
Abstract
Asymmetrical utilization of chiral compounds has been sought on Mars as evidence for biological activity. This method was recently validated in glucose. Earth organisms utilize D-glucose, not L-glucose, a perfect asymmetry. In this study, we tested the method in lactate and found utilization of both enantiomers. Soil-, sediment-, and lake-borne microbial communities prefer D-lactate but can consume L-lactate if given extra time to acclimate. This situation is termed imperfect asymmetry. Future life-detection mission investigators need to be aware of imperfect asymmetry so as not to miss relatively subtle signs of life. Key Words: Chirality—Homochirality—Life detection—Atacama Desert. Astrobiology 10, 397–402.
1. Introduction
T
An effort to test nutrient stereospecificity was recently initiated in our laboratory. Substrate stereospecificity on the enzyme level is well established. For example, Milton and colleagues (1992) showed that the native, L-form of the HIV-1 protease, composed of L-amino acids, interacts only with the L-form peptide substrate (composed of L-amino acids), not with the D-form. The reverse is true if the enzyme is in the D-form, synthesized from D-amino acids. It binds the D-form, not the L-form, of the substrate. Our objective is twofold: (1) to verify that what is observed with enzymes can be extrapolated to whole-cell physiology and (2) to determine how different compounds might differ in stereoselectivity. Our initial data (Sun et al., 2009), and data obtained independently by Warmflash et al. (2009), indicate that glucose is stereospecific. Representatives of all three known life-forms on Earth (Bacteria, Archaea, Eukarya) utilize D-glucose but not L-glucose.
The chiral biology of lactate, the subject of the present study, is more complicated. First, lactate formation is not homochiral. In most lactic acid bacteria, fermentation of carbohydrates yields a mixture of D- and L-isomers, although not necessarily in equal quantity (see review by Garvie, 1980 and references therein). Given that both isomers are available on Earth, natural selection would be expected to favor microorganisms that use both isomers over those that use one isomer exclusively. Indeed, some bacteria appear to be dual users and possess two versions of the lactate dehydrogenase, one specific for the D-isomer and one for the L-isomer. These enzymes are required to oxidize lactate to pyruvate, which is achiral and can be further oxidized to carbon dioxide via the tricarboxylic acid cycle (Garvie, 1980). How widespread such dual users are is unknown, but they are common at least among the medically or economically significant bacteria that have been studied. To complicate the matter further, some bacteria also possess a lactate racemase, which catalyzes the stereo-optical conversion of lactate from one enantiomeric configuration to another (Dennis and Kaplan, 1963; Hiyama et al., 1968; Stetter and Kandler, 1973; Hino and Kuroda, 1993; Goffin et al., 2005). These bacteria would be DL-lactate users even if they have only one type of the lactate dehydrogenase.
Therefore, there is a need for critical evaluation of lactate with respect to stereospecificity. This need cannot be met by the data available in the literature, which is qualitative or semiquantitative and limited to a small number of species of bacteria (Oren and Gurevich, 1995; Traudt and Kleinberg, 1996; Evans and Martin, 1997; Duncan et al., 2004; Goffin, 2004). We assessed stereospecificity in lactate with carefully controlled experiments based on natural soil microbial communities. We also studied individual bacteria, archaea, and eukaryotic fungi to compare different life-forms and to compare cultured and natural cells. Our results indicate that, unlike glucose, lactate is not a stereospecific substrate.
2. Materials and Methods
Bacteria
Micrococcus luteus was isolated from Las Vegas dust, Bacillus simplex and Arthrobacter globiformis from soil in the Atacama Desert, Chile. Pseudomonas fluorescens Pf1-1 was isolated by Compeau et al. (1988) from farm soil in Massachusetts, USA. Ochrobactrum anthropi (29243) was obtained from the American Type Culture Collection.
Archaea
Natrialba asiatica (70017) was obtained from the American Type Culture Collection.
Desert soils
Samples came from the Atacama Desert, near the city of Copiapó; Southern Nevada, on Mt. Charleston (Mojave #1) and on the desert floor in Hidden Valley, Southern Nevada (Mojave #2); the Egyptian desert, near Nubaria, Alexandria Governorate; South Australia, near Arkaroola (Australia #1 from underneath a shrub and Australia #2 under a cryptogamic crust); and Eastern Afghanistan, north of the town of Khowst.
Lake water and sediments
Samples originated from the Clark Country Wetland Park, Las Vegas (LV), and Baldwin Lake in the Los Angeles (LA) Arboretum.
Preparation of experiments
For isolates, late-log-phase, aerobically grown cells were collected by centrifugation, washed twice in phosphate-buffered saline or in the base buffer of the growth medium, and re-suspended in the same buffer. Soils were washed and suspended in phosphate-buffered saline in 1 g to 10 ml ratio. Cell density in lake water was increased 100-fold by collecting particulates and re-suspending them in less lake water. Each culture or sample was split into two equal parts to which pure isomers were added, followed by incubation at 20°C with continuous shaking. Lactate was quantified by the use of the colorimetric method of Barker and Summerson (1941). Because each sample contained only one isomer, chiral-specific determination was unnecessary. Cell density was determined by direct microscopy or by plate counts on nutrient agar.
3. Results
All five bacteria consumed D- and L-lactate. In each culture, quantities of the two isomers decreased simultaneously from an initial concentration of about 1 mM to around 0.1 mM, beyond which no further decline was observed (Fig. 1). These DL-lactate users include Gram-positive species Micrococcus luteus, Bacillus simplex, and Arthrobacter globiformis as well as Gram-negative species Pseudomonas fluorescens and Ochrobactrum anthropi.
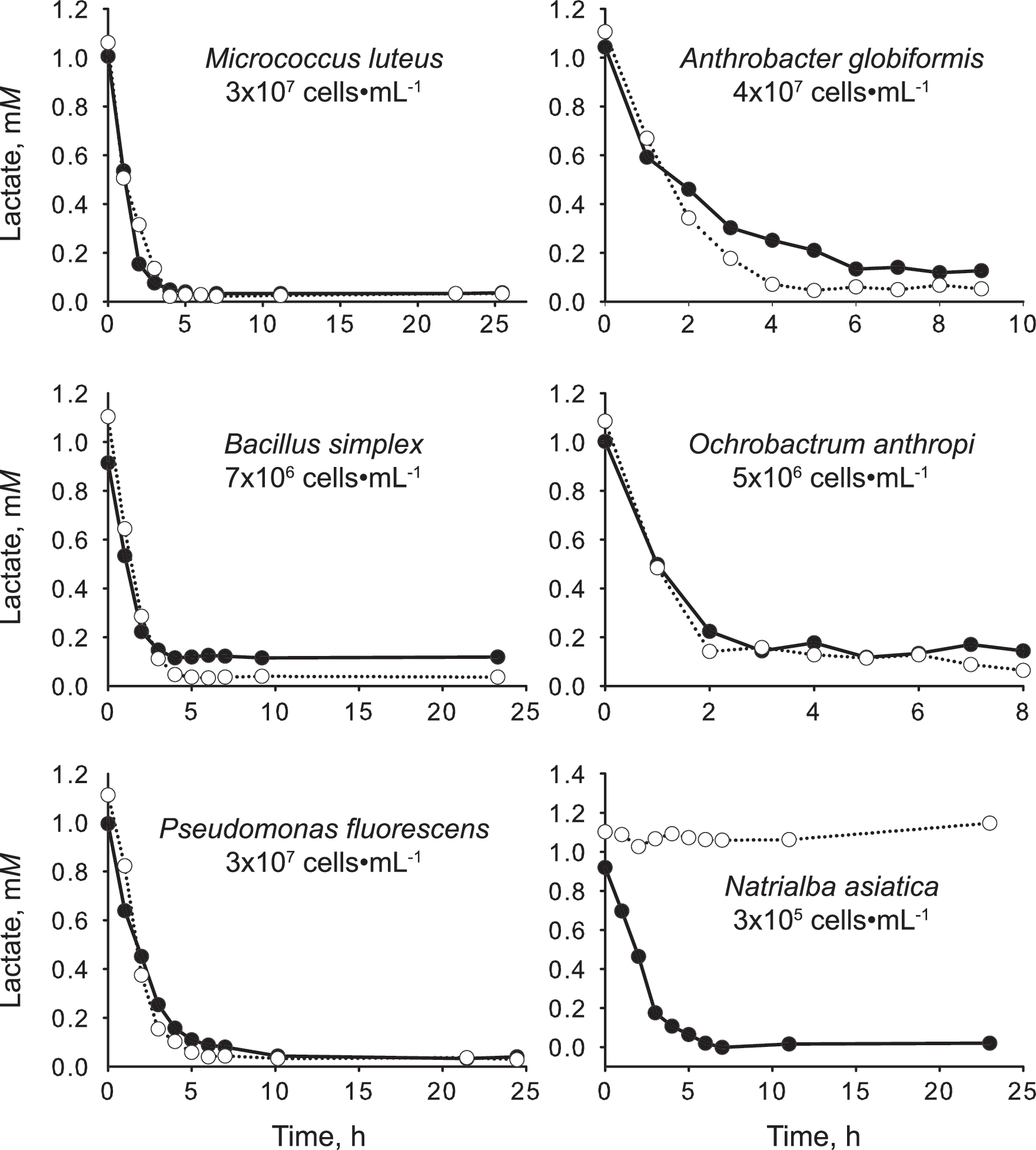
Consumption of D-lactate (filled symbol) and L-lactate (open symbol) by laboratory-grown bacteria and archaea. Only archaeon Natrialba asiatica utilized lactate in a stereospecific manner (using D-lactate but not L-lactate). All five bacteria used the two enantiomers indiscriminately.
The archaeon Natrialba asiatica used D-lactate but not L-lactate. Four other archaea, including Halostagnicola sp., Natronobacterium sp., Haloarcula mukohataei, and Picrophilus oshimae, were also studied. None of them consumed lactate under our experimental conditions.
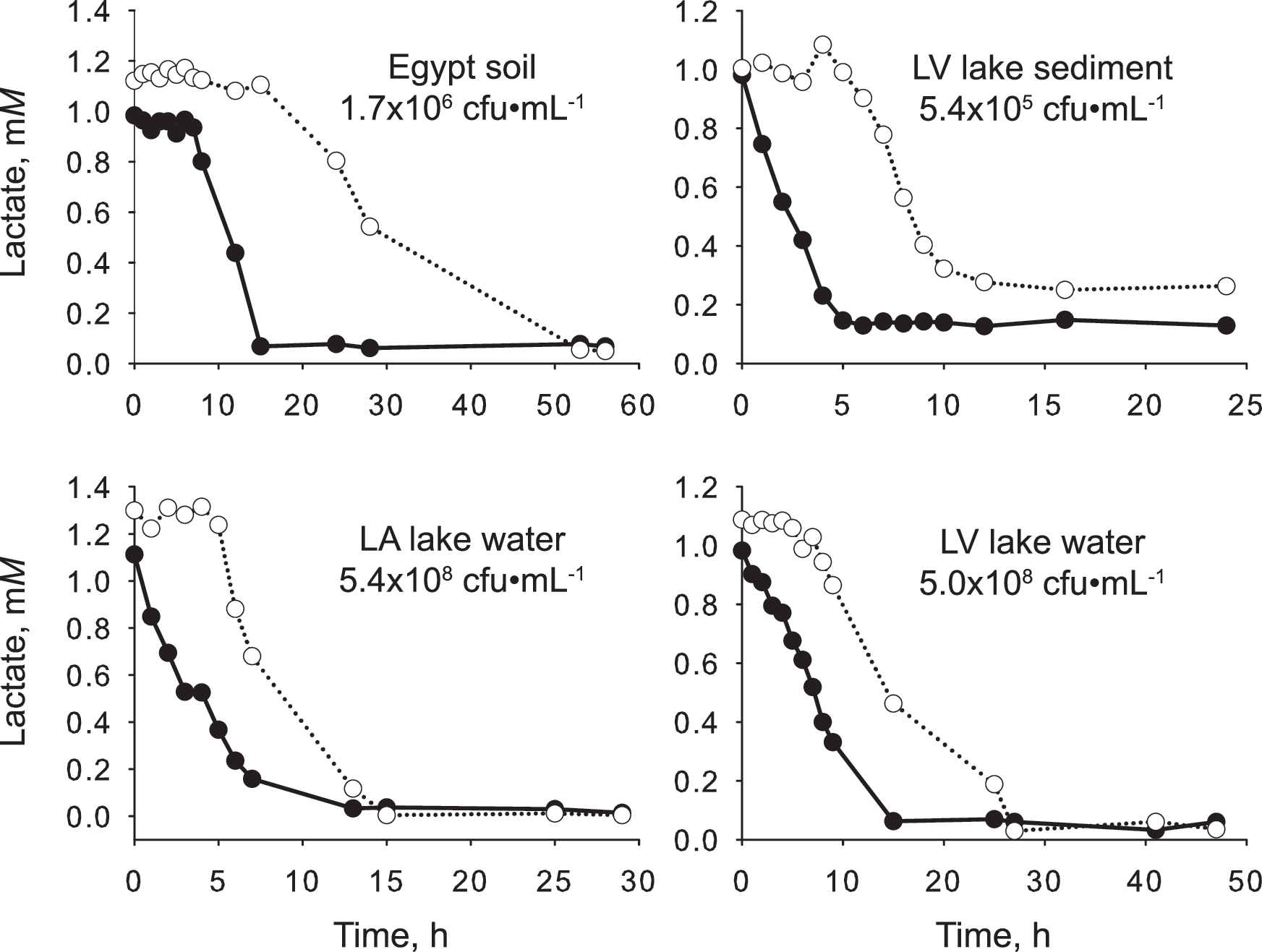
Soil, lake water, and sediment samples consumed the two isomers in an asynchronous manner (Fig. 2). D-lactate consumption started either immediately or after only a short time period. In contrast, L-lactate consumption started after a relatively longer preparation phase. The onset differential between the two isomers ranged from one hour in soil Mojave #1 to five hours in soil Mojave #2, soil Australia #2, LV lake, and LA lake.
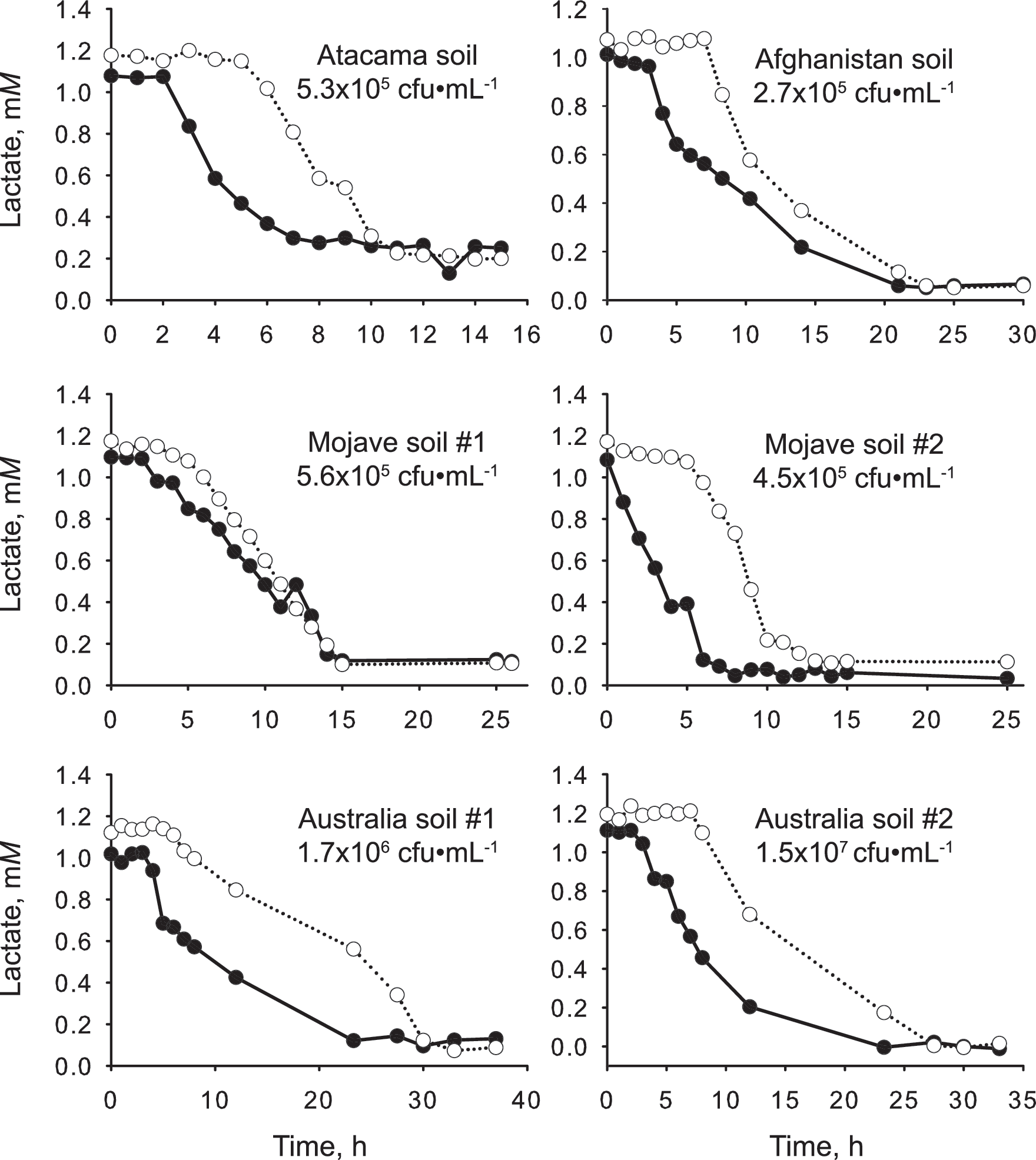
Consumption of D-lactate (filled symbol) and L-lactate (open symbol) in soil, sediment, and lake samples. In all samples, active D-lactate consumption began sooner than did L-lactate consumption. cfu, colony-forming units.
4. Discussion
Our data indicate that lactate is not a stereospecific substrate. The fact that six randomly selected bacteria all use L-lactate as well as D-lactate (Fig. 1) suggests that dual users are common in the domain Bacteria. While some microorganisms do take up lactate in a stereospecific manner, they do not always favor the same enantiomer. Hence, while the archaeon Natrialba asiatica is a D-lactate user (Fig. 1), the literature contains many examples of L-lactate bacteria (Garvie, 1980; Traudt and Kleinberg, 1996). Although complex, eukaryotic organisms are less relevant to the astrobiology of Mars, most invertebrates, plants, and animals metabolize only L-lactate (Cristescu, 2008; Passarella et al., 2008). Our knowledge about D-, L-, and DL-lactate users, especially among the domain Archaea, is incomplete and perhaps even inaccurate. This much is clear: lactate is not a stereospecific substrate as previously understood.
At the same time, lactate is not a stereo-indiscriminate substrate either. All ten soil-, lake-, and sediment-borne microbial communities studied displayed a preference for D-lactate; active consumption of D-lactate started consistently and significantly sooner than L-lactate (Fig. 2). Presumably, D-lactate dehydrogenase activity was constitutive or readily induced. In contrast, the enzymes for metabolizing L-lactate—the lactate racemase, the L-lactate dehydrogenase, or both—were relatively slower to induce. Ultimately, this implies that lactate production in the studied environments is biased toward the D-form. The fact that the bias is consistent in different habitats sampled across different continents is a strong indication that what we observed in these experiments may be a general characteristic of microbial communities on Earth.
The chiral biology of lactate is, therefore, different from that of glucose. In the latter case, chirality appears to be an insurmountable barrier to life. Consequently, sugar metabolism on Earth is confined to the dextrorotary side of the barrier, with no capability to access L-glucose (Sun et al., 2009; Warmflash et al., 2009). We refer to this situation as perfect asymmetry. Lactate, in contrast, is a case of imperfect asymmetry: life operates on both sides of the chiral barrier with unequal efficiencies. On Earth, D-lactate appears to be the primary isomer and L-lactate the secondary isomer. Apparently, for lactate, the chirality barrier has been breached more than once in the biological history of Earth, as the lactate racemase arose multiple times (Le Bras and Garel, 1991; Taguchi and Ohta, 1991; Kochhar et al., 1992). This explains the widespread presence of dual users among different microbial groups. Remarkably, these organisms are able to access both D- and L-lactate, but they do not make both D- and L-enzymes. As mentioned earlier, D- and L-enzymes are identical proteins that differ only in the enantiomeric configuration of the constituent amino acids. As shown with HIV-1 protease, D-enzymes only bind D-substrates, while L-enzymes only bind L-substrates (Milton et al., 1992). D-lactate and L-lactate dehydrogenases are both L-enzymes, composed of L-amino acids.
Lactate still may be considered in planetary chiral life detection, although the signal from a lactate experiment would be subtle compared to the signal from a glucose experiment. Instead of consuming only one of the two enantiomers supplied, a positive biological response would manifest as a difference in the consumption onset between the two enantiomers. Simultaneous onsets between the two reactions would be abiotic in nature. Constructing an in situ experiment with sufficient precision to distinguish the two outcomes would be an engineering challenge. If the challenge could be overcome, enantiomerically selective consumption of lactate could be deployed on Mars as part of a chiral LR. Such an effort is timely in light of the recent evidence for methanogenesis on Mars (Formisano et al., 2004) and the question with regard to its origin, whether it is biological or thermogenic. Methanogenic archaea alone cannot metabolize lactate. In syntrophic associations with bacteria, however, they can (Yang and Tang, 1991). Methanogens typically utilize simple carbon compounds such as formate, propanol, and acetate, which are achiral and of no value for chiral experiments. However, if methanogens are sustained on Mars, there has to be a supply of the substrates, either from a geochemical source or a biota. The latter question is amenable to a sugar-based, chiral life-detection experiment.
Footnotes
Acknowledgments
We thank the NASA EPSCoR Office (grant #NNX07AT65), the Exobiology Program (grant #NNX08AO45G to H.J.S), and the NASA Astrobiology Institute for funding; Doctors Johann Engelbrecht, Heba Ibrahim, and Penny Boston for soil samples; Doctor Eduardo Robleto for Pseudomonas strain; Lisa Wable for graphic assistance; and Roger Kreidberg for editing.
Abbreviations
D, dextrorotatory; GCMS, gas chromatograph–mass spectrometer; L, levorotary; LA, Los Angeles; LR, labeled release experiment; LV, Las Vegas; MOx, Mars Oxidant experiment.