
Abstract
Silicification of organisms in silica-depositing environments can impact both their ecology and their presence in the fossil record. Although microbes have been silicified under laboratory and environmental conditions, viruses have not. Bacteriophage T4 was successfully silicified under laboratory conditions that closely simulated those found in silica-depositing hot springs. Virus morphology was maintained, and a clear elemental signature of phosphorus was detected by energy-dispersive X-ray spectrophotometry (EDS). Key Words: Bacteriophage T4—Virus—Microbial silicification—Fossil record. Astrobiology 10, 569–576.
1. Introduction
V
Viruses are thought by many to be ubiquitous companions of all cellular life (Koonin and Martin, 2005; Forterre, 2006a; Koonin et al., 2006). Although there are numerous cellular organisms without known virus “companions,” this is thought to be due to the difficulties of finding viruses rather than any inherent lack of viruses in these organisms.
Although generally viewed as parasites, viruses can play other roles in the environment, such as promoting horizontal gene transfer between organisms, including organisms of different species (Ripp et al., 1994; Jiang and Paul, 1998; Filee et al., 2002; Weinbauer, 2004; Beumer and Robinson, 2005; Sullivan et al., 2006; Kenzaka et al., 2007). Another important ecological role viruses play is that of microbial predator; in that role, they recycle scarce nutrients and increase the genetic diversity of an ecosystem (Hennes and Simon, 1995; Riemann and Middelboe, 2002; Weinbauer, 2004; Jardillier et al., 2005; Ortmann and Suttle, 2005). This role is particularly important in ecosystems where the more “typical” eukaryotic microbial predators are unable to function. Such ecosystems include hydrothermal environments where the temperature is persistently above the limit for eukaryotic life (Baumgartner et al., 2002, 2003).
Many of these hydrothermal waters become oversaturated in silica as they cool to ambient temperature (White et al., 1956). Microorganisms in these silica-depositing waters can act as nucleation sites for silica polymerization, which leads to their being encased in silica (Schultze-Lam et al., 1995; Cady and Farmer, 1996; Renaut et al., 1998; Jones et al., 2000, 2004; McKenzie et al., 2001; Konhauser et al., 2004; Amores and Warren, 2007; Peng et al., 2007) and, after diagenesis, their preservation in the fossil record (Cloud and Licari, 1968; Schopf and Packer, 1987; Cady and Farmer, 1996; Reysenbach and Cady, 2001; Al-Hanbali and Holm, 2002; Geptner et al., 2005; Schopf, 2006).
In 1915, Charles Walcott was the first to report finding fossilized microbes: blue-green algae (now called cyanobacteria) in limestone of the Newland Formation in central Montana (Walcott, 1915). Since that time, there have been a number of similar findings in a variety of mineral settings (e.g., Barghoorn and Tyler, 1965; Cloud, 1965; Schopf and Packer, 1987; Altermann and Schopf, 1995; Schopf, 2006). Many of the most distinct microbial fossils are those that have been found in silica-rich rock, for example, cherts and flints. Despite vigorous debate about the oldest of these finds (Lowe, 1994; Garcia-Ruiz et al., 2003; Brasier et al., 2005), the consensus is that these fossils date the beginning of microbial life to earlier than 3 billion years ago.
Currently, microbes in hydrothermal springs are often accompanied by, and presumably infected with, a variety of viruses (Prangishvili et al., 1999; Reysenbach and Cady, 2001; Al-Hanbali and Holm, 2002; Breitbart et al., 2004; Geptner et al., 2005; Snyder, 2005; Stedman et al., 2006; Lee et al., 2007). It is reasonable to assume that these viruses would be silicified along with their microbial hosts in a silica-depositing environment. If viruses can be silicified along with their hosts, it is also possible that they might be present in the fossil record.
Discovery of fossilized viruses would be a boon not only to virologists but also to those interested in the origins of life on Earth and elsewhere. While it is supposed that viruses originated at approximately the same time as cellular life (Forterre, 2006a, 2006b; Koonin et al., 2006), there is no direct evidence of viruses in existence prior to the past century. There is, however, some indirect evidence of ancient viruses, such as the presence of structures that resemble polyhedrosis virus inclusions in insects trapped in amber dated to 100 million years ago (Poinar and Poinar, 2005). Phylogenetic analysis also supports the ancient nature of viruses (Hendrix et al., 1999; Filee et al., 2002; Holmes, 2003; Benson et al., 2004; Rice et al., 2004).
While silicification of viruses would be an important first step toward incorporation in the fossil record, it is only a single step. Whether recognizable fossil viruses or detectable biosignatures would survive taphonomic alteration is unknown. The oldest molecular biosignatures, from specimens billions of years old, are remnants of membrane lipids (e.g., Brocks et al., 1999, 2003). While bacteriophage T4 (used in this study) does not have a lipid envelope (the viral analogue to a cell membrane), many viruses, including some bacteriophage, do (Laurinavicius et al., 2007). Proteins (Nielsen-Marsh et al., 2009) and DNA (Noonan et al., 2006; Haile et al., 2009) have been sequenced from specimens tens of thousands of years old. Thus, it appears possible that some viral components could survive diagenesis. If viral lipids can be distinguished from their host membrane lipids, even after taphonomic alteration, it might be possible to identify a viral biosignature even when their morphology has been altered beyond recognition.
In this study, bacteriophage T4 (Fig. 1) silicified under conditions similar to those observed in the outflow channels of silica-depositing hot springs—room temperature and 300 ppm (5 mM) silica (White et al., 1956; Renaut et al., 1998; McCleskey et al., 2004). The methods used were similar to those used in the experimental silicification of bacteria (Westall et al., 1995; Toporski et al., 2002; Yee et al., 2003; Benning et al., 2004a, 2004b; Konhauser et al., 2004; Orange et al., 2009). Bacteriophage T4 was used because it has been extensively studied (Karam, 1994), and its genome (Miller et al., 2003), structure (Fokine et al., 2004; Leiman et al., 2004; Aksyuk et al., 2009), and host interactions (Karam, 1994; Mosig et al., 2001; Filee et al., 2005; Kenzaka et al., 2007) are well characterized. The silicified viruses have a clearly recognizable morphology and a detectable phosphorus signal on elemental analysis.

Simplified diagram of bacteriophage T4 showing the major subunits of the virus.
2. Methods
2.1. Virus stocks
Bacteriophage T4 stock was obtained from Carolina Biologicals at a concentration of between 5 × 108 plaque-forming units/milliliter and 2 × 1010 plaque-forming units/milliliter; it was stored at 4°C until use. Immediately prior to use, 100 μl of the virus stock was dialyzed against sterile deionized water with a Millipore VSWP (0.025 μm pore) membrane for 1 hour to remove salts and proteins from the storage media (Marusyk and Sergeant, 1980).
2.2. Silica analysis
Silica concentrations were determined colorimetrically by the heteropoly blue modification of the silicomolybdate method (Kahler, 1941; Fishman and Friedman, 1989). All reagents and samples were stored in polyethylene or polystyrene containers, and the colorimetric reaction was carried out in polyethylene microcentrifuge tubes.
2.3. Silicification
Silicification was performed at room temperature at a final silica solution of 300 ppm SiO2, which simulates the conditions of some outflow channels of silica-depositing hot springs. This solution was made fresh at the start of each experiment by using solid sodium metasilicate. The pH of the silicification solution was adjusted to between 6.8 and 7.2 with small amounts of 1.0 N HCl immediately before use. Nine hundred microliters of the pH-adjusted solution was then mixed with 100 μl of the virus stock and incubated at room temperature. A control specimen was simultaneously prepared with sterile water at a pH between 6.8 and 7.2 and incubated in parallel.
Silicification experiments were also carried out with sterile, filtered (0.2 μm) water from a known silica-depositing hot spring, “The Growler,” located near Lassen Volcanic National Park (Thompson, 1983). The silica content of this spring, measured at its surface outlet, was 230–270 ppm SiO2 (Thompson, 1983), but at the time of the experiment the silica content of the spring water sample had dropped to 104 ppm, probably due to being stored at room temperature. The silica concentration was adjusted to 300 ppm with the addition of freshly made sodium metasilicate concentrate, and the resulting solution was promptly used for silicification.
2.4. Electron microscopy
Specimens were prepared by dialyzing a 50–100 μl aliquot of the treated virus-containing solution against deionized water with a Millipore VSWP (0.025 μm pore) membrane for 1 hour to remove residual dissolved silica. A 5 μl drop of the dialyzed solution was placed on a 400 mesh copper grid with a carbon-Formvar film (Ted Pella, Inc.) and allowed to remain for 2 minutes before wicking away the remaining liquid. The grids were then negatively stained with 2% uranyl acetate for 30 seconds, after which time the remaining stain was removed by wicking.
Transmission electron micrographs were obtained by a JEOL 100CX transmission electron microscope (TEM) with a Gatan 782 ES500W CCD camera that used an accelerating voltage of 100 kV. High-resolution imaging and energy-dispersive X-ray spectrophotometry (EDS) were performed on an FEI Technai F-20 TEM (accelerating voltage 200 kV) with an EDAX EDS system, which provides “real time” monitoring of the X-ray peaks as the exposure progresses. EDS was performed with a raster-scanned 15 nm electron beam, and the elemental peaks were identified by their characteristic energy (Bearden, 1967).
The EDS was performed by scanning the electron beam over a rectangular region of the virus capsid and a control area of the grid of identical size that lacked virus particles. Comparison with unsilicified bacteriophage T4 was done with the same technique. A longer exposure time was used on the unsilicified bacteriophage T4 when no silicon peak was seen at shorter exposures; this was done to rule out the possibility that silicon in the interior of the virus was somehow “shielded” by the capsid.
3. Results
To determine whether a model virus with well-defined structure—bacteriophage T4 (Fig. 1)—would silicify in a simulated hot spring environment, it was incubated for 48 hours in a 300 ppm (5 mM) silica solution and then observed with the TEM. After exposure, the virus particles appeared to be coated with a conforming layer of silica, with the virus morphology clearly intact (Fig. 2B, 2C). With exposures of over 72 hours, the coating appeared to become thicker; by 120 hours, the viruses were not identifiable. In contrast, the bacteriophage T4 exposed to only water did not appear to be coated (Fig. 2A, 2C).
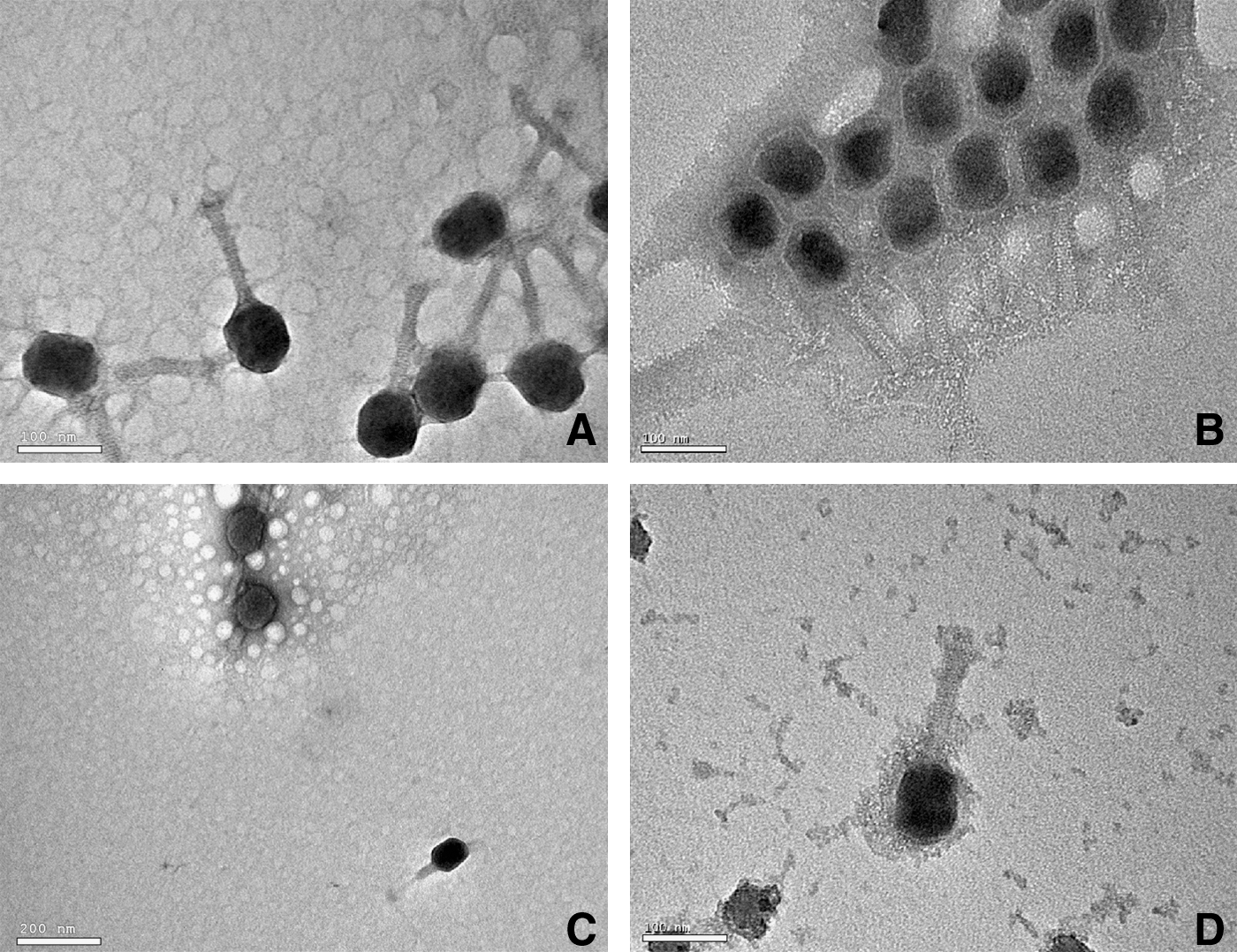
TEM images of control and silicified virus. (
The coating was most apparent around the heads of the virus as a lighter, nonstaining outline of the capsid. Tails of the silica-treated viruses appeared thicker than the untreated viruses. These differences were best observed when the treated and untreated viruses were mixed together in a 1:1 ratio immediately before placing them on the TEM grid and staining (Fig. 2C).
Similar results were obtained with actual hot spring water from the known silica-depositing hot spring, “The Growler” (Thompson, 1983). However, the stored Growler hot spring water used in the experiment contained large numbers of 5–10 nm suspended spheroids. When silica-augmented Growler hot spring water was used, the coating of the virus particles incorporated these spheroids, which gave the virus a “lumpy” appearance (Fig. 2D). Nevertheless, virus morphology was clearly maintained and was very similar to that seen when the simple silica solution was used, as discussed above.
An even more extreme situation was observed when the viruses were exposed to silica solutions in LB or Lysogeny Broth (Bertani, 1951). This medium, used for virus propagation, contains relatively high concentrations of proteins and other organic molecules. Under these conditions, large (50–150 μm) colloidal particles were formed that adhered preferentially to the virus capsid (data not shown). However, these concentrations of organic molecules are not typically seen in a hot spring environment (Wilson et al., 2000; Ball et al., 2002; McCleskey et al., 2004).
To further characterize the treated virus, high-resolution transmission electron microscopy with elemental analysis (EDS) was performed (Fig. 3). Elemental analysis showed a significant silicon peak in the silicified bacteriophage T4, which was absent both in the background of the silicified bacteriophage T4 specimen and in bacteriophage T4 that had been exposed only to water. A small, though clearly defined, phosphorus peak was also seen. Since bacteriophage T4 does not possess an envelope (analogous to the cell membrane of cellular organisms) and its proteins are not known to be phosphorylated, the phosphorus signature must be from the genomic DNA in the capsid. The copper peaks are from the copper grid, and the uranium peaks are from the uranyl acetate stain used to identify the virus particles for analysis.

EDS of the capsid of a single silicified bacteriophage T4, a control region of the same grid, and the capsid of a bacteriophage T4 that had not been treated with silica solution (15 nm spot size). Silicon (Si) and phosphorus (P) signals detected in the treated bacteriophage T4 are greatly reduced in the control region. Silicon was not detected in the untreated bacteriophage T4. The detection of copper is due to the TEM grid (i.e., a copper grid with carbon/Formvar film). Uranium peaks are due to the post-fixation uranyl acetate stain. Image (
4. Discussion
Geochemical considerations predict that the small size of viruses, ca. 100 nm, would preclude their silicification since even under supersaturation conditions, such as those found in silica-depositing hot spring ecosystems, the rate of silica dissolution would exceed its rate of deposition (Alexander, 1957; Kobayashi et al., 2005; Conrad et al., 2007). Results of this study have shown, however, that at silica concentrations below that needed for homogeneous (spontaneous) nucleation (Weres et al., 1980), virus particles can become coated in silica (Fig. 2). Ongoing virus silicification studies in our laboratory indicate that other viruses also become silicified under similar conditions (unpublished results).
The degree of silicification of the T4 viruses is yet to be determined, and it may be that they are not permineralized but only have a surface coating of silica. The capsid of bacteriophage T4 is composed of repeating interlocking proteins and is porous to small molecules, including the uranyl acetate used for staining. The decrease in staining of the capsid after silica treatment, seen most clearly in Fig. 2c, suggests that the silica coating is extensive enough either to reduce the porosity of the capsid or decrease its aqueous internal volume. Still, some internal space remains, as evidenced by the diffusion of stain into the capsid.
As viruses are often the only microbial predators in silica-depositing hot springs, their silicification could render them non-infectious and ultimately lead to a decrease in microbial diversity in these environments (Fuhrman, 1999; Riemann and Middelboe, 2002; Bouvier and del Giorgio, 2007; Suttle, 2007). A decrease in microbial diversity might manifest as a decrease in the number of species, or it could result in one species—or a small group of species—being dominant for a prolonged period of time, as has been reported in some studies in which hot spring microbial diversity was examined over time (Snyder, 2005; Snyder et al., 2007).
Another possible outcome would be for viruses that have adapted to silica-depositing environments to have modifications of their external surfaces that reduce or eliminate silicification. This is significant in light of previous studies that have shown that different peptide sequences and functional groups can have markedly different affinities for silica deposition (Coradin et al., 2002; Naik et al., 2002; Fang et al., 2008; Wallace et al., 2009). A few studies have shown that certain amino acids—histidine, lysine, and arginine—lead to increased affinity for silica binding (Coradin et al., 2002; Naik et al., 2002; Liang et al., 2009), whereas amino acids with alcohol groups—tyrosine, serine, and threonine—tend to reduce binding to silica (Fang et al., 2008). Therefore, structural characterization of indigenous hot spring viruses is a necessary next step in this research. Intriguingly, the coat proteins of fuselloviruses, common in acidic hot springs, are rich in lysine and arginine residues (Palm et al., 1991; Stedman et al., 2003; Wiedenheft et al., 2004; Redder et al., 2009), which should make them more susceptible to being encased in silica.
Although our experimental findings have shown that viruses will silicify under conditions similar to those found in silica-depositing hot springs, it remains to be seen whether silicified viruses can be found in the environment. Kyle et al. (2008) showed that viruses in the Río Tinto can become coated with iron oxides, which suggests that silicified viruses should occur when geochemical conditions favor silica deposition. Perhaps the most important implication of this work is that viruses coated with silica have the potential to become incorporated into the fossil record along with their microbial hosts. This study has also shown that the initial steps of the silicification process leave the virus morphologically recognizable, which raises the possibility that fossilized viruses might some day be found. In addition, elemental “fingerprints” (Fig. 3) may allow detection of silicified viruses in petrographic thin sections. The fossilization of viruses in mineralizing environments, and the potential to preserve their biosignatures in the geological record, has profound implications for the study of viruses and their antiquity.
Footnotes
Acknowledgments
The authors would like to thank Dr. Sherry Cady for her advice on the manuscript and support throughout. J.L. was supported by an NSF-IGERT fellowship. Research in the Stedman lab is supported by NASA (NNX07AJ26G and NNX07AT63A. Subaward G258-08-W1951) and NSF (MCB0702020). The authors would also like to thank Mr. Pete Seward for access to the Growler hot spring.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
EDS, energy-dispersive x-ray spectrophotometry; TEM, transmission electron microscope.