca and Mg became available in seawater for carbonate precipitation and thereby led to a decrease in atmospheric CO2 levels (Berner, 1993; Goddéris et al., 2007).
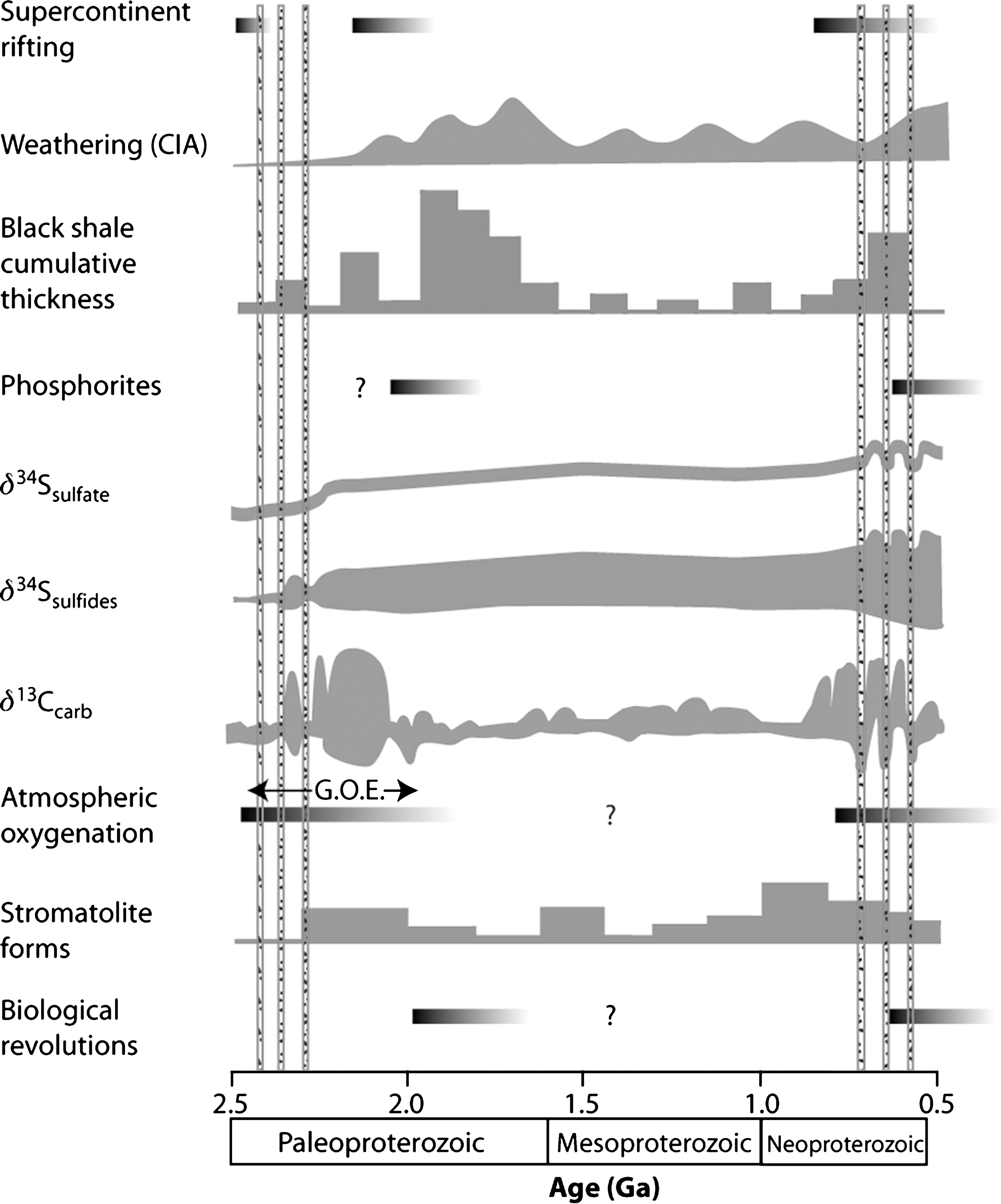
Proterozoic timeline of a selection of major biogeochemical changes including supercontinent rifting, weathering from chemical index of alteration and black shale cumulative thickness (after Condie et al., 2001), phosphorites, evolution of sulfur (after Canfield, 1998; Hurtgen et al., 2005; Halverson and Hurtgen, 2007) and carbon isotopes (after Shields and Veizer, 2002; Halverson et al., 2005; Bekker et al., 2006), and stromatolite forms (after Semikhatov and Raaben, 1996). Vertical bars represent the estimated timing of major glacial diamictites. See text for details. G.O.E., Great Oxidation Event.
Melting of a Snowball Earth event would likely have required intense greenhouse conditions, which would have led to widespread deposition of carbonates and a global transgression. Passive CO2 emissions from volcanic activity over timescales of tens of millions of years would have built up atmospheric greenhouse conditions during the glaciation, especially since the drawdown of CO2 by chemical weathering of Ca and Mg silicates would have been negligible during this time. The consequent enhanced chemical weathering may have been partly responsible for the negative carbon isotope excursions seen in “cap carbonates” through an increase of alkalinity into coastal seawater. In addition to these changes, a decrease in seawater Ni concentrations near the Archean-Proterozoic boundary may have stifled biological CH4 production and may have been a contributing factor to the subsequent glaciations and atmospheric oxygenation (Konhauser et al., 2009). Therefore, the Archean-Proterozoic boundary was a time when lowering greenhouse gases may have initiated a colder climate with more intense temperature fluctuations than previously in the Archean. Many Paleoproterozoic glacial deposits formed soon after the breakup and along the rifted continental margins of Kenorland.
Paleoproterozoic volcano-sedimentary sequences exhibit evidence of several major glaciations, with at least one proposed Snowball Earth event, during which the ice sheet extended to near the equator (Ojakangas, 1988; Evans et al., 1997; Bekker et al., 2001). The onset of the Paleoproterozoic glacial period is best represented and preserved in the Huronian Supergroup in Ontario, Canada. Similarities in lithologies and stratigraphy between the Huronian and Snowy Pass (Wyoming) supergroups suggest that these volcano-sedimentary successions were deposited in an epicratonic basin that formed from rifting between the Superior and Wyoming cratons of Kenorland (Roscoe and Card, 1993). The Huronian and Snowy Pass supergroups deposited between 2.45 and 2.22 Ga and preserve three discrete glacial diamictites (Fig. 3). The Bruce and Vagner formations are the second tillites of the Huronian and Snowy Pass supergroups, respectively, and are stratigraphically correlated (Young, 1973; Roscoe and Card, 1993). Both of these glacial horizons are stratigraphically overlain by a cap carbonate with similar carbon isotopic compositions as their younger Neoproterozoic counterparts (Bekker et al., 2005). Notably, after this second glaciation around 2.35 Ga, sulfides in sedimentary rocks do not contain significant mass-independently fractionated sulfur isotopes and have larger ranges of δ 34S values when compared to older rocks (Fig. 3), which suggests that atmospheric oxygenation occurred during interglacial and post-glacial periods in the Paleoproterozoic (Hattori et al., 1983; Papineau et al., 2005, 2007).
Other glaciations of this period are found in a similar geodynamic context. For instance, some Huronian glaciations have been stratigraphically correlated with several other now-distant glacial diamictites in North America such as in the Marquette (Michigan), Mistassini (Québec), and Hurwitz (Nunavut) supergroups (Ojakangas, 1988), and possibly also with Paleoproterozoic glacial diamictites in Fennoscandia (Marmo and Ojakangas, 1984; Strand and Laajoki, 1993; Ojakangas et al., 2001). Glacial diamictites are also preserved in other Paleoproterozoic sequences in South Africa and Western Australia, and these appear to have been related to a mantle plume that resulted in the emplacement of voluminous banded iron formations (Barley et al., 1997, 2005). These large banded iron formations are overlain by marine sedimentary sequences that include tillites in the Transvaal Supergroup in South Africa (Bekker et al., 2001, 2005) and Hamersley Supergroup in Western Australia (Martin, 1999; Barley et al., 2005) and may thus have also formed in response to a major tectonic event in the earliest Paleoproterozoic.
4.2. Chemical Weathering in the Paleoproterozoic
The consequences of high tectonic activity combined with glaciations in the Paleoproterozoic likely caused increased chemical weathering rates. Over geological timescales (i.e., tens of millions to a few hundreds of millions of years), increased delivery to seawater of sediments eroded from continental crust naturally brings more phosphate since rivers provide the most important source of phosphorus to the oceans (Föllmi, 1995, 1996). If periods of increased chemical weathering can be caused by elevated tectonic activity or major glaciations, or both, then greenhouse climate conditions can increase the supply of phosphorus to the oceans and provoke phosphogenesis.
Marine chemical sediments that preserve increases in 87Sr/86Sr may record periods of higher weathering rates, because this ratio increases with the chemical weathering of ancient continental rocks. Conversely, the 87Sr/86Sr ratio of marine chemical sediments decreases when Sr is input from hydrothermal circulation or the weathering of juvenile magmatic rocks. Major climatic and tectonic changes over geological timescales can thus modulate the global 87Sr/86Sr of seawater by influencing riverine fluxes of Sr or its isotopic composition. This process is the basis for suggesting that enhanced chemical weathering after major glaciations is responsible for short-term variations in Sr isotopes, while tectonic activity drives the long-term trends (Blum and Erel, 1995). The Sr isotope record of Paleoproterozoic carbonates shows an increase in 87Sr/86Sr from Archean levels (Veizer and Compston, 1976), which is consistent with higher weathering rates during the Paleoproterozoic Great Oxidation Event. In an attempt to exclude possible Sr contribution from detrital clay minerals, a reevaluation of this global shift based on the lowest 87Sr/86Sr for Paleoproterozoic carbonates shows a similar trend (Shields and Veizer, 2002; Shields, 2007), although there are some exceptions with relatively low 87Sr/86Sr during the Paleoproterozoic δ 13Ccarb excursion in a few localities (Bekker et al., 2003b). While the time resolution of Paleoproterozoic Sr isotope data published so far is insufficient to trace weathering rates in detail; for instance, in connection with glaciations, the rise in 87Sr/86Sr from 0.7022 to 0.7046 between 2.5 and 1.9 Ga is consistent with increased weathering rates.
Evidence for intense chemical weathering after the Paleoproterozoic glaciations also comes from a substantial increase in the chemical index of alteration (or CIA; see Fig. 3) in shales deposited after the glaciations (Nesbitt and Young, 1982; Condie et al., 2001) and the widespread distribution of mature quartz sandstones in almost all Paleoproterozoic post-glacial successions (Bekker et al., 2006). Oxidative weathering also leads to higher levels of cations (e.g., Ca2+, Mg2+, Na+, K+) and anions (e.g., PO4 3−, SO4 2−, Cl−) in seawater. The widespread deposition of sulfate evaporites after the Paleoproterozoic glacial period (Cameron, 1983; El Tabakh et al., 1999; Bekker et al., 2006) is also consistent with high chemical weathering rates under intense greenhouse conditions and higher atmospheric redox states.
4.3. High Productivity in the Paleoproterozoic
Newly formed intracratonic rift basins are ideal sites for high levels of primary productivity because of their proximity to nutrient sources from weathering. For instance, modern meromictic lakes, estuaries, and fjords are often associated with rift zones and host high levels of primary productivity fueled by high nutrient availability. Transgression and basin subsidence during rifting can create ideal conditions for phosphorite deposition by combining upwelled and weathered sources of phosphorus into partly restricted basin configurations. Active Paleoproterozoic rift basins often were sites where diverse and abundant stromatolites developed. δ 13Ccarb values up to +12‰ have been reported from shallow-marine carbonates deposited in such geodynamic context (Karhu, 1993; Karhu and Holland, 1996; Wanke and Melezhik, 2005). Enhanced weathering rates over geological timescales during Paleoproterozoic interglacial periods should have led to an elevated supply of nutrients in coastal seawater that would have resulted in high rates of primary productivity. In turn, these blooms would have produced significant quantities of atmospheric oxygen (Fig. 2) and thus may have played a key role in modulating atmospheric levels of CO2 and CH4 that possibly triggered subsequent glaciations (Papineau et al., 2005, 2007). Ultimately, the cyclic nature of glacial-interglacial cycles at both ends of the Proterozoic may have ended after the establishment of a new carbon-oxygen balance between oxygenic photosynthetic and aerobic heterotrophic organisms once oxygen levels reached a certain level or sufficient evolutionary time had passed for the emergence of new oxygen-breathing heterotrophic organisms, or both occurred concomitantly (Papineau et al., 2005). This was at the time of the final breakup of Kenorland around 2.1 Ga, when unprecedented events in the Paleoproterozoic carbon cycle occurred. Shortly after that, the oxygenation of the atmosphere ceased with the end of the carbonate carbon isotope excursion.
Changes in the Paleoproterozoic carbon cycle were profound between ∼2.25 and 2.06 Ga (Fig. 3), when worldwide carbonates exhibited significant δ 13C excursions that have been related to a major increase of atmospheric oxygen (Karhu and Holland, 1996; Melezhik et al., 1999; Bekker et al., 2003a). This connection stems from carbon isotope mass balance considerations that dictate that 13C-enriched carbonates can form when a large amount of organic matter is prevented from being re-oxidized (e.g., through increased burial rates), which thereby leaves a net excess of oxygen in the atmosphere. Possible connections between δ 13Ccarb excursions and high rates of primary productivity have been suggested before (Kump and Arthur, 1999; Melezhik et al., 1999; Aharon, 2005). Under high nutrient availability, intense biological activity can act to concentrate phosphorus and lead to local eutrophication conditions. Primary productivity in costal areas is expected to have remained high during post-glacial periods, especially with a continuously elevated flux of nutrients from weathering over geological timescales. Therefore, anoxic bottom waters in these environments may have created a positive feedback of diagenetically regenerated phosphorus to the water column and further enhanced deepwater anoxia (van Cappellen and Ingall, 1996; Colman and Holland, 2000; Aharon, 2005). Such a mechanism may also have contributed to produce elevated δ 13Ccarb values in the Paleoproterozoic.
Models have shown that increased chemical weathering fluxes of phosphorus to the oceans after major glaciations can stimulate photosynthetic oxygen production (Lenton and Watson, 2004), which may have been an underlying cause for the occurrence of δ 13Ccarb excursions in interglacial and post-glacial Paleoproterozoic carbonates. Notably, carbonates formed during this time period also preserve an exceptional increase in taxonomic diversity and abundance of stromatolite reefs (Melezhik et al., 1997, 1999; Grotzinger and Knoll, 1999; Semikhatov et al., 1999), which is consistent with high rates of primary productivity. The occurrence of phosphorites near the end of the worldwide Paleoproterozoic δ 13C excursion in carbonates (Melezhik et al., 1999; Bekker et al., 2003a) further suggests an important connection to high productivity and atmospheric oxygenation because phosphate deposits are absent from the Archean and Mesoproterozoic rock records (Fig. 3). Many Paleoproterozoic phosphorites formed in marine sediments stratigraphically above basal conglomerates (Table 1), which suggests that phosphogenesis occurs in dynamic depositional environments. Notably, Paleoproterozoic phosphorites formed significantly after the series of major glaciations, which tends to blur this possible link. However, it remains likely that intense chemical weathering was the ultimate source of phosphorus in these marine sedimentary deposits. Future geochronological constraints on these rock types will help to determine the duration of Paleoproterozoic phosphogenesis.
The relationship between phosphorites and primary productivity can potentially be traced through a multidisciplinary integration of the depositional environment, stratigraphic context, stable isotope data, and geobiology. For instance, the Jhamarkotra stromatolitic phosphorites (Fig. 1a; Table 1) occur in restricted environments and are part of a transgressive sequence. Regionally correlated dolomites without phosphorites from coeval open marine environments have δ 13Ccarb values up to +11‰ (Sreenivas et al., 2001; Maheshwari et al., 2002), which suggest that they were deposited contemporaneously with the worldwide Paleoproterozoic δ 13Ccarb excursions (Karhu and Holland, 1996) and may be related to a period of high primary productivity. Some shales stratigraphically above these 13C-enriched Jhamarkotra carbonates have relatively high δ 13Corg values that have been interpreted to indicate increased levels of primary productivity (Papineau et al., 2009). In addition, relatively high δ 13Corgvalues in the coeval nearby stromatolitic phosphorites have also been interpreted to indicate high primary productivity (Banerjee et al., 1986; Sreenivas et al., 2001). Therefore, along the rifted margin of the Paleoproterozoic Aravalli Supergroup, high levels of primary productivity and especially oxygenic photosynthesis were stimulated by high phosphorus availability. Phosphorites from the Ludikovi Group of Fennoscandia (Table 1) also preserve evidence of microbial activity during diagenesis and also occur in a transgressive suite. The Ludikovi phosphorites occur stratigraphically above stromatolitic dolomites with δ 13Ccarb excursions (Melezhik et al., 2007), which further suggest connections with high levels of primary productivity.
5.1. Setting the Stage for Neoproterozoic Phosphogenesis: Rifting and Glaciations
Phosphorites formed only sporadically during the Neoproterozoic and Phanerozoic (Cook and McElhinny, 1979; Cook and Shergold, 1984). Their occurrences coincided with major global biogeochemical changes as well as significant leaps in biological evolution. Similar correlations were described above for biogeochemical evolution in the Paleoproterozoic, but those for the Neoproterozoic-Cambrian periods are better preserved, more abundant, and even more striking. This perspective leads to the proposal of a comprehensive repeating model for the evolution of biogeochemical cycles and emergence of increasingly complex life at both ends of the Proterozoic.
Setting the stage for global biogeochemical changes in the Neoproterozoic was the onset of the breakup of the supercontinent Rodinia. Paleomagnetic evidence suggests that Rodinia had formed by ∼1.0 Ga and began to break up around 0.75 Ga (Powell et al., 1993; Weil et al., 1998). Rifting of Rodinia appears to have initiated a series of events at that time and culminated in the Neoproterozoic Snowball Earth glaciations (Goddéris et al., 2007). Global cooling in the Neoproterozoic, possibly triggered by a drawdown of CO2 from increased weathering during rifting, was recorded by thick, glacially influenced marine strata in newly created rift basins worldwide (Eyles, 2008). Between ∼0.750 and ∼0.580 Ga, several major glaciations or Snowball Earth events, or both, occurred in sequence and were preserved as glacially derived diamictites deposited on newly created passive rifted margins (Hambrey and Harland, 1985; Kirschvink, 1992; Powell et al., 1993; Young, 1995; Hoffman et al., 1998; Eyles, 2008). Examples of such Neoproterozoic glacial diamictites in rift basins are now found in western North America (Young, 1995), China (Wang and Li, 2003), Australia (Young and Gostin, 1991), South America (Pecoits et al., 2007), Oman (Allen, 2007), and many other areas. The combined effect of these tectonic and climatic perturbations must have included intense chemical weathering of continental crust.
5.2. Enhanced Chemical Weathering in the Neoproterozoic
Increased chemical weathering rates during this period of global rifting and glaciations were likely to have perturbed the carbon-silicate cycle and led to a decrease in atmospheric CO2 levels (Berner, 1993). Widespread weathering of continental crust in the Neoproterozoic is evidenced in part by an unprecedented global increase in 87Sr/86Sr (Veizer et al., 1983; Donnelly et al., 1990; Kaufman et al., 1993; Kennedy et al., 1998; Brasier et al., 2000; Shields and Veizer, 2002; Shields, 2007), high chemical index of alteration of shales (Fig. 3; Condie et al., 2001), and the widespread deposition of detrital clay minerals, an event that has been referred to as the “clay mineral factory” (Kennedy et al., 2006).
In addition to these observations, the Neoproterozoic sulfur cycle was strongly perturbed by high rates of oxidative weathering. Because rivers are the dominant sources of sulfate to the ocean, seawater sulfate concentration increases during periods of high chemical weathering rates and increased redox states. Sedimentary sulfides and sulfates in Neoproterozoic interglacial and post-glacial marine sedimentary rocks have an increased range of δ 34S values (Fig. 3), including 34S-enriched compositions (Gorjan et al., 2000; Hurtgen et al., 2005; Fike et al., 2006; Halverson and Hurtgen, 2007; Fike and Grotzinger, 2008), which is consistent in part with microbial sulfate reduction in increased sulfate concentrations (Habicht et al., 2002). The expected consequence of high weathering rates in the Neoproterozoic is higher fluxes of phosphate delivery by rivers to seawater that should have stimulated photosynthetic oxygen production (Lenton and Watson, 2004). The latter model also predicts that oxygen production will decrease if the normalized overall weathering rate is decreased.
5.3. Productivity and Phosphorites at the Neoproterozoic-Cambrian Boundary
Global glaciations and Snowball Earth events in the Neoproterozoic have been correlated on the basis of geochronology and carbon isotopic composition of carbonates (Fig. 3; e.g., Brasier et al., 2000; Halverson et al., 2005). In fact, it is unambiguous that worldwide Neoproterozoic interglacial and post-glacial carbonates have 13C enrichments, and this has been attributed to increases in the relative rate of organic carbon burial (e.g., Des Marais et al., 1992; Halverson et al., 2005). For instance, Neoproterozoic interglacial and post-glacial carbonates enriched in 13C, which occur almost everywhere in sequences of this age, are preserved (Kaufman et al., 1997; Shields and Veizer, 2002; Halverson et al., 2005; and references therein). High nutrient supply at the Neoproterozoic-Cambrian boundary has been correlated with episodes of carbon isotope excursions that were contemporaneous with hydrocarbon burial and evaporite deposition (Brasier, 1991). Evidence for high rates of primary productivity between, and after, the Neoproterozoic glaciations is preserved as various geochemical and geological signatures. Organic biomarkers have also been used to infer high rates of primary productivity after a Neoproterozoic Snowball Earth event and were proposed to be due to high nutrient availability from weathering (Elie et al., 2007). As shown in Fig. 3, the period between ∼0.800 and ∼0.600 Ga was also a time of worldwide black shale deposition (Condie et al., 2001) and unprecedented stromatolite proliferation and diversification (Grotzinger and Knoll, 1999). Notably, most Neoproterozoic-Cambrian phosphorites occurred during post-glacial periods during these major perturbations in the carbon cycle in China, northern India, South Australia, Namibia, Oman, and elsewhere.
There are numerous large phosphorite deposits in the late Neoproterozoic-Cambrian, which was a time interval when oceanic circulation experienced dramatic changes. Stagnation of the oceans during Snowball Earth events was followed by periods of vigorous circulation, which could have been conducive to phosphorite formation (Donnelly et al., 1990). Similar to the Paleoproterozoic, the combined effect of rifting of the Rodinia supercontinent and postglacial transgressions under intense greenhouse conditions in the Neoproterozoic would have resulted in high chemical weathering rates. Major Neoproterozoic-Cambrian phosphorite deposits occur in North and South America, Africa, Europe, Asia, and Australia (Notholt and Sheldon, 1986), and most of them stratigraphically overlie glacially derived sedimentary rocks (Cook and Shergold, 1984). Major phosphorites occur stratigraphically above glacial tillites in the Australian Adelaide Geosyncline (Howard, 1986), in the Chinese Doushantuo Formation (Yueyan, 1986), in the West African Volta and Taoudeni basins (Slansky, 1986; Flicoteaux and Trompette, 1998), in the Brazilian Bambuí Group and correlative sequences (Dardenne et al., 1986; Misi et al., 2006), in the Khubsugul Basin and related deposits of the Mongolian-Siberian region (Ilyin et al., 1986; Yanshin, 1986), and many other places in Vietnam, Kazakhstan, Pakistan, India, Europe, and North America (Cook and Shergold, 1984). Many of these phosphorites formed during major transgressions (Brasier, 1980) and along newly rifted continental margins.
Various types of sedimentological structures in Neoproterozoic-Cambrian phosphorites indicate biological involvement in their formation. Microorganisms were important in the formation of these phosphorites as evidenced in part by several examples of stromatolitic phosphorites (with columnar carbonate fluorapatite) of Cambrian age in Australia (Southgate, 1980; Schmidt and Southgate, 1982), Algeria (Bertrand-Sarfati et al., 1997), China (Yueyan, 1986), Russia (Yanshin, 1986), and elsewhere. Perhaps even more striking is the fact that many phosphorites of Late Neoproterozoic-Cambrian age preserve exceptional remains of multicellular organisms, including animal embryos (Xiao et al., 1998, 2007; Steiner et al., 2004), brachiopods (Cowan et al., 2005), and various other fossils of eukaryotic organisms (Cook and Shergold, 1984; Brasier, 1990, 2009). Collectively, these observations for the Neoproterozoic-Cambrian suggest strong connections between rifting and the ensuing greenhouse collapse, the consequent increase in chemical weathering rates and higher delivery of phosphate to seawater, the subsequent response of cyanobacterial blooms responsible for the accumulation of atmospheric oxygen, and, ultimately, the rise of complex multicellular oxygen-breathing organisms (Fig. 3).
6. Discussion of the Causes of Atmospheric Oxygenation and Significant Leaps in Biological Evolution
Phosphorite occurrences during periods of biogeochemical changes are likely to be related to higher fluxes of phosphorus to seawater from elevated chemical weathering rates over geological timescales. Although the specific mechanisms of apatite precipitation in sediments are not fully understood (Bentor, 1980; Notholt, 1980), the formation of phosphorites is related to a series of events that sequentially concentrate phosphorus such as high primary productivity, diagenetic processes in organic-rich sediments, and possibly direct microbial trapping or precipitation, or both (Fig. 2). In modern environments, phosphorites form in upwelling and non-upwelling areas, even though the oceanic concentration of phosphorus is less than 3 mM in both the Pacific and Atlantic oceans (Colman et al., 2005). Archean levels of oceanic phosphorus may have been lower than in the Paleoproterozoic, which could be a reflection of higher sequestration of phosphorus by adsorption onto iron oxides, such as in hydrothermal environments (Berner, 1972; Wheat et al., 1996), or a smaller flux of riverine phosphorus (Bjerrum and Canfield, 2002; cf. Konhauser et al., 2007), or both. In the aftermath of the Paleoproterozoic glaciations, however, enhanced greenhouse conditions led to widespread chemical weathering of continental crust and, therefore, to higher fluxes of phosphorus to the oceans. Because photosynthetic organisms quickly incorporated phosphorus, it is likely that the continuous riverine supply of phosphorus stimulated cyanobacterial blooms along coastlines and phosphogenesis occurred where basin configuration, paleoceanographic processes, and diagenetic conditions were adequate.
The model of increased post-glacial riverine phosphorus delivery for atmospheric oxygenation differs from other models proposed to explain the rise of atmospheric oxygen in the Paleoproterozoic. One such model suggests that the oxygenation of the atmosphere could have been caused by the delivery of hydrogen peroxide to the oceans in the aftermath of global glaciations (Liang et al., 2006). While this interesting model fits many geological observations, it is not easily compatible with positive δ 13C excursions in post-glacial carbonates nor with the occurrence of phosphorites and other lines of evidence that suggest high rates of primary productivity at both ends of the Proterozoic.
In another model, it has been suggested that the redox state of volcanic gases increased in the Archean and led to the Paleoproterozoic Great Oxidation Event (Kasting et al., 1993; Kump et al., 2001; Holland, 2002). A gradual increase in the oxidation state of volcanic gases and hydrothermal inputs could have occurred from the progressive loss of H2 from the Archean atmosphere (Kasting et al., 1993; Holland, 2002). Subducted oxidized crust may have penetrated deeply in the mantle and could have accumulated at the core-mantle boundary until a plume delivered the oxidized slab remnants to the surface (Kump et al., 2001). However, abundances of redox-sensitive elements such as chromium, vanadium, and iron in Archean volcanic rocks and minerals are similar to those of modern volcanic rocks and therefore do not support a change in the oxidation state of the mantle at ∼2.5 Ga (Canil, 1997, 2002; Delano, 2001). A corollary to this model is the possibility that there was a shift from predominantly submarine volcanism in the Archean to a more substantial role for subaerial volcanism in the Paleoproterozoic, which would have decreased overall sinks for atmospheric oxygen (Kump and Barley, 2007).
In another model, hydrodynamic escape of H2 to interplanetary space was proposed to have been important throughout the Archean and could have gradually left residual O2 in the atmosphere (Catling et al., 2001). Other simulations of hydrodynamic escape for H2 in a reducing atmosphere suggest that rates of hydrogen loss on early Earth were probably minimal (Tian et al., 2005). While these models of secular changes in volcanic outgassing and hydrodynamic escape have been proposed to explain Paleoproterozoic atmospheric oxygenation, they cannot explain the oxygenation of the Neoproterozoic atmosphere, which occurred under strikingly similar circumstances, and therefore do not support a stepwise oxygenation of the Proterozoic atmosphere.
An important similarity concerning the effects of atmospheric oxygenation is that both eras were followed by significant leaps in biological evolution. In the Paleoproterozoic, the accumulation of oxygen in the environment created new opportunities for large aerobic organisms (Fig. 3). Occurrences of millimeter- to centimeter-sized fossils in the ca. 1.9 Ga Marquette Supergroup in Michigan (Han and Runnegar, 1992), in the ca. 1.7 Ga Changcheng Group in northern China (Shixing and Huineng, 1995), in the ca. 1.6 Ga Vindhyan Supergroup in central India (Kumar, 1995), in the ca. 1.9 Ga Belcher Group in northern Canada (Hofmann, 1976), and in the ca. 1.9 Ga Pechenga Belt in northwest Russia (Akhmedov et al., 2000) constitute morphological evidence that multicellular oxygen-respiring eukaryotic organisms had evolved by the end of the Paleoproterozoic. It is reasonable to propose that the increase of atmospheric oxygen in the Paleoproterozoic atmosphere was beneficial to oxygen-respiring organisms, and therefore a new biogeochemical balance in the carbon cycle may have been established by the end of the global δ 13Ccarb excursion between oxygenic photosynthesis and aerobic respiration.
In comparison, life in the Late Neoproterozoic-Cambrian also diversified rapidly with the rise of metazoans, a development that was arguably the result of increased availability of oxygen. Shortly after the end of the Neoproterozoic glaciations, exceptional fossils of early animal evolution are preserved in marine sediments, and these include new macroscopic organisms shaped as spheres, disks, fronds, worms, and other radial and bilateral body architectures (e.g., Runnegar, 1991; Grotzinger et al., 1995; Jensen et al., 1998; Narbonne, 1998, 2005; Canfield et al., 2007). The Neoproterozoic rise in atmospheric oxygen was also responsible for the most significant leap in biological diversity and complexity in Earth history: the Cambrian explosion (Fig. 3). New organisms that emerged in the oceans of the Early Cambrian include sponges, brachiopods, protoconodonts, and trilobites, and many of these metazoans had biomineralized skeletons, complex shapes, and undulated/articulated bodies (Brasier, 2009).
Predictions from the model presented here will hopefully stimulate the search for phosphate accumulations in Paleoproterozoic sedimentary rocks (e.g., by using hydrochloric acid–ammonium molybdate solution in the field) and create opportunities for future quantitative modeling of feedback effects. The model proposed here is a simple natural series of cause-and-effect events consistent with global geological, geochronological, geochemical, and geobiological observations pertaining to the Paleo- and Neoproterozoic. The series of events that led to the oxygenation of the atmosphere and to fundamental changes in the evolution of global biogeochemical cycles was similar in both eras. Because of the striking similarities in global biogeochemical changes during both the Paleo- and Neoproterozoic, the causes of atmospheric oxygenation were likely similar and dependent upon an increased supply of nutrients to the oceans. During these eras, high chemical weathering rates must have led to higher riverine delivery of phosphorus to seawater, which would have increased rates of primary productivity that produced significant quantities of atmospheric oxygen. As a result, the accumulation of atmospheric oxygen paved the way for significant leaps in biological evolution in the Paleoproterozoic with the rise of macroscopic oxygen-breathing organisms and in the Neoproterozoic-Cambrian with the emergence of animals. The model of increased riverine phosphorus delivery leads to the conclusion that phosphorus was a key ingredient for the stepwise rise of atmospheric oxygen and for the evolution of life on Earth.
Footnotes
Acknowledgments
I would like to thank M.L. Fogel, J.A. Karhu, M. van Kranendonk, S. Hardy, and R.M. Hazen for constructive suggestions on the manuscript. Constructive criticism by G.A. Shields, L.R. Kump, and an anonymous reviewer are also gratefully acknowledged and helped to improve the manuscript. Support from the Geophysical Laboratory of the Carnegie Institution of Washington, Carnegie of Canada, and from the Fonds québécois pour la recherche sur la nature et les technologies (FQRNT) is gratefully acknowledged. This research was supported by the NASA Exobiology and Evolutionary Biology Program, and the NASA Astrobiology Institute through Cooperative Agreement NNA04CC09A.
Author Disclosure Statement
The author declares that no competing financial interests exist.