
Abstract
Cyanobacteria are capable of tolerating environmental extremes. To survive in extreme environments, cyanobacteria have developed the capability to adapt to a variety of stresses. For example, cyanobacteria have adopted a number of strategies with which to survive UV stress, including expression of UV-screening pigments and antioxidant systems. We have previously shown that several antioxidants are significantly expressed in Nostoc sp. by UVB irradiation. We report here that the content of UV-responsive biomarkers such as β-carotene and scytonemin can be easily detected by Fourier transform Raman spectroscopy with use of a small sample size and that the content of β-carotene is dependant on the UVB intensity and exposure time. Our results indicate that Raman spectroscopy may be a helpful tool to analyze UV-protective molecules of cyanobacterium in astrobiological studies without access to large sample sizes and complicated extractions, which are needed by other analytical techniques such as high-performance liquid chromatography and mass spectrometry. Key Words: Nostoc sp.—UVB—Raman spectra—β-carotene. Astrobiology 10, 783–788.
1. Introduction
S
To survive UV stress, cyanobacteria have developed a number of strategies, including synthesis of UV-screening pigment (e.g., scytonemins), migration to escape from UV radiation, expression of highly efficient antioxidant systems, and DNA repair systems (Quesada and Vincent, 1997; Sinha et al., 1998; Billi et al., 2000 Estevez et al., 2001; Dillon and Castenholz, 2003; Gorton and Vogelmann, 2003; Xue et al., 2005; Liu et al., 2008). For example, to tackle the detrimental effect of UVB radiation mediated via reactive oxygen species (ROS) (He and Häder, 2002; Shi et al., 2005), cyanobacteria have developed efficient ROS scavenging systems, including small-molecule antioxidants and antioxidant enzymes (Malanga et al., 1999; Tyagi et al., 2003; Wang et al., 2007). We have previously shown that antioxidant molecules, such as ascorbate and reduced glutathione, as well as the activities of the antioxidant enzymes such as superoxide dismutase, catalase, and peroxidase, are significantly expressed in Nostoc sp. in response to UVB irradiation (Wang et al., 2008).
Raman microspectroscopy has been increasingly used in biochemical and biological studies because it is nondestructive and requires only a small sample size (Edwards et al., 2004, 2005; Marshall et al., 2007; Bowden et al., 2010). It is feasible to acquire a range of valuable biochemical information from a single cell or just a few cells with Raman microspectroscopy (Choo-Smith et al., 2001; Heraud et al., 2007). The technique has been applied to a wide range of microbiological and astrobiological materials and has shown great potential for the spatial analysis of intact soil communities (Edwards et al., 2005). However, there are few reports for direct application of Raman microspectroscopy related to UV radiation on cyanobacteria. To investigate how and when the antioxidant molecules are expressed by UVB radiation in cyanobacteria, we analyzed the variation of antioxidant small molecules in a desert cyanobacterium Nostoc sp. with the simultaneous use of Raman microspectroscopy. Our results demonstrate that Raman microspectroscopy is a valuable tool for the analysis of the adaptation mechanism of cyanobacterium to UVB radiation. Further, Raman microspectroscopy has the potential for remote in situ measurement of biological processes in astrobiological materials, given that this non-intrusive technique requires only a small sample size and minimal sample manipulation to analyze organic and inorganic compounds simultaneously.
2. Experimental Approach
2.1. Cyanobacterial culture
Nostoc sp. was obtained from the FACHB collection (Freshwater Algae Culture Collection of Institute of Hydrobiology, The Chinese Academy of Sciences), which was originally isolated from Shapotou (37°27′N, 104°57′E) of Tengger Desert, Ningxia, China (Hu et al., 2003), and cultured in BG-110 (nitrogen-free BG-11; Rippka et al., 1979) media at 25 ± 1°C and 40 μmol photon m−2 s−1 light with bubbling air.
2.2. Ultraviolet B treatment
Ultraviolet B radiation was obtained from an Ultraviolet B TL 40W/12 tube (Philips, Netherlands), with its main output at 312 nm and a cellulose acetate filter to screen out UVC, in addition to regular fluorescent lamps (He and Häder, 2002). The UVB radiation (plus or minus) was measured with a double-monochromator spectroradiometer (OL 754, Optronic Laboratories, Orlando, FL, USA). Cyanobacterial samples in Petri dishes with quartz lids were irradiated with UVB for those times indicated and used for determination of photosynthetic activity. The samples were then transferred to 1.5 mL microfuge tubes and frozen in liquid nitrogen for Raman spectroscopy.
2.3. Determination of photosynthetic activity
The photosynthetic activity was assessed by measuring chlorophyll fluorescence, which was determined by a portable plant efficiency analyzer (Hanstech, UK). The determination was carried out at room temperature; the time of dark adaptation was at least 15 min. The stimulating light intensity was half the maximum intensity (ca. 1500 μmol photon m−2 s−1), and the value of Fv/Fm (Fv, variable fluorescence; Fm, maximum fluorescence) was used to indicate the photosynthetic efficiency.
2.4. Raman spectroscopy
Fourier transform Raman microspectroscopy of the Nostoc sp. samples was performed on a Nicolet Nexus Fourier transform Raman module with use of an Nd/YAG laser operating at 1064 nm and the methods of Edwards et al. (2003). The Raman spectra were recorded between 100 and 3800 cm−1, and 6000 scans were performed to obtain good spectra at 4 cm−1 resolution with wavenumbers accurate to 1 ± cm−1 or better. The laser power was typically maintained at 0.1–0.5 W to minimize sample degradation.
2.5. Statistics
All experiments were performed at least 3 times. Data were analyzed by one-way analysis of variance.
3. Results
3.1. Effect of UVB on the photosynthetic activity of Nostoc sp.
Shown in Fig. 1, the photosynthetic activity (Fv/Fm, a ratio of variable to maximum fluorescence indicating the quantum yield of photosynthesis) of the cyanobacteria without UVB treatment remained almost the same throughout the experimental period (6 h), while that of the cyanobacteria treated with 0.2 mW cm−2 UVB was decreased sharply and, at 6 h, reduced to only about 25% of the original ratio. When the cyanobacteria were treated with 0.1 mW cm−2 UVB, the Fv/Fm ratio was also decreased during the 6 h, but relatively more slowly than that of the 0.2 mW cm−2 UVB-treated cyanobacteria, and reduced to about 70% of the original activity at 6 h. The 0.05 mW cm−2 UVB treatment had an only limited effect on the cyanobacterial Fv/Fm ratio, which was only slightly decreased in the first 2 h and then stabilized. These data clearly show that the photosynthetic activity of Nostoc sp. was inhibited by UVB and the inhibition level was related to UVB intensity and exposure time.

Inhibition of photosynthetic activity of Nostoc sp. by UVB. The cyanobacterial culture was irradiated with UVB at different intensities for the times indicated, and the fluorescence of chlorophyll was recorded. The ratio of Fv/Fm was used to indicate the photosynthetic efficiency. Data are shown as mean ± standard deviation of three replicates.
3.2. Simultaneous detection of biomolecules in Nostoc sp. by Raman spectroscopy
Shown in Fig. 2, the spectral bands of β-carotene are at approximately 1002, 1160, and 1520 cm−1, which are the characteristic bands for, δ(C = CH), ν(C–C) and ν(C = C) of β-carotene, respectively (Edwards et al., 2005; Villar and Edwards, 2006; Heraud et al., 2007). The bands for scytonemin are at approximately 1630 cm−1 for ν(C = C) conjugated trans (Edwards et al., 2003; Villar and Edwards, 2006).
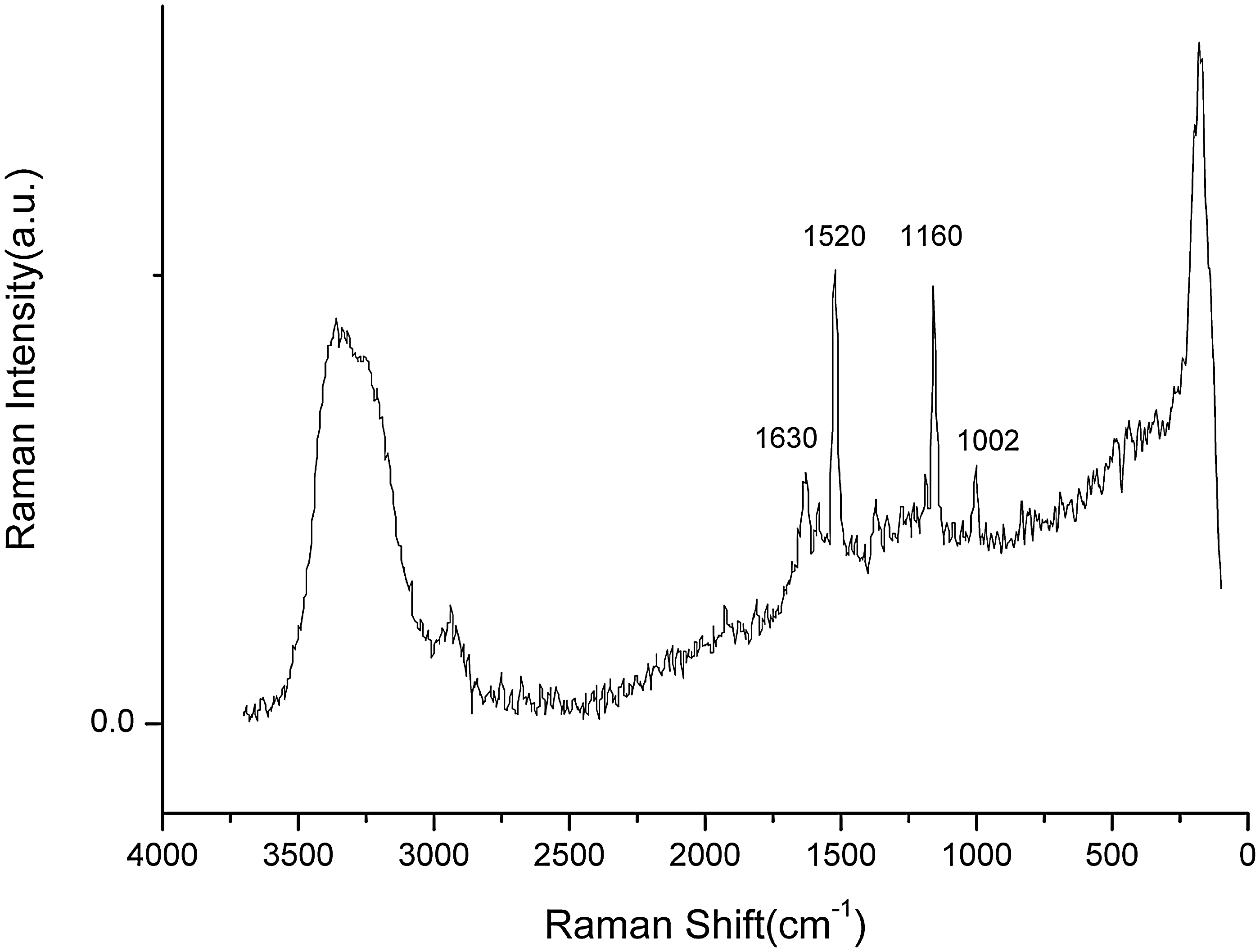
Raman spectra of UVB-treated Nostoc sp. The cyanobacterial culture was treated with 0.2 mW cm−2 UVB for 6 h, and Raman spectra were recorded between 100 and 3800 cm−1 at a spectral resolution of 4 cm−1. a.u., arbitrary units.
3.3. Effect of UVB intensity on the Raman spectra of Nostoc sp.
It was found by Raman spectroscopy that after UVB radiation the bands that correspond to β-carotene in Nostoc sp. are increased significantly when UVB intensity increases (Fig. 3), which indicates that β-carotene was expressed by UVB radiation. As is also shown in Fig. 3, the band of Raman spectra for scytonemin are increased after 0.05 and 0.1 mW cm−2 UVB radiation.
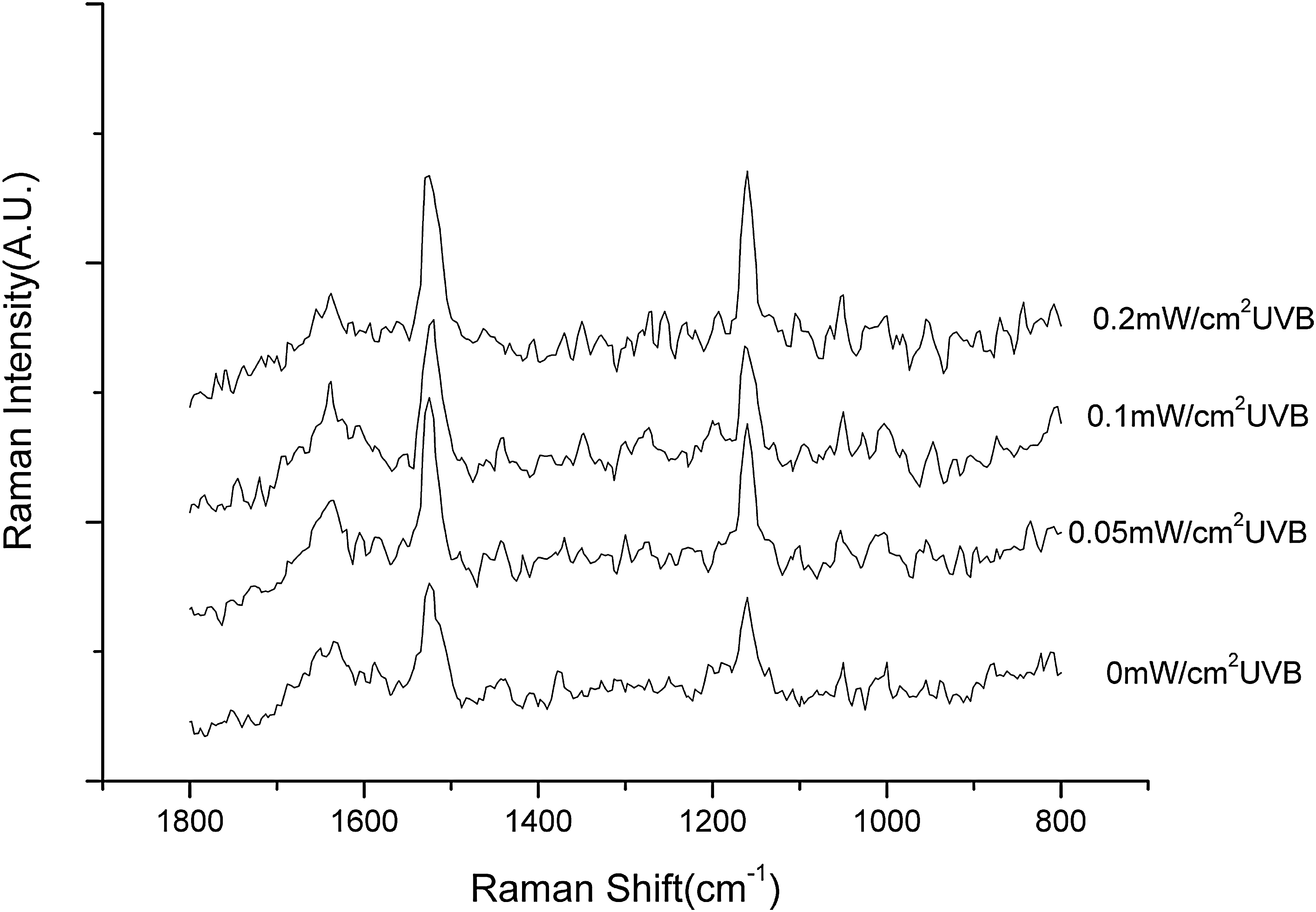
Effect of UVB intensity on Raman spectra of Nostoc sp. The cyanobacterial culture was treated for 6 h with UVB at different intensities as indicated, and Raman spectra were recorded at a spectral resolution of 4 cm−1. a.u., arbitrary units.
3.4. Effect of the exposure time to UVB on the Raman spectra of Nostoc sp.
As shown in Fig. 4, the bands at approximately 1160 and 1520 cm−1, which correspond to β-carotene, are increased at 2, 4, and 6 h then decreased at 12 and 24 h. On the other hand, the band for scytonemin at 1630 cm−1 is also slightly increased at 2 and 4 h (Fig. 4).

Effect of UVB exposure time on Raman spectra of Nostoc sp. The cyanobacterial culture was treated with 0.2 mW cm−2 UVB for different times as indicated, and Raman spectra were recorded at a spectral resolution of 4 cm−1. a.u., arbitrary units.
4. Discussion
Ultraviolet radiation is one of the factors that may harm Earth organisms in space exploration (Chen et al., 2003; Cockell et al., 2005). For instance, UVB may affect the metabolism of all organisms, particularly cyanobacteria and other photoautotrophs, which require solar radiation for normal life (He et al., 2002; He and Häder, 2002; Wang et al., 2006). Our data clearly indicate that the photosynthetic activity of Nostoc sp. was inhibited by UVB and the inhibition level was dependent on UVB intensity and exposure time. This shows that UVB is a harmful factor that affects the metabolism of cyanobacterium. To survive UV stress, cyanobacteria have adopted a number of strategies, including synthesis of UV-screening pigment, induction of a high-efficiency antioxidant system, and a DNA repair system (Quesada and Vincent 1997; Sinha et al., 1998; Billi et al., 2000; Xue et al., 2005). In addition to these strategies, the synthesis of small molecules (e.g., carotenoids, ascorbate acid) is also a very important aspect with regard to photoautotrophy. As effective quenchers of triplet-state photosensitizers, singlet oxygen, and peroxy radicals, carotenes have been proven to be effective against UV damage in organisms that suffer photooxidative stress (Albrecht et al., 2001; Lakatos et al., 2001; Laurion et al., 2002). For example, it has been reported that carotenes can provide significant protection against UVB in the cyanobacterium Synechococcus PCC7942 (Gotz et al., 1999; Schafer et al., 2006) and in Dunaliella spp. (White and Jahnke, 2002). Indeed, we found by Raman spectroscopy that after UVB radiation the bands that correspond to β-carotene in Nostoc sp. are increased significantly when UVB intensity is raised, which shows that expression of β-carotene may be an important adaptation mechanism of cyanobacteria to UVB radiation.
To investigate the effect of exposure time on the change of the content of the above molecules, Nostoc sp. culture was exposed to UVB for different times, and the Raman spectra were recorded at each time point. It was found that β-carotene content was increased at 2, 4, and 6 h time points. Since β-carotene is an effective quencher for ROS, its synthesis was significantly increased as the response to UVB-induced ROS production.
As an effective UV-screening pigment, scytonemin is found exclusively in cyanobacteria and has been shown to be expressed in Calothrix sp. and Chroococcidiopsis sp. by UV radiation (Dillon et al., 2002). In our experiment, we also found that scytonemin was synthesized by UVB radiation. However, we also found that the content of scytonemin was just slightly higher than that of the control, which may be due to the fact that synthesis of scytonemin takes a long time; it has been shown that it takes Chroococcidiopsis sp. more than 2 days and Nostoc punctiforme more than 5 days to synthesize scytonemin under UV radiation (Dillon et al., 2002; Fleming and Castenholz, 2007).
In summary, we found that the expression of UV-responsive biomarkers such as β-carotene could be easily detected in small sample sizes by Fourier transform Raman spectroscopy, which, in our study, revealed that the content of β-carotene was dependant on the UVB intensity and exposure time. These data indicate that, unlike analytical techniques such as high-performance liquid chromatography and mass spectrometry, which require large sample sizes and complicated extractions, Raman spectroscopy may be a particularly useful tool with which to study in situ desert cyanobacteria in an astrobiological context.
Footnotes
Acknowledgments
The work was supported by the Project of Chinese Manned Spaceflight, Natural Science Foundation of China (Nos. 30400093 and 30970688) and the National Major Programs of Water Body Pollution Control and Remediation (2008ZX07105-005).
Abbreviations
Fm, maximum fluorescence; Fv, variable fluorescence; ROS, reactive oxygen species.