
Abstract
Based on their unique resistance to various space parameters, Bacillus endospores are one of the model systems used for astrobiological studies. In this study, spores of B. subtilis were used to study the effects of galactic cosmic radiation (GCR) on spore survival and induced mutagenesis. In interplanetary space, outside Earth's protective magnetic field, spore-containing rocks would be exposed to bombardment by high-energy charged particle radiation from galactic sources and from the Sun, which consists of photons (X-rays, γ rays), protons, electrons, and heavy, high-energy charged (HZE) particles. B. subtilis spores were irradiated with X-rays and accelerated heavy ions (helium, carbon, silicon and iron) in the linear energy transfer (LET) range of 2–200 keV/μm. Spore survival and the rate of the induced mutations to rifampicin resistance (RifR) depended on the LET of the applied species of ions and radiation, whereas the exposure to high-energy charged particles, for example, iron ions, led to a low level of spore survival and increased frequency of mutation to RifR compared to low-energy charged particles and X-rays. Twenty-one RifR mutant spores were isolated from X-ray and heavy ion–irradiated samples. Nucleotide sequencing located the RifR mutations in the rpoB gene encoding the β-subunit of RNA polymerase. Most mutations were primarily found in Cluster I and were predicted to result in amino acid changes at residues Q469L, A478V, and H482P/Y. Four previously undescribed alleles in B. subtilis rpoB were isolated: L467P, R484P, and A488P in Cluster I and H507R in the spacer between Clusters I and II. The spectrum of RifR mutations arising from spores exposed to components of GCR is distinctly different from those of spores exposed to simulated space vacuum and martian conditions. Key Words: Bacillus—Spores—High-energy charged particle bombardment—Lithopanspermia. Astrobiology 10, 509–521.
1. Introduction
T
In interplanetary space, the radiation field is composed mainly of two groups: solar cosmic radiation (SCR) and galactic cosmic radiation (GCR) (Benton and Benton, 2001; Reitz, 2008; Ferrari and Szuszkiewicz 2009). In the vicinity of Earth, a third radiation component is present: radiation trapped by Earth's magnetosphere, the so-called van Allen belts (Reitz, 2008). For space life sciences, it is important to note that the radiation in space is a mixed radiation, composed of particles of different masses, charges, and energies (Hellweg and Baumstark-Khan, 2007). Charged particles, mainly electrons and protons (hydrogen nuclei), are steadily ejected from the upper atmosphere of the Sun. They are called the solar wind. Their energy is relatively low, about 1 keV (kilo electron volt). The solar wind creates the heliosphere, which surrounds our solar system. It produces a magnetic field that partially protects our solar system and its planets from galactic cosmic rays. Depending on the 11-year cycle of solar activity, the magnetic field is stronger during solar maximum and weaker during solar minimum (Rouillard and Lockwood, 2007). In addition to the quiet solar wind, there are sudden eruptions of high-energy particles. These events are composed primarily of protons with a minor component (5–10%) that is helium nuclei (alpha particles) and an even smaller part (1%) heavy ions and electrons. They are called solar particle events (SPEs), and they originate from magnetically disturbed regions of the Sun, which sporadically emit bursts of charged particles with high energies. These particles are ejected sporadically, last for hours to days, and can reach very high energies, in the worst cases up to several 10,000 MeV. SPEs are a sporadic component of radiation in space. They are more frequent during phases of solar maximum than during phases of solar minimum. SPEs are deflected by the magnetic field of Earth, so they do not reach astronauts in low Earth orbit (LEO), that is, at an altitude of about 400 km (Benton and Benton, 2001; Facius and Reitz, 2006; Reitz, 2008).
Radiation also reaches our solar system from outside the Solar System, from our galaxy, and even beyond. This GCR originates in cataclysmic astronomical events, such as supernova explosions. GCR consists of 98% baryons and 2% electrons (Hellweg and Baumstark-Khan, 2007; Reitz, 2008). The baryonic component is composed of 85% protons, with the remainder being alpha particles (14%) and heavier nuclei (about 1%). The latter component comprises HZE particles (particles of
Unlike Earth, Mars is devoid of an intrinsic magnetic field and, thus, most particles of GCR and SCR reach the outer atmosphere of Mars (Pissarenko, 1994). In addition, the carbon dioxide atmosphere of Mars of a pressure of about 600 Pa (6 mbar) is orders of magnitude less dense than that of Earth (100,000 Pa). This thin atmosphere provides considerably less shielding (about 16 g/cm2) than Earth's atmosphere (1000 g/cm2). As a consequence, the martian surface environment is exposed to the chronic radiation component (GCR and solar wind) as well as the sporadic component (SPE), both of which reach the martian surface nearly unfiltered (Pissarenko, 1994). A prominent feature of the surface radiation environment is the large number of neutrons produced as secondaries (interactions of the GCR and SCR with the regolith at the surface of Mars). Such neutrons can reach high frequencies and may even exceed those produced in the martian atmosphere (Badhwar and O'Neill, 1994). This radiation environment has to be taken into consideration when sending humans to Mars and looking for life (Horneck et al., 2006).
Radiation interacts with matter primarily through ionization and excitation of electrons in atoms and molecules. Biological effects are induced either through direct energy absorption by key biomolecules, such as proteins and nucleic acids, or indirectly via interactions of those molecules with radiation-induced radicals, which are, for example, produced by radiolysis of cellular water. With increasing density of ionizations, the number and magnitude of local damage in cells increases. This is especially valid for HZE particles of GCR that produce clusters of ions and radicals along their passage through a cell (Ballarini et al., 2008). Because HZE particles are considered to be important components among the cosmic rays that cause high levels of radiobiological damage (Ballarini et al., 2008), studies on biological effects of HZE particles of GCR in space as well as accelerated heavy ions at ground-based facilities are useful for estimating biological influences of space radiation environments. Due to their morphological and physiological attributes, bacterial endospores have been frequently used in HZE particle research. Endospores of Bacillus subtilis are highly resistant to inactivation by environmental stresses, such as biocidal agents and toxic chemicals, desiccation, pressure and temperature extremes, and high doses of UV and ionizing radiation (reviewed in Nicholson et al., 2000, 2002, 2005; Setlow, 2006). Spores of B. subtilis have been applied as biological dosimeters for terrestrial and extraterrestrial UV radiation (Horneck et al., 1996) and in outer space or in space simulation facilities for astrobiological studies on the likelihood of interplanetary transfer of life (Horneck et al., 1984, 1994a, 2001, 2002, 2008; Weber and Greenberg, 1985; Nicholson et al., 2000). Spores have been used especially for probing the biological effectiveness of different types of HZE particles as well as that of the impact parameter for each particle's trajectory, that is, the radial distance from the particle's trajectory. This was investigated in the first (micro)biological experiments within the Biostack device on NASA missions Apollo 16 and 17 (Horneck et al., 1974; Buecker et al., 1984). Since GCR is comprised of heavy ions of a wide spectral range in mass and energy, its mutagenic potential varies with physical properties of the ion under consideration. Previous experiments have indicated that the mutagenic effect of GCR in bacteria clearly depends on various parameters of the applied radiation and the biological target. The mutagenic efficacy of heavy ions in bacteria depends on physical (radiation dose, energy and charge of ion, track structure) and biological variables (bacterial strain, repair genotype of bacteria), and the endpoint investigated (type of mutation, induction of enzymes related to mutagenesis) (Facius et al., 1983, 1994; Kozubek et al., 1994; Schaefer et al., 1994).
To date, most of the biological endpoints studied in ionizing radiation–exposed biological systems, such as inactivation and DNA strand break induction, have shown similarities in their dependence on the particle parameters (Horneck and Buecker, 1983; Baltschukat et al., 1986a; Baltschukat and Horneck, 1991; Micke et al., 1992, 1994). The linear energy transfer (LET), that is, the energy transferred to material as an ionizing particle travels through it, is a generally used value to quantify the effects of ionizing radiation on biological systems. A main finding is that the efficiency per particle increases with increasing LET up to 100 keV/μm, independent of the ion applied. LET values from 200 to 1000 keV/μm show a saturated LET-response curve distribution, while particles with a LET above 1000 keV/μm describe the LET dependence as a separate function for each applied ion. Several radiobiological studies at accelerators and in space experiments have pointed out the assumption that at least two qualitatively different lesions are formed depending on the LET of the heavy ion irradiation (Takahashi et al., 1983; Horneck, 1993a, 1993b, 1994). Damage produced by low LET (<2 keV/μm) particles can be attributed to ionization and excitation processes or interactions with radicals. DNA strand breaks, base damage, or cross-linking between bases are likely to occur. For high LET (>2 keV/μm) particles, additional physicochemical processes, such as thermal spikes or shock waves, have been conjectured and may cause local destruction of ordered structures (Facius et al., 1983, 1994; Schaefer et al., 1994). Therefore, a bacterial spore hit by a heavy ion will either be killed, that is, lose the capability to germinate and give rise to a colony, or it will survive as a mutant. In the track core of HZE particles, spores will be inactivated with high probability, and mutations will not be produced. Mutations are most likely to be produced by high-energy secondary electrons (“δ-rays”) produced by interaction of the primary ions with matter (biological target). A particle of sufficiently high LET that passes through a spore is likely to kill it. Since such spores cannot give rise to mutants, it is reasonable to assume that mutants that arise are those induced by δ-rays from heavy ions passing near, but not through, the spore core. Further, it is reasonable to assume that δ-rays have an ability to induce mutations, which is comparable to that of low LET radiation (Hutchinson, 1985; Kozubek et al., 1994). Therefore, it is reasonable to distinguish between “direct hits” and “indirect hits” to the sensitive structure of the spore. Direct hit means the passage of the particle through the sensitive volume, that is, spore core, of a spore (0.2–0.3 μm2), whereas indirect hit means that the sensitive volume was hit by secondary electrons only; the particle itself has not passed through the spore core. The indirect hits are of great importance for the estimation of the effectiveness of accelerated, very heavy ions to induce mutations. The fraction of particle energy deposited in indirect hits obviously depends on the physical characteristics of the track of heavy ions and on the geometry of the sensitive structure of the spore (Facius et al., 1983, 1994; Schaefer et al., 1994; reviewed in Nicholson et al., 2000).
In this study, we examined the impact on spore survival of X-rays and four heavy ions (He, C, Si, and Fe) that differ in their LET values. To obtain more-detailed insights on the mutagenicity of these types of ionizing radiation, we also identified the mutagenic specificity in the nucleotide composition in the rpoB gene of B. subtilis that results in amino acid changes in RpoB, the β-subunit of RNA polymerase, one of the major enzymes that regulates the overall synthesis of RNA under the direction of DNA. Certain mutations in rpoB result in increased resistance to the antibiotic rifampicin (Rif ) (Wehrli et al., 1968; Campbell et al., 2001). Rifampicin-resistant (RifR) mutations have been isolated from numerous bacterial species and mapped to four small areas within the rpoB gene called Clusters I, II, III, and N (reviewed in Campbell et al., 2001; Severinov et al., 1993). To date, all the RifR mutations isolated in B. subtilis have been found to be single nucleotide substitutions that result in specific amino acid changes only within the Cluster I of the rpoB gene (Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins and Nicholson, 2008). Several lines of evidence indicate profound fundamental connections between the induced RifR and effects on the general microbial physiology, for example, growth, competence, sporulation, and metabolism (Maughan et al., 2004; Perkins and Nicholson, 2008). In this communication, we report on the spore sensitivity to HZE particle bombardment as a direct function of LET, and we report that ionizing radiation results in the induction of several novel RifR mutations in the B. subtilis rpoB gene.
2. Material and Methods
2.1. Bacillus subtilis spores, sporulation, purification, and sample preparation
Bacillus subtilis strain 168 (DSM 402) was obtained from the German Collection of Microorganism and Cell Cultures (DSMZ) GmbH, Braunschweig, Germany, and was used throughout. Spores were obtained by cultivation under vigorous aeration in double-strength liquid Schaeffer sporulation medium (Schaeffer et al., 1965), and the spores were purified and stored as described (Nicholson and Setlow, 1990; Moeller et al., 2006). Spore preparations were free (>99%) of growing cells, germinated spores, and cell debris, as determined by phase-contrast microscopy.
2.2. Spore exposure to sparsely ionizing radiation (X-rays)
Air-dried spore layers of 1 × 109 spores on 50 × 50 mm glass slides (as layers approximately two spores thick) were irradiated at room temperature with X-rays (150 keV/19 mA) generated by an X-ray tube (Mueller Type MG 150, Germany) at various doses up to a final dose of 0.5 kGy, as described previously (Moeller et al., 2007, 2008b).
2.3. Spore exposure to densely ionizing radiation (accelerated heavy ions)
Air-dried spore layers (in each case three samples of 5 × 108 spores on 25 × 25 mm glass slides; as layers approximately two spores thick) were irradiated simultaneously at room temperature with one of the four heavy ions—helium (He), carbon (C), silicon (Si), and iron (Fe)—in the LET range from 2 to 200 keV/μm, up to a final dose of 0.5 kGy. Further information (i.e., energy, LET, and beam intensity) of the applied accelerated heavy ions is shown in Table 1. Heavy ion irradiations were performed at the Heavy Ion Medical Accelerator (HIMAC) at the National Institute for Radiological Sciences in Chiba-shi, Japan, under the aegis of the HIMAC research project 17B463 “Gene activation of heavy ion treated Bacillus subtilis 168 (DSM 402) endospores during germination involved DNA-repair” in the years 2005–2009 (Moeller et al., 2008b). All irradiations were carried out in the Biological Irradiation Room, where a maximum beam diameter of 10 cm is obtained by using a pair of wobbler magnets and a scatterer. Reference doses were retrieved from a high-precision, Farmer-type ionization chamber of 1 mm water-equivalent thickness installed upstream of the target, which was also used to check uniformity of the beam in one dimension, yielding maximum fluctuations of <5% over the circular beam area for the employed ion species. Further details on the irradiation geometry of the HIMAC facility, beam monitoring, dosimetry, and dose calculations have been described in detail by Okayasu et al. (2006).
D37 value = dose (Gy) of X-ray and HZE particle irradiation leading to a 63% inactivation of the initial spore population (Moeller et al., 2008b).
k i value = spore inactivation constant (Gy−1).
k m value = mutation induction constant, determined from the slope of the exponential mutation induction curve (Fig. 2).
X-rays (150 keV/19 mA) were generated by an X-ray tube (see Material and Methods).
Value obtained by extrapolation.
Data are expressed as averages and standard deviations (n = 3). Asterisks (*) indicate D37, k i, and k m values that were significantly different (P values of ≥0.05) than the respective values of the X-ray irradiation.
2.4. Survival assay
To recover the spores from the quartz discs after irradiation, air-dried spore layers were covered by a 10% aqueous polyvinyl alcohol solution, and after drying the spore–polyvinyl alcohol solution layer was stripped off as described (Horneck et al., 2001; Moeller et al., 2007) and resuspended in 1 ml sterile distilled water, which resulted in >95% recovery of the spores. This procedure does not affect spore viability (Horneck et al., 2001). Spore survival was determined from appropriate dilutions in distilled water as colony-forming ability (of vital colony-forming unit) after incubation overnight at 37°C on nutrient broth (NB) agar plates (Difco, Detroit, USA), as described previously (Horneck et al., 2001; Moeller et al., 2007).
2.5. Assay for mutation induction to RifR
For studying the mutagenic effects of X-ray and heavy ion irradiation to RifR, treated spores and non-treated control spores were plated in appropriate dilutions in sterile distilled water on NB agar plates containing Rif (Merck KGaA, Darmstadt, Germany) at a final concentration of 50 μg/ml, as described previously (Nicholson and Maughan, 2002). RifR mutants that arose after overnight incubation at 37°C were counted. The radiation-induced RifR mutation frequencies (RifR frequency) were measured and calculated according to Horneck et al. (1994b) with the following equation:
CFU-RifR, colony formers carrying radiation-induced RifR mutations (after irradiation); control: colony formers carrying spontaneous RifR mutations (control sample); total CFU, total colony formers (after irradiation).
2.6. Sequencing and characterization of rpoB mutations
The mutagenic specificity of RifR mutations in rpoB was analyzed by polymerase chain reaction (PCR) as follows: RifR colonies were picked and incubated in 5 ml NB medium (containing 50 μg Rif/ml) overnight at 37°C. One milliliter of the culture was centrifuged at 12000 × g for 5 min at 4°C; the cell pellet was resuspended in 0.2 ml TE buffer (10 mM Tris-HCl, 1 mM Na2EDTA; pH 8.0). Template chromosomal DNA was prepared by heating the cell suspension at 95°C for 10 min, cooling to room temperature, extraction with 0.2 ml chloroform/isoamyl alcohol (v/v, 96/4; Merck KGaA, Darmstadt, Germany), and centrifugation at 14500 × g for 90 s in a microcentrifuge for removing cellular debris. The upper phase was transferred to a clean microcentrifuge tube and stored at 4°C until further analysis.
Rifampicin-resistant (RifR) mutations have been mapped to four small areas within the rpoB gene called Clusters I, II, III, and N (reviewed in Campbell et al., 2001). The N-cluster is located near the beginning of rpoB and consists of amino acid changes that alter the single amino acid V135 (Note: B. subtilis coordinates are used throughout). In the middle of the rpoB coding sequence are three closely linked regions: Cluster I, which spans amino acids 463-GSSQLS-QFMDQTNPLAELTHKRRLSALG-490; Cluster II, which spans amino acids 519-TPEGPNIGLIN-529; and Cluster III, which consists of the single amino acid R645 (Severinov et al., 1993; Campbell et al., 2001). To study the mutagenic specificity in the nucleotide composition in the rpoB gene of B. subtilis, a region of rpoB was amplified by PCR with the primer pair 5′-ACGGAGTAGGCGACACAGAT and 5′-AACGAGC-TACGATGCTGTCA flanking a 568 bp region spanning B. subtilis rpoB nucleotides 1198–1766, which corresponds to amino acids G394 to I588 in the deduced B. subtilis RpoB protein sequence and contains the RifR Clusters I and II as defined in E. coli rpoB (Severinov et al., 1993). Each PCR reaction was performed on 5 μl template DNA (0.15–0.2 μg DNA/μl), with each 1 μl (20 pmol) of the primer pairs, 25 μl H2O, and 20 μl PCR ready-to-use mixture (HotStart-PCR Kit, Qiagen, Hilden, Germany) in 35 thermal cycles of 95, 60, and 72°C for 45, 60, and 90 s, respectively (MiniCycler Bio-Rad, CA, USA). The PCR products were purified by using a column-based DNA cleanup system (MiniElute PCR Purification Kit, Qiagen, Hilden, Germany). The sequence determinations of the PCR products were carried out by AGOWA DNA sequencing service (AGOWA, Berlin, Germany). The DNA sequences were translated to the predicted amino acid sequence by using the DNAsis program (MedProbe, Oslo, Norway). The amino acid sequence was then compared with sequences from the non-treated control, respectively, as well as with sequences available from previous studies that reported mutation induction in rpoB (Nicholson and Maughan, 2002; Perkins et al., 2008). Alignment of the sequences was carried out with the CLUSTALW program (EMBL-EBI, Cambridge, UK). Chromosomal DNA was prepared from strains carrying rpoB mutations by the method of Cutting and Vander Horn (1990) and introduced by transformation into the initial wild-type B. subtilis strain 168 (DSM 402) as described previously (Maughan et al., 2004). RifR transformant strains resulting from these crosses were checked for all expected markers on NB agar plates with and without Rif (50 μg/ml) and streak purified; their rpoB regions were again PCR amplified and sequenced, as described above, to confirm that the desired mutations had been transferred successfully.
2.7. Numerical and statistical analysis
The surviving fraction of B. subtilis spores was determined from the quotient N/N 0, with N = the number of colony-forming units of the irradiated sample and N 0 that of the non-irradiated controls. To determine the curve parameters, the following relationship was used: ln (N/N 0) = − k i × D + n, with k i = spore inactivation constant (Gy−1), D = dose (Gy) and n = extrapolation number, that is, the intercept with the ordinate of the extrapolated semilog straight line. The constants k i and n were determined by linear regression. Spore survival was plotted as a function of X-rays and heavy ion irradiation dose. Data are reported as D37 values, that is, the dose lethal for 63% of the initial spore population, determined from the linear portion of the semilogarithmic curve [i.e., the reciprocal of the spore inactivation constant k i; according to Moeller et al. (2008b)]. Similar to the spore inactivation constant k i, the mutation induction constant for X-ray and each heavy ion exposure was obtained from the slope of each respective exponential mutation induction curve. The ratio of the spore inactivation constant (k i) and the mutation induction constant of the X-ray to the heavy ion–irradiated spores was used for data normalization to determine the sporicidal and mutagenic effectiveness of the applied HZE particle. These latter values reflect the biological impact of the LET of heavy ion irradiation on spore survival and induced RifR mutations. Each experiment was repeated at least three times, and the data shown are expressed as averages ± standard deviations. The results were compared statistically via the Student t test. Values were analyzed in multigroup pairwise combinations, and differences with P values of ≤0.05 were considered statistically significant (Moeller et al., 2007, 2008a, 2008b).
3. Results
To study the impact of X-ray and heavy ion irradiation as lethal stressor and potential mutation inducer, spores of B. subtilis 168 were exposed as air-dried spore layers to sparsely ionizing (i.e., X-rays) and densely ionizing (i.e., HZE particles) radiation.
3.1. Spore survival after exposure to X-ray and HZE particle irradiation
Survival curves for air-dried spores of B. subtilis 168 in response to X-rays and the heavy ions He, C, Si, and Fe, in an LET range of 2–200 keV/μm, are depicted in Fig. 1. After exposure to X-rays or HZE particles, spores exhibited strict exponential inactivation kinetics, as indicated by n (the ordinate of the extrapolated semilog straight line) approximating 1 (Fig. 1). Spore resistance was expressed as the D37 value (Table 1). Spores were significantly more sensitive to high LET irradiation than to low LET and X-ray irradiation. In comparison to the determined D37 values of the irradiated spores, the following order of radiation sensitivity to HZE particles and X-rays was obtained: Fe ions > Si ions > C ions > He ions > X-rays. These observations are in good agreement with the results from previous experiments with X-ray, He, and Fe ion irradiation (Moeller et al., 2008b).
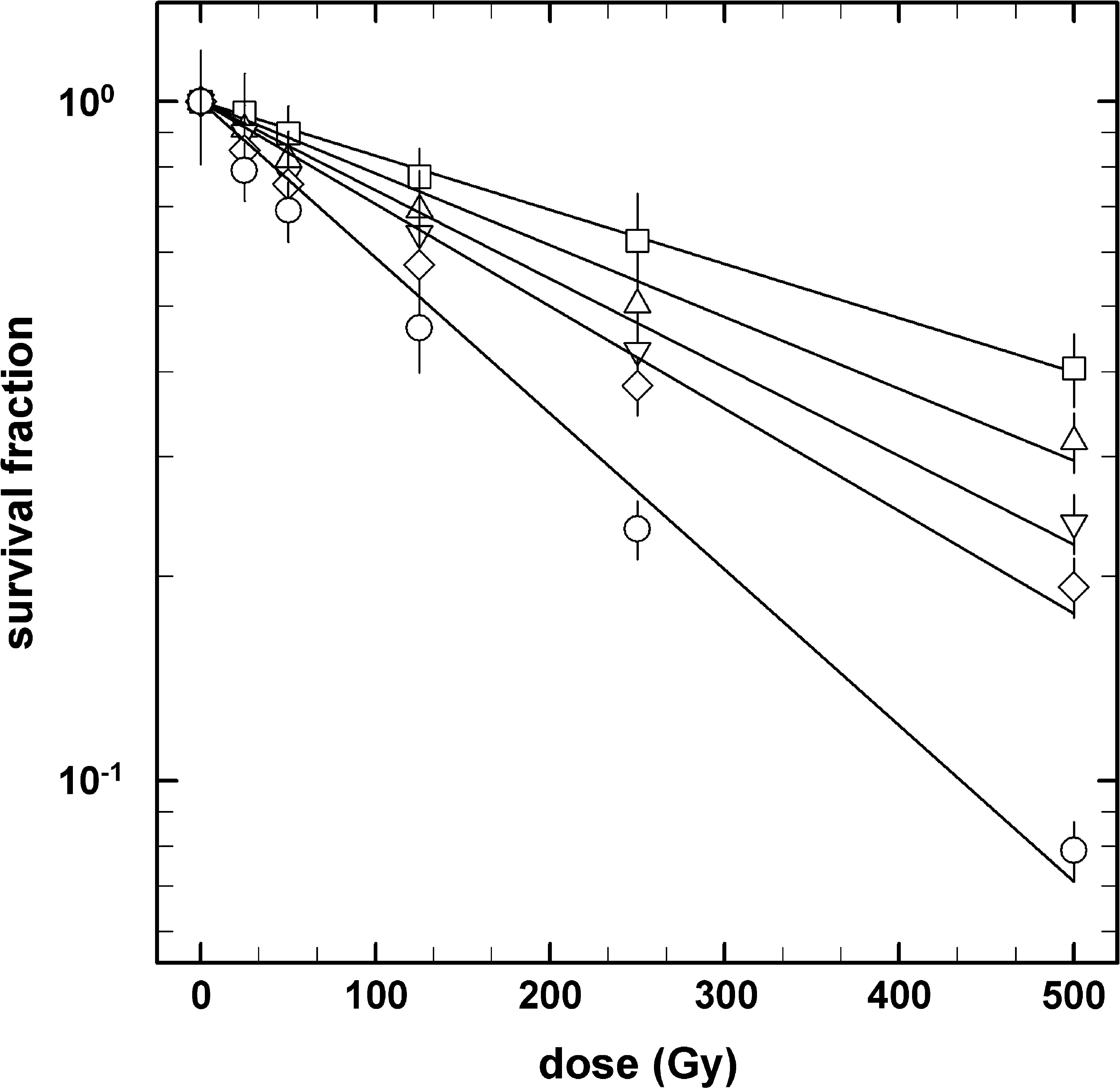
Survival curves of B. subtilis spores in response to X-rays (open squares) and accelerated heavy ions helium (open triangles up), carbon (open triangles down), silicon (open diamonds), and iron (open circles). Data are expressed as averages and standard deviations (n = 3).
Spores of B. subtilis 168 showed different sensitivities to X-rays and HZE particle bombardment, which is in good agreement with previous reports by Horneck and Buecker (1983) and Micke et al. (1994). Exposure of spores to He ions, C ions, Si ions, and Fe ions resulted in 1.4-, 1.7-, 1.9-, and 3.0-fold-greater inactivation, respectively, than did X-ray exposure, based on comparison of spore inactivation constants of each irradiation (Fig. 1 and Table 1). These results can be explained in terms of the LET values of the X-rays and HZE particles used. Spores were more sensitive to HZE particles than to X-rays and were more sensitive to HZE particles with higher LET values (Table 1), which is in good agreement with previous observations (Baltschukat and Horneck, 1991; Horneck, 1994).
3.2. Mutation to RifR by X-ray and HZE particle irradiation
It has been shown by various authors that exposure of B. subtilis spores to simulated and real space conditions, such as ultrahigh vacuum (UHV) and extraterrestrial solar UV radiation, leads to higher mutation frequencies in selected genetic markers, for example, specific amino acid auxotrophies and prototrophies (e.g., histidine revertants), sporulation ability (e.g., sporulation deficiency), and antibiotic resistance (e.g., resistance to nalidixic acid or Rif ) (Horneck et al., 1984, 1994a; Munakata et al., 1997, 2002, 2004; Fajardo-Cavazos et al., 2005). In our study, we investigated the impact of sparsely and densely ionizing radiation, either obtained by X-ray or heavy ion irradiation, on the mutagenic factor spores have to encounter in a putative travel through space. Specifically, we measured the mutation frequencies to RifR of X-ray– and HZE particle–exposed spores. Mutation to RifR was assayed on NB agar containing 50 μg of Rif per ml (as described in Material and Methods). The mutation induction as a function of the applied dose of X-ray and heavy ion irradiation is shown in Fig. 2. The non-irradiated controls showed a spontaneous mutation rate in the range of (2.9 ± 0.7) × 10−9 for wild-type B. subtilis spores, which is in good agreement with data from Nicholson and Maughan (2002). Within experimental errors, exponential mutation induction curves were obtained. With increasing LET of the applied ions, the mutation induction rate increased (Fig. 2). These findings confirm previous studies on mutation induction in B. subtilis spores—in that case to resistance to sodium azide—by accelerated heavy ions, which showed an increasing mutation induction rate with rising LET of the particle radiation (Baltschukat et al., 1986b). After irradiation at 0.5 kGy (the highest dose applied in this study), the mutation rates increased by about one order of magnitude for X-rays and by about 2.5 orders of magnitude for Fe ions, which leads to the notion that irradiation with high LET particles (>200 keV/μm) is more mutagenic to spores than low LET particle or X-ray exposure.

Dose-effect curves for mutation induction to Rif (rifampicin) in B. subtilis spores exposed to X-rays (solid squares) and accelerated heavy ions helium (solid triangles up), carbon (solid triangles down), silicon (solid diamonds) and iron (solid circles). Data are expressed as averages and standard deviations (n = 3).
3.3. Relative effectiveness of HZE particles on spore inactivation and mutagenesis
To determine the sporicidal effectiveness of HZE particles relative to X-rays for spores, the relative biological effectiveness (RBE) was calculated from the ratio of the spore inactivation constant of each heavy ion species to the spore inactivation constant of X-rays (Fig. 3). After exposure to the same doses of HZE irradiation, it was observed that spore survival is dependent on the LET of the applied heavy ion species. Likewise, when analyzing the ratio of the mutation induction constants from the HZE particle irradiation with the mutation induction constant of the X-ray-exposed spores, the relative effectiveness for the induction of RifR mutants was also observed to increase with increasing LET of the applied heavy ion species, similar to the sporicidal RBE (Fig. 3). The data for spore inactivation are well in agreement with previous studies, for example, by Powers et al. (1968), on heavy ion–irradiated dry spores of Bacillus megaterium, Takahashi et al. (1983) on spores of B. subtilis, as well as Suzuki et al. (1996) for inactivation of human embryonic fibroblast-like cells. For the inactivation of all systems tested, particles at LET of 100–200 keV/μm possessed maximum RBE reaching values around 3 (IAEA, 2008). Particles at higher LET values reach the “overkill” region where the amount of energy deposited in a cell by a single particle traversal is in excess of the amount required to kill the cell. It is interesting to note that in the spores the RBE of both inactivation and mutagenicity reach similar values, whereas in the fibroblasts the RBE for mutagenicity was about twice as high as that for cell inactivation (Suzuki et al., 1996).
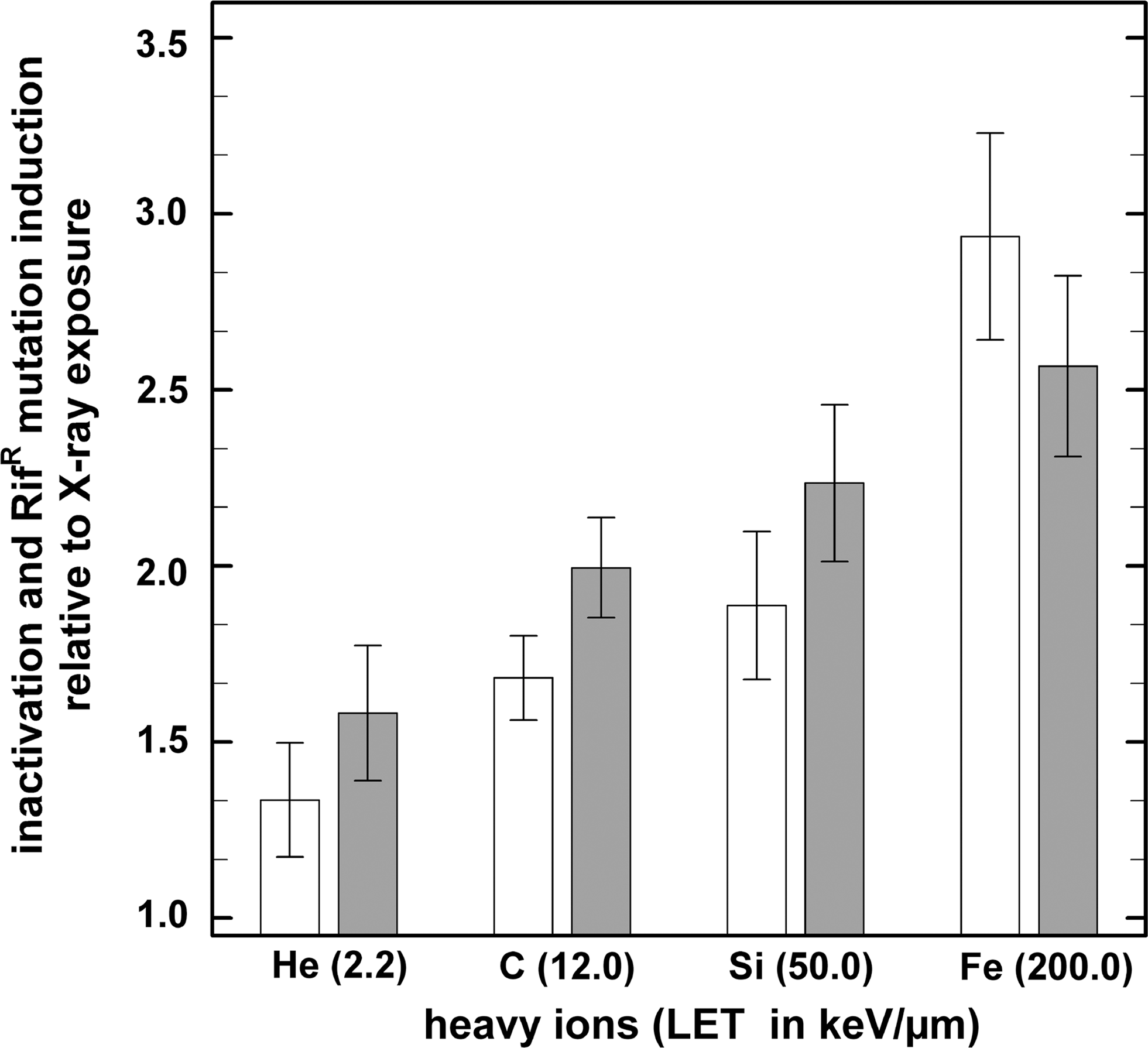
Inactivation (white bars) and RifR mutation induction (gray bars) of HZE particle–exposed B. subtilis spores relative to X-ray irradiation as a function of LET. The ratio of the spore inactivation constant (k i) and the mutation induction constant of the X-ray– to the heavy ion–irradiated spores was used for data normalization to determine the sporicidal and mutagenic effectiveness of the applied HZE particle. Data are expressed as averages and standard deviations (n = 3).
3.4. Spectrum of RifR mutations in the rpoB gene of B. subtilis
DNA sequencing was performed on a total of 21 isolates, which consisted of 3 and 18 RifR mutants isolated from X-ray– and HZE particle–irradiated samples, respectively (Table 2, Fig. 4). The majority of the mutations (20/21) were localized to Cluster I, in codons L467 (1/21), Q469 (5/21), A478 (3/21), H482 (5/21), R484 (3/21), S487 (2/21), and A488 (1/21) (Fig. 4). The remaining one RifR mutation, at H507, was located between Clusters I and II (Table 2). No mutations were found in the N-cluster or Clusters II or III (data not shown; it should be noted that other primer pairs, encompassing the entire rpoB gene, were used in these experiments). Codons Q469 (CAA) and H482 (CAC) were definite hot spots for RifR mutations, and each accounted for ∼24% of the total; codons A478 (GCT) and R484 (CGT) accounted each ∼14% of the total (Fig. 5).
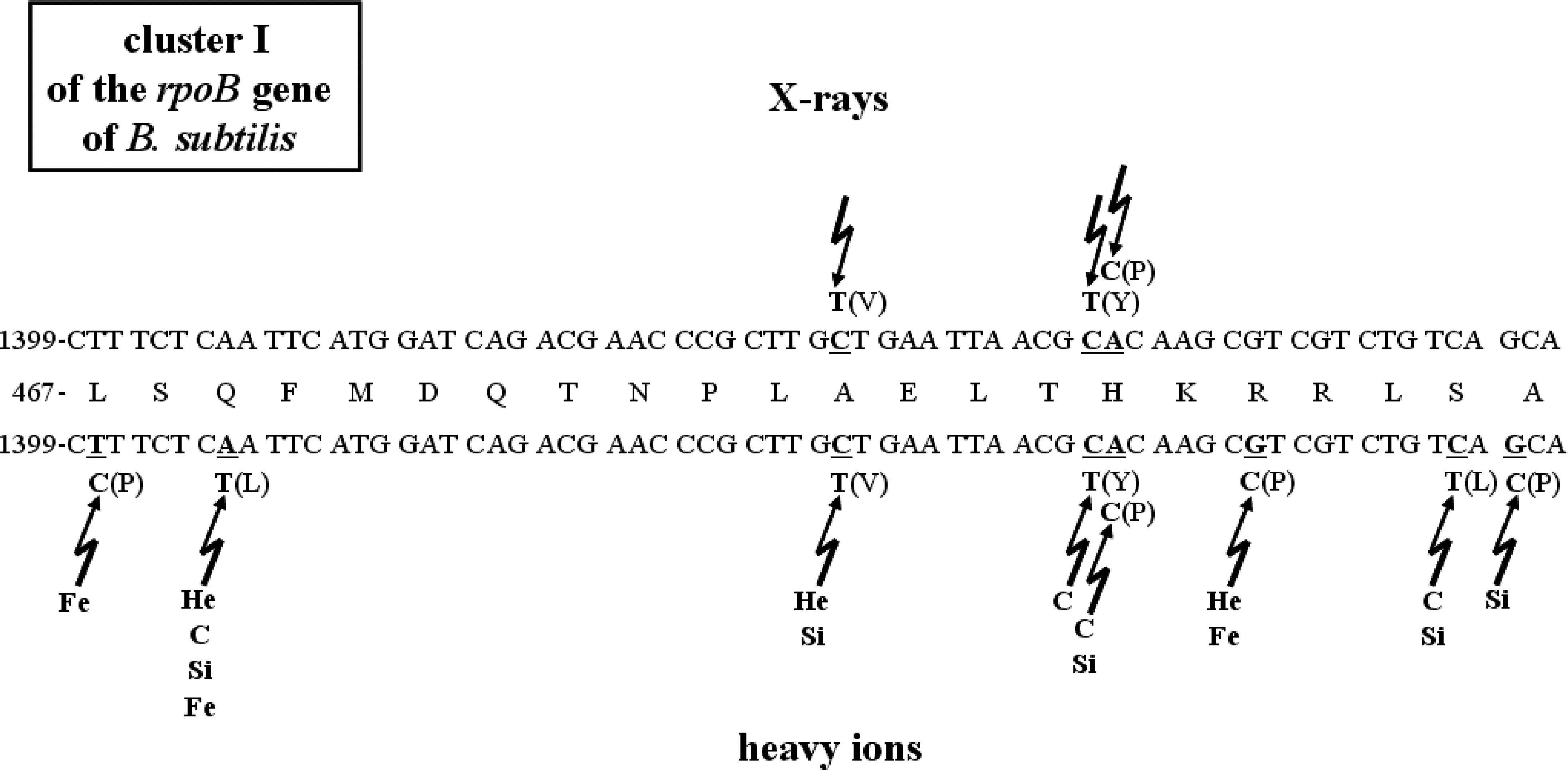
Mutations in Cluster I of B. subtilis rpoB gene from spores exposed to X-rays (top) and accelerated heavy ions (bottom). The central line depicts the wild-type amino acid sequence in Cluster I, with the wild-type nucleotide sequence above and below. Nucleotides suffering changes are denoted by underlined letters. Nucleotide changes are indicated, and the resulting amino acid changes are in parentheses.
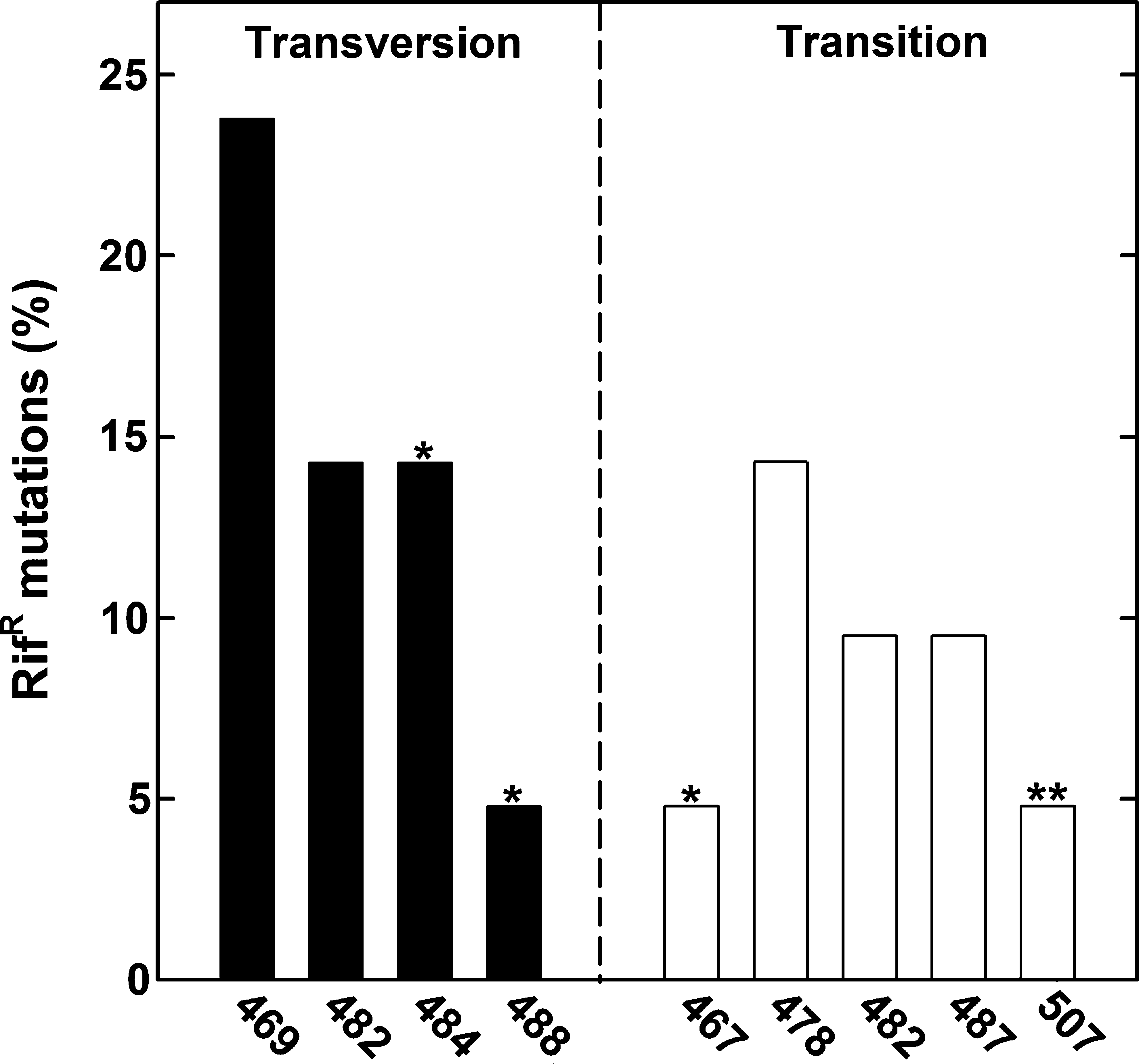
Frequency of RifR mutations (transversion or transition mutation) induced in B. subtilis spores by ionizing radiation (X-rays and accelerated HZE particles), determined in the sequenced part of Cluster I of the rpoB gene of B. subtilis 168. rpoB mutations newly identified in the framework of this study [marked by asterisk(s)] in Cluster I arising from irradiated spores were detected: L467P, R484P, and A488P, as well as one located codon change H507R (labeled with two asterisks) between Clusters I and II (according to Campbell et al., 2001).
Codon positions (CP) using B. subtilis 168 coordinates.
aa change = amino acid change.
1- and 3-letter code for the amino acids: A ala (alanine), H his (histidine), L leu (leucine), P pro (praline), Q gln (glutamine), R arg (arginine), S ser (serine), V val (valine), Y tyr (tyrosine).
21 RifR mutations were obtained and sequenced from X-ray– and heavy ion–irradiated B. subtilis spores after cultivation (germination and outgrowth) of their vegetative cells.
The spectrum of mutations to RifR in rpoB from spores irradiated with X-ray and accelerated heavy ions was compared (Table 2, Fig. 4). The majority of the RifR mutations were C-to-T transitions (∼34% of the total), A-to-T transversions (∼24% of the total), and G-to-C transversions (∼19% of the total) (Fig. 6). Interestingly, the A-to-T transversions were localized to only one hot spot within Cluster I: A1406T in the first nucleotide of codon Q469, which resulted in the amino acid change Q469L (Table 2, Fig. 5). Only one mutation was found with a T-to-C transition: T1401C in the second nucleotide of codon L467, which resulted in the amino acid change L467P (Table 2, Fig. 4). The second most abundant mutations were C-to-T transitions in three codons: G1434T, which resulted in the amino acid change A478V, C1445T in the amino acid change H482Y, and C1461T in the amino acid change S487L (Table 2, Fig. 4). The third most abundant category of mutations was G-to-C transversions (Fig. 6). This class of mutations was located in Cluster I as G1452C, which resulted in the amino acid change R484P, and G1463C, which resulted in the amino acid change A488P in RpoB (Fig. 4). G1463C marks a nucleotide substitution of guanine to cytosine at base position 1463, and thereof the A488P denotes a mutation of alanine to proline at site 488, where the surface charge for basic amino acids is conserved. Interestingly, one RifR mutation outside Cluster I, an A-to-G transition (A1521G), which resulted in the amino acid change H507R (Table 2), was isolated after high LET iron ion exposure. Out of the eight amino acids suffering changes as a result of spore exposure to HZE particle irradiation, fully half (L467, R484, A488, and H507) represent new RifR mutations not previously described in the B. subtilis rpoB gene (Rothstein et al., 1976; Boor et al., 1995; Ingham and Furneaux, 2000; Nicholson and Maughan, 2002; Perkins et al., 2008).
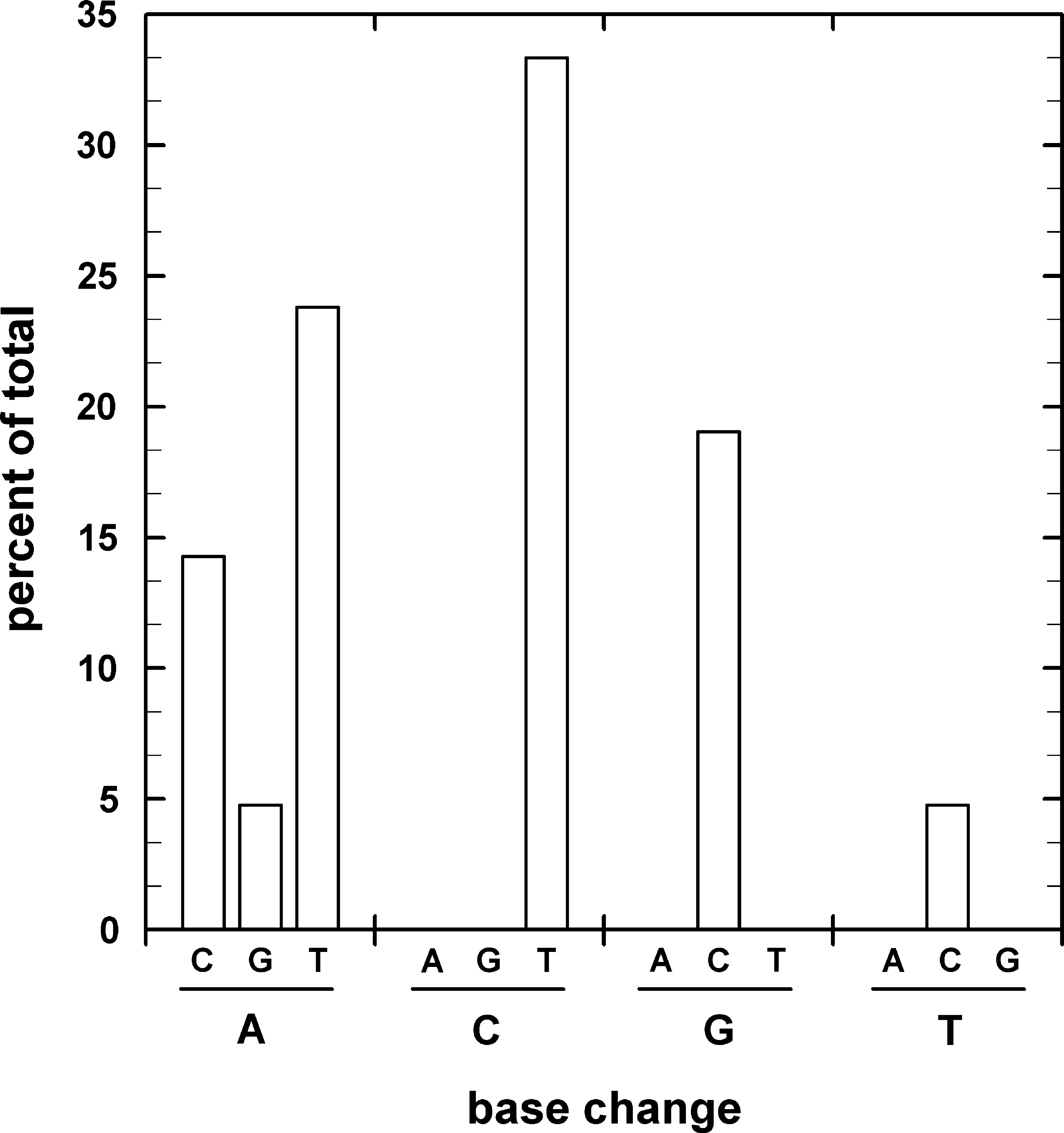
Relative frequency of mutations in rpoB of B. subtilis spores exposed to X-rays and accelerated heavy ions.
4. Discussion
Among the ionizing components of radiation in space, HZE particles are the most biologically effective species (reviewed in Horneck, 1993b). Bacterial spores are optimal models in experimentally addressing questions of the effects of cosmic radiation on biological systems. Such HZE particles of cosmic radiation are conjectured to set the ultimate limit on the survival of spores in space because they penetrate even thick shielding. However, since the flux of HZE particles is relatively low, a spore may exist in space for several hundred thousand years before being hit by an HZE particle (e.g., iron of LET >100 keV/μm) (Horneck et al., 2002). This time span complies with estimates of the time required for boulder-sized rocks to travel from one planet of our solar system to another, for example, from Mars to Earth (Melosh, 1988; Nicholson, 2009). However, only a few months have been estimated to be sufficient for an interplanetary transfer of a fraction of meteorites ejected from Mars (Melosh, 1988; Mileikowsky et al., 2000). As reported previously by Facius et al. (1983, 1994) and Weisbrod et al. (1992), B. subtilis spores may survive even a central hit of an HZE particle of cosmic radiation to a certain percentage. In our experiments on dormant spores of B. subtilis 168 irradiated with components of GCR, that is, X-rays and high-energy charged particles up to a dose of 500 Gy, we demonstrated that a substantial fraction of spores is capable of surviving the exposure of galactic cosmic rays that occur during the second stage of lithopanspermia, the travel through space. Assuming an average of 0.2–0.5 Gy per year (depending on various physical parameters, e.g., solar activity, contribution of high- and low-energy charged particles) in the open space environment, as reported by Cucinotta and Durante (2006), spores at the surface of a meteorite (but shielded from solar UV) would require approximately 1156–2890 years (for X-rays) and approximately 354–885 years (for high-energy-charged iron ions) to reach the D37 value (37% survival of the initial spore population). Concerning shielding against radiation in space, a few micrometers of meteorite material are sufficient to give efficient protection against solar UV radiation if the material is without cracks (Horneck et al., 2001), whereas higher shielding requirements are necessary to protect against X-rays and HZE particles (Mileikowsky et al., 2000; Nicholson et al., 2000; Valtonen et al., 2009). However, one has to bear in mind that radiation is only one of the harmful parameters in space and that for the success of a viable transfer the spores have ultimately to cope with the complex matrix of all environmental factors governing the space environment (Mileikowsky et al., 2000; Nicholson, 2009; Valtonen et al., 2009).
The cellular response to X-ray and heavy ion irradiation is complex due to the variety of targets in a cell (Goodhead, 1994; Kiefer et al., 1996). Ionizing radiation can damage cellular components through direct deposition of radiation energy into biomolecules and also indirectly by generating reactive oxygen species (Hutchinson, 1985). Hydrogen peroxide (H2O2) and hydroxyl radicals (HO·) are major oxidizing species produced by the radiolysis of water, and superoxide ions (
Previous studies with spores of B. subtilis exposed to accelerated heavy ions have shown an increased mutation frequency, based on the resistance to various chemical agents, for example, sodium azide and nalidixic acid, increasing with the applied dose and rising LET (Baltschukat et al., 1986b; Baltschukat and Horneck, 1991; Horneck et al., 1994b; Munakata et al., 1997). Reports of mutations in gyrA, the gene encoding DNA gyrase subunit A resulting in resistance to nalidixic acid, refer to the induction of unique tandem-base change mutation in one particular allele of gyrA (gyrA12; 5′-CA to 5′-TT) after irradiation with UV, vacuum UV, γ radiation, and soft X-rays (Munakata et al., 1997). In our experiments, we studied the mutagenicity of the rpoB gene after exposure to X-ray and heavy ion exposure. Mutations in the rpoB gene have dramatic effects on the general microbial physiology but refer as nonlethal events. Nevertheless, it should be kept in mind that lethal mutations are very likely after exposure to X-ray and HZE particle irradiation; this type of mutation leads to the inactivation of the irradiated spore. One of the most investigated mutations in B. subtilis is the loss of the individual sporulation capability, the so-called sporulation-defective (Spo−) mutation, as illustrated and described in detail by Fajardo-Cavazos et al. (2005). While the detection of Spo−, by visual inspection of the colony morphology and pigmentation, is quite simple, detailed information on the nature and type of mutations in Spo− cells are less available. Even though it is known that the loss of sporulation is a dramatic change in the life cycle of B. subtilis cells, it remains to be investigated whether and to what extent sporulation-defective are mutants induced by X-ray and HZE particle exposure. However, it should be noted that, in B. subtilis, sporulation genes comprise almost 5% of the genome (Maughan et al., 2007). In this regard, the induction of single-point mutations, as reported in the rpoB gene causing RifR, are more suitable for a detailed investigation of ionizing radiation-induced mutagenesis than lethal events such as Spo−. Besides the aspect of the induction of lethal mutations, it should be noted that beneficial mutations could also be induced (reviewed in Rainey, 1999; Perfeito et al., 2007). Beneficial mutations are known to increase microbial fitness, such as higher adaptation to environmental extremes (e.g., temperatures, salinity, pressure), specific resistance to chemicals (e.g., antibiotics), new metabolic pathways (Hall and Zuzel, 1980; Bennett et al., 1992; Papadopoulos et al., 1999; Imhof and Schlotterer, 2001).
Macromolecular machines, such as the replication, transcription, and translation complexes, can be viewed as central control devices that optimize the flow of genetic information to the various cellular tasks needed for sensing and responding to environmental cues. It has been well established for the Gram-positive spore-forming bacterium B. subtilis that growth and metabolism, as well as several developmental events (e.g., extracellular enzyme synthesis, motility, chemotaxis, competence, sporulation, spore resistance properties, germination, and outgrowth), are controlled in large part at the level of transcriptional activity (Maughan et al., 2004). RNA polymerase is a central macromolecular machine that controls the flow of information from genotype to phenotype, and insights into global transcriptional regulation can be gained by studying mutational perturbations in the enzyme. Mutations in the RNA polymerase β-subunit gene rpoB causing resistance to Rif in B. subtilis were previously shown to lead to alterations in the expression of a number of global phenotypes known to be under transcriptional control, such as growth, competence for transformation, sporulation, and germination (Maughan et al., 2004; Perkins and Nicholson, 2008). When B. subtilis spores were exposed to UHV that mimicked space conditions (∼10−5 to 10−2 Pa), a 4-fold increase in the induction of RifR mutations in rpoB was observed (Munakata et al., 1997, 2002, 2004). Exposure of B. subtilis spores to UHV not only increased mutation frequency in spores but also altered the spectrum of mutations observed. Similar UHV exposure experiments with B. subtilis spores showed a predominance of CA-to-TT mutations that resulted in the amino acid change S487F in the B. subtilis rpoB gene leading to RifR (Munakata et al., 2002, 2004). Previous experiments on RifR mutations in B. subtilis indicated that the rpoB gene is indeed sensitive to alterations in the intracellular environment of DNA (Nicholson and Maughan, 2002) and, further, these mutations in rpoB can confer novel phenotypes upon the resulting RifR mutants (Maughan et al., 2004; Perkins and Nicholson, 2008). Especially, the mutations Q469L, H482P, and S487L, which we have also found after HZE particle irradiation (Table 2), have been reported to exhibit dramatic alterations in metabolism pattern in the utilization of various common and novel substrate classes, for example, sugars, carbohydrates, and amino acids (Perkins and Nicholson, 2008). In regard to the differences in the metabolism pattern of specific sugars, for example, D-ribose, the backbone of RNA, the S487F mutation exhibits dramatic reductions in ribose utilization to below background level of the wild-type strain (Perkins and Nicholson, 2008). In particular, it was intriguing that replacement of S487 by the similar aromatic residues F versus Y or of Q469 by the similar basic amino acids K versus R should result in such disparate effects (Perkins and Nicholson, 2008). These and several other observations suggest the intriguing possibility that mutations induced in key targets, such as rpoB, during exposure to space radiation might endow the resulting mutants with enhanced fitness in new habitats, for example, Mars. Such new phenotypes include changes in key microbial events, for example, growth, competence for transformation, sporulation, and germination of B. subtilis. For example, the S487L mutation showed slower growth and weaker response to temperature shifts as well as absence of competence for transformation (Maughan et al., 2004), which clearly implies that mutations in global regulators such as RpoB have selective advantages or disadvantages, or both, on the microbial propagation when encountering new habitable environments.
Perkins et al. (2008) found that the frequency of RifR mutations was not significantly elevated in spores exposed to simulated martian conditions (pressure and atmosphere) compared to control sets of spores exposed in parallel to Earth. In general, the spectrum of rpoB mutations was found to be very similar in spores exposed to Earth versus simulated martian conditions (Perkins et al., 2008). Spores exposed to simulated martian conditions had the following mutations in rpoB: Q469L, H482Y, S487L (Perkins et al., 2008). Munakata et al. (2002, 2004) reported that the predominant rpoB mutation isolated from spores exposed to UHV was S487F, a tandem CA-to-TT mutation. In the present study, we found Q469L, A478V, H482P, and R484P as major rpoB mutations (Table 2, Fig. 4); these mutations can be considered here and from previous reports as well-documented preferential hot-spot regions in the rpoB gene (Munakata et al., 1997, 2002, 2004; Nicholson and Maughan, 2002; Maughan et al., 2004; Perkins and Nicholson, 2008). Further on, four new mutant alleles of rpoB, which have never before been reported in B. subtilis, were isolated in these experiments, which resulted in the amino acid changes L467P, R484P, A488P, and H507R (Table 2, Fig. 5). The results of this study strongly suggest that exposure to galactic cosmic rays may indeed result in a broader spectrum of mutations in spores than exposure to other space or martian conditions (Munakata et al., 2002, 2004; Perkins et al., 2008). Furthermore, these findings clearly indicate that mutagenesis within the rpoB gene shows a differential response to varying parameters encountered in the spaceflight environment, which have profound impact on the implications for the interplanetary transport of terrestrial microbes to Mars by impacts (i.e., lithopanspermia) or robotic or human planetary exploration missions (i.e., planetary protection). Further investigations of various RifR mutants are ongoing to elucidate detailed insights into the microbial physiology and competitive fitness to harsh terrestrial and extraterrestrial conditions.
Footnotes
Acknowledgments
The authors are very grateful to Takeshi Murakami, Takamitsu Kato, Hisashi Kitamura, Maki Okada, Momoko Takahashi, Yasuharu Ninomiya, Emiko Sekine, and all HIMAC operators for the excellent technical assistance during heavy ion irradiations. W.L.N. was supported by grants from the NASA Planetary Protection (NNA06CB58G) and Astrobiology: Exobiology and Evolutionary Biology (NNX08AO15G) programs. These results will be included in the research reports of the HIMAC project (20B463).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
GCR, galactic cosmic radiation; HIMAC, Heavy Ion Medical Accelerator; HZE, high-energy charged; LEO, low Earth orbit; LET, linear energy transfer; NB, nutrient broth; NHEJ, non-homologous end-joining; PCR, polymerase chain reaction; RBE, relative biological effectiveness; Rif, rifampicin; RifR, rifampicin resistance; SAA, South Atlantic Anomaly; SCR, solar cosmic radiation; SPEs, solar particle events; UHV, ultrahigh vacuum.